
Abstract
A novel preharvest method for producing antimicrobial kale in hydroponic systems was developed using a low-concentration plai oil nanoemulsion. The nanoemulsion was applied to the hydroponic solution at concentrations of 5, 10, 15, and 20 μL/mL for 12 h prior to harvest. Antimicrobial activity against Listeria monocytogenes was evaluated in both treated and control kale samples. In addition, bioactive compounds, volatile oil residues, and the potential mechanism of antimicrobial action were investigated. Application of 10 μL/mL plai oil resulted in a reduction of L. monocytogenes by more than 5 log10 colony-forming units per gram and completely inhibited disease incidence during storage at 4°C for at least 7 days. Although higher concentrations (15 and 20 μL/mL) also reduced bacterial counts, increased wilting was observed in the leaves. In contrast, the control group developed visible disease symptoms within 3 days of storage. Based on microbial suppression and visual quality, 10 μL/mL was identified as the optimal concentration. The preharvest treatment significantly enhanced the levels of bioactive compounds and antioxidant activity compared with control kale. Volatile constituents of plai oil, including thujene (35%), terpinene (21%), pinene (15%), and terpinene-4-ol (14%), were detected in treated kale. The antimicrobial mechanism was associated with membrane disruption, as evidenced by porous and shriveled cell walls observed in L. monocytogenes, along with decreased pH, increased conductivity, and elevated protein and amino acid leakage after 18 h of inoculation. Biofilm formation by L. monocytogenes on kale was also significantly inhibited. This preharvest strategy offers a practical and effective method for enhancing microbial safety and extending the shelf life of hydroponically grown kale.
Introduction
Kale (Brassica oleracea L.), often referred to as a “superfood” and the “queen of greens,” has been widely recognized for its high nutritional value and associated health benefits. It is rich in essential vitamins, minerals, and key nutrients (Gao et al., 2025). Additionally, kale provides phytochemicals such as lutein, violaxanthin, chlorophyll a, and chlorophyll b, along with dietary fiber and various antioxidants, including carotenoids, flavonoids, and folate (Ashenafi et al., 2022). This leafy vegetable can be cultivated through hydroponic systems, in which plants are grown in nutrient-enriched water (Setka et al., 2021). Under such conditions, the cultivation period typically ranges from 40 to 50 days. With the increasing demand, kale is widely grown and consumed, particularly in fresh salads. However, hydroponic systems, including irrigation water, nutrient solutions, seeds, seedlings, and substrates, have been identified as potential sources of microbial contamination, which may shorten the shelf life of the harvested vegetables (Klištincová et al., 2024).
The consumption of raw kale or its storage under refrigeration presents a potential risk of contamination by foodborne pathogens, particularly Listeria monocytogenes, which is capable of proliferating on refrigerated fresh produce. Although ultrasound treatment and sodium hypochlorite washing have been effectively used to sanitize fresh vegetables (Lee et al., 2014), the use of essential oils has emerged as a promising alternative. Essential oils exhibit broad-spectrum antimicrobial activity and, when applied at low concentrations, can also enhance the flavor profiles of vegetables (Parichanon et al., 2022). Among these, plai oil has shown promise due to its aromatic compatibility with vegetables and its strong antibacterial properties, which make it suitable for application in fresh produce (Matan and Kongchoosi, 2024). The concept of using essential oils at low concentrations to facilitate their absorption into plant tissues as natural antimicrobial agents has gained increasing interest. In the present study, a novel preharvest application of low-concentration plai oil was investigated in a hydroponic kale cultivation system to enhance the antimicrobial resistance and shelf life of kale after harvest and storage at 4°C for fresh kale salads. This approach is expected to benefit both farmers, by enabling the delivery of safer produce to the market, and consumers, by reducing spoilage and the risk of foodborne illness.
Materials and Methods
Medium and chemicals used
All chemicals for bioactive compound analysis and antioxidant activity were purchased from Millipore Sigma (Merck Co., Ltd., Thailand). Nutrient agar (NA), nutrient broth (NB), plate count agar (PCA), malt extract agar (MEA), De Man–Rogosa–Sharpe agar (MRS), and peptone water were obtained from Merck, Thailand. Compact Dry EC and LM were procured from Nissui Pharmaceutical Co., Ltd., Japan.
