
Abstract
Introduction
SMC3, a core component of the cohesin complex, plays diverse roles in tumorigenesis across different cancer types. However, its expression pattern, prognostic significance, and potential association with the immune microenvironment in gliomas remain largely unexplored.
Methods
In this retrospective study, we systematically analyzed SMC3 expression using multiple transcriptomic datasets, including CGGA (CGGA-325, CGGA-301), TCGA, and GEO (GSE4290, GSE50161), and validated protein-level expression in 44 clinical glioma specimens via immunohistochemistry. Associations with overall survival (OS) were assessed using Kaplan–Meier and Cox regression analyses. Immune microenvironment features were evaluated using ESTIMATE and TIMER2.0, and pathway enrichment was analyzed through GSEA. Core B cell–related genes and signaling pathways associated with SMC3 were further identified by intersecting SMC3-correlated genes with MSigDB C7 and KEGG B cell receptor signaling gene sets.
Results
SMC3 was significantly overexpressed in glioma tissues compared with normal brain, with higher expression in lower-grade gliomas (LGG) than glioblastoma (GBM). High SMC3 expression was associated with prolonged OS across CGGA-325, CGGA-301, and the Validation Cohort, and remained prognostically favorable in specific clinical subgroups, including MGMT-unmethylated and 1p/19q non-codeleted tumors. SMC3 high-expression tumors exhibited lower overall immune and stromal scores but showed significant positive correlations with B cell and plasma cell infiltration. GSEA revealed enrichment of PI3K–AKT–MYC– and MTORC1-related pathways in high SMC3 tumors. Intersection analyses identified four core B cell–related genes (AKT3, CHUK, GSK3B, PIK3CA) within the PI3K–Akt signaling pathway, suggesting SMC3 expression may be associated with B cell–related immune features.
Conclusions
SMC3 is highly expressed in gliomas and is associated with favorable prognosis. Its expression correlates with B cell–related immune infiltration and activation of PI3K–Akt–MYC signaling, highlighting a potential role in shaping local immune microenvironments. These findings provide novel evidence for SMC3 as a prognostic biomarker and suggest a potential association with B cell–mediated immune features in gliomas.
Introduction
Gliomas are the most common primary malignant tumors of the adult central nervous system (CNS), characterized by high incidence, aggressive behavior, and remarkable molecular and histological heterogeneity. In recent years, advances in molecular pathological diagnosis, surgical techniques, postoperative concurrent chemoradiotherapy, as well as certain targeted and immunotherapies, have modestly improved overall survival. However, the improvement remains limited, particularly for patients with glioblastoma (GBM), whose median overall survival is still less than 15 months.1,2 Currently, commonly used molecular markers in clinical practice, such as IDH mutation, MGMT promoter methylation, and 1p/19q codeletion, play important roles in glioma classification and treatment decisions but are insufficient to fully reflect the molecular heterogeneity of gliomas. Therefore, identifying novel molecular markers and elucidating their underlying mechanisms is of great significance for improving patient prognosis and developing new therapeutic strategies.
Structural Maintenance of Chromosomes 3 (SMC3) is a core component of the cohesin complex. Cohesin plays key roles in chromosomal cohesion, replication and segregation, DNA damage repair, and the three-dimensional topology of chromatin. 3 Recent studies have reported aberrant expression or functional alterations of SMC3 and other cohesin members in various solid tumors, potentially contributing to tumor progression through the regulation of chromatin structure and transcriptional activity. For example, SMC3 is a transcriptional target of β-catenin/TCF4 and is highly expressed in colorectal cancer tissues 4 ; its high expression is associated with poor prognosis in colorectal cancer patients. 5 Similarly, SMC3 is overexpressed in primary hepatocellular carcinoma and correlates with unfavorable outcomes6. In contrast, SMC3 mutations are frequently observed in hematologic malignancies such as primary acute myeloid leukemia and are associated with relatively favorable prognosis. 7 However, the expression pattern, functional role, and clinical significance of SMC3 in CNS tumors, especially gliomas, remain unclear.
The tumor immune microenvironment (TIME) is a critical factor influencing tumor progression and therapeutic response. Compared with peripheral solid tumors, gliomas present unique immunological challenges: protection by the blood–brain barrier, high tumor cell heterogeneity, and a markedly immunosuppressive local environment, which limit the efficacy of immune checkpoint inhibitors (ICIs). 8 Although multiple clinical trials of ICIs have been conducted in GBM, overall therapeutic improvement has been modest. 9 This limitation is associated with multiple mechanisms, including the barrier function of the blood–brain barrier, insufficient tumor antigen presentation, T cell exhaustion, and dominance of myeloid-derived immunosuppressive cells. 10 In this context, remodeling the immune microenvironment is considered a key approach to overcoming the bottleneck of GBM immunotherapy. Notably, although the CNS has long been considered an “immune-privileged” site, recent studies have identified the presence of tertiary lymphoid structures (TLS) in both mouse and human gliomas, suggesting that local immune states possess a degree of plasticity. 11 These findings provide new insights for improving glioma treatment through immune modulation.
Based on this background, the present study systematically evaluated the expression profile of SMC3 in gliomas and its association with prognosis by integrating multi-cohort transcriptomic analyses with immunohistochemical validation in clinical samples from the Validation Cohort. Additionally, we focused on the relationship between SMC3 and the tumor immune microenvironment, particularly B cell/plasma cell infiltration, and explored the key signaling pathways potentially affected, such as PI3K–AKT–mTOR and MYC. Through these investigations, we aim to characterize the potential role of SMC3 in glioma and provide evidence for its potential relevance to prognosis and immune features, thereby offering a basis for future molecular classification and immunotherapeutic strategies.
Materials and Methods
Data Acquisition and Expression Analysis
Transcriptomic data and clinical information of gliomas were obtained from The Cancer Genome Atlas (TCGA), the Chinese Glioma Genome Atlas (CGGA-325, CGGA-301), and the Gene Expression Omnibus (GEO; GSE4290, GSE50161). CGGA and GEO datasets were downloaded and curated from their respective official websites. Some expression plots were obtained from the TCGA online visualization platform. Differential gene expression analyses were performed using GraphPad Prism 9.0, with statistical comparisons conducted via two-sided t-test or one-way analysis of variance (ANOVA).The public transcriptomic datasets were mainly used for expression analysis, survival analysis, clinicopathological correlation analysis, immune microenvironment analysis, and pathway enrichment analysis.
