
Abstract
The Perseverance rover recently discovered sedimentary rocks reddened by ferric oxides and peppered with bleached spots lacking these oxides. Some of these spots are associated with phosphate, iron sulfide minerals, and organic matter and are regarded as “potential biosignatures,” suggestive of microbial iron- and sulfate-reduction and organic matter oxidation. Similar millimeter–centimeter-scale “reduction spots” occur in many ancient red beds on Earth. Although terrestrial reduction spots are widely considered biogenic, the available evidence is not decisive, and the proposed microbial mechanism of spot formation has not been tested experimentally. Here, we report a successful laboratory demonstration of bleached spot formation in ferruginous sediment. Millimeter–centimeter-scale rounded bleached spots appeared within weeks on the underside of anaerobic sand–ferrihydrite slurries inoculated with microbial communities from the reducing zones of Winogradsky columns, originally seeded with soil and pondwater. The spatial and temporal distribution of observed bleaching events, which did not occur in sterile controls, is best explained by a microbially induced process, and DNA sequencing confirms that bacteria of iron-reducing genera (e.g., Paradesulfitobacterium) are abundant in the bleached areas. These results strongly support the longstanding hypothesis that microbial colonies can indeed generate visibly bleached reduction spots in ferruginous sediments and rocks. Further experiments are needed to establish whether and how nonbiological processes can mimic these features and to search for features that disambiguate biogenic and abiogenic reduction spots.
Introduction
In mid-2024, during its exploration of the Neretva Vallis channel leading into Jezero crater, Mars, NASA’s Perseverance rover encountered bleached (ferric-oxide-depleted) spots in the ferric-oxide-pigmented sedimentary rocks of the Bright Angel formation (Hurowitz et al., 2025). Similar “reduction spots” are common in ferric-oxide-rich red beds on Earth and have long been considered a potential biosignature (e.g., Hofmann, 1990; Spinks et al., 2010).
Perseverance found two populations of spots. Brick-red mudstones of the Mount Spoonhead member contained pale greenish millimeter-scale halos around mottled dark cores. Pink/tan mudstones of the Cheyava Falls member contained organic matter and pale millimeter-scale “leopard spots” depleted in ferric iron, enriched in ferrous iron, and enriched in sulfide relative to the matrix. These spots were lined with dark ferrous phosphate, also present in accompanying submillimeter-scale “poppy seeds.” Both populations of spots may be explained by localized reductive dissolution and loss of ferric iron; at Cheyava Falls, reduction of iron and sulfate may have been coupled to the oxidation of organic matter, liberating phosphate (Hurowitz et al., 2025). The greenish spots in the Malgosa Crest abrasion patch in the Mount Spoonhead member seem especially similar in appearance to Earth’s reduction spots (Broz et al., 2025). Curiosity has also observed features resembling reduction spots, with white halos and dark cores, in the slightly pink Carolyn Shoemaker formation of Gale Crater (Seeger and Grotzinger, 2024).
On Earth, reduction spots are thought to form when microorganisms couple the oxidation of organic matter to the reduction of iron during sediment diagenesis (pre- or post-lithification), creating a redox front that expands radially from a microbial colony (e.g., Hofmann, 1990). However, an abiotic origin can rarely be excluded, and evidence for biogenicity is typically indirect. For example, Spinks et al. (2010) reported sulfide δ34S values in a population of Triassic reduction spots markedly lighter than co-eval sulfates, suggestive of microbial sulfate reduction but within the range of (thermochemical) abiotic sulfate reduction. Similarly, in a global sample of Mesoproterozoic—Recent reduction spots, McMahon et al. (2018) found a pattern of progressive increase in δ238U from matrix to halo to core, consistent with the enzymatic fractionation of uranium isotopes by metal-reducing bacteria (Stylo et al., 2015) but not unequivocally so (Brown et al., 2018).
Because the redox reactions that occurred at Cheyava Falls could have been driven either by microorganisms or by abiotic chemistry, Hurowitz et al. (2025) describe them as “potential biosignatures.” A phenomenon is a “potential biosignature” if, first, “biological processes are a known possible explanation” and, second, “potential abiotic causes have not yet been reasonably explored and ruled out” (Gillen et al., 2023). Reduction spots meet the second criterion: Potential abiotic causes have not yet been reasonably explored, although experiments have shown that abiotic reductants can leach iron from sediment (Moulton, 1926; Hofmann, 1991). Arguably, however, reduction spots have not hitherto met the first criterion: It has never been demonstrated that biological processes can produce them.
This study, therefore, aimed to determine whether microbes can in fact produce reduction spots. We used Winogradsky columns to enrich natural mixed communities from pondwater and soil, expecting these communities to contain iron-reducing microorganisms, which are ubiquitous in near-surface environments on Earth (Weber et al., 2006). Samples from the Winogradsky columns were introduced to ferric iron-supplemented sediment slurries in an anaerobic chamber, mimicking the diagenetic conditions under which reduction spots are thought to form. We present evidence that these experiments successfully produced biogenic reduction spots.