Plai nanoemulsion preparation
Plai oil was obtained from Thai–China Flavours and Fragrances Industry (Thailand). A 50 L nanoemulsion of plai oil at concentrations of 5, 10, 15, and 20 μL/mL was prepared using deionized water and homogenized with an ultrasonic homogenizer (Tefic Biotech, China) for 20 min until a stable emulsion was formed. The emulsions were used immediately for testing.
Effect of preharvest kale treatment with plai oil on growth of L. monocytogenes, disease incidence, and wilting stage
Kale seeds (100 seeds per concentration; three replications) were placed on moistened germination gauze for 5 days at 4°C to promote germination. The seeds were then incubated for 7 days on sponge cubes (2.0 cm × 2.0 cm × 2.0 cm) in a seedling tray placed above the hydroponic boxes at room temperature (30 ± 2°C, 80% relative humidity). Each hydroponic tray (60) contained 20 plants spaced 15 cm apart. The plants were grown hydroponically in a nutrient solution (electrical conductivity [EC] = 1 mS cm−1; pH 5.8–6.2) for 50 days. On day 50, before harvest, each concentration of plai oil nanoemulsion was added to the nutrient solution tank. A pump was used to feed the plai oil into the hydroponic tray, and the solution was circulated for 12 h to prevent the separation of plai oil and water. After 12 h, the pump system was turned off, and the kale was harvested (Fig. 1). The control group was irrigated with deionized water in the hydroponic system. The kale leaves were then measured for various parameters, as follows:
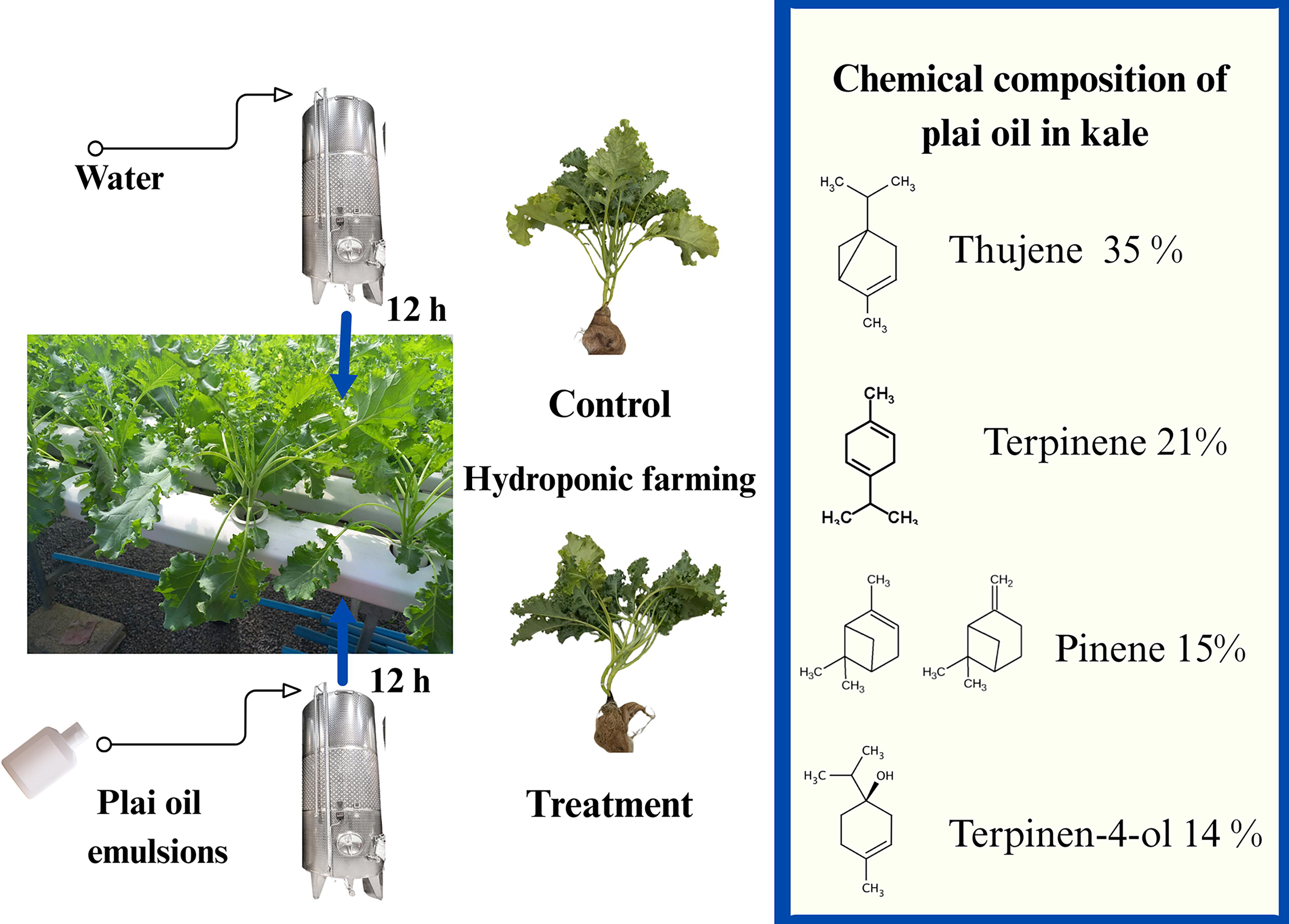
Preharvest experiments involving the feeding of plai oil into a hydroponic kale cultivation system and analysis of the chemical composition of plai oil detected in kale.
Survival of L. monocytogenes on kale
