
Abstract
James Lovelock was the first to lay out a physical and chemical basis for searching for life beyond Earth, based on the observation that the molecular features and patterns in classes of organics differ for compounds formed through biological versus nonbiological processes. This approach shaped the Viking molecular analysis experiment designed to search for organics, including potential signs of extinct or extant life, and it still remains fundamentally viable today. In the five decades since Viking, many missions have uncovered new information about the martian organic inventory, geological landscape, and changes to habitability over time, while advances in organic geochemistry on Earth enabled scientists to identify even more differences between biotic and abiotic organics, solidifying the utility of Lovelock’s physical and chemical approach in the search for extraterrestrial life. Key Words: Search for Mars’ Organics—Mars—Biosignatures—Life detection—Biomarkers. Astrobiology 00, 000–000
A Physical Basis for Life Detection Experiments
In 1965, James Lovelock published an article in Nature entitled “A Physical Basis for Life Detection Experiments.” He proposed a set of efficient and unequivocal experiments to search for life, based on the hypothesis that a nonsterile planet will be recognizable through physical phenomenology (Lovelock, 1965). Lacking a formal definition of what life is, we can at minimum recognize what life does to a planet, which is to (1) create ordered chemical and physical structures and (2) increase chemical disequilibrium. These features produced by biological activity are recognizable and unique because they defy thermodynamic and kinetic principles that dictate products of abiotic chemistry. The case of Earth demonstrates that life is a global phenomenon, one that began quickly and overtook the planet through an extended and growingly complex set of biochemical reactions, producing ordered and discrete sets of nonrandom organic molecules that form cellular structures, power metabolism, and encode information. As biology modifies the geologic landscape through these processes, it leaves behind physical and chemical evidence imprinted on the environment, including body fossils and molecular biosignatures preserved in the rock record that are distinctive from abiotically synthesized organic molecules, along with atmospheric disequilibrium (e.g., Summons et al., 2011; Lovelock, 1965).
Lovelock’s fundamental observation and proposed life detection measurements served as the foundation for molecular biosignature detection strategies that have guided search-for-life missions within our Solar System ever since. These experiments were designed to (1) search for physical order, including the structures and distributions of organic molecules that differ for biogenic versus abiogenic synthesis (Fig. 1A), (2) search for chemical disequilibrium, including surface-atmosphere exchange or the presence of atmospheric species that cannot coexist long-term in the absence of biogenic production (Lovelock, 1965). This approach is broadly applicable (Fig. 1B), as it does not require detection of the same set of specific compounds, structures, or metabolic products that may be unique to terrestrial organisms, but instead searches for deviations from the abiotic norm, which should be a universal feature of carbon-based life as we understand or expect it to be (Dorn et al., 2011; Georgiou and Deamer, 2014; Glavin et al., 2018; Buckner et al., 2024). While planetary-scale signatures of chemical disequilibrium can indicate the presence of extant life, physical signatures of directed biosynthesis are encoded in molecular structures themselves, and recalcitrant organics preserved in rock can serve as signs of either extant or extinct biota, making this approach particularly suited to the search for life on Mars (Vago et al., 2015; Buckner et al., 2024; Saeedi and Buckner et al., 2025; Buckner and Mojarro, 2026).