To validate the expression of SMC3 protein, surgical specimens were collected from 44 glioma patients pathologically confirmed at the Second Hospital of Hebei Medical University between January 2016 and March 2021. This retrospective study was approved by the Research Ethics Committee of the Second Hospital of Hebei Medical University, Shijiazhuang, Hebei, China (Ethics Approval Number: 2019-R191). Written informed consent for the use of tissue samples and clinical information was obtained from the patients and/or their family members. All patient details were de-identified before analysis.Tumor tissues were formalin-fixed and paraffin-embedded (FFPE) for immunohistochemistry (IHC) using a rabbit anti-SMC3 polyclonal antibody (bs-5735R, Bioss Antibodies) at a dilution of 1:200. Histological images were acquired using a Leica inverted optical microscope and a digital whole-slide pathology scanner from Ningbo Jiangfeng Bio. Semi-quantitative scoring of IHC slides was performed according to the Allred scoring system 12 by two independent pathologists. The Allred score comprises the proportion score (PS) and intensity score (IS): PS ranges from 0–5 (0 = 0%; 1 = <1%; 2 = 1–10%; 3 = 11–33%; 4 = 34–66%; 5 = ≥67%), and IS ranges from 0–3 (0 = no staining; 1 = weak; 2 = moderate; 3 = strong). The total score is calculated as PS + IS (0–8), and samples were subsequently classified into high- or low-expression groups based on the total score.The Validation Cohort was selectively included according to the following criteria: histopathologically confirmed glioma after surgery, WHO grade II–IV, age ≥18 years, receipt of standardized comprehensive treatment including surgery, radiotherapy, and chemotherapy, and complete clinical records. Patients with incomplete clinical data or other malignant tumors were excluded. The Validation Cohort was primarily used for protein-level validation by immunohistochemistry and supportive survival analysis. The reporting of this study conforms to REMARK guidelines. 12
Survival Analysis
To evaluate the prognostic impact of SMC3 expression, survival analyses were performed using CGGA-325, CGGA-301, and the Validation Cohort. Patients were divided into high- and low-expression groups based on the median SMC3 expression. Kaplan–Meier survival curves were generated using the R software (version 4.3.3) with the survminer and survival packages, and differences were compared using the log-rank test. Overall survival (OS) was defined as the time from the date of pathological diagnosis to death from any cause or the last follow-up.Univariate and multivariate Cox regression analyses included clinical variables (age, sex, WHO grade, IDH mutation status, 1p/19q codeletion, etc.), with results reported as hazard ratios (HRs) and 95% confidence intervals (CIs).Because of the substantial missingness of key molecular variables in the Validation Cohort, multivariable Cox regression was not used for definitive prognostic inference in that cohort.
Stratified Survival Analysis
To further assess the prognostic value of SMC3 across different clinical subgroups, stratified analyses were conducted based on the CGGA-325 dataset. Subgroup variables included IDH mutation status, 1p/19q codeletion status, MGMT promoter methylation status, age (≤42 years vs. >42 years), sex, and tumor grade. Kaplan–Meier curves and log-rank tests were performed in R.
Correlation Analysis With Clinical Features
The association between SMC3 expression and clinical features was analyzed using CGGA-325 and CGGA-301 datasets. Variables included 1p/19q codeletion status, IDH mutation status, MGMT methylation status, age, and tumor grade. Between-group comparisons were conducted using the Wilcoxon rank-sum test or Kruskal–Wallis test, and visualization was performed using the ggplot2 package in R.
Immune Microenvironment Analysis
To evaluate the relationship between SMC3 and the tumor immune microenvironment, StromalScore, ImmuneScore, ESTIMATE Score, and Tumor Purity were calculated using the ESTIMATE algorithm (R package “estimate”). Enrichment patterns of immune-related pathways were visualized using the GSVA package.
Furthermore, the TIMER2.0 online platform (https://timer.cistrome.org/) was used to assess correlations between SMC3 expression and immune cell infiltration, incorporating multiple algorithms, including TIMER, EPIC, and MCPCOUNTER. Correlations were calculated using Spearman’s correlation coefficient, with a significance threshold of P < 0.05.
Pathway Enrichment and Key Gene Intersection Analysis
Differentially expressed genes (DEGs) were identified in the CGGA-325 cohort by grouping samples according to SMC3 expression using the limma package, followed by gene set enrichment analysis (GSEA). Significance criteria were normalized enrichment score (NES), FDR < 0.25, and adjusted P value < 0.05.
SMC3-related gene sets were intersected with the MSigDB C7 immune signature gene sets (B cell-related) to identify overlapping B cell-associated genes. These genes were further intersected with the KEGG_B_CELL_RECEPTOR_SIGNALING_PATHWAY gene set to obtain a core pathway gene set.
Statistical Analysis
All statistical analyses were conducted using R software (v4.3.3) and GraphPad Prism (v9.0). Between-group comparisons were performed using Student’s t-test or Mann–Whitney U test according to data distribution. Correlations between gene expression and immune cell infiltration or clinical features were analyzed using Spearman or Pearson correlation tests. Survival analyses were conducted using Kaplan–Meier curves and log-rank tests, with Cox proportional hazards models applied to evaluate prognostic factors. ESTIMATE analyses were performed using the “estimate” R package, and immune cell infiltration data were obtained from TIMER2.0. All tests were two-sided, and P < 0.05 was considered statistically significant.
Results
SMC3 Is Highly Expressed in Gliomas
Baseline Characteristics of Datasets
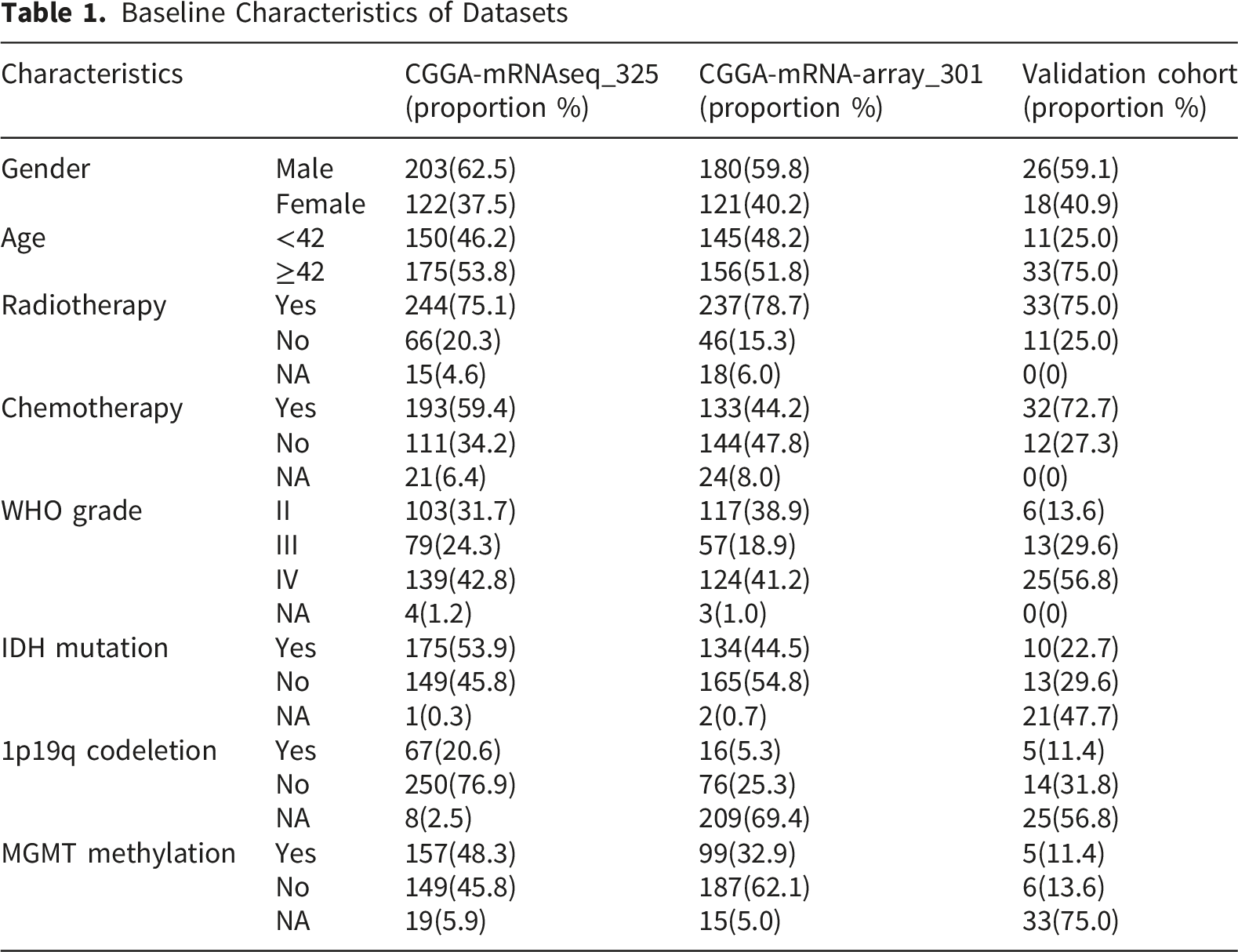
Expression of SMC3 in Glioma.(A) CGGA-325 dataset.(B) GEO dataset (GSE4290).(C) GEO dataset (GSE50161).(D) TCGA dataset.(E) Immunohistochemical image showing significantly enhanced SMC3 expression in a case of grade III glioma tumor tissue from the Validation Cohort.(F) Comparison of SMC3 expression between high-expression and low-expression groups.(G) Comparison of SMC3 expression in tumor tissues from GBM and LGG
To validate protein-level expression, we performed immunohistochemistry (IHC) on 44 glioma specimens collected at the Validation Cohort. SMC3 was predominantly localized in the tumor cell nucleus. A representative image from a WHO grade III glioma demonstrated markedly stronger staining in tumor tissues (Figure 1E). Quantitative scoring confirmed that high-expression cases had significantly elevated SMC3 levels compared with low-expression cases (P < 0.0001; Figure 1F). Consistent with the transcriptomic data, LGG samples exhibited higher SMC3 expression than GBM (P = 0.016; Figure 1G).
