
Abstract
The ambiguity of the Viking lander life-detection experiments left microbiology and microbial ecology out of space missions for 50 years. However, this was largely because the scientific process inherent in the search for life was misunderstood by a space exploration community that wanted clear-cut answers. Since Viking, enormous advances have been made in our knowledge of microorganisms, their biochemistry, and physical and chemical limits. Coupled with improvements in methods for culturing microorganisms and measuring their active metabolism, this has made possible more sophisticated experiments to search for life in extreme environments on Earth and on other planetary bodies. By overcoming misunderstandings surrounding the Viking lander experiments, microbiology and microbial ecology can take center stage in renewed efforts to seek life elsewhere. Furthermore, new knowledge in these fields should be used to overhaul assumptions in planetary protection, and it can be used to achieve permanent human space settlement.
Introduction
When microbiology experiments on other planets were being conceived, leading up to the flight of the two NASA Viking missions and their landers in the 1970s (Lederberg, 1960; Sagan, 1964; Levin, 1965; Imshenetsky et al., 1967), the design of these experiments was based on a more limited set of human-related and environmental microorganisms than we possess today. Furthermore, the detection of extant microorganisms and their component parts (i.e., life detection) was much more rudimentary than it is today and was largely confined to whole-cell assays for pathogens and other microorganisms associated with human-occupied environments. Our knowledge of the microbiology of extreme environments was comparatively limited, although new steps were being taken to culture extremophiles and understand the microbial ecology of extreme environments (Straka and Stokes, 1960; Brock and Freeze, 1969; Gochnauer and Kushner, 1969).
The 50 years since the Viking missions have witnessed astonishingly rapid advances in microbiology.
First, there has been scientific progress, largely in four aspects: (1) Our vastly greater knowledge of the diversity of microbial life. This has allowed us to appreciate the phylogenetic variety of microorganisms on Earth and what we might look for elsewhere (Hug et al., 2016; Shu and Huang, 2022), although our knowledge of the diversity of microbial life is still far from complete. (2) The much greater knowledge about microbial growth at a variety of extremes, including temperature, pH, pressure, radiation, desiccation, and chemical toxins. This allows us to make better predictions of where life might exist and from which environments we think life might be excluded (Merino et al., 2019; Cockell, 2025). An understanding of these limits, as with our knowledge of microbial diversity, will no doubt improve. (3) A much greater range of environments have been characterized with respect to their microbial biota and the energy sources (redox couples) and substrates that microbes can use to sustain themselves, potentially over long time periods (Lloyd, 2021), thus improving our predictive knowledge of where life can successfully maintain itself (Hoehler and Jørgensen, 2013). (4) Our knowledge of the biochemical mechanisms that allow for the diversity and limits of life listed in (1) to (3) has deepened, enabling better insights into what sorts of molecules we might try to detect as evidence of life (Neveu et al., 2018; Chou et al., 2021). Machine learning and artificial intelligence analyses of large quantities of data will aid these efforts (Scharf et al., 2023), as well as the steps that might be taken to evaluate our confidence in life detection (Green et al., 2021).
Second, technological progress has been made in parallel with empirical and theoretical scientific advances encouraged by the need to study more extreme and challenging environments, such as the deep subsurface.
These technological advances have occurred both in the laboratory and the field. In the laboratory, new cultivation methods have enabled a greater range of organisms to be enriched, studied, and detected (Zengler et al., 2002; Lewis et al., 2021; Salam et al., 2021). The characterization of microorganisms has been enabled by improvements that include, but are not limited to, developments in molecular sequencing methods, cell detection, and biochemical characterization methods (Hedlund et al., 2014; Tighe et al., 2017; Satam et al., 2023). In the field, miniaturized methods for the detection, isolation, and biochemical study of microorganisms (including sequencing) have become available (Kallmeyer, 2011; Roggo and van der Meer, 2017). Although still in their early development, the low energy, low mass, miniaturized design, and ruggedized nature of this field apparatus has obvious implications for translating this technology into life detection apparatus on spacecraft.
On the 50th anniversary of the Viking landers, it is time for microbiology and microbial ecology to take center stage in the search for life beyond Earth and for microbiologists to play a prominent role in the development of instruments and missions. I suggest that planetary protection needs an overhaul and should be tackled from a more empirical microbiological approach than current approaches. The emerging use of microbiology and microbial ecology in space settlement is similarly developing at a rapid pace. In summary, we could recognize a triumvirate (Fig. 1) that encapsulates the maturing expansion of microbiological sciences beyond Earth.
