
Abstract
Vibrio parahaemolyticus is a common foodborne pathogen with drug-resistant strains now widespread. The use of phages to control resistant bacteria is a promising means of reducing public health pressure. In this study, V. parahaemolyticus phage 460s1 was isolated, which was active over a wide range of temperature (30–50°C) and pH (3–10). It exhibited a specific host range, lysing 16 of the 23 strains of V. parahaemolyticus. The phage 460s1 can effectively inhibit the growth of V. parahaemolyticus in shrimp at 15°C and 25°C, and bacterial count decreased by 3.46 log colony-forming unit/g at multiplicity of infection of 1000 at 24 h under 15°C. The phage 460s1 genome was sequenced, and it contains 105 open reading frames (ORFs), of which ORF 53 encodes putative endolysin. The ORF 53 protein was successfully expressed in Escherichia coli and reduced the optical density at 600 nm of host bacteria from 0.82 to 0.58 within 60 min at a concentration of 10 μmol/L. The endolysin also showed lytic activity against 20 strains of antimicrobial-resistant V. parahaemolyticus. Therefore, bacteriophage 460s1 and its endolysin are of great significance for the prevention and control of V. parahaemolyticus.
Introduction
With economic development, the aquaculture industry is also increasingly prosperous. Therefore, the frequency and scope of bacterial diseases of aquaculture organisms are gradually increasing, and the diseases caused by Vibrio parahaemolyticus in seafood are serious (Li et al., 2018). Many cases and studies have confirmed that V. parahaemolyticus is the bacterium that causes lesions and even mass deaths in different species of shellfish, including abalone, clams, and oysters (Zhang et al., 2018). In addition, V. parahaemolyticus is considered to be the main pathogen of acute gastroenteritis in humans caused by the consumption of raw or undercooked seafood.
Antimicrobials have been widely used in the control of bacterial diseases since 1950 (Aarestrup and Wegener, 1999). Antimicrobials and disinfectants are nowadays mostly used in health care and food industry chain to prevent and control contamination by pathogenic bacteria (Christaki et al., 2020). However, the prolonged and excessive use of antimicrobials led to the development of bacterial resistance, and the resistance of V. parahaemolyticus to antibiotics has also gradually increased, which has the potential to cause serious public health and economic problems (Sienkiewicz et al., 2014).
The use of phages for microbial control and rapid detection has become a popular research nowadays (Tan et al., 2016). Endolysins are peptidoglycan-degrading enzymes encoded by phages and are produced within the infected host bacteria at the end of the proliferation cycle (Kashani et al., 2018). It disrupted the osmotic pressure balance of the bacteria, leading to bacterial death, and new phages were released from the dead bacterial cell (Oliveira et al., 2016). The use of endolysins in food is considered safer than bacteriophages as they do not create gene transfer problems and do not lead to the emergence of antimicrobial-resistant bacteria. Because of their specialized lysing properties, their application in the prevention and control of antimicrobial-resistant bacteria, and so on showed a greater prospect of development.
In this study, the biological characteristics of V. parahaemolyticus phage were determined, and the effect of phage on the control of V. parahaemolyticus in food was investigated. The whole genome was analyzed, and endolysin was expressed in Escherichia coli, as well as the lytic ability of endolysin was explored. This study provided a certain theoretical basis for V. parahaemolyticus phage and its endolysin in controlling V. parahaemolyticus contamination.
Materials and Methods
Bacterial growth and phage isolation
Phages were isolated from the sewage of retail markets in Wuhan, Hubei, China, with V. parahaemolyticus 460 as the host bacterium. In brief, the samples were filtered, centrifuged at 7104 g for 10 min, and the supernatant was filtered with a 0.22 μm filter membrane (Millipore). The phage filtrate was added to Luria Broth (LB) medium (Oxoid) containing logarithmic phase host bacteria and incubated for 12–18 h at 37°C. The above steps were repeated at least 2–3 times for double-layer plate assay. The purification of phage was completed when phage spots of uniform shape, size, and transparency were present on the plate. The purified phage was named 460s1.