Association of SMC3 Expression With Prognosis
Kaplan–Meier survival analyses across the CGGA-325, CGGA-301, and Validation Cohort demonstrated that patients with high SMC3 expression had significantly longer overall survival (OS) than those with low expression (CGGA-325: P < 0.001, Figure 2A; CGGA-301: P = 0.006, Figure 2C; Validation Cohort: P = 0.018, Figure 2E). Relationship between SMC3 Expression and Prognosis in Glioma Patients.(A) Survival analysis in the CGGA-325 dataset.(B) Univariate Cox regression analysis in the CGGA-325 dataset.(C) Survival analysis in the CGGA-301 dataset.(D) Univariate Cox regression analysis in the CGGA-301 dataset.(E) Survival analysis in Validation Cohort.(F) Univariate Cox regression analysis in the Validation Cohort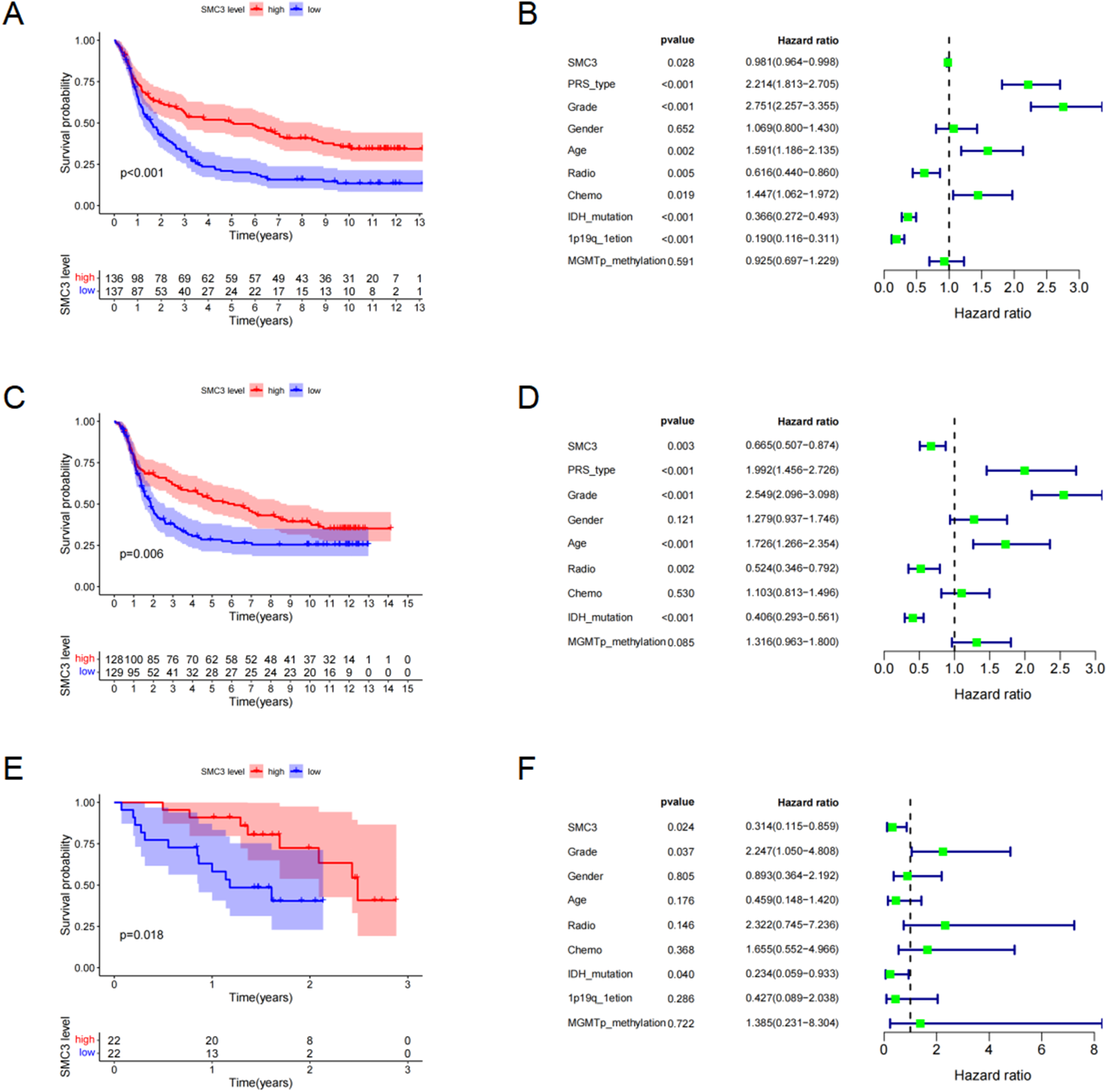
Univariate Cox regression further supported the protective role of SMC3 expression (CGGA-325: HR = 0.981, 95% CI: 0.964–0.998, P = 0.028; Figure 2B; CGGA-301: HR = 0.665, 95% CI: 0.507–0.874, P = 0.003; Figure 2D; Validation Cohort: HR = 0.314, 95% CI: 0.115–0.859, P = 0.024; Figure 2F). These findings suggest that high SMC3 expression is associated with favorable prognosis in glioma.
However, multivariate Cox regression adjusting for age, sex, WHO grade, IDH mutation status, and 1p/19q codeletion showed that SMC3 expression did not retain independent prognostic significance across the public datasets (Supplementary Figure 1), suggesting its prognostic value may be confounded by established clinical parameters.Because the Validation Cohort contained substantial missing data for several key molecular variables, it was not used for definitive multivariable prognostic inference.