Methods
Microbial inoculum
Sediment was collected in polypropylene cylinders from the Hermitage of Braid and Blackford Hill Local Nature Reserve on May 12, 2025. Soil was collected at 55°55′31.5”N 3°11′39.5”W at ∼5 cm depth; dark pond mud was collected from the bank of Blackford Pond at 55°55′32.9”N 3°11′39.6”W. Sediment was sieved through a 1-mm mesh.
Six Winogradsky columns were prepared in polypropylene cylinders (8.5 cm diameter and 18 cm height). Each contained (1) an enriched slurry of 1.67 g CaCO3, 1.67 g CaSO4, 6.67 g blended newspaper and tap water (1:3), and 33.3 mL of the environmental sample, following Lalla et al. (2021); and (2) 350 g of sample sediment and 100 mL of tap water. Four columns also contained 0.5 g ferrihydrite (synthesized according to Smith et al., 2012). The columns were capped with cling film and rubber bands and labeled according to sample source (S, soil; P, pond), presence (F) or absence (NF) of ferrihydrite, and growth conditions (L, light; D, dark), that is, Soil, no ferrihydrite, light (SNFL); soil, ferrihydrite, light (SFL); soil, ferrihydrite, dark (SFD); pondwater, no ferrihydrite, light (PNFL); pondwater, ferrihydrite, light (PFL); and pondwater, ferrihydrite, dark (PFD). “Light” columns were placed on a south-facing windowsill and exposed to a natural day/night cycle.
Deep black layers typical of microbial sulfate reduction (Rogan et al., 2005) developed at the base of all six columns by day 20 and were respectively 4.6 cm (PFD), 5.9 cm (PFL), 6.3 cm (SFD and SFL), 9.3 cm (PNFL), and 9.2 cm (SNFL) thick (Supplementary Fig. S1).
Sediment bleaching experiments
In Experiment 1, 57 days after the columns were established, 1 mL of inoculum was transferred anaerobically by sterile Pasteur pipette from the upper region of each black layer in each column into an autoclaved 1-mm ball of absorbent cotton wool (CW) (see Fig. 1 and below). One cotton wool ball received 1 mL of sterile pond water instead of a microbial inoculum. In Experiment 2 (day 114), the black layer in columns SFL and PNFL was sampled as before. In addition, a sterile control was prepared whereby the inoculum from the column was passed through a 0.22 µm nylon syringe filter before soaking into the cotton ball. In Experiment 3 (day 137), the black layers in columns SFL and PNFL were again sampled as before. In Experiment 4, the cotton ball was removed from the SFL dish in Experiment 1 (80 days after the original setup) and transferred to a new dish. Finally, an experiment was performed using a pure culture of the iron-reducing bacterium Geobacter bemidjiensis cultivated on Geobacter Medium 579.