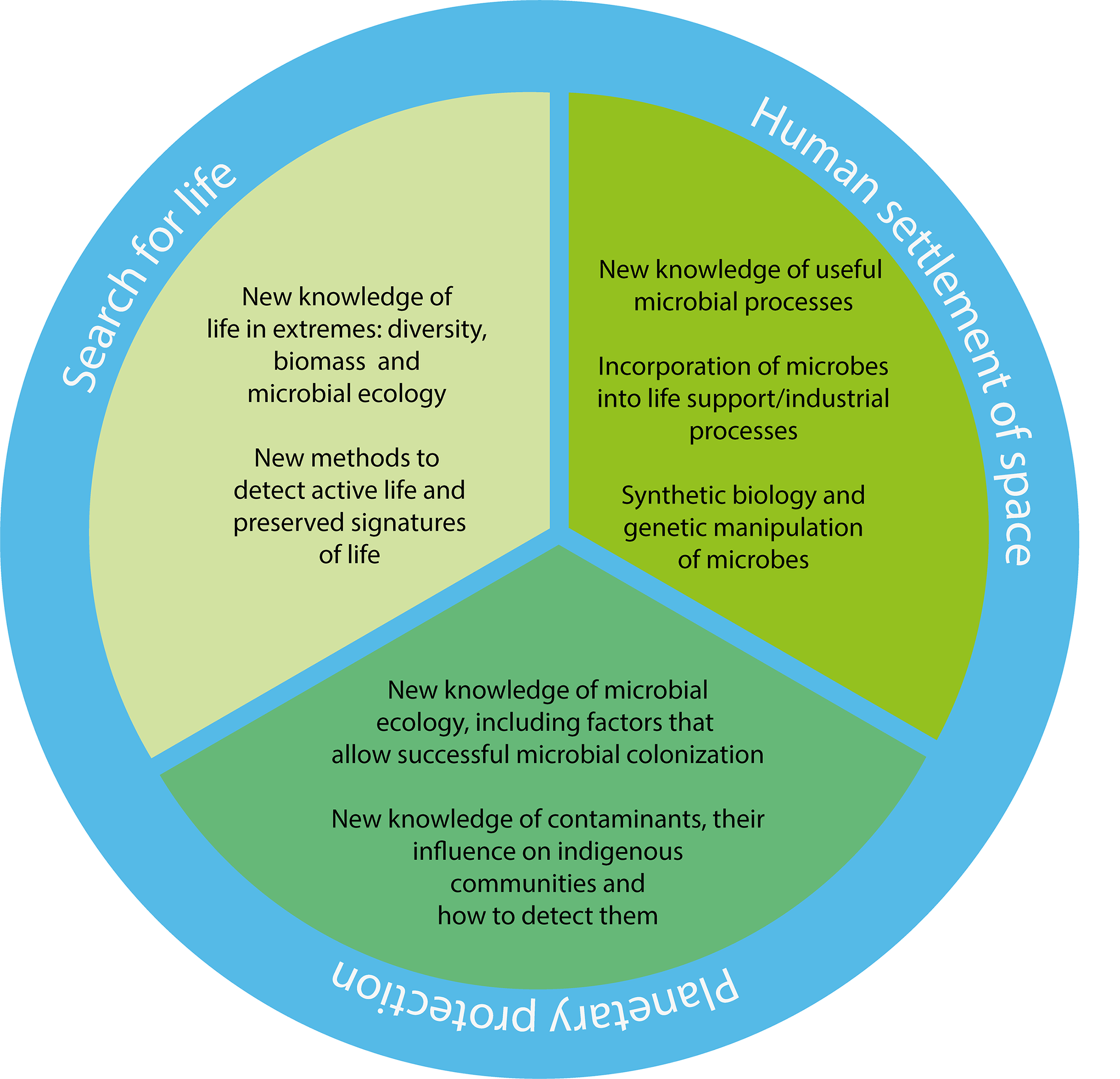
A microbiological triumvirate. Fifty years after the Viking missions, advances in microbiology and microbial ecology can be applied to at least three different areas of space exploration and settlement.
An unfairly maligned mission
Before discussing the modern promise of microbiology in space, it is necessary to slay the dragon that has breathed fire on (and successfully incinerated) almost all attempts to introduce microbiology into space missions since the Viking missions.
The Viking experiments were outstanding in their scientific design and use of controls (Klein, 1978). They drew on the existing knowledge of microbial metabolism to test for heterotrophy, autotrophy, including photosynthesis, and to seek evidence of carbon fixation associated with life. Technically, the life-detection apparatus on Viking was remarkable. It was packed into a space of 28.3 dm3 and had a mass of 15.5 kg, making it the first miniaturized set of instruments to seek life on another planet. The experiments worked. The device returned the data it was designed to obtain.
However, the results of the experiments were not conclusive on the question of whether there were actively replicating microorganisms in the Martian soil. Note that this fact does not contradict the previous paragraph. An experiment that works perfectly does not guarantee that its results will provide clear answers, let alone ones desired by the observers or their funders. Other papers have adequately addressed the Viking results (Klein et al., 1976; Levin and Straat, 1977; and, more recently, McKay et al., 2025). It is not my intention to discuss those experiments here or to express a view on whether the landers did or did not detect life. There is a more general point about this controversy with respect to what happened to microbiology afterwards. The ambiguity in the results led to a retreat from continuing similar experiments, despite the fact that ambiguous results in good experiments should instead lead to more experimentation. Prevalent was a sense that the experiments had “failed.” Several corrective observations about how those experimental results have been interpreted by the wider community may be useful.