Stratified Survival Analyses
To clarify the prognostic role of SMC3 across clinical subgroups, we performed stratified analyses in the CGGA-325 cohort. High SMC3 expression predicted significantly improved OS in the MGMT promoter unmethylated subgroup (P < 0.0001) and in the 1p/19q non-codeleted subgroup (P = 0.035; Figure 3A–B). Age-stratified analyses revealed consistent survival benefits in both younger (≤42 years: P = 0.0067) and older (>42 years: P = 0.022) patients (Figure 3C–D). Furthermore, survival benefits were evident in male patients (P < 0.0001) and in patients with LGG (P = 0.0071; Figure 3E–F). No significant survival differences were observed in other subgroups. Stratified Survival Analysis of SMC3 Expression.(A) Non-methylation subgroup.(B) Non-1P19q deletion subgroup.(C) ≤42 years subgroup.(D) >42 years subgroup.(E) Male subgroup.(F) Low-grade glioma subgroup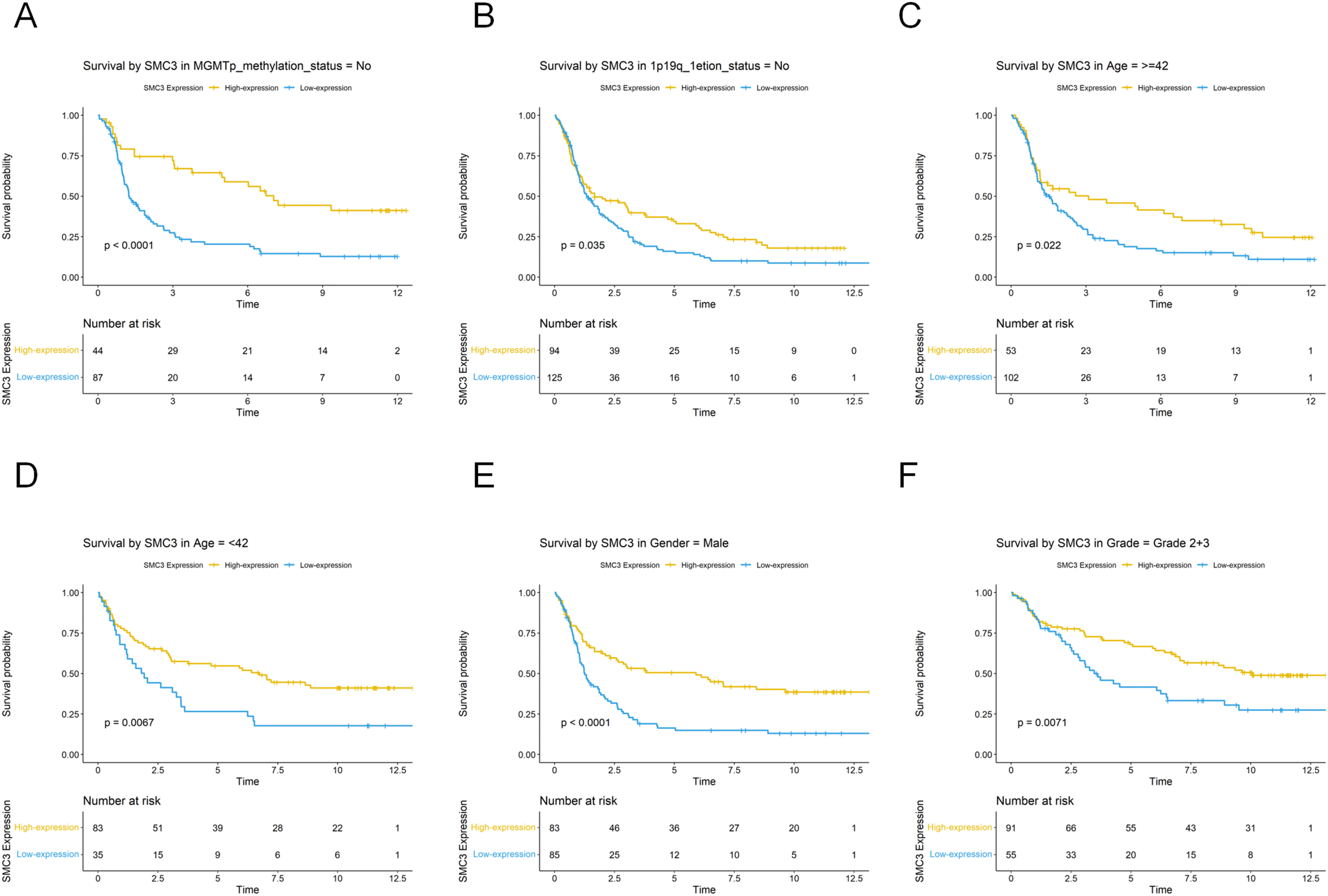
These findings indicate that although SMC3 was not an independent prognostic marker in multivariate analysis, it retains prognostic relevance in specific clinical contexts. Thus, SMC3 may serve as a useful biomarker for risk stratification and precision management of glioma.
Correlation of SMC3 Expression With Clinical Characteristics
We next evaluated associations between SMC3 expression and clinicopathologic features. In the CGGA-325 cohort, SMC3 expression was significantly higher in patients with 1p/19q codeletion (P < 0.001; Figure 4A), age <42 years (P < 0.001; Figure 4B), MGMT promoter methylation (P < 0.01; Figure 4C), and IDH mutation (P < 0.01; Figure 4D). LGG patients also expressed higher levels of SMC3 than GBM patients (P < 0.01; Figure 4E). These associations with age, IDH mutation, and tumor grade were further validated in the CGGA-301 cohort (all P < 0.05; Supplementary Figure 2). Collectively, these results highlight that SMC3 expression is closely linked to both tumor grade and molecular subtypes. Correlation of SMC3 Expression with Clinical Features.