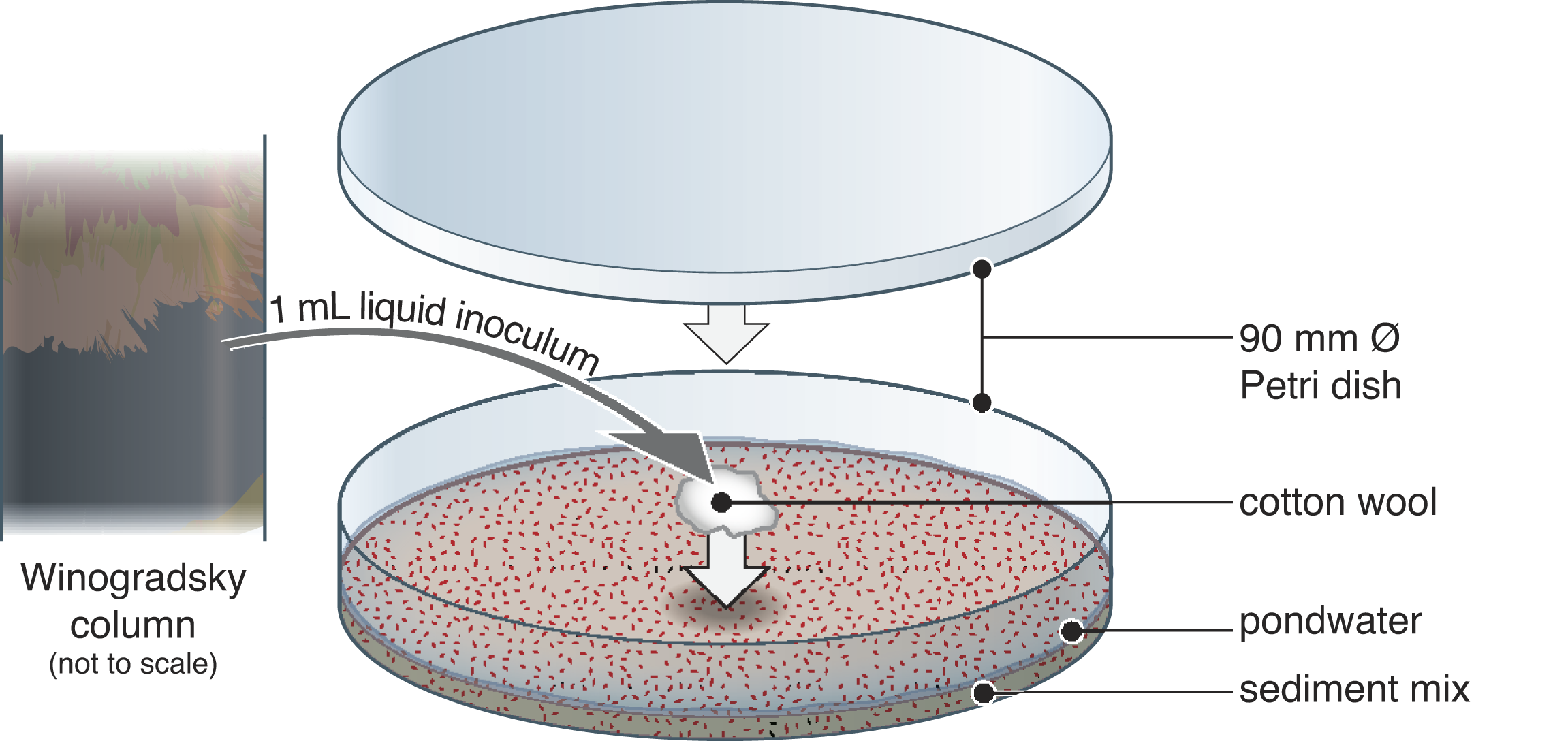
Schematic illustration of experimental set-up. In an anaerobic chamber, material from the reducing zone of Winogradsky columns (initially prepared with pondwater and soil) was introduced to balls of cotton wool, which were placed on a slurry of ferric-iron-rich sediment and sterile pondwater in Petri dishes in an anaerobic chamber under 20% CO2, 80% N2. The sediment mix consisted of 98 wt% quartz powder and 2 wt% synthetic ferrihydrite. Dishes were sealed with parafilm. See Supplementary Figure S1 for photographs of the Winogradsky columns.
Under a laminar flow hood, “quartz sand” powder (particle size <63 µm, Sigma-Aldrich; sterilized at 150°C for 16 h in a Teflon bottle) was mixed with 2 wt% ferrihydrite following Vixseboxse et al. (2024) and dispensed into 90-mm-diameter Petri dishes (22 g per dish; Experiments 1 and 2) or 40-mm dishes (3 g per dish; Experiment 3). Each dish was moistened with 10 mL (3mL in smaller dishes) of sterilized (filtered <0.22 µm) diluted pond water (collected from Blackford Pond, 11 June 2025) using a sterilized spray bottle, producing a < 1 mm supernatant above the saturated sediment. Dishes exchanged gas for 3 days in an anaerobic chamber (Coy; Vinyl Anaerobic Airlock Chamber, MI, USA; 20% CO2 and 80% N2 atmosphere).
In the anaerobic chamber, the inoculated cotton balls were placed gently on top of the sediment in the middle of each dish using sterile tweezers. All dishes were parafilm-sealed. Water lost by evaporation was replenished twice monthly to maintain the supernatant. Dishes were infrequently removed from the anaerobic chamber during the experiment for photography (∼3 min) and flatbed scanning (∼5 min, using a XEROX AltaLink C8035 scanner).
The methods used for DNA extraction and sequencing, resin impregnation, and electron microscopy (including energy-dispersive X-ray spectroscopy, EDX) are described in the Supplementary Data.
Development of bleached spots
In Experiment 1, bleaching occurred only in dishes inoculated from columns SFL and PNFL (Table 1). On day 22, bleached spots were noticed on the undersides of the sediment in both dishes, corresponding with the positions of the CW (Fig. 2). In the PNFL dish, the spot was circular, faint with indistinct margins, and ∼6 mm across. By day 36, it had expanded to ∼8 mm, become less faint, and acquired a slight orange halo; no further change occurred (Fig. 2). Patches of rust-orange material formed at the sediment–water interface in (only) the same two dishes, directly above the bleached zones (Supplementary Fig. S2).
