
Abstract
Biological evolution confronts situations in which the modification of a trait to improve the performance of one function may diminish the performance of another. Similar trade-offs occur in economics and engineering, where they are evaluated with the concept of Pareto optimality. “Pareto optimal” solutions are solutions such that performance cannot be improved for any task without sacrificing performance for another task. Solutions outside the Pareto optimal set are likely to be uncompetitive in that they could be improved without negative consequences. Biologists have argued that optimization for multiple biological functions restricts the variety of evolutionarily stable phenotypes to a Pareto set within traitspace, while promoting diversity within this set. Here, we consider whether evidence for such optimality could serve as evidence of life in astrobiology. We propose that objects whose properties lie demonstrably within a region constrained by trade-offs between biologically relevant functions are more likely to be biogenic; examples discussed here include bacterial morphology, mycelial networks, and the selection of molecules. Conversely, objects with comparable characteristics to known forms of life that exist outside the relevant Pareto set are less likely to be biological. We conclude that the detection of Pareto optimality may disclose functionality and, hence, biogenicity in unfamiliar materials.
Introduction
Consider the following hypothetical scenario. A number of microscopic objects shaped like highly curved rods, fully mineralized and lacking in any organic matter, are found in a sample of rock from Mars. Some astrobiologists interpret these objects as potential fossil microorganisms on the basis of their shape, and because evolution on Earth has frequently produced curved-rod-shaped motile bacteria whose morphology is thought to balance swimming efficiency, construction cost, and chemotactic efficiency (Schuech et al., 2019). But other, more skeptical astrobiologists object that the Martian rods are much more curved than curved-rod-shaped bacteria on Earth. Now, this objection is weak: Evolution might simply have taken a different course on Mars. However, a stronger argument may be available to the skeptic. Perhaps the Martian rods are so curved that they would demonstrably perform badly at the very trade-offs for which curvature is known to be an adaptation: Swimming, resource cost, and chemotaxis.
Differences between Earth life and Mars life may be contingent, but the underlying physics of bacterial motility and the resulting trade-offs should be the same on both worlds. If the shape of the Martian rods is plainly maladaptive, this seems a good (although not, perhaps, absolutely decisive) reason to discount them as fossils. Conversely, if a large population of Martian rods were found to exhibit diverse curvatures, all within the set of optimal morphologies for swimming, resource cost, and chemotaxis, this would support the view that they are the Martian equivalent of fossil bacteria. This thought experiment leads to the suggestion that the presence (or absence) of adaptive optimality, to the extent that it can be detected and measured, might serve as evidence for (or against) life in astrobiology. We explore this idea here.
Evolution by natural selection has produced a startling diversity of forms from molecular to macroscopic scales. However, this diversity is neither infinite nor random: The life we observe represents only a small subset of the space of all possible forms (McGhee, 2006). One way to conceptualize this is to construct the relevant theoretical morphospace (McGhee, 2006; Mitteroecker and Huttegger, 2009; Budd, 2021), trait space, or phenotype space. Points within these spaces represent possible morphologies or other phenotypic traits that an organism can exhibit. This space can be used to study the evolution of life and its theoretical limits in a quantitative way, including phenomena such as convergent evolution (McGhee, 2006). Relevant trait spaces for astrobiology may include, for example, the morphology of bacteria, the chemical properties of biomolecules, or the network properties of mycelium (Fig. 1).