L. monocytogenes, isolated from vegetables, was obtained from the Microbiology Laboratory at Walailak University, Thailand. A 100 mL culture was grown in NB at 37°C for 24 h to achieve a concentration of 106 colony-forming units (CFU)/mL. Treated and control kale leaves were then dipped in the L. monocytogenes solution and placed in a biosafety cabinet (Thermo Fisher Scientific, MA) for 1 h to allow the pathogen to adhere. The quantity of L. monocytogenes on the kale at different concentrations and in the control group was then counted using NA at 1, 3, 6, 9, 12, 15, 18, 21, and 24 h. The results were reported as log10 CFU/g. Ten kale plants were randomly sampled, and the experiment was repeated three times.
Disease Incidence
The measurement of disease incidence, observed as disease spots appearing on the surface of kale, was evaluated using the method of Parichanon et al. (2022) with minor modifications. Disease incidence was assessed on days 1, 3, 5, and 7 after storage at 4 ± 2°C. Kale from each treatment group (n = 10) was replicated three times. Disease incidence (%) was then calculated using the following Equation 1.
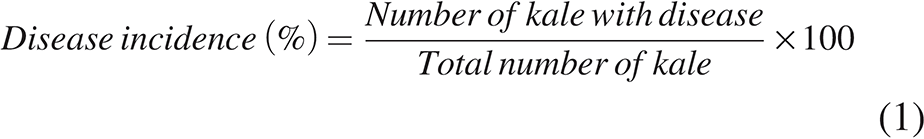
Wilting stages of kale
The wilting stages of kale after harvest and during storage at 4°C for 1, 3, 5, and 7 days were recorded using the wilting stage scale described by Fields et al. (2014). Ten kale plants were sampled and evaluated by comparison to the scale and scored as stage 1 (initial wilt), stage 2 (moderate wilt), or stage 3 (heavy wilt).
Gas chromatography–mass spectrometry analysis
Kale was extracted into a solution, and the plai oil components present in the kale were analyzed using gas chromatography–mass spectrometry (Agilent 5977A; Agilent Technologies Inc., CA) equipped, following the method described by Promwee and Matan (2025).
Total phenolic content, total flavonoid content, and antioxidant activity
The total phenolic content (mg gallic acid equivalents [GAE]/100 g) and total flavonoid content (mg catechin equivalents [CE]/100 g) in kale were analyzed using the method of Matan and Kongchoosi (2024). In addition, the scavenging capacities of 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) and 2,2-diphenyl-1-picrylhydrazyl (DPPH) radicals were evaluated according to Matan and Kongchoosi (2024) and reported as percentages (%).