Since the Viking lander experiments, we have learned that the detection of life is a complicated task. Three factors are important: (1) Small quantities of active biomass are often spatially difficult to detect. The physically small size of microorganisms (prokaryotes are typically on the order of a micron) mixed in with soil and rocks makes their direct detection challenging. (2) Small quantities of active biomass have low concentrations of biomolecules in a given volume of the substrate in which they are found. This challenges the detection limits of our instruments. (3) Active microbial life at low biomass often produces disequilibria or other chemical perturbations to the environment that are difficult to distinguish from the abiotic background.
These limitations mean that, in many locations on Earth, there are debates about the presence of actively metabolizing life. Studies in the Atacama Desert have resulted in a series of papers that have reported contrasting accounts of microbial activity in extremely desiccated soils (Navarro-González et al., 2003; Drees et al., 2006; Connon et al., 2007; Crits-Christoph et al., 2013). In Antarctica, microbial activity in dry permafrost near the limits of life generated controversy about the status of actively metabolizing microorganisms (Goordial et al., 2016,2017; Wood et al., 2024). Ice-free, arid, salty soils in Antarctica have similarly generated uncertainty as to whether they support actively metabolizing life (Dragone et al., 2021).
In deep marine and subsurface continental low water activity and high chaotropicity brines (Hallsworth et al., 2007; Payler et al., 2019), controversy surrounds the presence of cells or biomolecules, such as RNA, whose origin from in situ metabolically active cells remains unclear (Hallsworth et al., 2007; Yakimov et al., 2015; Steinle et al., 2018). Similarly, analyses from hypersaline, near-surface, low-water-activity pools, such as Don Juan Pond in Antarctica (Meyer et al., 1962; Cameron et al., 1972; Peters et al., 2014) and ponds in the Dallol Rift, Ethiopia, where low water activity and high chaotropicity brines in geothermal pools can be found (Belilla et al., 2019), have resulted in similarly equivocal microbial activity.
The microbiological community’s response to these challenges is not to declare the ambiguous results as “unsuccessful” in the way in which Viking was treated. Instead, scientists return to these field sites with better instruments. If they continue to fail to resolve the ambiguity of active life, they continue on a journey of improving instruments and refining knowledge of the lowest detection limits to life until they arrive at an answer.
A second important point is that often experiments seeking to detect life reveal the potential for false-positive or false-negative signatures that subsequently improve our understanding of the abiotic baseline. For example, in the case of the Viking experiments, we learned that the addition of aqueous solutions to Martian soils could potentially result in the release of oxygen (Mazur et al., 1978), similar to what would be expected from the activity of microorganisms capable of oxygenic photosynthesis. This potential false-positive signature of photosynthetic autotrophy is an important chemical characteristic of the Martian environment that would have been difficult to predict a priori.
In recent years, we have also learned that the potential for perchlorates to destroy organic material at high temperatures could lead to a false-negative detection of life on Mars (McKay et al., 2025). Therefore, we must consider the presence of perchlorates when developing life detection technologies for Mars that avoid false-negative results.
When seeking life in environments with different physical and chemical characteristics to many environments on Earth, it may take several attempts and several missions to establish the abiotic baseline from which to detect biology. In cases of extremely low biomass, similar to our experiences in extreme deserts on Earth, we may need to iteratively improve our theoretical frameworks and observational methods before reaching a definitive conclusion. One could therefore argue that scientists should focus on designing missions to explore the chemical environment until the abiotic baseline has been satisfactorily explored, then turn their attention to life detection. However, it is often only by deliberately seeking the signs of life that we discover how the chemical or physical characteristics of an environment might masquerade as biological signatures. We often underestimate the potential diversity and complexity of the abiotic baseline of physical and chemical processes and the diversity of microbial metabolic possibilities. Only by searching for life can we attempt to advance both our understanding of this baseline and the potential for life.
The programmatic implication
From a programmatic point of view, biology rarely lends itself to the (often) black and white demands of science traceability matrices, through which high-level mission objectives are linked to what a mission will accomplish (Weiss et al., 2005). Scientists and mission planners should embark upon life detection missions with an understanding that a definitive conclusion from a single mission may not be possible. This may ultimately require a change in the culture surrounding mission science traceability matrices. A corresponding moderation of public expectations from life-detection missions, along with a clear explanation of why an iterative series of missions may be required, is essential.