(A) Correlation analysis between 1P19q deletion status and SMC3 expression.(B) Correlation analysis between age and SMC3 expression.(C) Correlation analysis between MGMT methylation status and SMC3 expression.(D) Correlation analysis between IDH mutation status and SMC3 expression.(E) Correlation analysis between glioma grade and SMC3 expression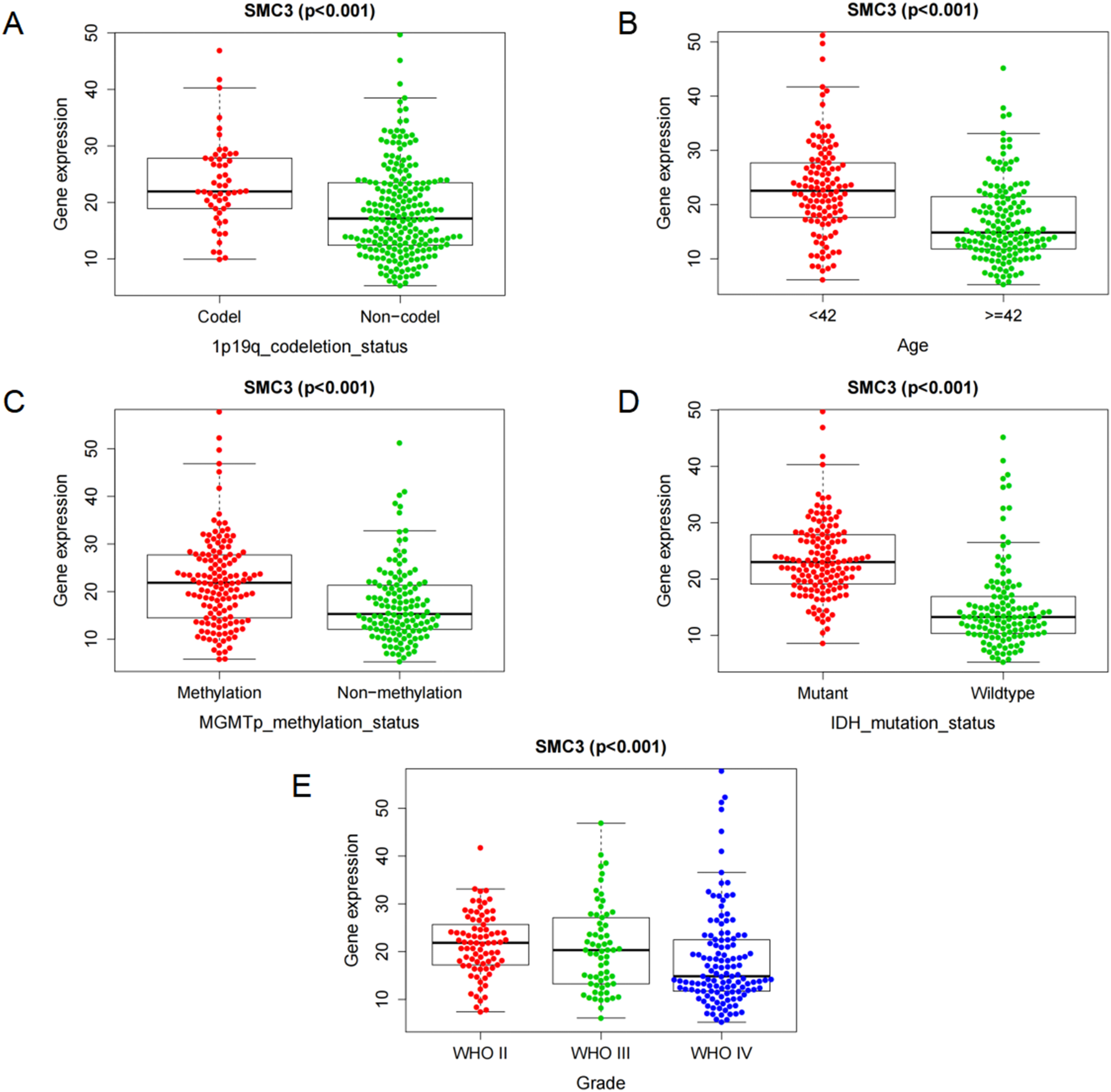
Association of SMC3 Expression With the Immune Microenvironment
Using the ESTIMATE algorithm, we found that low SMC3 expression was associated with significantly higher ImmuneScore, StromalScore, and ESTIMATE score, but with lower tumor purity in both the CGGA-325 (Figure 5C–F) and CGGA-301 cohorts (Figure 5G–J; all P < 0.001). Heatmaps further revealed that multiple immune-related pathways were enriched in the low-expression group across both cohorts (Figure 5A–B). These findings suggest that low SMC3 expression corresponds to a more immune- and stroma-rich microenvironment, whereas high SMC3 expression is linked to increased tumor cell proportion. Relationship between SMC3 Expression and the Immune Microenvironment.(A) Heatmap from the CGGA-325 dataset.(B) Heatmap from the CGGA-301 dataset.(C) Tumor purity in the CGGA-325 dataset.(D) StromalScore in the CGGA-325 dataset.(E) ESTIMATE score in the CGGA-325 dataset.(F) ImmuneScore in the CGGA-325 dataset.(G) Tumor purity in the CGGA-301 dataset.(H) StromalScore in the CGGA-301 dataset.(I) ESTIMATE score in the CGGA-301 dataset.(J) ImmuneScore in the CGGA-301 dataset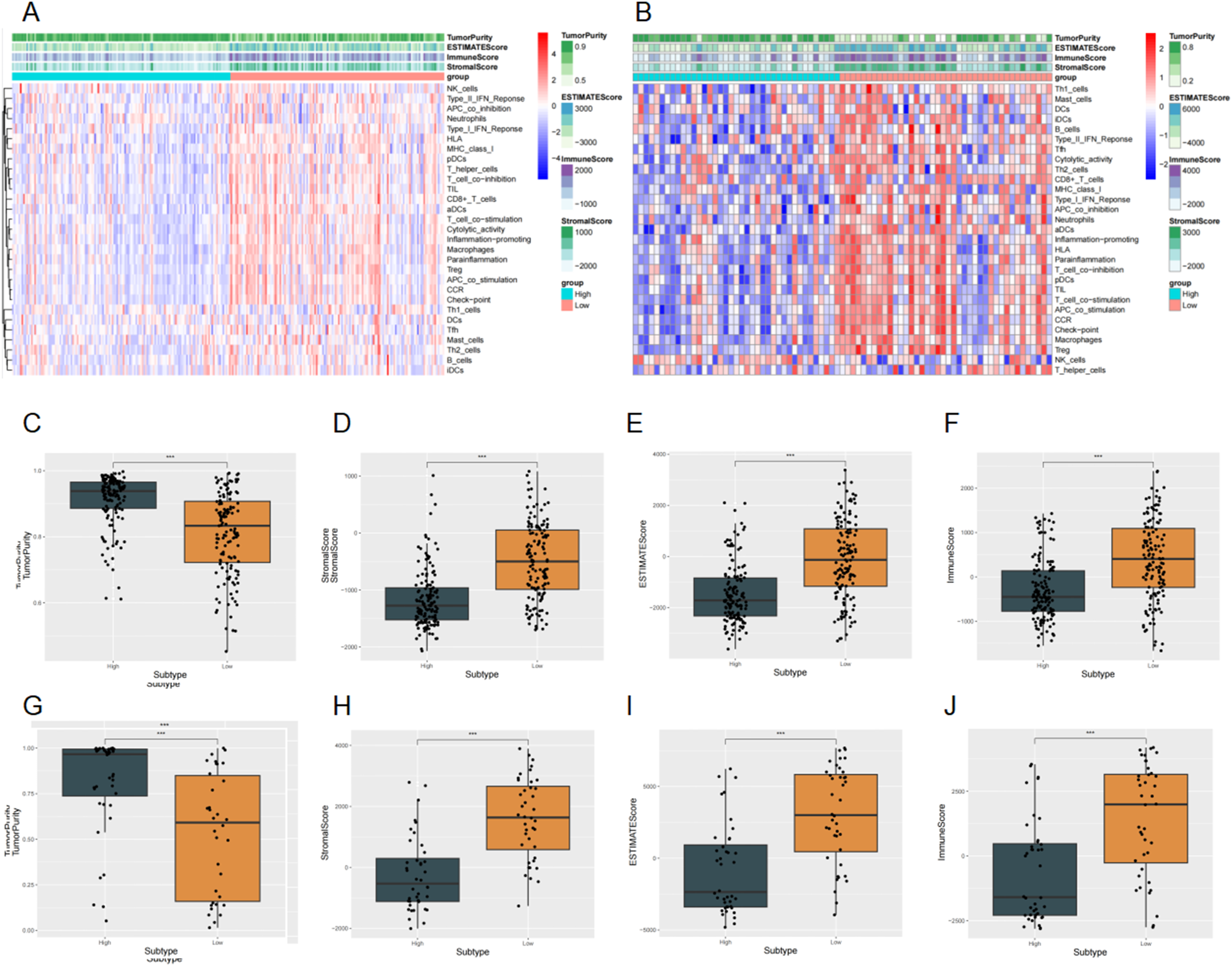
Consistently, analysis via TIMER2.0 revealed a significant positive correlation between SMC3 expression and B cell infiltration (Spearman R > 0.25, P < 0.001; Figure 6A, C–D), with results stable across multiple algorithms. Moreover, SMC3 expression correlated with plasma cell infiltration (Spearman R = 0.358, P < 0.001; Figure 6B), whereas associations with most T cell subsets (e.g., CD8+ T cells, Th1 cells) were weak or nonsignificant. Correlation of SMC3 Expression with Immune Cell Infiltration.