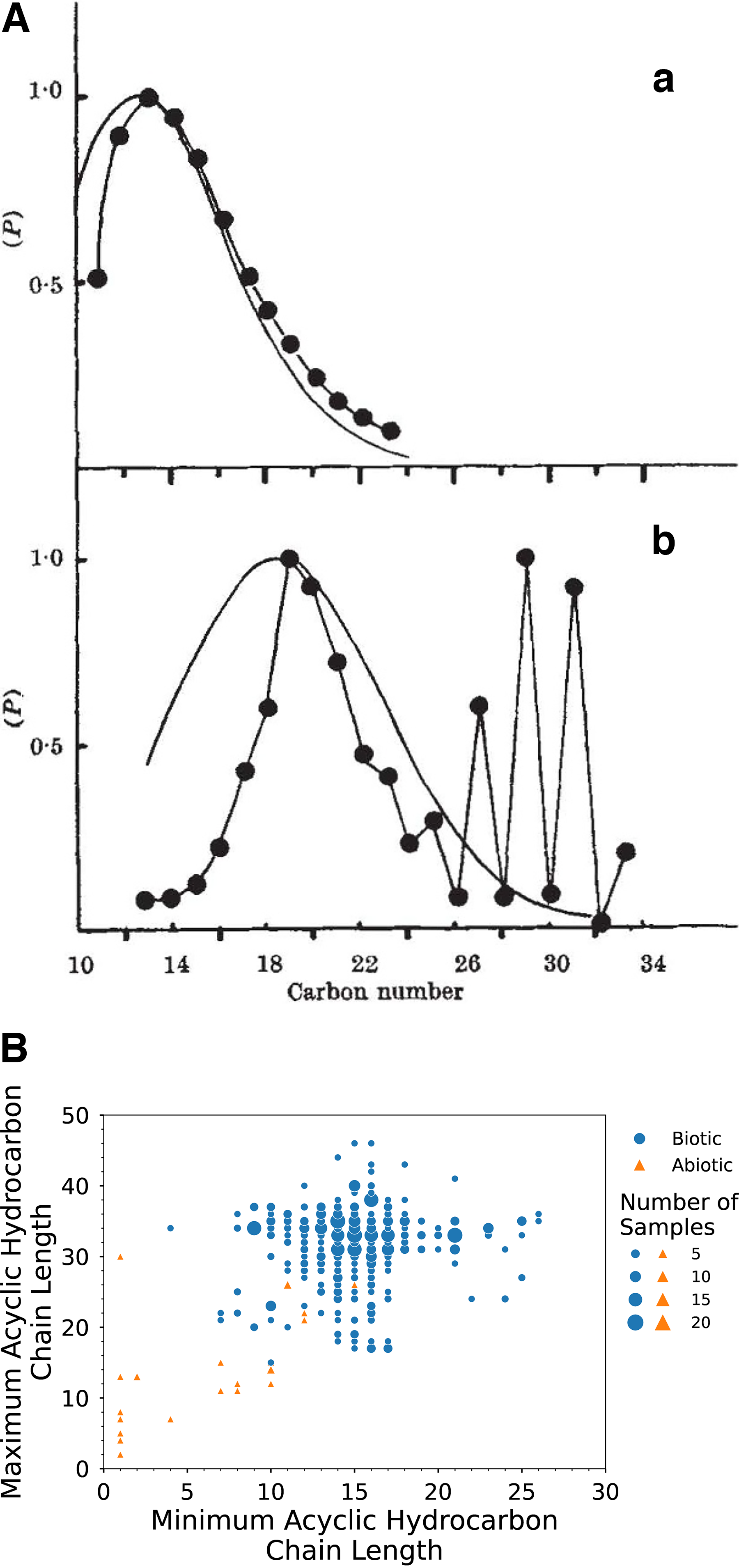
Penned before humankind ever reached Mars and less than a decade after the field of organic geochemistry was formally established, Lovelock’s framework for the search for physical and chemical order as signs of life was pioneering (e.g., Kvenvolden, 2006; Dorn et al., 2011; Neveu et al., 2018; Buckner et al., 2024; Hoehler et al., 2025; Malaska et al., 2025; Saeedi and Buckner et al., 2025). Over five decades later, much of this approach to searching for extraterrestrial biochemistry remains foundational for life detection mission strategy and experimental design (Des Marais et al., 2008). This work established the idea that molecular patterns could differentiate products of biotic from abiotic chemistry, and in the following decades, continued analysis of terrestrial and extraterrestrial samples have uncovered numerous additional hallmark molecular distributions and features, reaffirming the utility of this approach (Fig. 1B) (Cronin and Chang, 1993; Pizzarello et al., 2001; Eigenbrode, 2008; Summons et al., 2008; Georgiou and Deamer, 2014; Mißbach et al., 2018; Buckner et al., 2024; Malaska et al., 2025; Saeedi and Buckner et al., 2025). Abiotic organics decrease in abundance with increasing molecular weight and display random structures with no isomeric preference, while biology displays a preference for specific conformations, chain lengths, and subsets of isomers. Some of these key molecular features and patterns that can indicate how an organic molecule was synthesized—through biologic or nonbiologic processes—include subsets of monomers (e.g., 20 amino acids), preference for specific and non-random molecular conformations/isomerizations (e.g., chain length, position, and number of unsaturations and branch points, presence of cyclic moieties, and functional groups for hydrocarbons and amphiphiles), and enantiomeric excess (i.e., preference for either the left [L-] or right [D-] handed conformation for chiral compounds) (Summons et al., 2008; Glavin et al., 2018; Buckner et al., 2024).