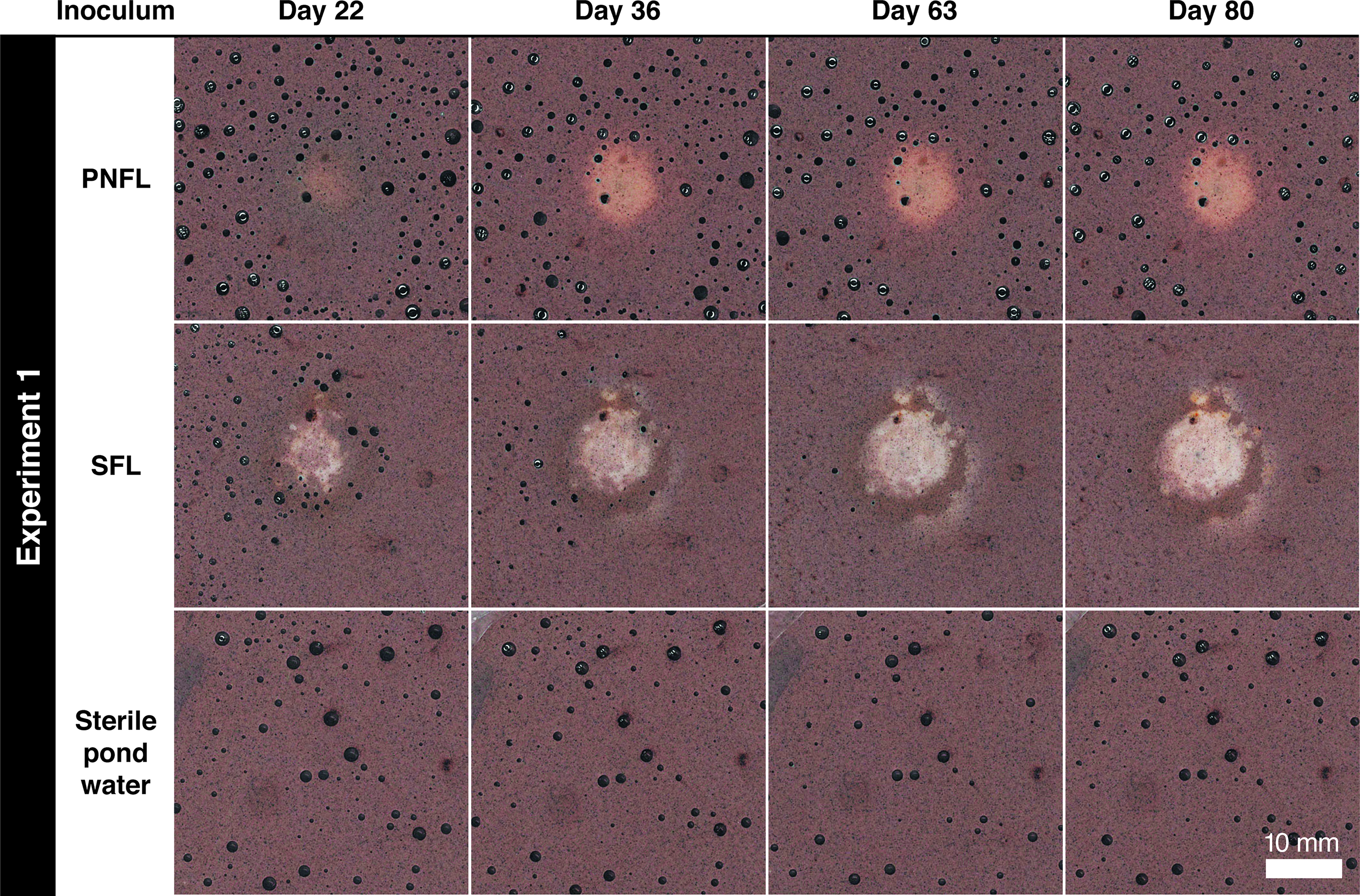
Time series of flat-bed scans showing central bleaching of the undersides of Petri dishes in Experiment 1. Numbers of days refer to time since dishes were inoculated. Fields of view are aligned across time points. Dark circles are air bubbles. Spots developed directly beneath inoculated cotton wool in the dishes inoculated from Winogradsky columns: pond sediment, no ferrihydrite, light (PNFL) and soil, ferrihydrite, light (SFL). The PNFL spot has less distinct margins than the SFL spot, which by day 36 had developed a secondary bleached halo at a distance of several mm from the original spot. Both PNFL and SFL spots showed some localized orange coloring to the margins.
Occurrence of Bleaching on the Undersides of Dishes

Numbers of days refer to time since Winogradsky columns were established. For each Winogradsky column, the original source of inoculum (soil or pondwater), presence or absence of ferrihydrite, and presence or absence of light are indicated. Note that bleaching occurred in all three dishes inoculated from the black layer in column SFL and did not occur when the inoculum from the same layer was filter-sterilized. However, the bleaching initially observed in the sample inoculated from column PNFL could not be replicated.
PNFL, pondwater, no ferrihydrite, light.
In the SFL dish, the central spot was ∼8 mm across when first noticed and irregularly shaped with orange peripheral patches. By day 36 it had expanded to a ∼10 mm circle with well-defined margins and a secondary bleached halo at a distance of several millimeters (Fig. 2). By day 36, additional rust-orange material formed at the sediment–water interface along the margins of the same dish (Supplementary Fig. S2); corresponding bleaching of the underside occurred in discrete ∼1-mm spots as well as larger blemishes (Fig. 3a). Optical imaging (Fig. 3b) and energy-dispersive X-ray spectroscopy (EDX) mapping (Fig. 3c) (after resin embedment) confirmed that the bleached zone was depleted in fine ferric particles compared with the matrix. No other elemental differences were apparent (e.g., in phosphorus or sulfur; Supplementary Fig. S3).