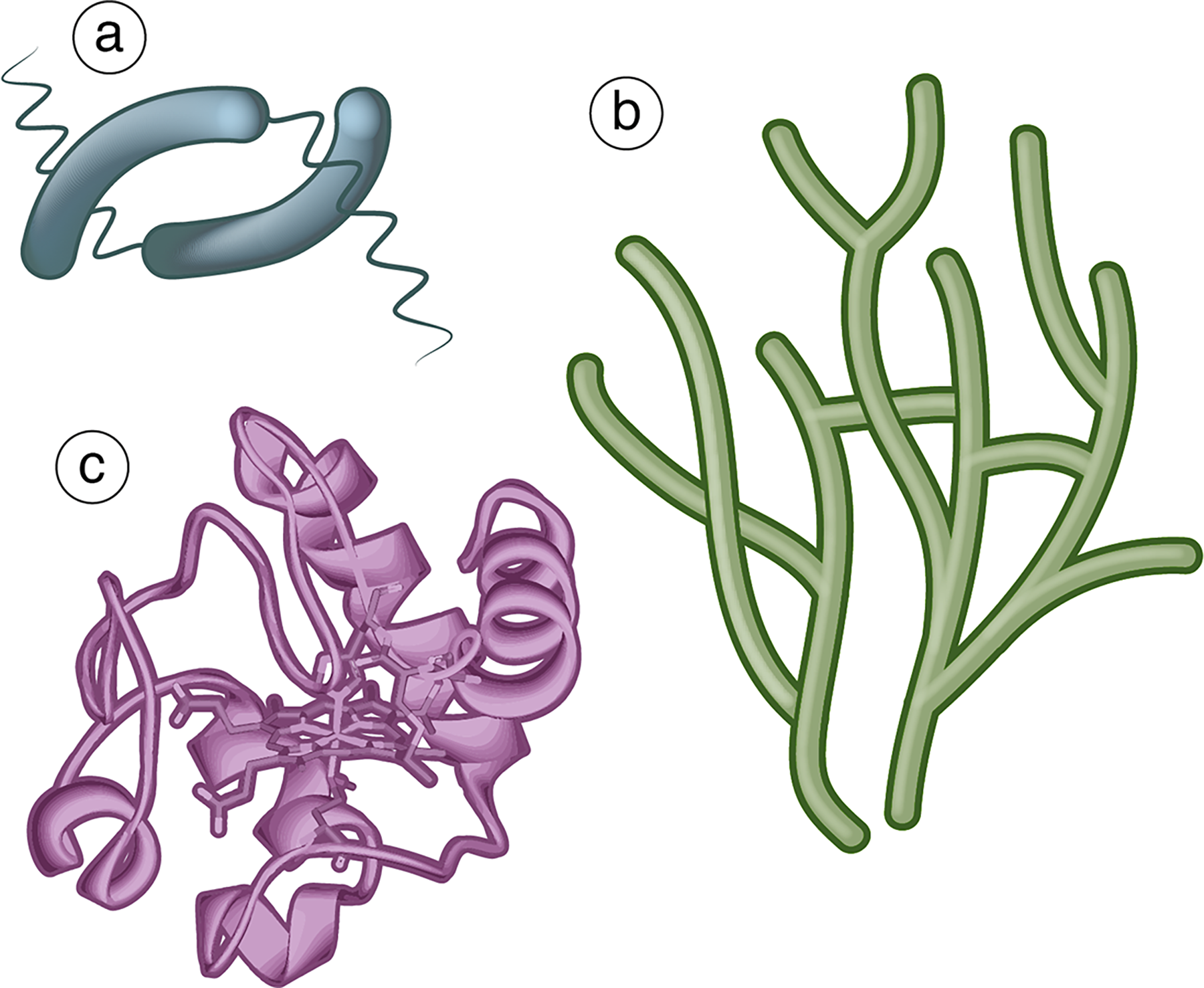
Examples of biological objects subject to trade-offs when optimizing their functional fitness.