The ability to search for ordered molecular structures was made possible with the advent and commercial availability of the gas chromatograph–mass spectrometer (GC-MS); the combined GC and MS was first demonstrated in 1955 by Fred McLafferty and Roland Gohlke from Dow Chemical (Gohlke, 1959). This technology became a cornerstone of the field of organic geochemistry and was widely applied for analysis of both synthetic and natural samples. Lovelock and others noted that while the same classes of compounds are found in both biogenic materials from Earth and abiogenic materials from meteorites and laboratory synthesis experiments, a clear distinction in the structures and distributions of individual molecules sets the two apart (Meinschein, 1963; Hayatsu, 1964; Studier et al., 1965). Abiotic synthesis reactions observed in the interstellar medium, within primitive accreting bodies, and through atmospheric and geochemical reactions in planetary environments produce functionalized organics with random structures and stochastic distributions, including short-chained and highly branched carboxylic acids, more than 100 amino acids with both D- and L-conformations, and a variety of nucleobases, N-heterocycles, and sugar derivatives (Glavin et al., 2018). On the other hand, life on Earth preferentially synthesizes discrete sets of monomers with specific structures to build complex biomolecules and cellular structures, including predominantly straight-chained C16 and C18 fatty (i.e., carboxylic) acids that form lipids, 20 homochiral L-amino acids that make up proteins, and five nucleobases with D-ribose and D-2-deoxyribose that assemble into RNA and DNA (Peters et al., 2005; Summons et al., 2008). Even if extraterrestrial organisms utilize different molecular suites or conformations and biopolymers unique from life as we know it, nonrandom organic structures and preference for specific conformations within organic compound classes that are also synthesized in abiotic settings could signal life (Dorn et al., 2011; Buckner et al., 2024; Saeedi and Buckner et al., 2025; Malaska et al., 2025).
The Viking Molecular Analysis Experiment
As a complement to the Viking biology experiments discussed elsewhere in this special collection each lander carried a GC-MS capable of analyzing atmospheric gases and detecting organics at parts per million to parts per billion concentrations and elucidating their molecular structures. This process involves pyrolyzing martian samples to thermally liberate organics from mineral matrices. Volatilized compounds are passed through the GC column with a flow of gas and undergo chromatographic separation; then they exit the column and enter the MS, where they are ionized and accelerated toward the detector, fragment, and are identified based on their mass-to-charge (m/z) ratios (Anderson et al., 1972; Grayson, 2016). Lovelock contributed significantly to enabling the GC-MS to fly on Viking by developing a gas transmodulator that significantly reduced the mass of the instrument for flight, along with a separator for removing the carrier gas from the system so molecules can efficiently pass into the MS (Lovelock et al., 1969; Simmonds et al., 1970; Anderson et al., 1972; Biemann et al., 1976; Grayson, 2016).
According to the 1972 overview article that describes the Viking molecular analysis experiment and GC-MS, The purpose of the organic analysis experiment of the first Viking mission is, therefore, an understanding of the quantity and complexity of organic compounds at the surface of Mars. One can then attempt to deduce their origin and the extent to which they may be capable of evolving into or from living systems. In this respect, it provides valuable support of the active biology experiment of this mission. Furthermore, the molecular analysis experiment should yield knowledge of the level and type of Martian organic chemistry necessary to design a second-generation biology experiment based on a Martian rather than terrestrial biological model (Anderson et al., 1972).
The GC-MS onboard Viking 1 analyzed two surface samples from Chryse Planitia. Instead of finding an array of indigenous organics as expected, the science team reported that the only compounds detected in the first sample were methyl chloride and freons (Biemann et al., 1976). Two surface samples from Utopia Planitia analyzed by Viking 2 produced similarly surprising results, with detections that included methylene chloride (but no methyl chloride), freons, siloxanes, toluene, xylene, acetone, benzene, and alkylbenzene (Biemann et al., 1977). In both of these reports, all organics detected were assigned as terrestrial contaminants (Biemann et al., 1976, 1977). Most compounds were observed in preflight and flight tests, with the exception of methyl chloride and methylene chloride. The science team concluded that: The methyl chloride, or part of it, could conceivably be indigenous to Mars. However, if it were, one would expect that other related compounds like ethyl chloride or methyl bromide would also be formed, but none were detected. The abundance ratio of m/z 50 to m/z 52 was about 3:1, corresponding to the terrestrial isotope ratio of chlorine, but this does not necessarily confirm its origin, because there is no reason to predict a different ratio for Martian chlorine. Considering all these facts, we tend to believe that all the methyl chloride is from terrestrial sources (chlorinated solvents or from adsorbed traces of methanol and HC1) (Biemann et al., 1977).