Mechanism of action of treated kale on L. monocytogenes
Kale treated with 10 μL/mL of plai oil and untreated control kale were selected for this study. L. monocytogenes was inoculated onto the kale surface, and analyses were conducted following the method of Matan and Khunjan (2025). For protein and nucleic acid leakage, intracellular electrolyte leakage, and pH analysis, the L. monocytogenes suspensions were centrifuged at 4000 × g for 5 min at 4°C, and the supernatants were filtered through a 0.22-μm membrane. Absorbance at 260 and 280 nm, conductivity, and pH were measured at 25°C. Biofilm formation was assessed by staining with crystal violet, eluting with methanol, and measuring the absorbance at 600 nm using a spectrophotometer. These parameters were assessed at 1, 6, 12, 18, and 24 h. The experiment was performed in triplicate with three replications per treatment.
Surface morphology was examined using a scanning electron microscope (SEM; Zeiss Merlin Compact SEM; Carl Zeiss Microscopy GmbH, Germany).
Microbiological quality of kale during storage
The microbiological quality of kale stored at 4°C for 7 days was evaluated by analyzing total mesophilic and psychrophilic bacteria, yeasts and molds, coliforms, and lactic acid bacteria. A 25 g sample of kale was homogenized in 225 mL of 0.1% peptone water and serially diluted (10−1 to 10−7). One milliliter from each dilution was plated on PCA for mesophilic bacteria (incubated at 35 ± 2°C for 48 h; Binder, Germany) and psychrophilic bacteria (7 ± 2°C for 10 days); MEA for yeasts and molds (25 ± 2°C for 3 days); Compact Dry EC for Escherichia coli; and Compact Dry LM for L. monocytogenes (35 ± 2°C for 1 day). Microbial counts were reported as log10 CFU/g. Each measurement was conducted in five replicates (n = 5).
Statistical analysis
All data are expressed as mean ± standard deviation. Statistical analysis for the survival of L. monocytogenes, wilting scale, and microbial quality during storage was carried out using one-way analysis of variance, followed by Duncan’s post-hoc test to determine statistically significant differences (p < 0.05). Data on the mechanism of action, bioactive compounds, and antioxidants were evaluated using an independent t-test. Statistical software (StatSoft, OK) was used for all analyses.
Results
Figure 2A illustrates the survival of L. monocytogenes on kale. Treatment with plai oil at concentrations of 15–20 μL/mL completely inhibited the growth of L. monocytogenes within 24 h. A concentration of 10 μL/mL significantly reduced the bacterial population by 5 log10 CFU/g compared with the control. Antimicrobial activity was detected as early as 3 h postinoculation and became clear after 9 h. As shown in Figure 2B, disease incidence in the control group exceeded 20% by day 3 and reached 63% by day 10 of storage. In contrast, kale treated with 10 and 15 μL/mL of plai oil exhibited only 10% disease incidence, while no disease symptoms were observed at 20 μL/mL. However, the wilting scale results (Fig. 2C) indicated that plai oil at 15 and 20 μL/mL caused moderate wilting from day 1, progressing to severe wilting (stage 3) by day 3. In the control group, moderate wilting appeared on day 3, with severe wilting occurring by day 10. Notably, kale treated with 10 μL/mL of plai oil exhibited only mild wilting throughout the 7 days in storage. Collectively, treatment with 10 μL/mL of plai oil was found to be the most effective concentration for inhibiting L. monocytogenes, reducing disease incidence, and minimizing wilting in kale during cold storage at 4°C for 7 days.
Survival of Listeria monocytogenes
Following the introduction of plai oil into the hydroponic system and subsequent harvesting, components of the plai oil were absorbed into the kale leaves. At a plai oil concentration of 10 μL/mL, the leaf tissue was found to contain thujene (35%), terpinene (21%), pinene (15%), and terpinene-4-ol (14%) (Fig. 1). These results corresponded with elevated levels of total phenolic content (956 mg GAE/100 g) and total flavonoids (746 mg CE/100 g) in the treated kale, which were higher than those in the control group (823 mg GAE/100 g and 664 mg CE/100 g, respectively). Additionally, antioxidant activity, as measured by ABTS and DPPH scavenging assays, was significantly enhanced in the treated kale, with values of 47.14% and 49.6%, respectively, compared with 25.7% and 32.5% in the control (Table 1).