(A) SMC3 expression is significantly positively correlated with B cell infiltration (EPIC).(B) SMC3 expression is significantly positively correlated with plasma cell infiltration (XCELL).(C–D) SMC3 expression is significantly positively correlated with B cell infiltration (QUANTISEQ)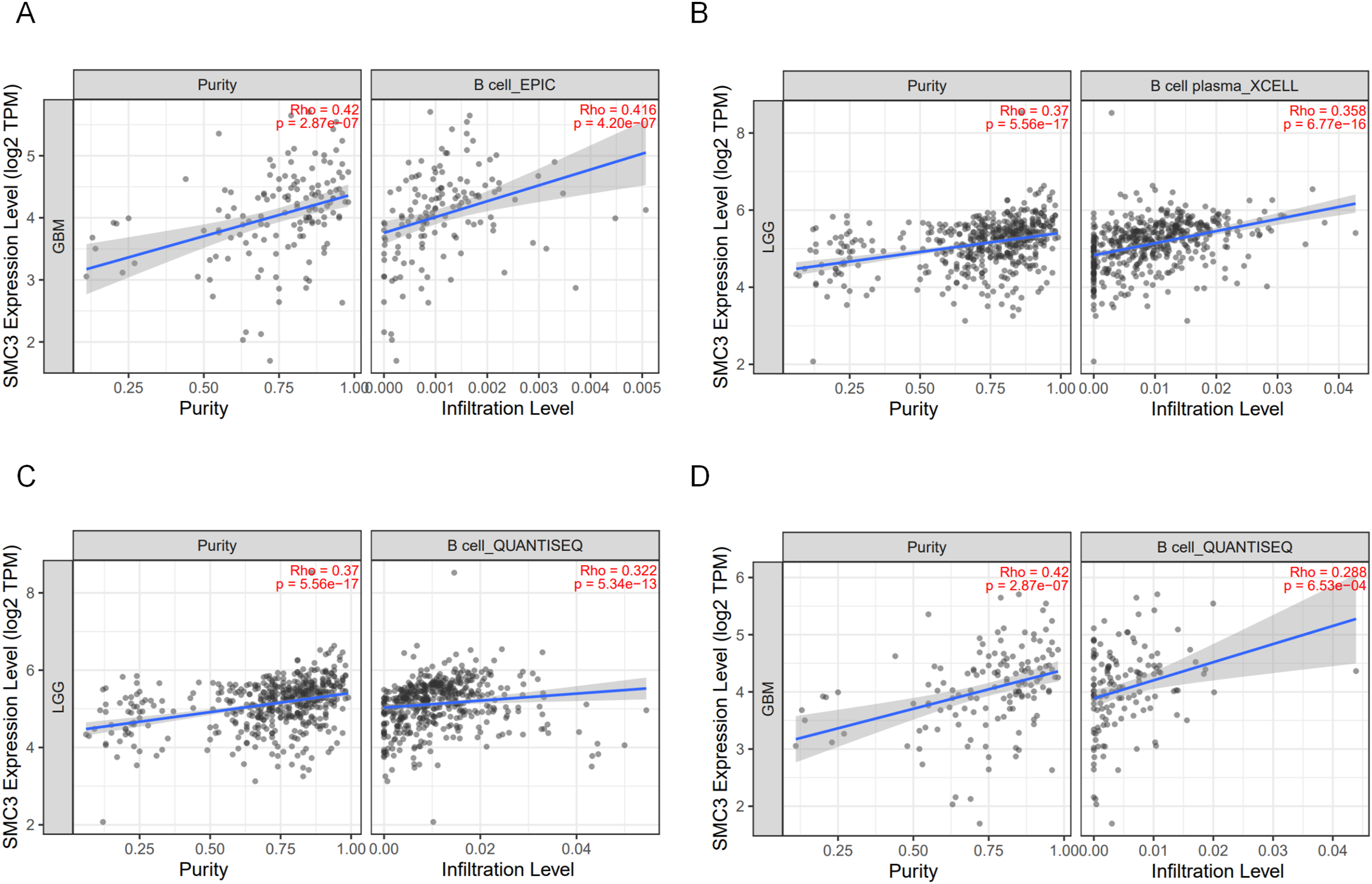
Taken together, these findings indicate that SMC3 expression reflects not only tumor-to-stroma/immune composition but also is associated with glioma immune responses, particularly those related to B cell–associated mechanisms, thereby influencing clinical outcomes.
Pathway Enrichment and Key Gene Intersection Analysis
To further investigate the molecular mechanisms underlying SMC3 function in glioma, we performed GSEA in the CGGA-325 cohort stratified by SMC3 expression. Gene sets enriched in the SMC3 high-expression group were predominantly involved in B cell–related signaling pathways, including the PI3K–Akt signaling pathway (Figure 7A), MTORC1 signaling (Figure 7B), MYC_TARGETS_V1 (Figure 7C), and MYC_TARGETS_V2 (Figure 7D). Pathway Enrichment Analysis Related to SMC3.(A) PI3K-Akt signaling pathway.(B) MTORC1 signaling pathway.(C) MYC_TARGETS_V1 signaling pathway.(D) MYC_TARGETS_V2 signaling pathway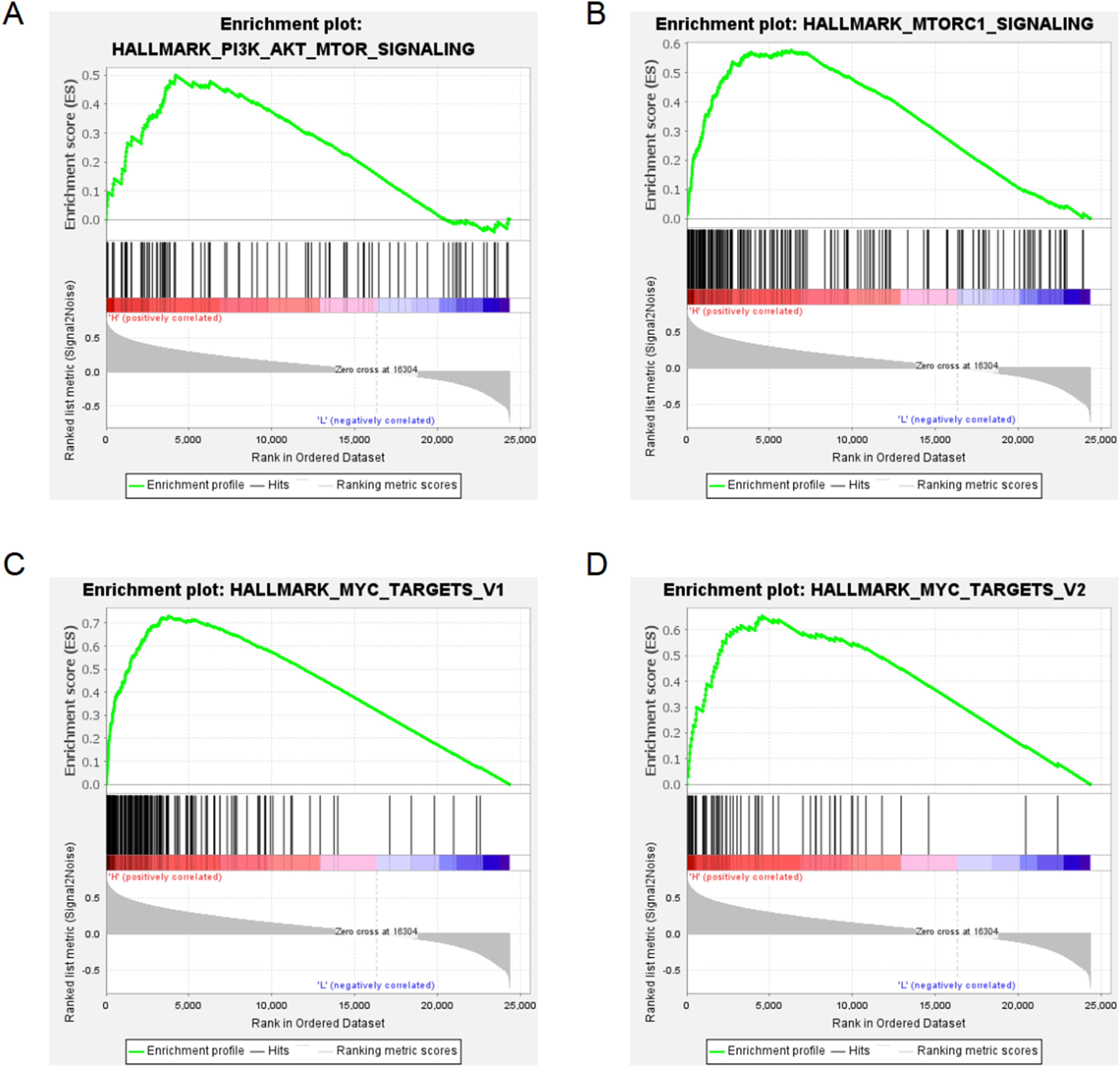
Correlation analysis (Cor > 0.5) identified a set of genes significantly associated with SMC3 expression. Intersecting these with MSigDB C7 B cell–related gene sets yielded 629 overlapping genes, suggesting strong relevance of SMC3 to B cell–related immune features. Further intersection with the KEGG B cell receptor signaling pathway (KEGG_B_CELL_RECEPTOR_SIGNALING_PATHWAY; Figure 8A) identified four hub genes: CHUK (Figure 8B), GSK3B (Figure 8C), AKT3 (Figure 8D), and PIK3CA (Figure 8E). Notably, all four converge on the PI3K–Akt signaling pathway. Core Genes Related to SMC3 Expression.