With the results of the molecular analysis and biology experiments, Mars was deemed uninhabited, interest in astrobiology waned, and surface exploration entered a hiatus for two decades. However, these first landed missions to Mars demonstrated successful robotic exploration of another planet, including operation of the GC-MS, and critical environmental and geochemical information was obtained by the payload’s instrument suite and cameras. Prior to launch, the molecular analysis team acknowledged that
“Unfortunately, our knowledge of the organic chemistry on the surface of Mars is currently nil and it is indeed the chief purpose of this experiment to obtain useful information in any of a wide range of possible situations” (Anderson et al., 1972; Soffen, 1977). Initial findings were unexpectedly disappointing to the scientific community, but ultimately, this experiment was a success, and the head of the NASA Jet Propulsion Laboratory (JPL) bioscience division, Norman Horowitz, stated the Viking GC-MS was “probably the most important single instrument on the Mars lander” (Watts, 2024). Later missions built a greater understanding of the martian geochemical landscape, and analysis of organic biosignatures in hyperarid Mars analog settings on Earth enabled reanalysis of the molecular analysis experiment data (Navarro-González et al, 2003; Hecht et al., 2009; Quinn et al., 2013; Guzman et al., 2018). Interpretations of what the Viking GC-MS discovered have since changed.
Mars Exploration Post-Viking
After Viking, questions remained about the apparent non-detection of native organics by the GC-MS, because even if Mars was sterile, organics should still be pervasive due to in situ production and exogenous infall from meteorites, comets, and interplanetary dust particles (IDPs), which together deliver an estimated 105 kg of organic carbon to the surface of Mars every year (Chyba and Sagan, 1992; Flynn, 1996; Ten Kate, 2010; Frantseva et al., 2018). Hypothesized reasons for the missing organics included photolysis from short-wavelength UV radiation, degradation through interactions with H2O2, other soil oxides, and superoxides produced through atmospheric photochemical pathways, and oxidation to metastable nonvolatile carboxylic acid salts that would not have been detectable by a GC-MS (Oyama and Berdahl, 1977; Zent and McKay, 1993; Stoker and Bullock, 1997; Benner et al., 2000). Meanwhile, the field of organic geochemistry continued to advance, and in 1996 NASA announced the discovery of potential mineralogical, morphological, and organic biosignatures in ALH 84001, a 4.5 billion-year-old martian meteorite (McKay et al., 1996). Subsequent reanalysis of ALH 84001 found abiotic explanations for the features McKay et al. (1996) attributed to biology, but the detection of indigenous organics renewed interest in martian history and astrobiological potential (Jull et al., 1998; Golden et al., 2004).
These results coincided with NASA’s return to the red planet under the Mars Exploration Program (MEP), conceived in 1993 to expand on geochemical and climatic knowledge built by Viking, but with lower-cost and faster-turnaround orbital and landed missions. The MEP’s goals are to (1) determine if life ever arose on Mars, (2) characterize the climate of Mars, (3) characterize the geology of Mars, and (4) prepare for human exploration (Shirley and McCleese, 1996). The Mars Pathfinder mission launched in December 1996, and sustained surface and orbital exploration under MEP have continued to the present. Early missions did not conduct any experiments relevant to the search for life, but in 2008, the Phoenix lander discovered perchlorates heterogeneously distributed in martian topsoil samples, providing an answer to Viking’s apparent nondetection of indigenous organics (Hecht et al., 2009). Since Viking’s GC-MS utilized pyrolysis to thermally extract organics from mineral matrixes, subsequent laboratory experiments demonstrated that heating organic molecules in the presence of oxychlorines leads to their rapid oxidation and modification (e.g., Navarro‐González, 2010). Re-analysis of Viking mission data indicated that the methyl chloride and methylene chloride signal initially attributed to chlorinated solvents or adsorbed methanol and HC1 were likely breakdown products of these reactions, revealing that Viking did detect native organics after all (Quinn et al., 2013; Guzman et al., 2018).