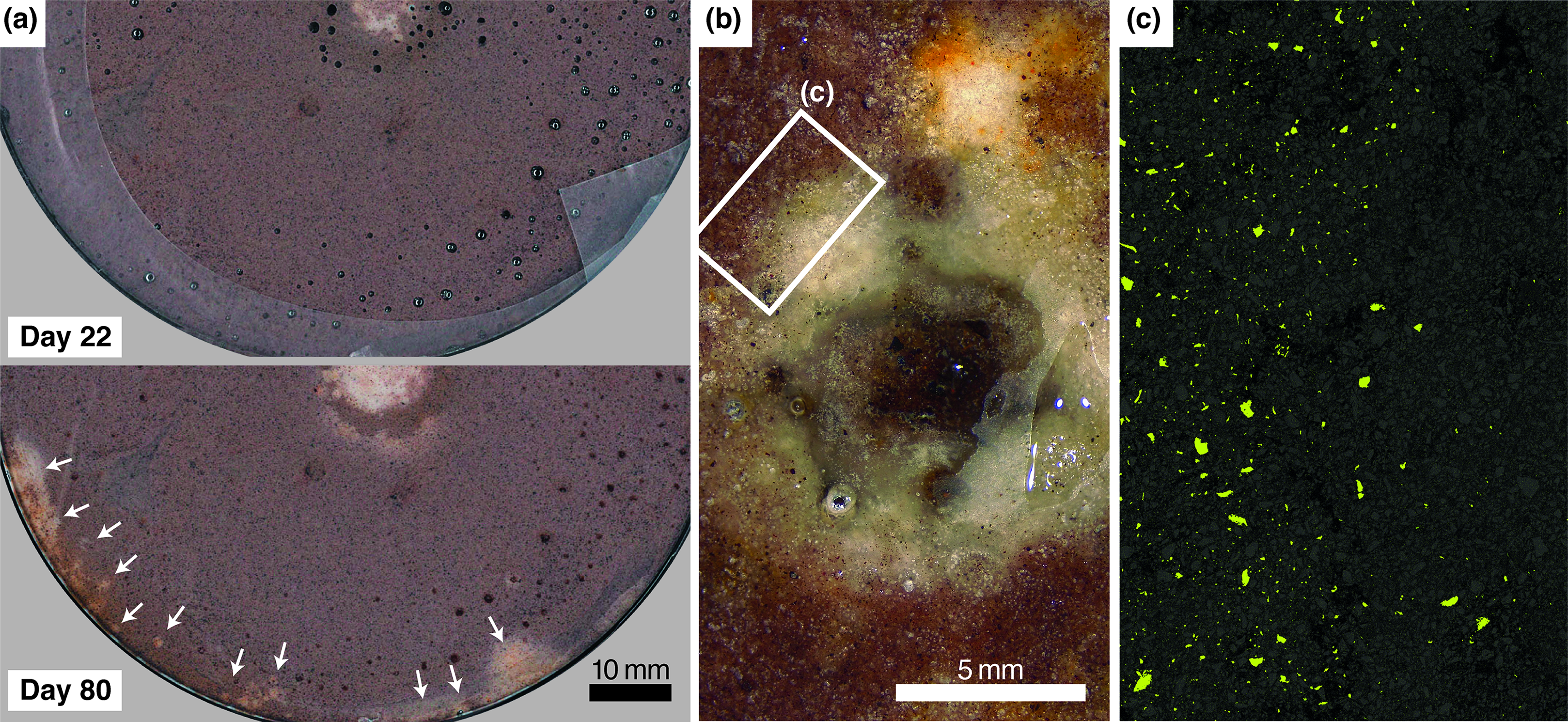
Additional observations from Experiment 1, dish SFL.
In Experiments 2 and 3, bleaching occurred only in dishes inoculated from column SFL (Fig. 4). Experiment 2 produced a faint, fuzzy spot with a dark orange-brown rim. An accompanying filter-sterilized control did not produce bleaching. Experiment 3 produced multiple spots with fuzzy margins and some orange coloring. No bleaching occurred in the dish inoculated with G. bemidjiensis. In Experiment 4, CW was transferred from the SFL dish in Experiment 1 (day 80) to a new dish. No spot was evident when this new dish was examined 90 days later, but after a further 60 days a very pale spot had appeared and grown to a size of about 20 mm, again with some associated orange stain. This spot continued to grow for months, becoming rounder and more sharply defined, while the surrounding sediment became increasingly orange (Fig. 4).