The scientifically correct response to the Viking life-detection results would have been to plan a follow-up mission in the late 1970s with improved elemental detection capabilities (especially for low-mass elements, including carbon), more metabolic substrates, and other methods to test the hypothesis that certain signals were chemical rather than biological in origin. The nonexistent Viking Lander 3 was a missed opportunity for NASA to kick off a series of missions that today would have put us in a very different position with respect to assessing Mars as an abode for life. By now, we might either have found life or have robustly concluded that the surface and near-surface regions of Mars are lifeless (with a vastly improved understanding of its chemical and physical environment). We can correct that error by embarking on the next iteration of life-detection missions. If we find a new set of ambiguous results, we must use these results to develop hypotheses, and improved instruments and missions, to address those uncertain results, inching forwards until we find the answer.
Another consequence of the programmatic miss-direction following the Viking landers should be corrected. This is the tendency to exclude microbiologists from planetary missions because of concerns that these missions will be interpreted as “Viking-like” life-detection missions. In some missions, microbiologists cannot make direct microbiological observations, for example, if they are geologically focused planetary missions. However, by being involved in mission observations, microbiologists can learn about the physical and chemical environment of an extraterrestrial location, which then informs the development of life detection instrumentation. This experience may be used to propose and develop better missions and instruments to search for life in the future. Always have at least one microbiologist and a microbial ecologist on the science team of your planetary mission, whatever its scientific focus and whether it be orbiters or landers.
Time for Microbiology and Microbial Ecology in the Search for Life
Other papers in this issue set out the methods that microbiologists have developed since the 1970s to detect life. In particular, a recent Keck Institute for Space Studies Workshop considered the search for extant life on Mars and new concepts for instruments that draw upon advances since the Viking era (Perl et al., 2026). These papers provide evidence that microbiology has now reached a state of sophistication with which we can attempt a new search for extant life on Mars and elsewhere. I refer the reader to those papers.
It is significant that many of the suggestions for instrumentation build upon the Viking lander biology experiments, which emphasizes how well designed the Viking experiments were and how they provided a foundation and direct line of heritage to new efforts. Novel methods for culturing a wide range of organisms using many new redox couples that had not been identified in the 1970s have become possible (Zengler et al., 2002; Lewis et al., 2021; Salam et al., 2021). More sophisticated and sensitive approaches for detecting the metabolic products and processes of life have become available (Perl et al., 2026). In their scientific rationale, these instruments advance beyond, yet build upon, the Viking experiments, which used the production of gases and the fixation of carbon as proxies for microbial activity.
The search for “agnostic” life signatures has become a focus in recent life-detection concepts (Johnson et al., 2018; Chou et al., 2021; Georgiou et al., 2023). The Viking biology experiments sought to detect a range of gases from metabolic processes that were independent of the types of organisms that might be responsible. The labeled-release experiment made no assumptions about the nature of the biochemistry of active organisms (Levin and Straat, 1977), other than they would use electrons derived from organic matter as free energy. In that sense, they were agnostic to the biochemical structure of life. Today, we can improve the range and types of metabolic products and processes that we seek by using the much greater knowledge we have of microbial metabolisms in extreme environments (Hystad et al., 2025; Perl et al., 2026).
Alongside fundamental microbiology, the search for life in the universe can be aided by the enormous advances in the science of microbial ecology since the 1970s. Studies of extreme deserts on Earth have revealed the patchiness of life and have improved our understanding of the biogeographical distribution of microorganisms (Warren-Rhodes et al., 2007, 2023; Duan et al., 2022; Prescott et al., 2022). The patchiness of life may imply that we need to plan missions to sample in multiple locations and not in a single location that may be, by bad luck, a region of biologically depauperate substrate. We require many more samples and larger masses of samples than those investigated by the Viking landers. Warren-Rhodes et al. (2022) note that <23 grams of Mars have so far been directly analyzed in situ, including for the presence of life, by Mars missions. This is a quantity that would typically represent a single sample from a single site in many terrestrial microbiology analyses. A robust statistical understanding of the distribution of life in different extreme environments, its biomass, and its metabolic activities can improve our ability to design missions that adequately sample at different spatial scales to help us achieve a definitive conclusions with regard to the presence of life. These data can be used to refine robotic sampling (including sample return efforts) or human sampling strategies. Thus, one productive direction of research is to more thoroughly link microbial ecology with the development of life-detection missions.
If, at the end of this effort, we find no life or environments that are habitable with respect to known life but uninhabited, these discoveries themselves are significant to microbiologists and microbial ecologists (Cockell, 2025). They will improve our understanding of the biogeographical distribution of life on an interplanetary scale and how habitability can be decoupled from the presence of life, with implications for the abundance of both habitable conditions and life in the universe.
Time for Microbiology and Microbial Ecology in Planetary Protection
Another concern in space sciences that intersects with microbiology is planetary protection. This, too, is often rooted in assumptions that first emerged in the Viking era, and it is worth revisiting some of these assumptions (Fig. 1).