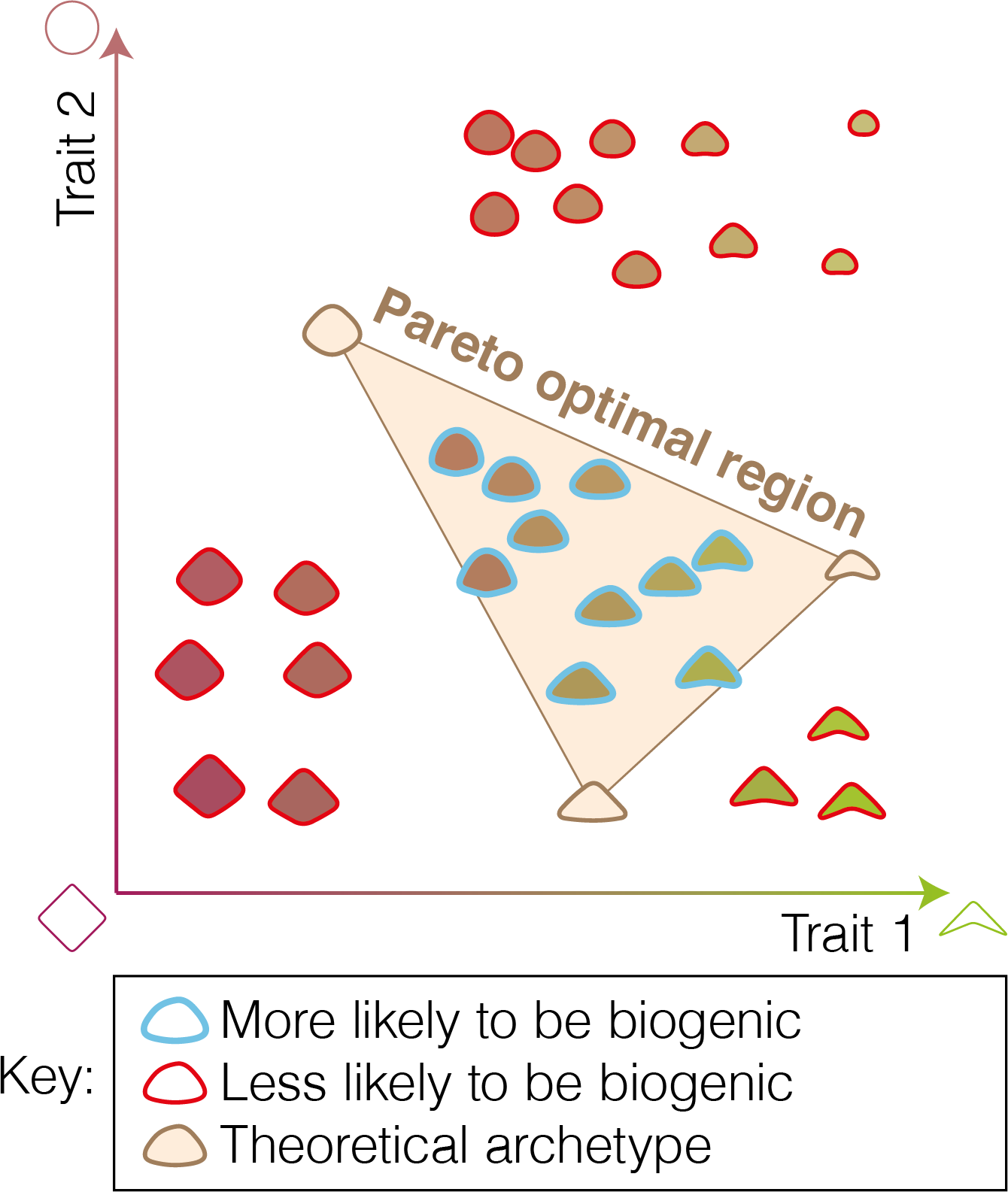
Schematic showing the Pareto optimal region for three tasks in a mock morphospace. Objects inside the region are more likely to be biogenic while objects outside the region are less likely to be biogenic. Archetypes, objects which maximize performance at a single task, are at the vertices of the region.
The boundaries of the region of trait space occupied by life are constrained by various physical phenomena (McGhee, 2006; Cockell, 2018; Johnston et al., 2022; Solé et al., 2024), by the limitations of genetic and developmental machinery, by the fundamental features of life (Lineweaver and Chopra, 2012), by historical contingency (Lineweaver and Chopra, 2012), and by the motion toward regions of higher fitness through natural selection (Banga and Sager, 2024). The evolutionary pressure to increase adaptive fitness is a multiobjective optimization problem that requires trade-offs between different tasks. In general, such trade-offs can be quantified by the concept of Pareto optimality (Shoval et al., 2012; Alon, 2019), which is commonly used in economics and engineering. It can be shown that management of competing objectives restricts the space of possible solutions (Sheftel et al., 2013), which implies that life may be constrained to lie within Pareto optimal regions of trait space. Given that extraterrestrial life is likely a product of natural selection, the notion of Pareto optimality could inform our conceptions of extraterrestrial life and the signatures it might produce.