Bioactive Compounds and Antioxidant Activity of Control and Plai Oil-Treated Kale
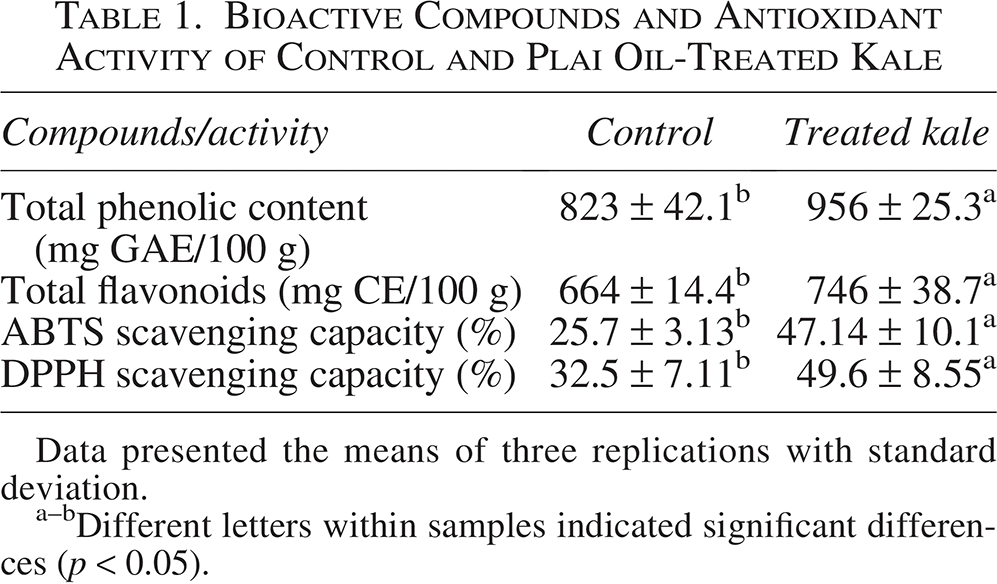
Data presented the means of three replications with standard deviation.
Different letters within samples indicated significant differences (p < 0.05).
The mechanisms involved in the inhibition of L. monocytogenes on kale with and without plai oil are shown in Figure 3. The pH of the test tubes containing L. monocytogenes and kale with plai oil was found to decrease starting at 18 h and significantly declined to pH 5.6 at 24 h. In contrast, the pH in the control group remained stable between 6.1 and 6.2 throughout the 24 h. This trend corresponded with the conductivity measurements, where the group containing treated kale exhibited increased membrane leakage of L. monocytogenes, as indicated by a conductivity value of 241 μS/cm. Protein release was detected with an optical density (OD)280 of 0.055, and amino acid release was measured with an OD260 of 0.049, both higher than the values observed in the control group (conductivity ∼ 215 μS/cm, OD280 ∼ 0.049, OD260 ∼ 0.033). Biofilm formation was also affected; L. monocytogenes in tubes with treated kale exhibited reduced biofilm production, with an OD600 of 0.079, which was notably lower than the OD600 of 0.092 observed in the control. The morphology of L. monocytogenes was found to be consistent with the results shown in Figure 3. Cells on the surface of kale in the control group (Fig. 4A) appeared as regular, bacillus-shaped cells with smooth surfaces. In contrast, L. monocytogenes in the treated kale group (Fig. 4B) exhibited deformed, shrunken structures with clearly wrinkled surfaces.