(A)Venn diagram showing the intersection of SMC3-related gene sets and B cell-related gene sets.(B) Correlation analysis between SMC3 and CHUK.(C) Correlation analysis between SMC3 and GSK3B.(D) Correlation analysis between SMC3 and AKT3.(E) Correlation analysis between SMC3 and PIK3CA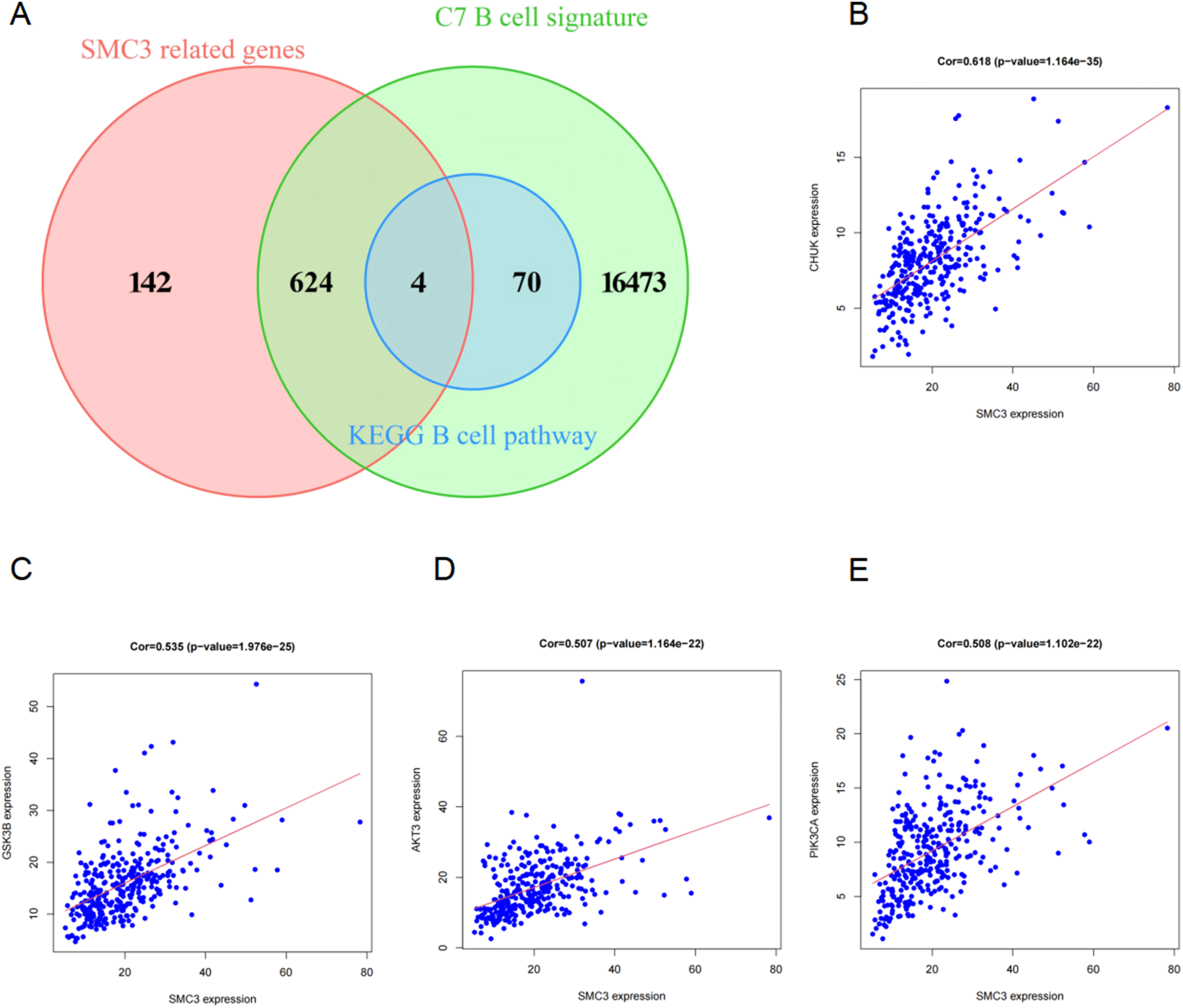
Collectively, these results suggest that SMC3 may be associated with glioma progression and B cell–related immune features, in part through the PI3K–Akt signaling pathway.
Discussion
SMC3, as a key subunit of the cohesin complex, exhibits distinct functional roles across different tumor types. Previous studies have shown that in colorectal cancer, the β-catenin/TCF4 pathway directly drives SMC3 transcription, and its high expression promotes cellular transformation and proliferation, supporting its tumor-promoting role in WNT-driven cancers 4 . However, the function of SMC3 is not consistent across tissues. For example, in pancreatic ductal adenocarcinoma (PDAC), SMC3 can bind to enhancer regions of the Rab27a gene to promote its expression, whereas epigenetic silencing of SMC3 (e.g., via methylation) reduces Rab27a expression, thereby promoting PDAC progression, suggesting that loss of SMC3 function in this context is associated with disease advancement. 13 In lung cancer, although mechanistic studies are limited, both in vitro and in vivo experiments indicate that SMC3 is associated with cell proliferation and drug responsiveness, such as hydrogen treatment downregulating SMC3 and inhibiting tumor growth. 14 Therefore, SMC3 may act as either a tumor-promoting factor or, when functionally lost, contribute to disease progression depending on tissue context, highlighting its tissue-dependent roles. Currently, the function and molecular mechanisms of SMC3 in gliomas remain largely unexplored, representing an important area for both research and potential translational application.