Transmission electron microscopy of phages
The method was experimented with reference to Gradaschi et al. (2021). Appropriate amount of purified phage suspension solution was dripped onto the copper mesh and adsorbed naturally for 5–10 min and then stained on the copper mesh with 20 µL of 2% (vol/vol) phosphotungstic acid for 10 min, dried at room temperature, and photographed under transmission electron microscopy (TEM) observation.
Host spectrum
Referring to Marquioni et al. (2022) who studied the host profile of V. parahaemolyticus phage, V. parahaemolyticus and other strains were plate streaked, and single colonies were picked for liquid culture to obtain bacterial suspension. The obtained bacterial suspension and phage were mixed and plated. The host extent was determined by the presence or absence of visible plaques.
Optimal multiplicity of infection and lytic activity determination
The method of Yu et al. (2013) and Sharma et al. (2021) was referred and modified for this experiment: V. parahaemolyticus strain 460 was cultured to logarithmic phase, diluted to 107 colony-forming unit (CFU)/mL, and then mixed with phage at different multiplicity of infection (MOI) of 1000, 100, 10, 1, 0.1, 0.01, and 0.001. The mixture was incubated at 37°C for 3.5 h, and the phage potency was determined by double-layer plate method. The optimal MOI was the MOI value corresponding to the highest peak in the titer of the phage.
About 100 µL of V. parahaemolyticus 460 overnight culture was added to 200 µL of phage in 96-well plates (Corning) at MOIs of 1000, 100, 10, 1, 0.1, 0.01, and 0.001 and incubated at 37°C. LB medium and host bacteria alone were used as controls. Optical density at 600 nm (OD600) was measured at 1 h intervals for 24 h to determine the lytic activity of phage.
Optimal adsorption rate and one-step growth curve
The methodology was referred to Evseev et al. (2020) with minor modifications to determine optimal adsorption rate: The phage 460s1 suspension was incubated with host bacteria at the optimal MOI for 30 min at 37°C. Samples were taken every 2 min and centrifuged at 7104 g for 1 min, and the supernatant was used for titer measurement.
Phage 460s1 suspension was mixed with host bacteria at the optimal MOI and incubated at the optimal adsorption rate. After adsorption was completed, the mixture was centrifuged, and the precipitate was resuspended using LB medium. The suspension was incubated at 37°C for 2 h. Samples were taken at 10-min intervals to determine the phage titers.
Thermal and pH stability tests
Thermal stability of phage 460s1 was determined by incubation at 30–70°C, with samples taken at 0, 30, and 60 min. pH determinations were referenced to Thung et al. (2017) with modifications: Phage stability was determined by incubation with phosphate buffered saline buffer at pH of 2–12 for 2 h, and phage titers were measured.
Phage 460s1 genome analysis
Phage genomic DNA was extracted with reference to Guo et al. (2019), with minor modifications: The sodium dodecyl sulfate (SDS)–proteinase K protocol was applied to extract the genomic DNA of the phage 460s1 from high-titer phage particles. The genome was sequenced using the Illumina HiSeq platform (Illumina), and the raw sequence was assembled using SOAPdenovo2 (Luo et al., 2012). The online tool GeneMark was used to determine its open reading frame (ORF). Functional annotation was performed by NCBI-blast against GeneBank database and UniProt database (https://www.uniprot.org/). The virulence and resistance profiles of phage genomes were assessed by the Pathogenic Virulence Factor database (https://www.mgc.ac.cn/VFs/main.htm) and the CARD database (Yang et al., 2019) (https://card.mcmaster.ca/; http://sourceforge.net/projects/brig). Comparison of V. parahaemolyticus phage genome sequence with its cognate phage was performed using Easyfig (Sullivan et al., 2011). MEGA 7.0 software was used to construct a phylogenetic tree based on phage conserved terminase large subunit sequences (Kumar et al., 2016).