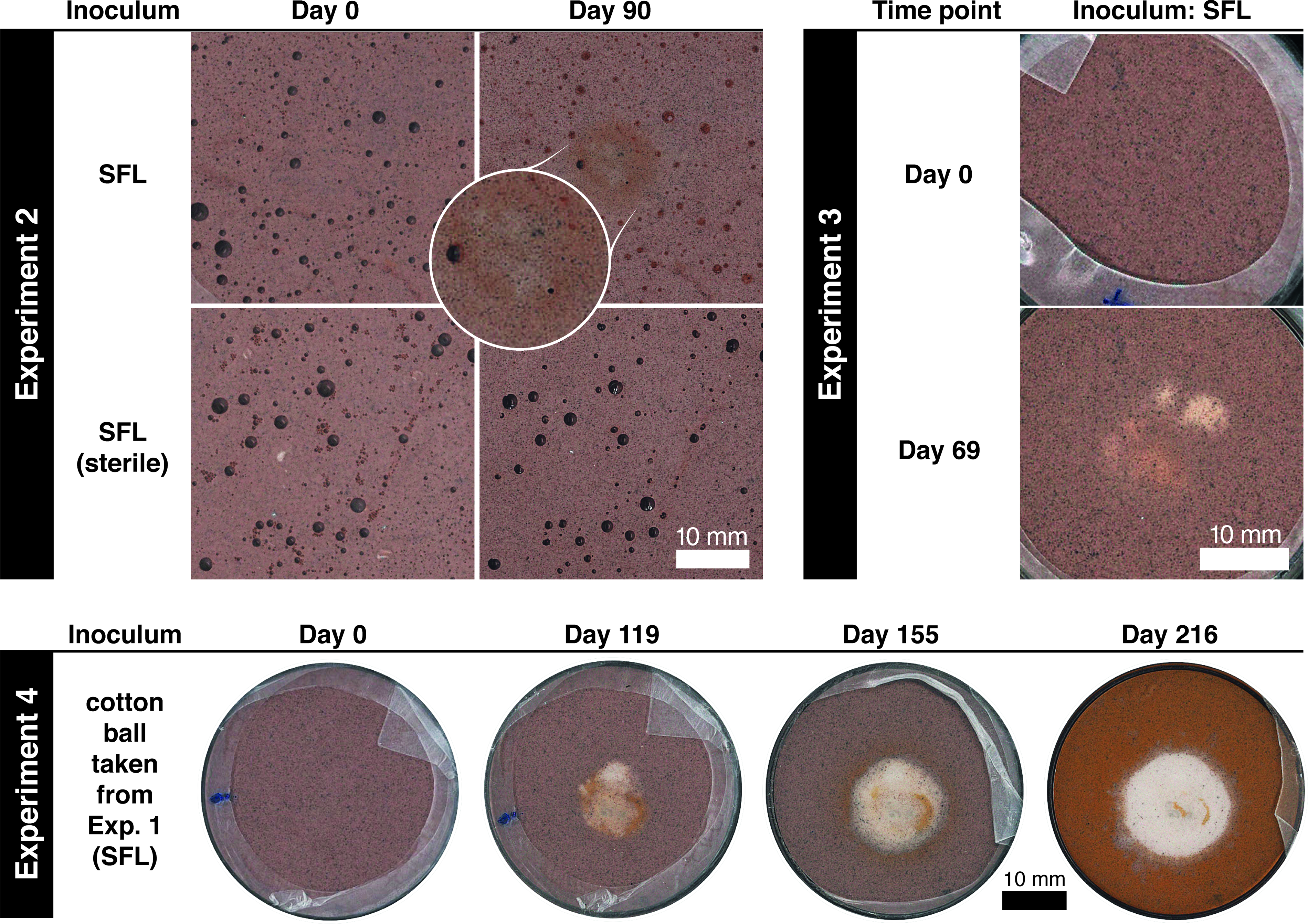
Flat-bed scans of Petri dishes showing bleaching in Experiments 2, 3, and 4. Fields of view are aligned across time points. Experiment 2 produced a faint, fuzzy spot with a dark orange-brown rim. No bleaching occurred in the filter-sterilized control. Experiment 3 produced multiple spots with fuzzy margins and some orange coloring (40 mm Petri dishes were used for this experiment). Experiment 4 contained the ball of cotton wool transferred from Experiment 1 dish SFL. The matrix gained an orange stain as the reduction spot grew.
Slurry from Experiment 1 dish SFL was sampled from the sediment above the peripheral bleached zone (PBZ) and from around the central “CW” for 16S rRNA gene sequencing (see Supplementary Data). Briefly, 488 amplicon sequence variants were identified across the two samples, indicating 136 distinct bacterial genera (Fig. 5). Of those genera accounting for at least 1% of reads, genera typically characterized as Fe(III) reducers (Paredesulfitobacterium and Desulfosporospinus, both strictly anaerobic) jointly comprised 52.7% in “PBZ” and 16.8% in “CW” (e.g., Li et al., 2021). Other genera containing at least some Fe(III)-reducing species comprised 30.2% in “PBZ” and 36.6% in “CW.” Of these, the most abundant was Cellulomonas (a facultatively anaerobic cellulose degrader (Stackebrandt and Schumann, 2014), some of whose strains are known to perform dissimilatory Fe(III) reduction (Sani et al., 2002)). Genera not known to contain any Fe(III)-reducing species comprised 10.3% of reads in “PBZ” and 36.1% in “CW.” Genera accounting for <1% of reads comprised 6.8% in “PBZ” and 10.5% in “CW.”