In this study, we systematically analyzed SMC3 expression and its prognostic significance in gliomas by integrating multiple transcriptomic cohorts and validating findings with clinical samples. Our results demonstrated that SMC3 is generally highly expressed in glioma tissues, with expression higher in LGG than in GBM. Kaplan–Meier and Cox regression analyses indicated that patients with high SMC3 expression had longer overall survival, suggesting a potential protective role in gliomas. Although SMC3 did not emerge as an independent prognostic factor in multivariate analyses across the overall cohort, high expression remained associated with improved prognosis in specific clinical subgroups, such as MGMT-unmethylated and 1p/19q non-codeleted tumors. This pattern contrasts sharply with the typical “high expression = poor prognosis” observed in most solid tumors,5,6suggesting that the biological significance of SMC3 may be context-dependent in glioma. This discrepancy may partly reflect the unique chromatin organization of glial lineage cells, the distinct transcriptional regulatory landscape of the central nervous system, and the specific molecular background associated with lower-grade gliomas. In addition, because SMC3 expression in our cohorts was associated with several favorable clinicopathological features, its prognostic significance in glioma should be interpreted within this disease-specific biological context rather than directly extrapolated from other solid tumors. 15
Further analysis revealed that high SMC3 expression was associated with lower immune and stromal scores, indicating generally lower immune infiltration in glioma tissues and an “immune-cold” phenotype. This is consistent with prior observations that T cell infiltration in gliomas is generally insufficient, which limits the efficacy of immune checkpoint inhibitors (ICIs) in unselected patients.16,17 Interestingly, within this immunosuppressive environment, high SMC3 expression was significantly positively correlated with B cell and plasma cell infiltration, suggesting a potential role in local immune regulation. Subsequent GSEA indicated that SMC3 high-expression–related genes were enriched in PI3K–AKT–mTOR, MYC, and MTORC1 pathways, which are central to B cell activation, differentiation, and antibody production.18-21 These findings suggest that SMC3 may be associated with a distinct local immune context characterized by B cell– and plasma cell–related features.
Previous studies have demonstrated that cohesin plays a critical role in chromatin remodeling and regulation of immune-related gene expression,22,23 and extensive cross-cancer evidence indicates that B cells and tertiary lymphoid structures (TLS) are closely associated with enhanced ICI response and survival benefit.24-26 For instance, in melanoma and renal cell carcinoma, patients with abundant TLS-associated B cell infiltration show higher ICI response rates and improved survival compared with TLS-deficient patients. 24 Similarly, TLS-related gene signatures in sarcoma and breast cancer are associated with better immunotherapy outcomes and long-term survival.25,26 These studies suggest that B cells and TLS are not merely bystanders but important components of anti-tumor immunity. Functional TLS have also been identified in brain tumors, indicating that B cells may play a key role in local antigen presentation and humoral immunity, forming structures analogous to peripheral lymphoid organs.11,27,28 This underscores the plasticity of local immune responses. In this context, the positive association observed here between SMC3 expression and B-cell/plasma-cell infiltration raises the possibility that SMC3 may be linked to TLS-related local immune organization in glioma. However, because the present study did not directly evaluate TLS markers, spatial immune architecture, or multiplex pathological features, this interpretation should be regarded as hypothesis-generating rather than mechanistically established.
Integrating molecular pathways and immune infiltration features, we propose a possible mechanistic model: at the topological level, SMC3, as a critical component of the cohesin complex, may influence chromatin loop structures and enhancer–promoter interactions, selectively enhancing transcription of immune-related genes; at the signaling level, SMC3 expression may be associated with downstream PI3K–AKT–MYC pathway activation following BCR signaling, which is central to B cell proliferation, differentiation, and antibody production; at the cellular level, previous animal studies indicate that Smc3 expression affects B cell fate decisions and immune function within germinal centers. 29 These mechanisms align with our clinical observations that high SMC3 expression correlates positively with B cell and plasma cell infiltration, suggesting that SMC3 may represent a potential link between chromatin structure regulation with immune function.
Notably, the biological effects of the PI3K–MYC pathway are cell type–dependent: in tumor cells, activation typically drives metabolic reprogramming and proliferation, promoting tumor progression; in B cells or TLS regions, activation is more likely to reflect enhanced humoral immune responses, facilitating antigen presentation and antibody production, thereby improving immune efficacy.30-32 This duality may explain our observations in gliomas, where high SMC3 expression is associated with PI3K–MYC pathway enrichment yet correlates with better prognosis.
This study has several limitations. First, the mechanistic depth remains limited; we only observed correlations between SMC3 and B cell/plasma cell infiltration in multi-cohort analyses, and enrichment of PI3K–AKT–MYC/MTORC1 pathways in GSEA, without mechanistic validation at the cellular or animal level. Second, our analyses are primarily correlative and enrichment-based, precluding causal inference, which requires functional validation through SMC3 and key pathway perturbation studies.Third, the Validation Cohort was relatively small and contained incomplete clinicomolecular data, which limited more comprehensive prognostic modeling and restricted its role to supportive clinical validation.
Conclusion
This study shows that SMC3 is generally highly expressed in gliomas and is associated with improved patient survival. Unlike the typical “high expression=poor prognosis” pattern in most solid tumors, SMC3 shows a distinct prognostic association in gliomas. Further analyses suggest that high SMC3 expression, although associated with lower overall immune infiltration, is positively correlated with B cell and plasma cell infiltration and enriched in B cell–related pathways. These findings indicate that SMC3 may be associated with B cell–related immune responses through chromatin structure and immune signaling–related mechanisms. This study provides novel evidence for the potential role of SMC3 in gliomas and its potential as a prognostic and immune biomarker.
Supplemental Material
Supplemental material - SMC3 Is Highly Expressed in Gliomas and Is Associated With Favorable Prognosis and B Cell–Related Immune Features
Supplemental material for SMC3 Is Highly Expressed in Gliomas and Is Associated With Favorable Prognosis and B Cell–Related Immune Features by Wei Han, Lei Tian, Haonan Li, Yadong Liu, Xuetao Han, Liubing Hou, Zhiqing Xiao, Huandi Zhou and Xiaoying Xue in Technology in Cancer Research & Treatment.
Footnotes
Acknowledgments
The authors thank all colleagues who contributed to this study.
Ethical Considerations
This retrospective study was approved by the Research Ethics Committee of the Second Hospital of Hebei Medical University, Shijiazhuang, Hebei, China (Ethics Approval Number: 2019-R191). Written informed consent for the use of tissue samples and clinical information was obtained from the patients and/or their family members.
Author Contributions
Wei Han, Huandi Zhou, and Xiaoying Xue conceived and supervised the study. Wei Han, Lei Tian, and Haonan Li collected the data. Wei Han drafted the manuscript. Huandi Zhou and Xiaoying Xue critically revised the manuscript for important intellectual content. All authors read and approved the final manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publicationof this article: This work was supported by Hebei Province Medical-Research-Enterprise Joint Innovation Special Project Program (Grant Number LH20250063), the Medical Science Research Project of Health Commission of Hebei Province (Grant Number 20211393) and the Youth Natural Science Foundation of Hebei Province (Grant Number H2025307026).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The datasets are available from the corresponding author on reasonable request.
Supplemental Material
Supplemental material for this article is available online.
Appendix
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.