Expression of ORF53 protein
The amplification of ORF53 was performed by polymerase chain reaction (PCR) using a forward primer containing a NdeI restriction endonuclease site and a reverse primer with XhoI restriction endonuclease site. Purified PCR products were digested and ligated into the pET28a expression vector. The recombinant plasmid was transformed into competent E. coli strains. The bacterial solution was incubated to an OD600 value of 0.8, and 1 mM isopropyl β-
Lytic activity and lytic spectrum of ORF53 protein
Referring to the study of Vaara (1992), chloroform (0.5%) was added to an overnight bacterial culture and left for 20 min, then centrifuged at 1776 g for 10 min, and the precipitate was washed with sterilized deionized water for more than 3 times. The bacterial sediment was suspended in 50 mmol/L Tris‐HCl buffer containing 0.1% Triton X‐100 (pH 8.2), and the OD600 was adjusted to 0.8–1.0. About 195 µL of host bacterial suspension was mixed with 5 µL of ORF53 protein in a 96-well plate, while the one without protein was used as a negative control. The OD600 value was measured every 5 min for 1 h.
Temperature resistance of endolysin and the effect of metal ions on endolysin
The endolysin was separately subjected to 25, 30–90°C in a water bath for 30 min to determine the changes in the ability to lyse the host bacteria after treatment at different temperatures. The endolysin was supplemented with final concentrations of 0.1, 1, and 10 mmo/L of NaCl, CaCl2, and KCl, respectively, to explore the effects of Na+, Ca2+, and K+ metal ions on the lysing ability of endolysin.
Statistical analysis of data
Statistical analysis was performed using SPSS 22.0 (SPSS Inc.). One‐way analysis of variance was used to analyze the data. p < 0.01 or 0.05 was considered statistically significant.
Results
Morphology and host profile of phage 460s1
The morphology of the phage 460s1 was observed by TEM (Fig. 1). The length of the icosahedral head of phage was 111 ± 3 nm, the width was 60 ± 2 nm, and the tail was 17 ± 1 nm. Therefore, it was shown that the phage belonged to the Caudovirales phage family.
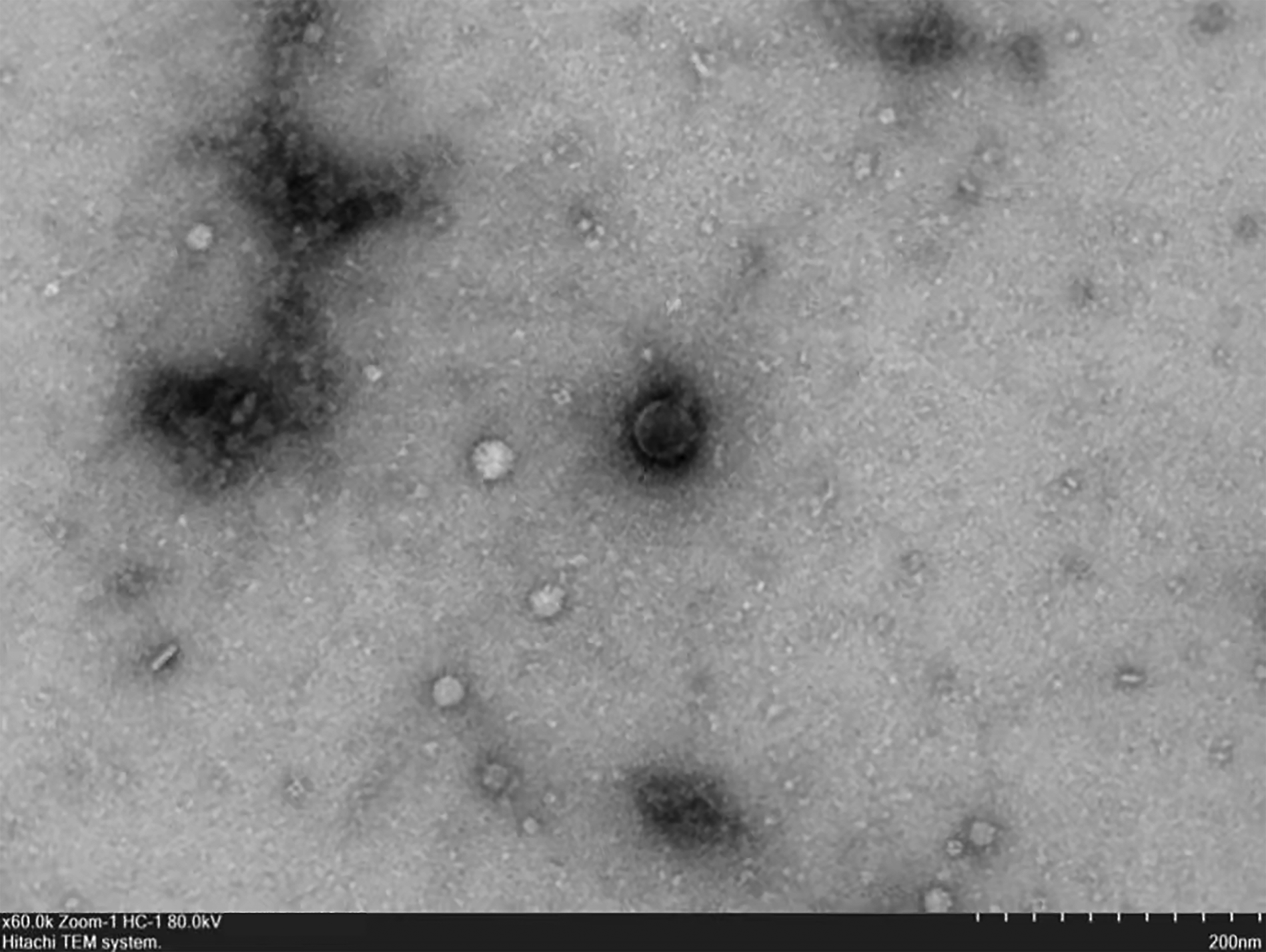
Transmission electron microscopy of phage.
In order to determine the host range of the phage, tests were performed on 23 strains of V. parahaemolyticus, 1 strain of E. coli, 1 strain of Salmonella, and 1 strain of Pseudomonas fluorescens (Table 1). Phage 460s1 had lysis ability against 16 of V. parahaemolyticus strains, which were resistant to at least 3 antimicrobials, including ampicillin, amoxicillin/clavulanic acid, cefuroxime, nalidixic acid, ciprofloxacin and chloramphenicol. No lytic activity was found against E. coli, Salmonella, and P. fluorescens, indicating specific lytic ability for V. parahaemolyticus (Table 1).