Following this discovery, the Curiosity rover returned a GC-MS to Mars in 2012 and successfully found organics at numerous locales within ancient lake sediments of Gale crater, including chlorobenzene and other chlorinated species, N- and S-bearing heterocycles, benzoic acid, and aliphatic and aromatic hydrocarbons. The forms and distributions of these molecules are consistent with degraded meteoritic infall, while long-chain n-alkanes have been interpreted as possible diagenetic products of fatty acids (Eigenbrode et al., 2018; Freissinet et al., 2025; Pavlov et al., 2025). A targeted search through Viking GC-MS data for the compounds detected by the Sample Analysis at Mars (SAM) instrument found a signal consistent with chlorobenzene at sub-part-per-billion levels, further confirming that native martian organics were detected by the molecular analysis experiment (Guzman et al., 2018). In 2021, the Perseverance rover landed on Mars, carrying the Scanning Habitable Environments with Raman and Luminescence for Organics and Chemicals (SHERLOC) instrument, which is a deep ultraviolet (DUV) Raman and fluorescence spectrometer to continue the search for organics. Unlike GC-MS, this optical technique measures molecular vibrational modes, so while organic structures are not resolvable to the same detail, its benefit lies in an ability to identify both organic functional groups and minerals and to spatially map their associations, providing geochemical contextual information (Bhartia et al., 2021). Perseverance detected macromolecular material and polycyclic aromatic hydrocarbons at multiple locations across Jezero crater, and organo-mineral associations are consistent with carbon of varying provenance, including abiotic in situ synthesis through igneous processes and reduction reactions potentially consistent with biological activity (Sharma et al., 2023; Fornaro et al., 2025; Hurowitz et al., 2025). Organics interpreted to have formed through various abiotic geochemical pathways have also been identified in martian Shergottites, Nakhlites, Chassignites (SNC) meteorites, providing further evidence for dynamic carbon cycling on Mars (Steele et al., 2012; Lin et al., 2014; Steele et al., 2018; Jaramillo et al., 2019). Together, these detections highlight the potential for preservation over several billion years on Mars, even after 105 to 106 years of exposure to ionizing radiation (e.g., Pavlov et al., 2012; Fox et al., 2019). If life did not arise on Mars, organics are still expected to remain at the near-surface, sourced from extraplanetary material delivered by meteorites, comets, and IDPs (Flynn, 1996; Mahaffy et al., 2012; Frantseva et al., 2018) and/or endogenous compounds generated in situ through shock synthesis, igneous processes, water-rock reactions, photochemistry, or electrochemical reduction of CO2 (Chyba and Sagan, 1992; Steele et al., 2018; Fornaro et al., 2025).
The Utility of Organics in Life Detection
In the present era, identifying where and how organics are synthesized throughout the Solar System and searching for biosignatures on Mars are two of NASA’s key near-term goals laid out by the 2023–2032 Planetary Science Decadal Survey (National Academies of Sciences, Engineering, and Medicine, 2018). Types of potential biosignatures include macro- to micro-morphological structures, microtextures and fabrics, mineralogical indicators, chemical distributions, organics, and isotopic signatures. Of this set, molecular biosignatures are particularly powerful and information-rich, as they (i) are fundamental to life as we know it (i.e., carbon-based) or can imagine it, (ii) possess one or more structural features and distributions that are distinct from abiotic counterparts (e.g., subsets of monomers, preference for non-random conformations, specific carbon chain lengths and branching patterns), (iii) display forms and conformations that reflect evolution and cellular function (e.g., hydrocarbons and amphiphiles that can form cell membranes and walls, components for information-encoding biopolymers similar to RNA and DNA), (iv) exhibit preservation potential over geologically relevant timescales (i.e., hydrocarbon skeletons billions of years old), and (v) are readily detectable by instrumentation available on rovers/landers or in the laboratory (Buckner et al., 2024 and references therein).
