
Abstract
The emergence of protocells, membranous compartments with encapsulated genetic material, was a crucial step in life’s origin and evolution. The hot spring hypothesis for the origin of life suggests that protocells with the capacity to encapsulate organic matter could have formed in hot spring pools during wet–dry (WD) cycling of hydrothermal fluids. Previous investigations have focused on mimicking WD cycles within a single pool, which precludes simulation of many hydrothermal field conditions, such as different mineralogies and variable temperature, pH, and water flow within and between multiple hot spring pools. Here, we present a modular 3D-printed hydrothermal field simulator that mimics many more aspects of the complex nature of hot spring fields by controlling the temperature, pH, and mineralogical variability of a series of linked pools. Furthermore, the pools can be programmed to experience fluid mixing between proximal pools and periodic WD cycling events. Results with the prototype hot spring field design demonstrate the ability to spontaneously form lipid vesicles that encapsulate organic matter within membranous compartments comprised of decanoic acid:decanol (4:1) or the phospholipids POPC:POPG (1:1). We observed that the vesicles formed during multiple WD cycles in the simulator pools displayed variation in their size distribution and differences in the number of membrane layers. Cargo encapsulation was favored in giant unilamellar vesicles and oligolamellar vesicles. Overall, the hot spring simulator offers a novel and customizable approach for studying multiple processes within hydrothermal field dynamics that include prebiotic chemical reactions, mineral surface catalysis, and the complexities of fluid mixing between proximal hot spring pools.
Introduction
Extant hot spring fields commonly comprise multitiered pool networks interconnected by natural surface channels and subsurface vein networks, with fluid flow driven by gravity, splashing geysers, and periodic evaporation–precipitation cycles (Djokic et al., 2017; Damer and Deamer, 2020). These complex systems result in a range of local microenvironments, each with distinct hydration states, residence times, and exposure to sunlight, while enabling wet–dry (WD) cycling. Terrestrial geothermal hot spring pools on early Earth have been hypothesized as plausible sites for the origin of life in particular for their ability to support WD cycles. As a consequence, hot springs are able to concentrate material that could facilitate prebiotic chemical reactions (Powner et al., 2009; Becker et al., 2019; Tran et al., 2020; Zhao and Wang, 2021) and the self-assembly of protocells with encapsulated organic materials (Djokic et al., 2017; Deamer et al., 2019; Damer and Deamer, 2020). Hot spring pools could have served as open reaction chambers for prebiotic chemical reactions, with rock surfaces providing microenvironments (Saha et al., 2022) for prebiotic chemistry. Moreover, the occurrence of (semi)periodic WD cycles could have helped drive processes that are thermodynamically unfavorable under aqueous conditions, such as the polymerization of nucleotides and amino acids (Benner et al., 2012; DeGuzman et al., 2014; Frenkel-Pinter et al., 2020). Experimentally modeling the full complexity of terrestrial hot spring fields, however, remains a difficult challenge.
Hot spring fields, particularly those resembling ancient terrestrial hydrothermal systems—such as those found in the Pilbara Craton (Western Australia)—are characterized by significant spatial and temporal heterogeneity in their physical structure, temperature gradients, fluid dynamics, and mineral compositions (Djokic et al., 2017; Van Kranendonk et al., 2021; Van Kranendonk, 2026). This variability plays a central role in their proposed suitability as environments for prebiotic chemistry and the emergence of life. Indeed, the overall physicochemical complexity (e.g., pH, salinity, element availability, mineralogical variability, and temperature), WD cycling dynamics, and the presence of mixing zones between individual pools within a hot spring field allow for combinatorial molecular interactions under variable selection pressures (Damer and Deamer, 2020). Localized differences in temperature, mineralogy, and solute concentration are proposed to drive molecular evolution by fostering diverse reaction pathways, differential stability of protocells, and the selection of more robust chemical systems (Damer and Deamer, 2020; Van Kranendonk, 2026). For example, thermal gradients occur across individual pools (Mast, 2024; Matreux et al., 2024), along their outflow and runoff channels, and between connected pools. Meanwhile, individual pools can have temperatures that range from ambient to near-boiling (∼100°C). Differences in pool temperatures and thermal gradients are influenced by their proximity to geothermal vents, evaporation rates, and solar exposure (Power et al., 2018). Along with variations in geochemistry, such differences will influence solute concentrations, phase separation, and consequently, vesicle stability when considering prebiotic scenarios.