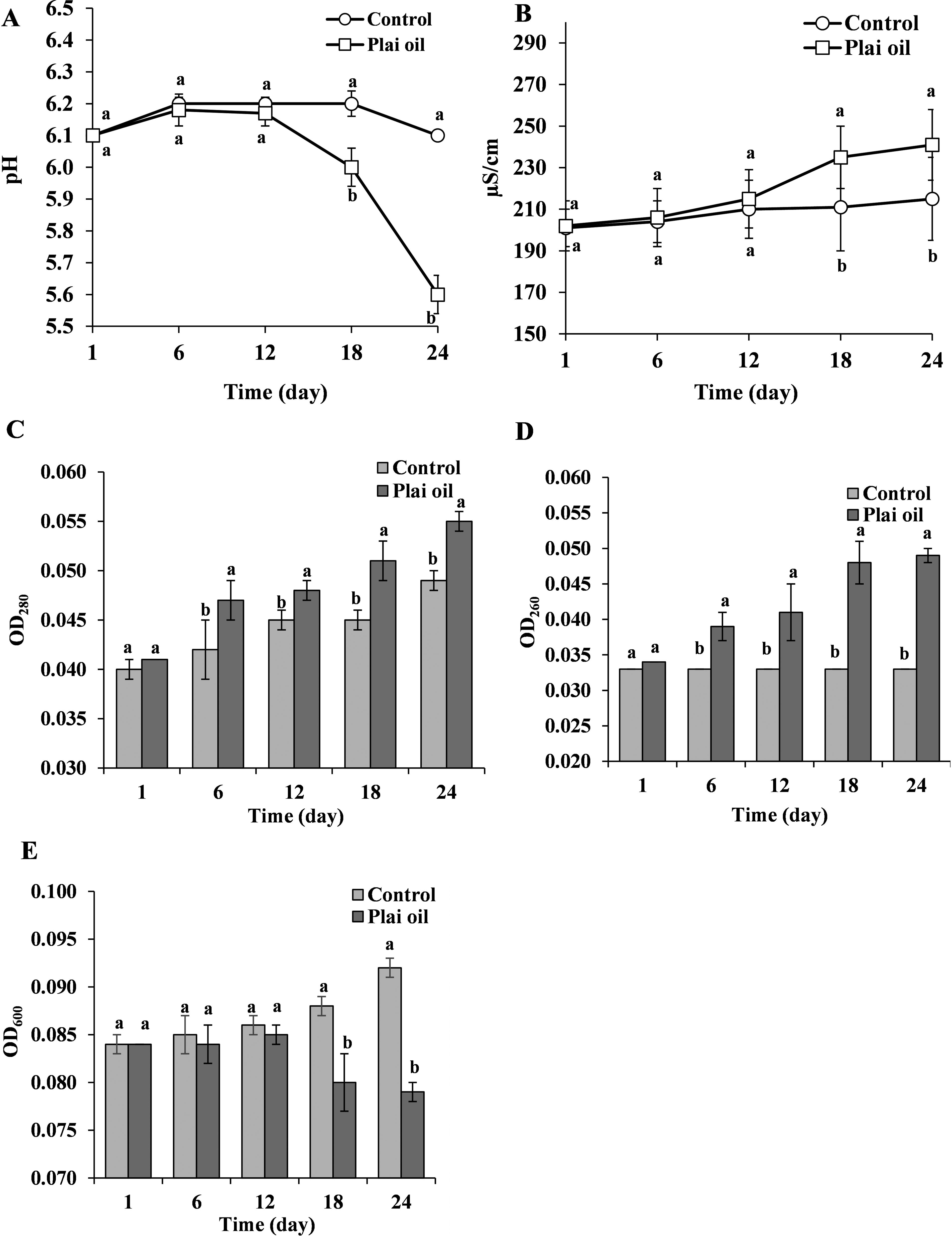
Changes in pH
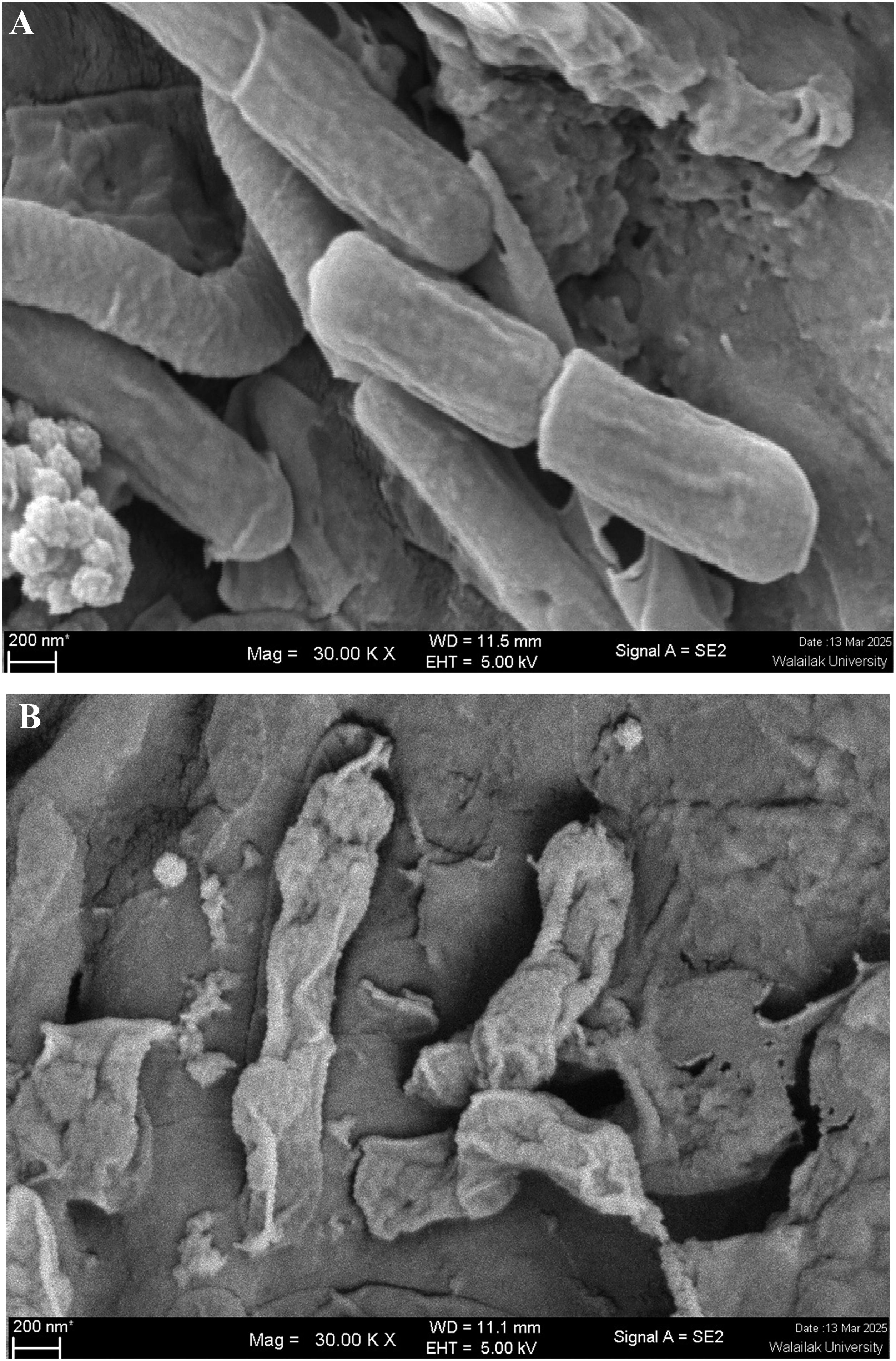
Morphology of Listeria monocytogenes on control kale
Spoilage-related microbial counts in fresh kale during storage were assessed, including mesophilic bacteria, psychrophilic bacteria, yeasts/molds, coliforms, and lactic acid bacteria (Table 2). At 7 days, kale treated with 10 μL/mL plai oil exhibited significantly lower microbial loads than the control (p < 0.05). Reductions of ∼3–4 log10 CFU/g were observed for mesophilic and psychrophilic bacteria, and reductions of ∼1.5 log10 CFU/g were recorded for yeasts/molds. E. coil and L. monocytogenes, which are naturally occurring and considered beneficial, were completely inhibited in the treated group, compared with ∼3.7–3.8 log10 CFU/g in the control.
Changes in Microbiological Quality of Control and Treated Kale During Storage at 4°C for 7 Days

Data presented the means of five replications with standard deviation.
Different letters within samples indicated significant differences (p < 0.05).
Different letters indicate statistically significant differences (p < 0.05) within the samples stored for different durations.
Discussion
A novel preharvest method to enhance the antimicrobial function of kale was confirmed by introducing 10 μL/mL plai oil into a hydroponic kale system, with the roots exposed to the solution 12 h before harvest. The absorbed oil was systemically transported to the leaves, which subsequently exhibited enhanced antibacterial properties. Key bioactive compounds in plai oil, including thujene, terpinene, pinene, and terpinene-4-ol, were detected in the treated leaves. These compounds were associated with antibacterial activity and contributed to increased levels of total phenolics and flavonoids. As a result, the antioxidant activity of the kale was significantly enhanced. These findings were consistent with Promwee and Matan (2025), who reported that preharvest application of citrus oil at 0.006% in chili plants reduced both physical injury and disease incidence by 50% after harvest. High levels of bioactive compounds and antioxidant activity were also observed in the harvested chili.