Assumptions about “optimality” have previously arisen in debates about microscopic magnetite crystals found in the famous Allan Hills 84001 (ALH84001) meteorite from Mars. It was explicitly assumed (Thomas-Keprta et al., 2000,2002) that, if these crystals were biogenic, they would resemble those produced by magnetotactic bacteria on Earth, given that natural selection acting on both systems would solve the same optimization problem in ways constrained by the universality of physics (Thomas-Keprta et al., 2000,2002). More generally, the idea that restricted occupancy of possibility spaces can indicate that biogenicity has often been assumed implicitly or explicitly in astrobiology (McKay, 2004; Brasier et al., 2006; Davies et al., 2009).
Pareto optimality can be defined as follows. Consider a set of
Under certain assumptions (Sheftel et al., 2013), the dimensionality of the Pareto optimal region should be one less than the number of tasks, so for two tasks it is a line and for three tasks it is a two-dimensional object (Sheftel et al., 2013). This implies that the recognition of Pareto optimality may act as a dimension reduction technique. Even in a high-dimensional trait space, if the number of tasks is small, then the phenotypic data points may be placed on a low-dimensional object embedded in the space (Sheftel et al., 2013), consistent with the manifold hypothesis (Bengio et al., 2013) for natural data.
When simplifying assumptions are made, the optimal regions are polytopes with straight edges (Sheftel et al., 2013); however, relaxing these assumptions can lead to more complex shapes (Sheftel et al., 2013), as observed in computed regions for real systems (Schuech et al., 2019).
An archetype is the set of traits that maximize performance at a single performance function related to a single task. The archetypes may be located at the endpoints of the Pareto front if the region is a line or at the vertices that define the higher dimensional shape, depending on the assumptions made (Sheftel et al., 2013; Alon, 2019). If the performance function is maximized at multiple points, then the archetypes may correspond to a set of points (Sheftel et al., 2013). Complex optimal regions can be observed in real data (Pollock et al., 2025). The nature of the archetypes can be used to infer which tasks are involved in evolutionary trade-offs. In biological systems, trade-offs between resource allocation and the performance of beneficial activities are expected to be common.
The proposal that organisms can only be found within restricted, Pareto optimal regions of trait spaces covers a wide range of organism types, scales, and traits, which motivates the expectation that this is likely to be a general feature of life, occurring whenever and wherever environmental and ecological conditions vary slowly enough for the set of relevant tasks to stabilize.
Biologists have claimed (sometimes controversially) that many measured traits of living and extinct organisms lie in regions that demonstrably correspond to Pareto optimality. Examples of this include the morphology of ammonite shells (Tendler et al., 2015), vertebrate jaws (Deakin et al., 2022), and leaf-cutter ants (Shoval et al., 2012); the “sabre-teeth” of extinct animals (Pollock et al., 2025) and beaks of Darwin’s finches (Shoval et al., 2012); the wing aspect ratio and body mass of bats (Shoval et al., 2012); gene expression in tumors (Hausser et al., 2019), human single cells (Korem et al., 2015), and Escherichia coli (Shoval et al., 2012); human brain networks (Avena-Koenigsberger et al., 2014) and wider nervous systems (Pallasdies et al., 2021); language (Seoane and Solé, 2018); the morphology of curved bacteria (Schuech et al., 2019); sperm length and mass (Koçillari et al., 2024); neurons (Jedlicka et al., 2022); life-history traits (Szekely et al., 2015); biological regulatory systems (Szekely et al., 2013; Adler et al., 2022); E. coli growth strategies (Mori et al., 2019); plant (Conn et al., 2017) and fungi networks (Aguilar-Trigueros et al., 2022); species diversification (Lewitus and Morlon, 2016); and polymorphisms (Sheftel et al., 2018). Controversy has focused on the use of particular numerical methods to identify bounded optimal regions in trait data (Hart et al., 2015; Blais and Jeukens, 2024); some methods are allegedly prone to a high rate of false positives in detecting such regions (Mikami and Iwasaki, 2021; Sun and Zhang, 2021). This debate, however, appears unresolved (Adler et al., 2022). Of course, the Pareto framework cannot be applied to all traits, since not all traits are entirely adaptive and many are certainly historically contingent or constrained in some other way (see Gould and Lewontin, 1979, for a classic discussion of this issue). However, the literature above suggests that the Pareto optimality framework can, in fact, explain the distribution of many adaptive traits.