The local geological substrates that comprise the walls and floors of hot spring pools and outflows, such as basalt, rhyolite, clay-altered equivalents (kaolinite and montmorillonite bearing substrates), introduce additional geochemical variability when such systems first develop at the surface. These rocks and their glassy or mineral assemblages directly influence the type of catalytic reactions, the surface energy, and adsorption properties of these systems. For example, montmorillonite can catalyze RNA polymerization and promote lipid vesicle formation, while kaolinite and basalt offer different pH buffering capacities and ion exchange properties (Ferris and Ertem, 1992; Hanczyc et al., 2007), along with the ability to catalyze RNA synthesis (Jerome et al., 2022). Iron sulfide (FeS) phases, relatively abundant in hydrothermal systems, are also considered prebiotically significant because they can catalyze a range of processes and participate in redox reactions (Wächtershäuser, 1988; Herschy et al., 2014; Yamaguchi et al., 2014; Roldan et al., 2015; White et al., 2015; Nan et al., 2024).
To build a systematic understanding of the ways in which the complex physicochemical parameter space of hot spring fields could have contributed to prebiotic chemistry, we have built a novel prototype hydrothermal field simulator capable of mimicking many of the key dynamic features of interconnected hot spring pool systems. The design of our simulator was informed by our expert understanding of the geological context of terrestrial hot spring fields, their ancient analogues, and the potential variability of different parameters often studied in hot spring systems (Van Kranendonk, 2026). In nature, WD cycles due to wetting and evaporation are caused by the natural rise and fall of fluid levels in hot spring pools driven by variable pressure in the subsurface fluid flow system, by the splashing that occurs around geysers, from the boiling and turbulence of more active hot spring systems, and by cycles of rain and dry weather patterns. Thus, having the ability to rehydrate using either freshwater or other solutions (as opposed to just freshwater) is more ideal when recapitulating the hydration step.
Our simulator consists of 3D-printed “cups” that serve as models for individual hot-spring pools. Modular in design, these cups can be interconnected via channels that allow for fluid exchange. The prototype design integrates temperature and water control systems to each individual cup, affording simulation of dehydration and rehydration events. The surface of the cups can be coated with different minerals characteristic of extant hot springs. In principle, this design enables each cup to contain aqueous solutions with independently controlled pH, temperature, and solute composition, lined by distinct mineral powders, while also allowing these solutions to mix in a systematic and programmable manner.
The proof-of-concept test case we illustrate here demonstrates the feasibility and durability of the prototype under multiple WD cycles to form fatty acid vesicles, as well as to demonstrate the formation of lipid bilayer membranes capable of encapsulating organic material. This test case simulates previously published laboratory experiments that have shown that dehydration and subsequent rehydration of phospholipids can entrain DNA (Deamer and Barchfeld, 1982). In this work, we used a short-chain fatty acid because their presence on early Earth is generally accepted (Cohen et al., 2023). Indeed, biochemistry is not required to synthesize fatty acids, evidenced by the presence of short-chain fatty acids identified in the Murchison meteorite (Lawless and Yuen, 1979; Deamer and Pashley, 1989). The ability of fatty acids and even meteorite extracts to spontaneously self-assemble into primitive vesicles and to uptake and retain inner contents delineated from the external environment are features that closely resemble the activity of modern biological cell membranes and have been explored extensively elsewhere (Deamer and Pashley, 1989; Walde et al., 1994; Morigaki et al., 2003; Steller et al., 2022).
Specifically, our experiments with the hot spring simulator demonstrate that the vesicles assembled from decanoic acid (DA), DA:decanol (DO), or phospholipids can encapsulate and retain fluorescent dye after multiple WD cycles. We also demonstrate that encapsulation is favored in thin-walled vesicles and that during WD cycling, an anionic dye and RNA can also be encapsulated. We then propose further research and discuss how our prototype could unveil novel avenues of research relevant to addressing the hot spring hypothesis for the origin of life.