Planetary protection emerged from the 1967 Outer Space Treaty’s requirement that states prevent the “harmful contamination” of outer space environments. It was encouraged by a growing realization in the scientific community that contamination of other planets could compromise scientific investigations by, for example, generating false-positive detections of life or compromising local indigenous biota (Lederberg, 1960). In recent years, this practical concern has been augmented by a more ethically motivated concern about compromising the integrity of indigenous ecosystems and biospheres, if they exist (Rummel et al., 2012). However, Lederberg (1960) explicitly considered the ethical dimension of planetary protection. The procedures and processes that have emerged from this have been overseen by Committee on Space Research (COSPAR) (Kminek et al., 2020; Coustenis et al., 2023; McKaig et al., 2024).
Since the beginnings of planetary protection, a central plank of the effort to develop policy has been a probabilistic approach to the assessment of contaminating other planets (forward contamination) and bringing extraterrestrial life back to Earth (back contamination). For forward contamination, this method assesses various component probabilities that a microorganism will be transferred to another planetary body and replicate there. These combined probabilities yield a final probability that is then assessed against some threshold of probability deemed acceptable to the community. These probabilities are often combined into a single equation; the original formulation is referred to as the Coleman–Sagan formulation (Sagan and Coleman, 1966; Barengoltz, 2005).
There are several observations to be made about this approach. First, many of these probabilities, such as the probability that a certain type of microorganism will be present on a spacecraft, are difficult to determine empirically with any certainty. Many of these probabilities are not independent; for example, the fraction of organisms that survive the cruise phase of a mission will depend on the types and fraction of organisms that survive the bioburden reduction regimen.
These difficulties were noted by a National Academy of Sciences committee that considered the planetary protection of icy solar system bodies (National Research Council, 2012). They stated: “The lack of independence for many bioload reduction factors and the minimal precision obtained when assigning values for the initial number of microbes within or on a spacecraft compromise the utility of the Coleman–Sagan formulation as a framework for incorporating planetary protection requirements into mission design.” The report went on to conclude that “current knowledge makes it impossible to confidently assign values for these factors within orders of magnitude of their true value. Multiplication of uncertain overestimates of bioload reduction factors can lead to unsubstantiated, low estimates of likely contamination.” (These concerns seem important, and it is interesting that since they were published in 2012 they have been largely ignored by the planetary protection community. 1 ).
Even if we do obtain a total probability estimate, it is difficult to understand how this can be used in any meaningful way. For the sake of argument, consider that we work out the probability that a particular microorganism will be transferred to Mars and will replicate there to be 0.0367. We then spend several million dollars carrying out studies on that microorganism, the limits of its growth, its ability to attach to spacecraft surfaces, its prevalence in spacecraft clean rooms, and so on. At the end of that research effort, we discover that the probability that it will be transferred and replicate on the Martian surface is now 0.0528 rather than 0.0367. So what? Even if this number had any microbiological meaning, it can only be useful from a policy point of view if it is compared with some probability threshold that is deemed to be acceptable by the community. Herein lies a second problem. A probability widely used by COSPAR in planetary protection studies is 10−4 as a probability threshold above which the contamination risk is considered to be unacceptable (Sherwood et al., 2019). However, this value was established based on mission success probabilities in the early years of spaceflight and has little foundation in microbiology (for a history, see Sherwood et al. [2019]).
Requiring engineers to make multimillion-dollar commitments to the cleaning of spacecraft based on questionable probabilities, which themselves are assessed against a probability threshold that is not based on microbiological principles, does not seem an optimal approach, even if these demands come from a motivation of precautionary concern.
The probabilistic approach to planetary protection suffers from a potentially bigger problem rooted in microbiology. Although the chances of a given microorganism being on a spacecraft might be estimated probabilistically, the final term in the Coleman–Sagan formulation, the probability of growth on a planet (Pg), does not reflect the realities of microbial growth.
If the growth of a microorganism was probabilistic, then over the last four centuries of microbiology, microbiologists would have obtained empirical probability data for given microorganisms and their growth in a particular substrate. If a scientist wanted to grow a microorganism in a culture medium, they would consult tables of probabilities assembled by microbiologists. For the sake of argument, they might find in their table that the probability of growth of microorganism X in a medium of interest is 0.1. Every time they wanted to grow this microorganism in the laboratory, they might make up 10 flasks using that medium in the hope that perhaps one of them would yield growth. However, this is not how microbiology works. Microbiologists make up a medium, they place their organism within it, and it either grows or it does not. Although probabilistic approaches can be used to model certain microbial growth phenomena, for example, the germination of spores (Li et al., 2025), the suitability of a microbial growth medium to support growth for a given organism is largely a binary property. Thus, Pg is, for practical purposes relevant to planetary protection, usually 0 (it does not replicate in a given environment) or 1 (it replicates).
The planetary protection community often assumes that the probability of growth is 1 (although not because the problem mentioned above is recognized, but rather out of a conservative approach to planetary protection) and then concerns itself with determining the other factors in the Coleman–Sagan formation to try to reduce them to an acceptable level (Brener and Cockell, 2025). In some sense, they escape the problem inherent in Pg, but nevertheless, the problems with the other terms in the formulation noted in the National Research Council report still apply.