Application to Astrobiological Scenarios
The hypothesis that life tends to produce objects whose traits are restricted to Pareto optimal regions implies a new way to test the biogenicity of candidate biosignatures. Consider an object, substance, or pattern for which a biological interpretation is available and that interpretation implies a corresponding set of biological tasks on which the fitness of the organism depends. In theory, measurement of the relevant properties of the object may reveal that it lies within the Pareto optimal region (supporting the inference of biogenicity) or not (challenging the inference of biogenicity) with respect to those tasks (see Figure 2). Here, we provide some examples of scenarios in which this general approach might be useful.
First, we return to the introductory thought experiment of the hypothetical curved rods. This idea is based on the work of Schuech et al. (2019), which establishes a two-dimensional morpho-space defined by a curvature parameter and an elongation parameter, in which diverse species of curved motile bacteria are placed using data extracted from images.
In the work of Schuech et al. (2019), performance landscapes were computed for tasks related to swimming efficiency, construction cost, and chemotactic signal-to-noise ratio to derive the resulting region of Pareto optimality. It was then shown (Schuech et al., 2019) that all of the species of bacteria lie in or near this region. If we were presented with an object morphologically similar to a curved bacterium from a suitable context, could this region be used to evaluate its biogenicity? The answer depends partly on whether construction cost, swimming efficiency, and chemotactic signal-to-noise ratio are considered to be sufficiently universal and partly on how robustly the Pareto optimal region can be calculated with respect to these tasks given what is known about the environment, for example, whether the relevant medium is water. Still, if the object is far outside the optimal region, that is at least a defensible reason for caution about interpreting it as a motile bacterium. Similarly, if the object lies inside the optimal region, that is a reason for optimism.
Curved biomorphs have been found in polyextreme geothermal environments on Earth (Belilla et al., 2019) alongside curved mineralized microbial remains (Belilla et al., 2019). The curved abiotic biomorphs illustrated by Belilla et al. (2019) (Fig. 4m) appear to be outside the optimal region of Schuech et al. (2019). However, a mineralized microbe (Fig. 4l in Belilla et al. (2019)) also appears to be outside of this region. It is not easy to explain the observed morphologies of the objects in the work of Belilla et al. (2019). It may be that the mineralized microbe in question is not a curved rod-shaped cell but a multicellular filament, and it may have been deformed by postmortem processes. Such context-dependent complications are in general to be expected with real data.
Many biological networks (e.g., slime molds, biochemical or biologically mediated reaction networks, and fungal mycelia) also confront trade-offs. By constructing a trait space from parameters that describe networks (Koutrouli et al., 2020), one can analyze whether they lie in a region of Pareto optimality with respect to relevant tasks (Seoane and Solé, 2015). For example, networks may trade off benefits from improvements in properties like transport efficiency and robustness with construction cost. It has been argued that many observed complex networks lie in a small region of parameter space (Avena-Koenigsberger et al., 2015; Seoane and Solé, 2015), including neural networks (Avena-Koenigsberger et al., 2014), mycelium (Aguilar-Trigueros et al., 2022), and plant architectures (Conn et al., 2017). When presented with a network structure of unknown origin, one might evaluate its biogenicity by considering which functional trade-offs are implied by biological interpretations and then evaluate whether the network lies in or near the optimal region of trait space for these trade-offs. Relevant complex network properties include, for example, small-worldness (short paths and high clustering) or prevalence of hubs. Such properties may place biogenic networks in a distinct region of trait space from abiotic structures or suitable null models, which do not experience this set of trade-offs and, hence, are unlikely to have the same set of properties.
Very diverse network systems subject to trade-offs can be restricted in trait space by Pareto optimality, as shown by the work of Santoro et al. (2018), which presents a multilayer network representation of transportation systems. This optimality approach could complement existing approaches and advance debates about mineralized networks interpreted as fungus-like fossils (Hofmann et al., 2008; Bengtson et al., 2017; McMahon, 2019). The optimality approach could build on the work of Aguilar-Trigueros et al. (2022) on the optimality of fungal networks or complement the use of atmospheric chemical reaction networks as biosignatures (Wong et al., 2023).