Experimental Section and Hot-Spring Simulator Design
Materials and reagents
DA, DO, and both phospholipids (POPC 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine and POPG 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoglycerol) were purchased from Sigma-Aldrich and used as received. All water used was purified to 18 MΩ (Milli-Q). A stock solution (0.1 M) of phosphate buffer (PB) was prepared by dissolving appropriate amounts of NaH2PO4 and Na2HPO4 in water, followed by adjustment of pH (7.4) using either 5 M HCl or 5 M NaOH (Lowy solutions). Similarly, 50 mM HEPES [4-(2-hydroxyethyl)−1-piperazineethanesulfonic acid] buffer, which was used for samples that contained phospholipids, was prepared by dissolving HEPES in water and adjusting the pH to 7.4. The basalt sample was collected from a recent, fresh lava flow just outside Reykjavik, Iceland (exact coordinates not recorded). The kaolinite sample was collected from altered basaltic rocks of the 3.49 Ga North Star Basalt that underlies the stromatolite-bearing Dresser Formation at the Dresser Mine site in the North Pole Dome, Pilbara Craton (UTM Zone 50 K, E752698, N7659202). The pyranine/HPTS (8-hydroxypyrene-1,3,6-trisulfonic acid trisodium salt, Sigma Aldrich Australia) stock solution was prepared in water and used as an encapsulation dye for both fatty acid and phospholipid-based vesicles. Either 0.1 M PB or water was used to rehydrate the lipids during the WD cycles. For experiments requiring the use of RNA, 1 mg mL−1 of RNA (ribonucleic acid from torula yeast, Type VI, Sigma-Aldrich) solution was prepared using 0.1 M PB (pH 7.4) containing 10 mM EDTA (ChemSupply Australia). The RNA solution was adjusted to pH 7.4 with 5 M NaOH. The fluorescent dye in a QuantiFluor® RNA System kit (Promega) was used for imaging RNA. Adjustment of pH for all samples was carried out using an Orion Star A121 pH meter with an Orion 8103BN mini-probe (Thermo Fisher).
Designing, 3D printing, and Pool configuration
To mimic hot spring pools on early Earth, a modular and scalable hot-spring pool simulator was developed (Fig. 1, Supplementary Figs. S1 and S2). The design aimed to produce small volume (2–3 mL), thermally responsive, and chemically robust units capable of integration into interconnected arrays for studies on prebiotic chemistry and protocell dynamics.

Photograph of the model hot spring pools system showing the assembled pools with temperature probes placed in the cups and heating pads inserted underneath for controlling temperature. Power supply provides electricity for heating pads, and an Arduino Uno is connected via USB-type-C cable to a laptop for PID (Proportional-Integral-Derivative) control. Blue-colored LEDs in the Arduino Uno turn off when the setpoint is reached and back on to maintain the temperature in the rehydration phase of the WD cycles. Water flow control is managed by the mini-peristaltic pumps connected to the water control unit.
SolidWorks® CAD (Computer-Aided Design) software was used for parametric 3D modeling of the pool units (cups). The design prioritized (1) low internal volume to both minimize reagent consumption and allow efficient heating, (2) interconnectivity between cups to emulate natural flow pathways found in volcanic terrace systems, (3) mechanical robustness for repeated WD cycling and heating to temperatures up to 100°C, and (4) low hydraulic resistance to support passive or pump-assisted flow of solution between cup modules.
Each cup was designed with a hemispherical internal geometry and incorporated fluid channels at the top rim. These channels or silicone tubing, which are hemispherical in cross-section, were positioned to enable gravity-driven drainage and minimize surface tension effects and contact-line pinning that can disrupt flow at small scales. CAD files will be provided upon request.
3D printing
Cups were fabricated via stereolithography (SLA) 3D printing using Formlabs Durable Resin. This material was selected for its high dimensional stability, smooth surface finish, and thermal resistance. The use of SLA enabled the fine detail resolution (∼50 μm layer height) required for the small-volume pool and channel structures. Postprint processing involved washing with isopropanol to remove uncured resin, followed by UV curing under 405 nm light for mechanical and thermal stabilization. A conformal waterproof coating (epoxy or silicone-based) was then applied to the internal surfaces to prevent seepage during prolonged heating.
Cups were also fabricated via fused deposition modeling (FDM) 3D printing using polylactic acid (PLA). This material was selected for its high dimensional stability, thermal resistance, and compatibility with the applied rock coating.
Test prints validated rock coating compatibility, dimensional tolerances, and fluidic behavior. Successive design iterations adjusted channel diameters and angles to ensure reliable unidirectional flow and avoid overflow or stagnation under gravity-fed conditions.
Physical configuration and modularity
The cup units were arranged on a custom-designed platform that allowed for both standalone testing and future integration into multi-pool/cup systems. Inter-cup alignment was facilitated by custom inserts to standardize spacing and tilt angle. The modular nature of the design allows researchers to configure arrays in various topologies (e.g., linear cascades, radial trees) depending on the experimental goals, such as simulating directional flow, temperature gradients, or compartmentalized chemistry.
The downscaled footprint (∼20 mm diameter per cup) supports rapid thermal cycling and minimizes thermal inertia. These features are essential for experiments that involve WD cycles, where temporal control over dehydration and rehydration is critical to test the conditions required for prebiotic polymer synthesis and encapsulation.