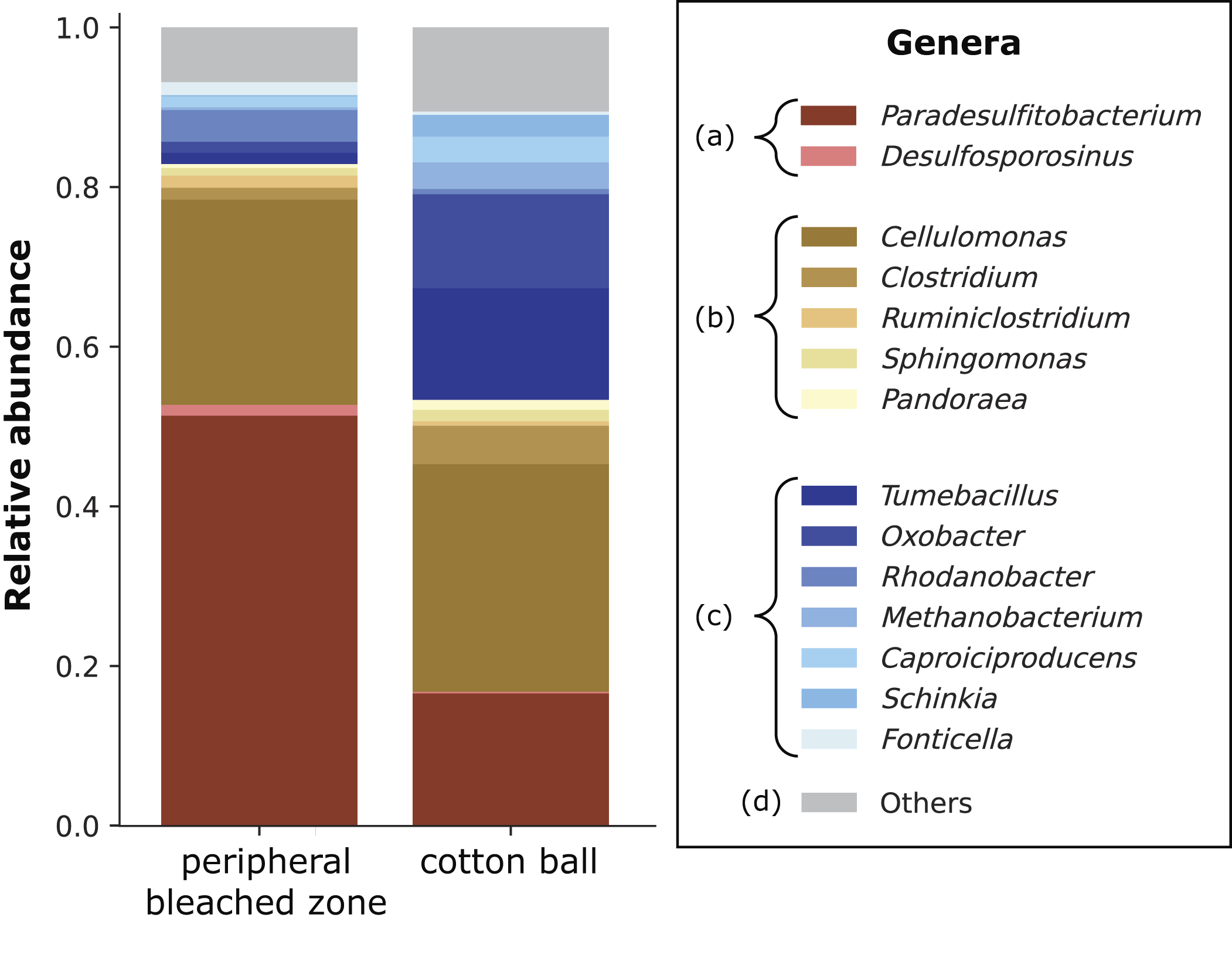
Genera occurring above reduction spots and association with Fe(III) reduction in Experiment 1, dish SFL. Wet sediment samples were taken from above the peripheral bleached zone (shown in Fig. 3) and from around the cotton wool 64 days after the dish was inoculated from a Winogradsky column prepared with soil, ferrihydrite, and natural light. Genera are grouped as follows: (a) all reported taxa in these genera can reduce iron; (b) some reported taxa in these genera can reduce iron; (c) no reported taxa in these genera can reduce iron; (d) low-ranked/others. Genera of <1% abundance across both samples have been grouped in the category “Others.”