Multiobjective optimality could also be considered when evaluating the biogenicity of molecules. For example, it has been argued that the standard genetic code is optimized (or partially optimized) for multiple objectives compared with some theoretical alternative codes (Freeland et al., 2000; Itzkovitz and Alon, 2007; de Oliveira et al., 2015, 2018; Wnętrzak et al., 2019,2018). It has also been argued that the set of proteins used by E. coli represents a Pareto optimal solution to trade-offs related to solubility and hydrophobicity (Koçillari et al., 2018). This highlights how we may expect the molecules used by life to inhabit a small region of the space of possible molecules (McKay, 2004). The small set of proteinogenic amino acids would also be an example (Philip and Freeland, 2011; Ilardo et al., 2015; Meringer and Cleaves, 2017; Mayer-Bacon and Freeland, 2021). However, debates about the standard genetic code highlight potential difficulties; the standard code is likely more optimal than a random code but less optimal than specific codes for certain tasks (Itzkovitz and Alon, 2007; de Oliveira et al., 2015, 2018; Wnętrzak et al., 2019, 2018).
The arrangement and crystallographic properties of intracellular magnetite grains produced by magnetotactic bacteria have also been argued to result from evolutionary optimization with respect to resource cost and magnetic moment (Thomas-Keprta et al., 2000; Faivre and Schüler, 2008; Kopp and Kirschvink, 2008; Lin et al., 2020). Trade-offs likely constrain these biogenic crystals to a small subset of the shapes, structures, and arrangements possible for magnetite. Nevertheless, while magnetite crystals discovered in Martian meteorite ALH84001 were argued by some authors to be biogenic, others have claimed that abiotic processes can match the relevant features (Golden et al., 2004; Jimenez-Lopez et al., 2010; Martel et al., 2012). Future research into the signatures of adaptive optimality in biogenic magnetite may advance related debates (Shen et al., 2023). Claims of optimality also appear in the recent literature on so-called “giant magnetofossils” (Harrison et al., 2025).
As trade-offs are ubiquitous in biology, further research may elicit more examples where adaptive optimality can be used as evidence of life or as a constraint on the possible forms of life. For example, the morphospace of possible extremophiles may be constrained by the fact that resources that could be used for cellular construction may be required for homeostasis in extreme environments. It is relevant that Pareto optimality has been thought to play a role in mechanisms for bacteria to respond to heat shock and to repair deoxyribonucleic acid (DNA) (Szekely et al., 2013). It may also be worthwhile to compare corkscrew-shaped biomorphs, such as those reported by Cosmidis and Templeton (2016) against the optimal region in the morphospace of spirillar bacteria.
Discussion
We propose that evidence for Pareto optimality may be detected in candidate biosignatures and may serve as evidence for biogenicity. This proposal is distinct and complementary to the idea that complexity may be a biosignature (Marshall et al., 2017,2021; Bartlett et al., 2022). Biological objects clearly exhibit complexity, but certain minerals (Krivovichev et al., 2022; Hazen et al., 2024) and many natural patterns that result from far-from-equilibrium self-organization are also notably complex (García Ruiz et al., 2002; Barge et al., 2015; Nakouzi and Steinbock, 2016; Knoll and Steinbock, 2018; Criouet et al., 2021; Cartwright et al., 2024; Jordan et al., 2024; Jenewein et al., 2025). However, biological complexity is not arbitrary but functional (Hazen et al., 2007); complexity is constrained by evolutionary trade-offs and resource-cost efficiency. The search for Pareto optimality in measurable traits is a search for the fingerprints of functional complexity.
Optimality is, in general, not agnostic in the sense of recently discussed “agnostic biosignatures” (Johnson et al., 2018; Chou et al., 2021; Marshall et al., 2021; Bartlett et al., 2022; Cleaves et al., 2023; McKaig et al., 2024). Each Pareto region in trait space is specific to a set of trade-offs between particular biological tasks, and it is not possible to construct a priori the relevant Pareto region for evaluating a candidate biosignature without making assumptions about the nature of these tasks and trade-offs. However, there are two reasons why a search for Pareto optimality may be useful in evaluating candidate signs of exotic “life as we don’t know it” (Grefenstette et al., 2024). First, it is at least theoretically possible that Pareto regions might be detected in multiparameter datasets just from their shape (low-dimensionality polytopes) even before the underlying trade-offs or biological functions are known. It is currently debated whether this has already been accomplished in some biological datasets (Hart et al., 2015; Alon, 2019; Mikami and Iwasaki, 2021; Sun and Zhang, 2021; Adler et al., 2022; Blais and Jeukens, 2024). Second, it can be argued that some tasks and trade-offs, such as resource cost and those arising from thermodynamics and mechanics, are universal in nature. However, while the physics and chemistry of fluids are clearly universal, the constraints imposed by the local environment and available solvents are not and must be carefully accounted for in seeking to define Pareto optimal regions in specific contexts.