Programmed water and temperature control for multipool systems
To simulate the dynamic physicochemical environment of ancient volcanic hot spring terraces, a programmable control system was developed to regulate water levels and temperatures independently across multiple interconnected cup units. This system enables precise spatial and temporal control over key experimental parameters critical to prebiotic chemistry research, which include WD cycling, thermal gradients, and fluid exchange.
System architecture
The control architecture was based on an Arduino Uno microcontroller, interfaced with temperature sensors, heating elements, and future provisions for fluid-handling components (e.g., peristaltic pumps and solenoid valves). The system was designed to be modular, scalable, and capable of operating independently for each cup within the array.
Temperature monitoring and feedback
Temperature monitoring was implemented using K-type thermocouples connected via a MAX31855 thermocouple-to-digital converter shield. This allowed each cup to be monitored with a spatial resolution of one thermocouple per unit, providing direct temperature readout with ± 2°C accuracy across a range of –50°C to 250°C.
Data from the thermocouples were transmitted to the Arduino Uno and visualized via a custom-designed graphical user interface developed using Windows Forms. This interface provided real-time streaming of temperature values, graphical plotting via a Python-based live grapher, and CSV export for postexperiment analysis. Data sampling frequency was tuned to 0.33 Hz (1 sample every 3 s) to avoid serial buffer overflows and ensure reliable long-term acquisition.
Temperature control strategy
Although a complete heating control loop was not yet implemented at the time of this study, the architecture was designed for integration with resistive heating elements placed beneath each cup, isolated via a thermally conductive barrier (e.g., silicone pad or Aluminium block) to prevent direct contact with the printed resin.
Modularity of the codebase ensures that new subsystems (e.g., pumps or additional sensors) can be integrated with minimal disruption to the existing framework.
Rock coating of the model pools to mimic surficial early Earth pools
To simulate the mineral surfaces of early Earth hot springs, a robust and repeatable rock coating methodology was developed for application on 3D-printed cup interiors. The aim was to reproduce chemically and structurally relevant mineral surfaces (e.g., basalt and kaolinite) that could endure WD cycling without degradation, thereby enabling prebiotic chemistry experiments on realistic geological substrates. Individual cup geometry facilitated balanced rotational motion during coating, improving coating uniformity and minimizing structural distortion during oven curing.
The rock coating process was executed in three distinct stages: (1) preparation of a PDMS intermediate layer, (2) formulation of the rock slurry, and (3) application and curing of the coating. Particular attention was paid to coating uniformity, adhesion, and resistance to aqueous dissolution.
Polydimethylsiloxane (PDMS) served as a hydrophobic adhesive substrate for the rock particles. A 10:1 ratio of PDMS base to curing agent was mixed and diluted with hexane in a 1:1 solvent-to-PDMS ratio to reduce viscosity and promote even spread on curved cup surfaces. Approximately 0.5 g of this solution was applied per cup, manually rotated to ensure even coating, and cured at 60°C for 2 h. Following curing, plasma treatment was performed using a Harrick plasma cleaner for 5 min to temporarily convert the PDMS surface from hydrophobic to hydrophilic to enhance adhesion of the aqueous rock slurry.
Comparative testing of the coating behavior on PLA compared with printed Durable Resin cups indicated superior adhesion and uniformity on PLA (Supplementary Fig. S3). The resolution of FDM printing was demonstrated to be more advantageous for our application, as the coarser surface finish increased the surface area of the cups and enabled greater adhesion of the rock coating to the cup surfaces, compared with using a durable resin material.
The geologically relevant materials kaolinite and basalt were selected for testing. Submillimeter-sized grains were prepared by a Fritsch Jaw Crusher and a Beige Ring mill, then ground with an agate mortar and pestle for 5 mins. The powders were then passed through a 106 µm sieve to remove the largest particles. Slurries of each powder were mixed with water and lime [Ca(OH)2] as an additive to improve adhesion and reduce cracking during drying. The slurry was applied and manually spread across the PDMS-treated surface. Drying was carried out at 40°C for 3–5 min to avoid rapid solvent evaporation and associated cracking. Microscopic examination at 10× and 50× magnification confirmed that a 5% lime slurry (rock:water: lime at 1:2.5:0.18) yielded the most homogeneous coatings, with minimal cracking and high surface coverage (Supplementary Fig. S4). Qualitative water droplet and submersion tests demonstrated coating resilience for the 5% lime slurry samples under fluid delivery, with minimal delamination observed after multiple wetting cycles (Supplementary Fig. S5).