The National Research Council study (2012) concluded with the following recommendation: Approaches to achieving planetary protection should not rely on the multiplication of bioload estimates and probabilities to calculate the likelihood of contaminating Solar System bodies with terrestrial organisms unless scientific data unequivocally define the values, statistical variation, and mutual independence of every factor used in the equation. Approaches to achieving planetary protection for missions to icy Solar System bodies should employ a series of binary decisions that consider one factor at a time to determine the appropriate level of planetary protection procedures to use.
Can our knowledge of microbiology and microbial ecology developed since the Viking days be used to upgrade planetary protection? Accepting the binary view of the National Research Council study (2012), one direction would be to take a conservative approach and assume that, at some point, in the case where spacecraft are not completely cleaned of all microorganisms, the probability of a microorganism arriving on a planetary body and then replicating in a given extraterrestrial environment is 1. One could then ask: Assuming this to be the case, would this critically compromise future scientific experiments, potential life on the planet, or pose a risk to human health and safety (for example, on Mars)? This relieves us of the Coleman–Sagan formulation and allows us to advance the discussion.
Consider Mars. There are some environments that can help us address the question of what would happen to microbial contaminants and what their effects would be on local microbial ecosystems and, thus, on potentially future scientific investigations. One such environment is Antarctica.
A study of contaminant microorganisms in several field sites in Antarctica yielded little or no detectable evidence of either Escherichia coli or Clostridium clostridiforme in either legacy sites (Canada Glacier camp site, occupied from 1976 to 1988) or in presently occupied sites such as Lake Fryxell summer base camp, where modern cleanliness protocols are implemented (such as removal of waste). In that study, the only conclusively contaminated site was the sewage outflow of Scott Base (Sjöling and Cowan, 2000). Similarly negative results were obtained in attempts to amplify bacterial DNA from the environment around Halley Research Station (Upton et al., 1997).
The study of human commensal microorganisms in the soils of the McMurdo Dry Valleys (Victoria Land) of Antarctica (Ah Tow and Cowan, 2005; Hawes et al., 2025) showed the presence of Staphylococcus epidermis in areas of high human activity, yet undetectable numbers in nearby pristine regions. This suggests that these microorganisms were unable to survive in the environment and that their distribution was local. However, in simulated inoculation experiments, these researchers did show that although organisms could survive no more than 7 weeks from the time of inoculation, the DNA associated with them could persist for longer.
The localization of microbial contaminants is consistent with the study of Clostridium perfringens in outflow liquids from Murdo Station, Antarctica. These organisms were detected within a few hundred meters of the station but were undetectable 3 km away from the station (Edwards et al., 1998). Even this localization is remarkable given that, in the past, the station discharged between 58,000 and 300,000 L of untreated sewage into McMurdo Sound over a 7-month period each year (Lisle et al., 2004). Clearly, for robotic and human explorers on Mars, this situation is avoidable, and the corresponding contamination would be considerably less.
Similar qualitative conclusions have been obtained in the Arctic, where a study of microbial dispersal during a crewed traverse was found to be minimal in the surrounding environment, despite the vehicle harboring diverse microbial taxa (Schuerger and Lee, 2015).
In a frozen state, as might be relevant to the surface of Mars, microorganisms such as Bacillus spp. spores have been found in pony dung frozen for more than 80 years at the Antarctic base camps associated with Shackleton and Scott (Nedwell et al., 1994). Under the surface of Mars, we might expect spores protected from UV radiation and in a desiccated state to survive for many decades, but these organisms would not be reproducing and could be distinguished from indigenous biota, just as they can readily be distinguished in Antarctica.
Bearing in mind that Antarctica, even in the Dry Valleys, is more clement than the surface of Mars with respect to liquid water and organics availability, these data suggests that contamination is likely to be highly localized, potentially reversible, and that, with cleanliness protocols on Mars being much more rigorous than those in the early years of Antarctica research, much of this contamination can be prevented in the first place.
It is interesting to note that during five decades of human exploration, no paper has yet been published from Antarctic Dry Valleys research claiming that a scientific study of the indigenous microbial communities has been either compromised or rendered impossible by previous contamination. There is no persuasive evidence that widespread disruption of ecosystems has occurred from either human-associated contaminant organisms or the presumably vast number of other nonhuman-associated microorganisms attached to the surfaces of helicopters, tents, scientific equipment, and all the other unsterilized paraphernalia of exploration. There remains the possibility that we do not know whether the communities have been compromised because we do not have measurements taken before human presence. However, the microbial community information that has been published shows taxa consistent with other pristine polar sites (Anderson et al., 2025).