The space of functional and efficient objects is clearly a subset of the space of complex objects, and it is likely that a biological object would be drawn from this subset of the space of complex objects. As in the case of bacteriogenic magnetite crystals, there may be some overlap between the space of functional and efficient objects and the space of abiotically producible objects. This overlapping region could present potentially problematic biomorphs likely to cause false positives particularly when accounting for preservation processes (Golden et al., 2004; Jimenez-Lopez et al., 2010; Martel et al., 2012). In general, the more parameters needed to describe a structure, the less likely it is that an abiotic mimic will land in the Pareto optimal region. We note that, as with other indicators of biogenicity, optimality is most robustly assessed in large populations of objects. When the relevant data are morphological, this assessment could complement population-level statistical morphometry along the lines proposed by Rouillard et al. (2021). It would be interesting to discover whether any areas within the optimal morphospaces for microorganisms are particularly difficult for abiotic “biomorphs” to reach. More generally, we speculate that the parameter distributions of populations of some abiotic structures may turn out to be confined to Pareto-like subsets of trait space, which result purely from physicochemical constraints on their formation, but this remains to be tested.
Optimality considerations might also be useful in evaluating candidate technosignatures, as the space of useful, functional, and cost-effective technological artifacts should be much smaller than the space of possible alternatives. Such evaluation would require assumptions about the intended function of the technology in question.
As already hinted, populating trait spaces with real data can be challenging. It may be difficult to assess whether gaps in the data are the result of the suboptimality of a group of traits or from a lack of data. Real biosignatures tend to be at least one step removed from the living organisms that produced them (consider fossils, collections of organic molecules, or suites of atmospheric gases). Thus, intermediating processes may modify the measurable distributions of traits (e.g., fossil preservation modifies the apparent morphology of organisms, and chemical reactions modify the abundance distributions of functional groups within collections of molecules). Such processes may obscure any original fit to a Pareto optimal region in trait space. For complex objects such as stromatolites, abiotic influences may so overwhelm biological optimality that the Pareto approach has little value.
We conclude that the detection of Pareto optimality in a suitable dataset may serve as evidence for life (and the lack of such optimality can serve as evidence against biogenicity, sometimes called an “antibiosignature” (Walker et al., 2018)). Suitable datasets could include measurements of morphology or chemical properties that relate to the performance of the proposed organism at the set of tasks. However, the strength of this evidence depends on how robustly and universally the optimal region can be identified and understood in terms of function, along with the quality of the data, the specificity of the Pareto region within trait space, the goodness of fit between the data and the Pareto region, and of course the likelihood that an abiotic process or null model (Pie and Weitz, 2005) could generate objects with the same parameters, which may differ on a case-by-case basis (Gillen et al., 2023). We hope that this work will provoke further research into how the theoretical parameter space of life is filled and constrained by trade-offs, particularly those that may be universal.
Footnotes
Acknowledgments
The authors thank the handling editor, N. Sleep, for the suggestion to include discussion of magnetite in ALH84001. For the purpose of open access, the author has applied a Creative Commons Attribution (CC BY) license to any Author Accepted Article version arising from this submission.
Author Contribution Statement
N.R.: Conceptualization, investigation, methodology, writing—original draft, and writing—review and editing. C.C.L.: Conceptualization, methodology, and writing—review and editing. H.J.C. and M.A.v.Z.: Funding acquisition, project administration, and writing—review and editing. S.M: Conceptualization, funding acquisition, project administration, investigation, methodology, visualization, supervision, writing—original draft, and writing—review and editing.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
N.R., H.J.C., M.A.v.Z., and S.M. acknowledge the support of the
Associate Editor: Norman H. Sleep