Vesicle preparation
Vesicles were made via the thin-film hydration method. The appropriate amounts of lipid [DA, DA:DO (4:1), and phospholipids] were dissolved in chloroform (Sigma-Aldrich), and then the chloroform was evaporated under a gentle stream of nitrogen followed by the addition of a hydration buffer (0.1 M PB). Samples of the resulting solutions were then imaged to confirm vesicle self-assembly. Samples undergoing WD cycles were heated to a temperature of ∼60°C–70°C in the 3D-printed cups until all the solvent evaporated, after which the materials were then rehydrated with the respective buffer or water. As a control to compare morphological differences between experiments, one WD cycle was also performed in a glass vial that contained lipids and was heated at the same temperatures used for the model pool system experiments.
To obtain contrast for imaging, fatty acid vesicle samples were diluted 3× in water and phospholipid vesicles (2.5 mM) diluted 6× in 50 mM HEPES. Stock solutions of DA vesicles (150–200 mM) were diluted such that the final concentration of the vesicles remained above the critical vesicle concentration (CVC ∼40 mM). For RNA-containing DA samples, the pH was adjusted to 7.35–7.40, and the vesicles were either washed twice using 100 kDa molecular weight cut off filters (5000 g for 10 mins) or diluted 3× in 0.1 M PB to obtain improved contrast for imaging. Experiments were repeated at least twice; for each repeat, all vesicle samples were freshly prepared on the same day as the subsequent WD cycling and imaging.
Microscope imaging
Images were captured using a Nikon Eclipse TE-2000 inverted microscope with a 100× Ph3 objective [Plan Fluor, numerical aperture (NA) = 1.3] equipped with a camera (pco.edge 4.2 sCMOS). For all images captured, the focus was set to the middle planes (solution phase) of the sample to image the products representative of the experimental solutions and avoids imaging vesicles that had been immobilized by the glass cover slips or glass slides, or those that had grown on the glass surfaces. Image analysis such as adjustment of brightness or contrast as well as insertion of scale bars was done using ImageJ/FIJI.
Results
Modular design considerations
The design of our initial hot-spring simulator prototype was based on an interconnected modular cup-based design, with each cup representing one hot-spring pool, that is, one set of conditions. This prototype consists of a three-pool system in which each cup is connected and fluid flow is driven by mini-peristaltic pumps and gravity (Fig. 1). Heating pads, which can be inserted under each of the cups, are covered with a layer of sand to minimize any direct burning of the cup resin and allow for an efficient exchange of heat between the cup and the heating pad. Two adjacent cups are connected via a closed channel and kept at the same surface level, while a third cup is at a lower height and connected only to the second cup via an open terraced channel to simulate conditions observed in natural hot spring systems. This unique design can simulate dynamic water flow that, along with temperature control within individual cups, can promote prebiotic chemical reactions in the different cups. The design enables different prebiotically relevant chemical reactions to occur in different pools due to WD cycling of fluids in the different cups (e.g., condensation of monomers and encapsulation of polymers such as RNA in lipid vesicles), and the collection of samples for analyses after the different fluids and reaction products are mixed in the third cup. Because various minerals in the weathering zones of acidic hot springs, particularly montmorillonite, have also been shown to be important in prebiotic chemistry (Ferris, 2006; Markússon and Stefánsson, 2011), the cups were designed so that they could be coated with rock and/or mineral particles for a more faithful simulation of the conditions of actual hot spring pools, as discussed in Section 2.4 and Supplementary Figures S3–S5.
Vesicle formation was not affected by the 3D-printed material (PLA vs. durable resin) of the cups
We initially investigated whether the 3D-printed resin of the cups affected the formation of vesicles (Fig. 1). PLA and durable resin were used to print the cups as they are among the most commonly used polymeric materials for 3D printing. These polymers differ in their material properties that include biodegradability, flexibility, strength, and durability, resistance to heat, and surface finish. It was found that the surface finish, which can significantly affect how lipids will assemble, was visibly smooth for durable resin and rough for PLA. Nonetheless, surficial features smaller than the wavelength of visible light, such as roughness at the nanometer scale, could be similar.
The 3D-printed cups were tested at elevated temperatures to mimic the geothermal environment of hot spring pools and geysers (∼60°C–70°C). Evaporation rates and any deformations of the cups were monitored during the experiments. Placing a thin layer of sand between the cups and the heat source (a heating pad) provided a more even distribution of heat in the cups as monitored by temperature probes positioned in the cups. An even heat distribution makes the evaporation of water more efficient. Complete evaporation of the fluid resulted in adsorption of lipids to the cups’ surfaces. Although PLA and durable resin are known to be inert, the cups were immediately rewetted after complete dehydration to avoid potential degradation and any chemical reactions with and between the adsorbed lipids.