Our knowledge of microbial contamination and the potential for displacement of indigenous microorganisms by contaminant microorganisms is still underdeveloped. There are many caveats to the observations made above, including the limited number of studies and samples. The studies cited above primarily involve the study of nonindigenous human-associated microorganisms. It would be wrong to counter some of the dogmatic concerns of planetary protection with an equally dogmatic statement that no terrestrial microorganism could ever threaten the existence of a community of Martian microorganisms, especially as data are sparse on the introduction and establishment of contaminant, noncommensal organisms adapted to life in extreme environments.
For example, Fairén and Schulze-Makuch (2013) suggest that if terrestrial life has already arrived on Mars in the past via material launched and transferred by asteroid and comet impacts, then we do not need to be concerned about taking it there on spacecraft. However, the existence of Martian life related to terrestrial life would not rule out a disruptive effect of newly introduced terrestrial microbiota on existing Martian ecosystems. On Earth, we know that the introduction of some nonindigenous invertebrates can potentially have disruptive effects on ecosystems at the limits of life (Hughes et al., 2020).
Considering these knowledge deficiencies, but adopting a microbiologically conservative view that we will contaminate Mars, especially when humans venture there, we are led to a different set of knowledge gaps than those invoked by the Coleman–Sagan formulation. These include the following:
How do nonindigenous microorganisms become established in microbial ecosystems? What founder and priority effects and principles control their success at becoming established in extreme environments, including nonlinear effects, and are they predictable? How do microbial community dynamics work in extreme environments, and what roles do dormancy and other factors play in the distribution, biomass, and function of life and introduced organisms in extremes (Kawata, 2002; Tscherko et al., 2003; Nicol et al., 2005; Caruso et al., 2011; Cutler et al., 2014; Kelly et al., 2014; Debray et al., 2022; Wetherington et al., 2022; Brener and Cockell, 2025; Maurice et al., 2025)? Would these principles apply to organisms with different biochemistries (i.e., the putative indigenous life) if terrestrial life were to come into contact with it, and what reasons, if any, would we have to believe that the fundamental principles would be different? (The answers to this question have implications for the more speculative scenario of the back contamination of terrestrial ecosystems with extraterrestrial life.) In addition to planetary protection, these advances in microbial ecology will inform the search for indigenous life. Assuming the presence of reproducing microorganisms under optimal Martian conditions (protected from surface radiation and with liquid water), for example in so-called “special regions” (Kminek et al., 2010), what biomass can we expect to observe over given periods of time (Van Horn et al., 2013)? Given the presence of nonindigenous microorganisms, what limits their spread, and what factors are responsible for limits to microbial disruption, as seems to be observed in Antarctica and the Arctic? How are patchiness (biogeographical distribution) and the functions of microorganisms, including dormancy, survival, and resuscitation over temporal and spatial scales, determined in extreme environments (Warren-Rhodes et al., 2007,2023; Duan et al., 2022; Prescott et al., 2022)? Given answers to (1) to (4), what contaminant microbial numbers could we expect in a sample, and would they be detectable by our instruments? Could we detect indigenous biomass reliably with this background cell number? What sampling number, quantity, and across what time frame would we need to detect them?
From the point of view of human exploration, knowledge gaps might include the following:
What methods can we use to mitigate microbial growth in a localized environment (for example, regular soil turnover to expose soils to UV radiation and desiccation), in other words, to prevent effectively irreversible contamination (McKay, 2009)? How might surface extremes act to limit microbial spread and survival (Schuerger and Lee, 2015)? How can we effectively document microorganisms introduced onto Mars through regular environmental monitoring? If localization of microbial contamination can be shown, could one solution be to designate some areas as sites of special scientific interest to maintain them in a pristine condition, similar to the motives behind the Antarctic Specially Protected Area (ASPA) concept, while allowing robotic and human exploration in other areas (Wright et al., 2024)? These areas would be different from COSPARs “special regions” in that not all areas where microorganisms could potentially replicate might be designated off bounds.
Some of these knowledge gaps are already the focus of attention in Antarctica (Hughes et al., 2015, 2018, 2025) and could be applied to extraterrestrial bodies. As well as microbial cleanliness, they can be applied to concerns about chemical contamination (Kennicutt et al., 2010). Certain knowledge gaps, such as a desire to learn more about what microorganisms can be transported on spacecraft (Coustenis et al., 2023) remain valid under this binary paradigm, since we want to know what microorganisms we might introduce into an environment, but they are not motivated by the need to establish probability estimates.
At least one criticism of this approach could be that we know that all organisms we examine on Earth are terrestrial. On Mars, we might have ambiguity in whether the organisms are novel Martian organisms or terrestrial contaminants. On Mars, we might detect contaminant organisms from Earth that are not yet characterized on Earth, leading us to falsely conclude they are novel Martian organisms (see Rummel and Conley, 2017 as a response to Fairén and Schulze-Makuch, 2013 and Fairén et al., 2017). These are valid concerns, but at the same time, they are the very problems that planetary protection efforts should address.