Kale leaves treated using this approach effectively suppressed L. monocytogenes. When surface contamination occurred, the number of viable L. monocytogenes cells on treated kale gradually declined. In samples with initial contamination levels below 5 log10 CFU/g, bacterial growth was completely inhibited. Additionally, no black spots or discoloration appeared on the surface of the leaf for more than 7 days, helping to maintain visual quality and reduce the likelihood of consumer rejection. The concentration of plai oil absorbed through the roots was found to be critical for preserving postharvest leaf turgor. Concentrations above 15 μL/mL resulted in accelerated wilting, emphasizing the importance of an optimal dosage. In this study, 10 μL/mL was identified as the most effective concentration. At lower levels, such as 5 μL/mL, no antimicrobial effect against L. monocytogenes was observed. These findings support earlier studies suggesting that essential oils can inhibit microbial growth even at low concentrations. However, slight increases beyond the optimal range can negatively affect plant tissues, leading to discoloration or undesirable texture (Chaidech and Matan, 2023). The use of low concentrations also provided cost advantages and minimized sensory changes in the treated vegetables.
The mechanism of action was clearly demonstrated. Once absorbed into kale leaves, plai oil disrupted the cell membranes of L. monocytogenes, leading to leakage of intracellular proteins and amino acids. This disruption was confirmed by increased EC and a reduction in pH of the surrounding medium. Morphological analysis revealed deformed and shrunken bacterial cells, consistent with membrane damage. Furthermore, biofilm formation on the leaf surface was significantly reduced during cold storage, confirming the role of absorbed essential oils in inhibiting biofilm development. The antibacterial activity was mainly attributed to specific components of plai oil found in the kale leaves. Most were identified as terpenoids, including monoterpene alcohols and acrylic terpinene alcohols. These compounds are known to inhibit nucleic acid synthesis and may also help address biofilm-mediated antibiotic resistance (Ghosh et al., 2022). These observations align with the findings of Matan and Matan (2025), who showed that lime oil absorbed by sprouts inhibited both E. coli and L. monocytogenes by damaging cell walls, causing nucleic acid leakage, and suppressing biofilm formation. Their method extended the shelf life of ready-to-eat sprouts by at least 5 days, compared with just one day for untreated control. Similarly, Matan and Kongchoosi (2024) confirmed that plai oil played a critical role in producing pathogen-free, high-quality broccoli.
Preharvest application of plai oil in hydroponic systems offers substantial benefits for vegetable producers. In addition to enhancing the antimicrobial properties of kale, this method extended its postharvest shelf life. Microbiological analysis showed that kale stored at 4°C remained within acceptable safety limits for ready-to-eat vegetables for at least 7 days. In contrast, control samples exceeded recommended thresholds within 3 days, reaching approximately 7 log10 CFU/g for aerobic plate count and over 2 log10 CFU/g for E. coli and Listeria (Faour-Klingbeil et al., 2016). These findings support the potential of this approach to produce microbiologically safe kale from cultivation through to consumption.
Conclusions
This experiment discovered a novel method for introducing low-concentration plai oil into a hydroponic kale cultivation system prior to harvest. A concentration of 10 μL/mL of plai oil was found to be optimal for minimizing disease incidence and preventing wilting of kale after harvest. Kale treated with this method exhibited enhanced resistance to L. monocytogenes and other microorganisms. The treated kale maintained its quality for up to 7 days longer than the control group when stored under cold conditions after harvest. The primary mechanism of action was identified as causing leakage in L. monocytogenes cells, leading to the release of intracellular substances and a reduction in biofilm formation.
Statement
During the writing preparation of the article, the authors use ChatGPT to check the grammar. After using this tool/service, the authors reviewed and edited the content as needed and took) full responsibility for the content of the publication.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The present work was supported by Walailak University (Contract No. WU-COE-68-06), Nakhon Si Thammarat, Thailand.