Our observations confirm that bleaching occurred via the reductive dissolution and diffusive loss of ferrihydrite. Orange-brown blemishes near the bleached zones and orange-brown crusts appearing on dish upper surfaces (Supplementary Fig. S2) probably indicate re-oxidation of iron derived from the ferrihydrite. Vixseboxse et al. (2024) produced comparable orange halos around decaying organic matter in similar experiments. No other mineralization was observed.
We are not aware of any previous report of the production of sedimentary reduction spots in the laboratory. Moulton (1926) found that crude oil was unable to bleach powdered ferruginous sediment. Hofmann (1991) successfully leached hematite from sedimentary rock with dithionite, a powerful artificial reducing agent. More generally, it is well known that sulfides and organic acids can abiotically reduce and dissolve Fe(III), especially in poorly or noncrystalline phases, in days (Moulton, 1926; Jauregui and Reisenauer, 1982; Miller et al., 1986; Torres et al., 1990; Ionescu et al., 2015), (including ferrihydrite; Miller et al., 1986). Organic acids adsorbed onto Fe-oxide surfaces form inner-sphere metal chelates; reduction via ligand-to-metal electron transfer results in the dissolution of the iron oxide and removal of ligand-bound Fe(II) complexes from the Fe-oxide surface, driving continued dissolution.
Other than the activity of microbes, our initial experiments contained at least two sources of sulfides and organic acids, namely, the 1 mL of material transferred from the Winogradsky columns and the pondwater in which all experiments were performed. Nevertheless, we infer that bleaching observed in our experiments was microbially mediated, for six reasons:
Of the four dishes inoculated from the same layer in column SFL, spots appeared in three and failed to appear only in the one dish whose inoculum was filter-sterilized, a process that would have removed cells but not organic acids or other reductants. The sequencing data revealed abundant bacteria of iron-reducing genera in the sediment above the bleached areas in the SFL dish from Experiment 1 (64 days after inoculation); such genera were especially dominant in the peripheral area. All samples used to inoculate all six Petri dishes were reducing, sulfidic, and rich in organic matter, being taken from the black zone of Winogradsky columns. Nevertheless, inocula from only two columns caused bleaching, which must, therefore, require a more specific (and temperamental) agent than organic matter alone, for example, certain microbial strains. Bleaching apparently accelerated in Experiment 1, suggesting a lag period and/or gradual propagation of bleaching downward through the sediment, both typical of microbial growth. Abiotic solubilization of iron would have occurred rapidly after the introduction of material to the dishes and slowly thereafter (e.g., Miller et al., 1986). In contrast, dish PNFL bleached faster between days 22 and 36 than between days 0 and 22 (Fig. 1). Visible bleaching of the margin of dish SFL did not begin until after day 22. Similar delays occurred in the other experiments. The late marginal bleaching observed in Experiment 1 dish SFL occurred at multiple discrete millimeter-scale points as if induced by immobile localized agents such as microbial colonies (Fig. 3a). Abiotic organic acids would not have been spatially focused in this way. When the cotton ball was removed from Experiment 1 dish SFL and transferred to a new dish, a new bleached spot formed after several months, with no diminution of effect (Fig. 4). This suggests a successful “subculturing” of the original spot-forming population.
Although limited in size and scope, this study represents a first step towards a more exhaustive exploration of the mechanisms that might explain the occurrence of reduction spots in red rocks on Earth and Mars. Our general experimental approach is facile and easily replicated and extended. To enhance the realism of our experiments, the method could be improved by replacing granular ferrihydrite with a more cement-like phase precipitated directly onto the sand grains. It would also be worthwhile to incorporate pressure–temperature conditions, fluid compositions, and iron (oxyhydr)oxide phases characteristic of (later) diagenesis. Finally, future work should seek diagnostically useful differences between bacteriogenic and abiotic reduction spots and their associated morphological, geochemical, and mineralogical fingerprints.
Our results show that iron-reducing microorganisms can produce discrete, rounded, millimeter–centimeter-scale bleached spots in ferruginous sediments, suggesting that those found commonly in red beds on Earth—and those recently observed by Perseverance on Mars—are indeed “potential biosignatures.” The demonstration that microbes can produce reduction spots under some conditions does not imply that all reduction spots are biogenic. It is now essential to investigate potential nonbiological pathways to reduction spot formation on Earth and Mars. Nevertheless, our results lend weight to the suggestion that reduction spots in Neretva Vallis may be the first visible sign of life discovered on another world.
Footnotes
Acknowledgments
The authors thank Kevin Dodd for resin-embedding samples; Nicola Cayzer for assistance with SEM-EDX; Charles Cockell, Niall Rodgers, and Seán Jordan for advice, support, and discussion; and the three anonymous reviewers of this article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Part of this work was funded by the
Supplemental Material
Associate Editor: Radu Popa
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.