It was found that vesicles formed after multiple WD cycles (Fig. 2) independent of the composition of the cup material. The PLA resin, possessing a larger degree of surface roughness, was found to be more conducive for coating with mineral particles, and is thus recommended for ongoing work.
Pairs of images showing bright-field (left) and fluorescence (right) images of DA and POPC:POPG (1:1) vesicles before and after WD cycles in the glass vials and model pools (cups).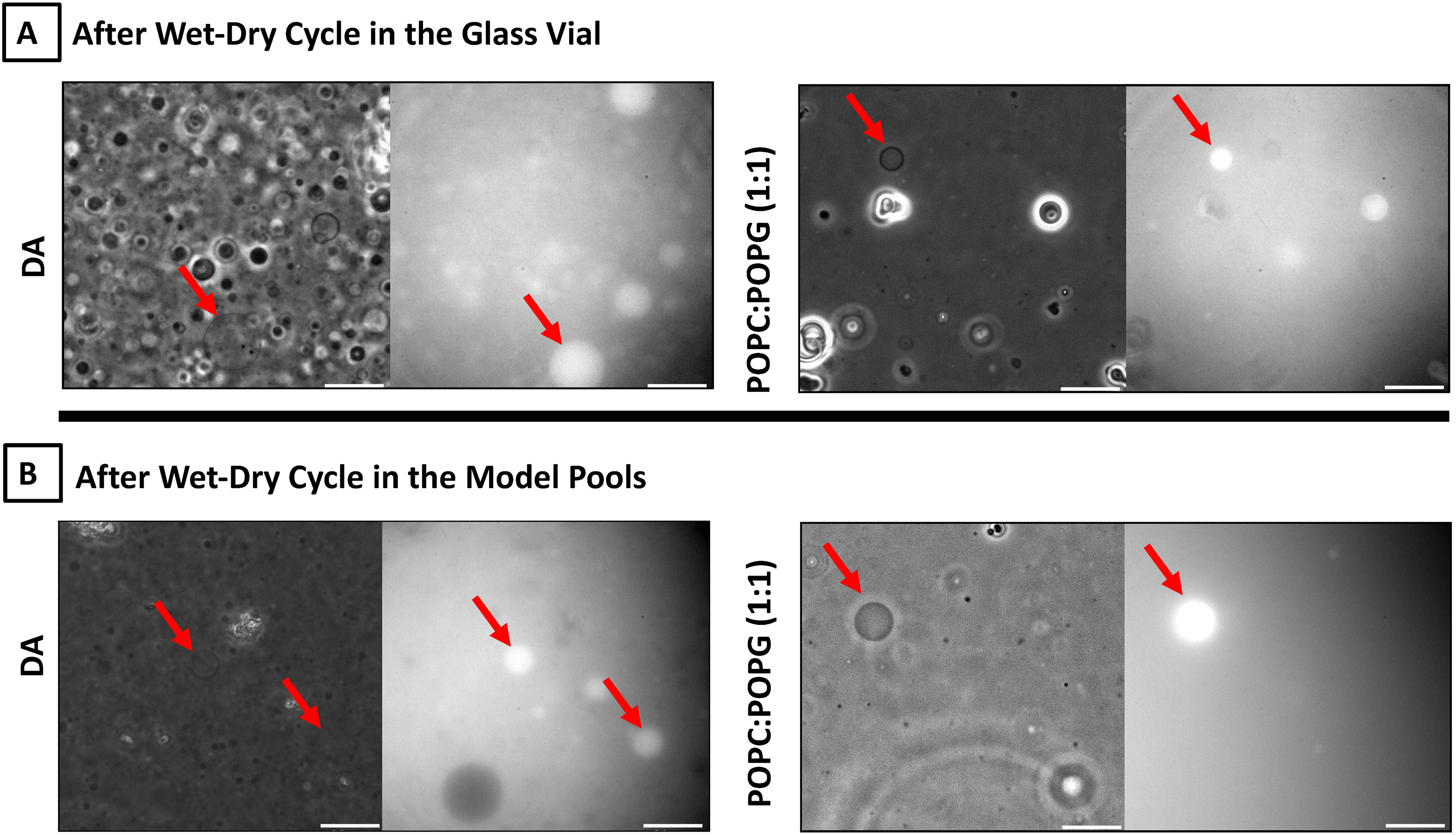
We performed WD cycles both in a glass vial and in the cups in order to test whether the WD cycles in the cups yielded vesicles with different morphologies. DA was used instead of long-chain oleic acid or other fatty acids to mimic the prebiotically plausible short-chain fatty acids. The morphological variation of the vesicles was compared with that of phospholipid vesicles also subjected to WD cycles.