Organics are universal to all life on Earth and should be fundamental for similar life-forms elsewhere. These compounds are synthesized through both biotic and abiotic processes and are ubiquitous throughout the Solar System, detected on Mars, Titan, Enceladus, meteorites and asteroids, IDPs, and comets, and observed throughout the interstellar medium (National Academies of Sciences, Engineering, and Medicine, 2018). All life is made of organics, but not all organics are formed by life, and functionalized organics with increasing complexity are synthesized through stellar death and birth, accretionary processes in the protoplanetary disk, and geochemical activity on and within planetesimals and planets. These processes are recorded in organics-rich primitive exogenous materials that have been delivered to the inner planets over the Solar System’s history. Carbonaceous chondrites and samples returned from asteroids Ryugu and Bennu contain a vast array of soluble organics with tens of thousands of molecular formulae, including a nearly complete inventory of prebiotic building blocks used by life on Earth (Schmitt-Kopplin et al., 2010, 2023; Glavin and Dworkin et al., 2025). Targeted analyses of these samples have identified 15 of the 20 proteinogenic amino acids, all five canonical nucleobases in RNA/DNA, additional components required for RNA synthesis (e.g., ribose, phosphate), bio-essential sugars and sugar derivatives, fatty (i.e., carboxylic) acids and other amphiphiles C8 and longer that are capable of forming protomembranes, and other CHONPS-bearing functionalized molecules required for metabolic processes (Deamer, 1986; Dworkin et al., 2001; Glavin and Dworkin et al., 2025; Mojarro et al., 2025; Furukawa et al, 2026). These discoveries highlight a potential link between exogenous infall and life on Earth, and because Mars received the same infall, these building blocks could have seeded life there as well; alternatively, geochemical and photochemical reactions occurring in situ could have produced similar monomers (Buckner and Mojarro, 2026). Because the structural form of organics synthesized by biology reflects their cellular function, organics could serve as indicators for life that is similar to, yet slightly distinct from, terrestrial organisms. Potential examples include a different set of amino acids, nucleobases, and sugars; hydrocarbons with non-random structures unique from those produced by life on Earth; or a preference for other functionalized molecules. In this case, a thermodynamic or kinetic deviation from the distributions produced by abiotic processes can potentially signal directed synthesis.
The search for order in physical and chemical structures remains a practical technique in astrobiological exploration on Mars, because while atmospheric disequilibrium is a sign of active metabolism, organics can persist in the rock record over long geologic timescales and can therefore record evidence of both extant and extinct life. In particular, hydrocarbons and lipids are extraordinarily well-preserved in the rock record on Earth, and they record the activity and evolution of organisms that lived billions of years ago (Brocks et al., 2003; Summons et al., 2011). The geologic longevity of certain of organic classes is comparable in magnitude to the age of the sediments laid down during the most habitable surface conditions on Mars, and preservation is further enhanced under cold and desiccating conditions that have dominated Mars for the majority of its geologic history, especially in the absence of thermal metamorphism driven by tectonism that is largely responsible for organic degradation on Earth (e.g., Georgiou and Deamer, 2014; Wilhelm et al., 2017, 2018; Buckner et al., 2024). Due to their geologic recalcitrance and diagnostic molecular structures, these compounds are ideal molecular targets in the search for ancient life on Mars (e.g., Vago et al., 2015).
Conclusions
Lovelock expected organics on Mars regardless of their origin, and after 50 years of Mars exploration, his predictions have been confirmed with detections of organics within martian meteorites, reanalysis of GC-MS data from the Viking molecular analysis experiment, and in situ organics measurements from surface and near-surface samples across the planet with subsequent landed missions. Additionally, when Viking embarked on the search for extant life, scientists had not yet discovered just how long-lived organics could be, and the last 50 years of organic geochemistry research has revealed that rich biological histories are recorded by molecular biomarkers preserved in rocks from ancient terrestrial environments (Brocks et al., 2003; Peters et al., 2005; Brocks and Schaeffer, 2008; Lee and Brocks, 2011). These same principles are particularly applicable to Mars, especially as one of the key findings of Mars exploration is that surface habitability declined over time as the planet cooled and dried, but those geological parameters also enhance organic preservation potential. It turns out that organic biosignatures are even more useful than Lovelock or the Viking team could have anticipated. In the coming decades, interrogating the structures, distributions, and diversity of martian organics to determine their origin and diagenetic history is imperative for the search for life and is long overdue. Analyzing organics may provide the best chance to determine whether life ever arose on Mars.
Footnotes
Acknowledgments
The authors would like to thank Penny Boston for helpful discussions and useful input from two anonymous reviewers.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the NASA Postdoctoral Program (NPP) and Oak Ridge Associated Universities (ORAU), funded by Award NNH-09ZDA007O, and under Contract NNM10AA11C issued through the NASA New Frontiers Program.
Associate Editor: Michael A. Meyer