The possibility that contamination would stymie our ability to detect indigenous life was a greater concern to planetary protection in the Viking era than now because molecular characterization methods had not so well developed. It was more difficult to separate contaminant organisms from indigenous organisms. This has often led to a view that we should attempt to completely prevent the contamination of extraterrestrial environments, especially environments where organisms might replicate.
Since the Viking era, sophisticated sequencing methods allow us to differentiate contaminants from indigenous biota. In extreme environments with low biomass, it is usually inescapable that there are contaminant DNA sequences, such as from the sampling apparatus or the kits used to sequence the DNA of the microorganisms. Microbiologists use databases to remove contaminant sequences from environmental samples, thus achieving separation of contaminant from environmental DNA and avoiding false positives (Laurence et al., 2014; Sheik et al., 2018).
In the case of observing a DNA sequence associated with an environmental (nonhuman commensal) microorganism, we could study whether that sequence is observed in any other samples and the number of those sequences, allowing us to assess whether it is likely to be a contaminant. These methods are not failsafe, and they still present challenges, especially in low biomass settings; nevertheless, they have greatly advanced since the Viking experiments (Eisenhofer et al., 2019; Karstens et al., 2019; Weyrich et al., 2019). These methods can be applied to molecules other than DNA, for example lipids (Hedrick et al., 2007). New life detection missions can motivate their further development.
The binary approach can allow us to make more pragmatic decisions on contamination. On Mars, the lack of a vigorous hydrological cycle suggests that, like regions in Antarctica, contamination could remain localized and therefore manageable, even in the case of local microbial replication. However, the contamination of a contiguous water body (such as an icy moon ocean; Doran et al., 2024) with organisms capable of replicating could cause irreversible contamination of an entire ocean. In this case, we might choose to achieve complete sterilization of spacecraft destined to land in these environments until we have carried out a more thorough investigation of the biological status of such an ocean and whether its physical and chemical conditions overlapped with known microbial growth on Earth. Note again, these decisions can be made in the absence of probability assessments (National Research Council, 2012).
As the human presence in space expands, we will inevitably take microorganisms with us. Rather than considering them as dangerous contaminants, or at best passive hitchhikers, it has become increasingly clear that microorganisms can be used in a variety of ways to augment the human settlement of space (Lopez et al., 2019; Koehle et al., 2023). These growing efforts represent the third part of a microbiological triumvirate (Fig. 1).
Microorganisms have long been understood to be a useful components of bioregenerative life-support systems (Roberts et al., 2004; De Micco et al., 2023), but in the last decade, with the expansion of the sciences of genetic engineering and synthetic biology, microorganisms have been proposed in other applications.
Microorganisms can be used as miniature factories to produce useful compounds, from building materials to pharmaceuticals (Llorente et al., 2022; Santomartino et al., 2023). Photoautotrophic microorganisms can be used to synthesize oxygen and biomass (Verseux et al., 2021).
Microorganisms can recycle materials in space and achieve a more efficient reuse of elements that might be valuable in space (such as carbon locked up in materials such as food waste and plastic packaging). Their ability to secrete organic acids, metal-chelating compounds, and a variety of other biochemicals that play a role in the natural dissolution and sequestration of nutrients from rocks gives them roles in processes such as biomining or soil formation (Cockell, 2010). Many of these new approaches and potential uses of microbiology and microbial ecology to support human space settlement have been adequately reviewed by others (Santomartino et al., 2023).
Alongside the growing quantity of work on the applications of microorganisms in space production and settlement, there is a rapidly developing knowledge of the human microbiome in space, its modification by space conditions, and the health and contamination implications of microorganisms and microbial ecosystems in spacecraft and surface stations (Siddiqui et al., 2021; Etlin et al., 2024). This area of work is connected to industrial production and space settlement in that similar microbial principles are involved: the characterization, manipulation, and control of microorganisms in space environments.
Thus, since the Viking mission, not only have microbiology and molecular ecology advanced in ways that can contribute to the search for life beyond Earth and the protection of other planets, but these fields can increase the chances of successfully establishment of permanent human settlements beyond Earth. Any design of spacecraft, space stations, or bases on other planetary bodies that support humans should always involve microbiologists and microbial ecologists.
Conclusion
Microbiology and microbial ecology have largely been left out of planetary exploration efforts over the last 50 years because of the ambiguity of the Viking results and concerns about repeating that outcome. However, a failure to embrace the uncertainty of the Viking results as an expected outcome of the search for life has unnecessarily caused this impasse. A better appreciation of this problem, combined with improved knowledge and instrumentation in microbiology (and other fields), enables us to continue the search for life in the universe. Furthermore, our growing knowledge of microbiology and microbial ecology should enable us to overhaul outdated concepts in planetary protection. Microbiology and microbial ecology can help us successfully establish human presence beyond Earth. An era in which microbiology and microbial ecology take a more prominent role across the whole landscape of space science and technology has arrived.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was funded by the
Associate Editor: Michael A. Meyer