It was observed that lipids subjected to WD cycles produced heterogeneous populations of vesicles such as thin-walled unilamellar or oligolamellar vesicles and thick-walled multilamellar vesicles as well as lipid aggregates (Fig. 2). WD cycling in the cups yielded fewer vesicles compared with the glass vial. Glass vials are known to have smooth and negatively charged surfaces, which could trigger the release of lipids upon rehydration as a result of electrostatic repulsion between the glass surface and negatively charged headgroups of the phospholipids and fatty acids. In contrast to glass vials, the rougher surfaces of the cups may have a greater tendency to keep the lipids adsorbed and/or vesicles trapped in surficial microstructures.
Encapsulation is favored in thin-walled vesicles
We then investigated whether different vesicle morphologies influenced the encapsulation of organic material such as RNA. For ease of investigation, we first used the fluorescent reporter dye HPTS as a model prebiotic material and evaluated its encapsulation in both fatty acid and phospholipid-based vesicles after multiple WD cycles. As observed in the initial experiments, the cups triggered the formation of multiple types of vesicle populations, especially thin-walled oligolamellar or unilamellar vesicles (Fig. 3). We were particularly interested in the thin-walled unilamellar vesicles as they best resemble extant biological cells. We previously demonstrated that one WD cycle in an Eppendorf tube leads to the formation of some unilamellar vesicles and enhanced encapsulation of both HPTS and fluorescently labeled RNA (Steller et al., 2022). Unilamellar vesicles have various advantages over their multilamellar counterparts, such as ease of permeation of nutrients and waste across the bilayer, a potentially necessary feature for early protocells to sustain internal metabolic reactions.
Bright-field (left) and fluorescence (right) images of DA and DA:DO (4:1) vesicles in 0.1 M phosphate buffer (pH 7.4) with HPTS dye before and after WD cycles in the model pools (cups). Before the WD cycle, dye was not efficiently encapsulated in the vesicles. However, after the WD cycle, dye encapsulation was favored in the giant thin-walled vesicles formed by DA and DA:DO (4:1) with the latter showing more encapsulation than with DA alone.
Here, we observed that multiple WD cycles yielded many oligolamellar or unilamellar vesicles, and fluorescence microscopy revealed enhanced encapsulation of HPTS in these vesicle morphologies compared with thick-walled multilamellar vesicles. As explained by Steller and coworkers, this result is potentially a consequence of the apparent pKa of the bilayers changing during WD cycling and the thin-walled structures being more osmotically accessible than thicker lipid stacks (Steller et al., 2022). During the dehydration phase of the WD cycles, the ionic strength of the solution increases, leading to a decrease in apparent pKa that would favor the formation of a bilayer phase even at lower pHs. When the salt concentration is high, recruitment of protons to the bilayer surface is expected to become less efficient, leading to a decrease in apparent pKa and thereby enabling lipids to stack in a layer-by-layer fashion while incorporating cations (e.g., Na+) along with other encapsulated solutes. Such physical preorganization of lipids in layers is considered crucial in the formation of giant unilamellar vesicles, as well as in the encapsulation of solutes. The thinner bilayer stacks containing the most solutes should swell fastest upon rehydration, consistent with the observation of thinner-walled vesicles encapsulating the most dye (Steller et al., 2022).
The morphology of the vesicles can vary depending on their lipid composition and external environment (e.g., pH, temperature, and salt concentration). For instance, it is known that the addition of DO to DA vesicles stabilizes the membranes by facilitating hydrogen bonding between hydroxyl and carboxylate/carboxylic acid head groups, respectively, even at higher pHs (Apel et al., 2002).
To investigate whether vesicles that contained mixtures of fatty acid and fatty alcohol affected the encapsulation of solutes, vesicles were prepared using a 4:1 mixture of DA and DO and subjected to WD cycles in the model pools (Fig. 3). We observed that the addition of 20 mol% DO promoted the formation of oligolamellar or unilamellar vesicles and enhanced the encapsulation of HPTS, while also lengthening the shelf-life of the high concentration samples (∼400 mM). In contrast, DA alone was prone to an abrupt decline in pH at high concentrations due to protonation and formation of neat oil at the air-water interface. Some of these differences can potentially be attributed to the different critical packing parameters of the membrane constituents (Israelachvili, 2011).
Encapsulation of RNA in vesicles is similar to dye encapsulation
After optimizing the experimental setup with HPTS and developing an analysis protocol, we next evaluated the encapsulation of RNA in the vesicles under similar conditions.
In this experiment, DA was used to form vesicles in 0.1 M PB, and the encapsulation of RNA from torula yeast (0.05–0.1 mg mL−1) was investigated. The RNA was dissolved in a 0.1 M PB that contained 10 mM EDTA, which serves as a chelating agent for trace divalent cations such as Ca2+ and Mg2+ and thereby prevents RNA degradation by nucleases. The DA along with RNA, Promega RNA dye (dilution factor indicated in figure captions, e.g., 800×), and buffer (0.1 M PB + 10 mM EDTA) were combined, and the pH was adjusted to ∼7.4 using 5 M NaOH. The mixture was then subjected to one WD cycle in a glass vial, rehydrated with water, readjusted to pH 7.4, and analyzed with fluorescence microscopy. Similar to the HPTS dye encapsulation that we observed in the DA vesicles, encapsulation of RNA in the inner core of the vesicles was observed after one WD cycle (Fig. 4). However, higher concentrations of RNA and prolonged storage after sample preparation both resulted in a significant drop in pH and the formation of clumped vesicles (Supplementary Figs. S6 and S7). These results confirmed that HPTS is a good proxy for encapsulation of low concentrations of larger anionic solutes, such as RNA, and can provide an inexpensive means for further experimentation.
Bright-field (top row) and epifluorescence (bottom row) images of DA vesicles in 0.1 M phosphate buffer (pH 7.35) with RNA and Quantifluor dye (800X) after one WD cycle. RNA is encapsulated in vesicles and remains encapsulated for at least days, after which aggregation was observed (Fig. S6).
We have successfully designed, built, and tested an integrated three-pool prototype terrestrial hot spring pool simulator. Although we have completed initial testing of the system, further features and modifications are suggested for customization and to optimize the efficiency of future experiments, such as fine-tuning the time required for a single WD cycle and improving the temperature response of the system (Supplementary Fig. S8).
Programmed water flow control
Preliminary work for automated fluid control included design simulations of water transfer using peristaltic pumps and solenoid valves governed by PID control, as implemented in LabView in previous iterations. Future integration will involve pump control via Arduino-driven relays, which will allow researchers to program inflow/outflow cycles, emulate WD cycles, and coordinate flow across pools at predetermined intervals or thresholds (e.g., temperature or time-based triggers).
The system design anticipates the need for directional flow between pools facilitated by top-mounted channels and programmable volume displacement. Such features are critical to experiments involving protocell transfer, dilution, and mineral diffusion across a synthetic hydrothermal network.
Rock coatings to simulate hot spring pool surfaces
In the future, we are focused on performing vesicle WD experiments that utilize the mineral-coated surfaces to closely simulate the rocky surface of geothermal hot spring pools. The use of different surface topographies, chemistries, porosities, and surface roughness could influence the versatility of the vesicles’ population and encapsulation of RNA, peptides or other polymers. This added feature of rock-coating in the pools will aid our understanding of how different early-Earth pools harbored vesicles and facilitated the evolution of protocells during WD cycles.
Conclusions
We have designed, constructed, and tested a lab-scale model multipool system that mimics geothermal hot spring pools. The pools were 3D printed using Durable Resin and PLA, and the pools were connected to a temperature and water control system. We used pure DA, DA:DO (4:1), and POPC:POPG (1:1) as model lipids to investigate the formation of vesicles, their morphological variations and ability to encapsulate fluorescent dye (HPTS) under multiple WD cycles. The formation of vesicles occurred in all cups regardless of the type of material the cups were 3D printed from, indicating the inertness of the cups’ surfaces. The cups favored encapsulation of HPTS in the thin-walled oligolamellar and cell-like giant unilamellar vesicles, an observation attributed to the efficient rehydration of the lipids on the cups’ surfaces. Furthermore, addition of 20 mol% DO in DA stabilized vesicles and promoted enhanced encapsulation of HPTS. Similarly, DA-based vesicles were able to encapsulate model prebiotic RNA under WD cycling, which indicated that encapsulation is not unique to HPTS. This model hot-spring pool simulator is ideally suited for experiments focused on understanding protocell formation and polymer encapsulation studies. The modular nature of the simulator will allow for more advanced prototypes to be designed and tested for future experiments with the aim of bridging geochemical complexity with prebiotic chemistry.
Footnotes
Acknowledgments
A.S. thanks the Australian Institute of Nuclear Science and Engineering (AINSE) for financial support through the Early-Career Researcher Grant in 2023.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
A.W. acknowledges support received under the Human Science Frontier Program (RGP0029/2020) and Australian Research Council (DE210100291). A.C.F. acknowledges support from an ARC Future Fellowship (FT220100757) and Discovery Project (DP210102133).
Associate Editor: Mary Beth Wilhelm