Host Profiles of Phage 460s1 and Lytic Spectrum of Lys460s1
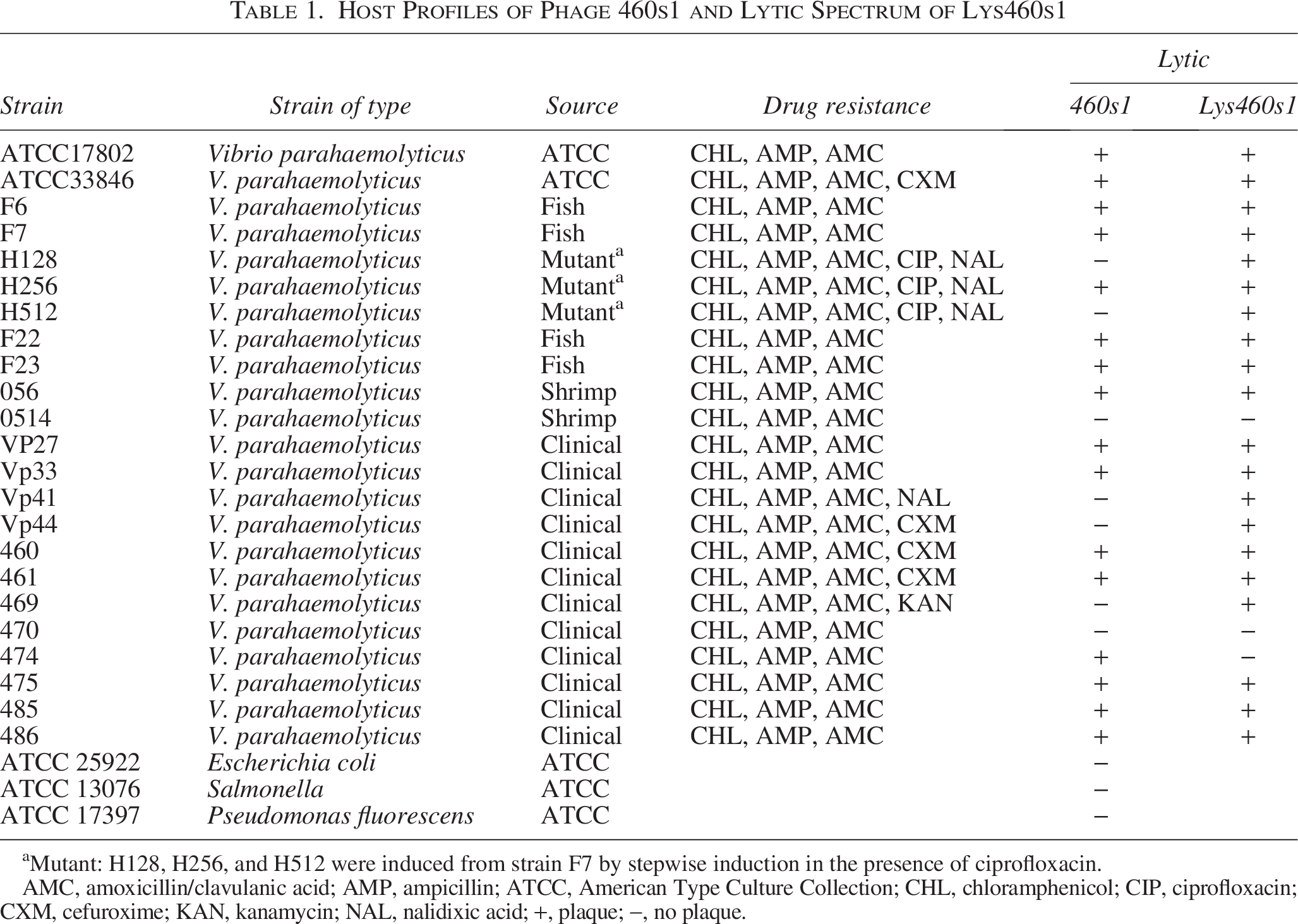
Mutant: H128, H256, and H512 were induced from strain F7 by stepwise induction in the presence of ciprofloxacin.
AMC, amoxicillin/clavulanic acid; AMP, ampicillin; ATCC, American Type Culture Collection; CHL, chloramphenicol; CIP, ciprofloxacin; CXM, cefuroxime; KAN, kanamycin; NAL, nalidixic acid; +, plaque; −, no plaque.
Biological characterization of phage 460s1
The highest titer of phage 460s1 was observed when the MOI was 0.01, that is, the optimal MOI for phage 460s1 (Table 2). Phage 460s1 was incubated with host bacteria with different MOIs to assess the lytic ability of phage 460s1 (Fig. 2A). The inhibition effect of MOI = 1000 and MOI = 100 groups was significantly higher than other groups, especially when the MOI was 1000, the OD600 value was always <0.2 during 3–23 h. Overall, phage 460s1 showed a strong inhibitory effect on V. parahaemolyticus (Fig. 2A). The one-step growth curve of phage 460s1 showed that its latency period was 0–20 min and lysis period was 20–80 min (Fig. 2B).
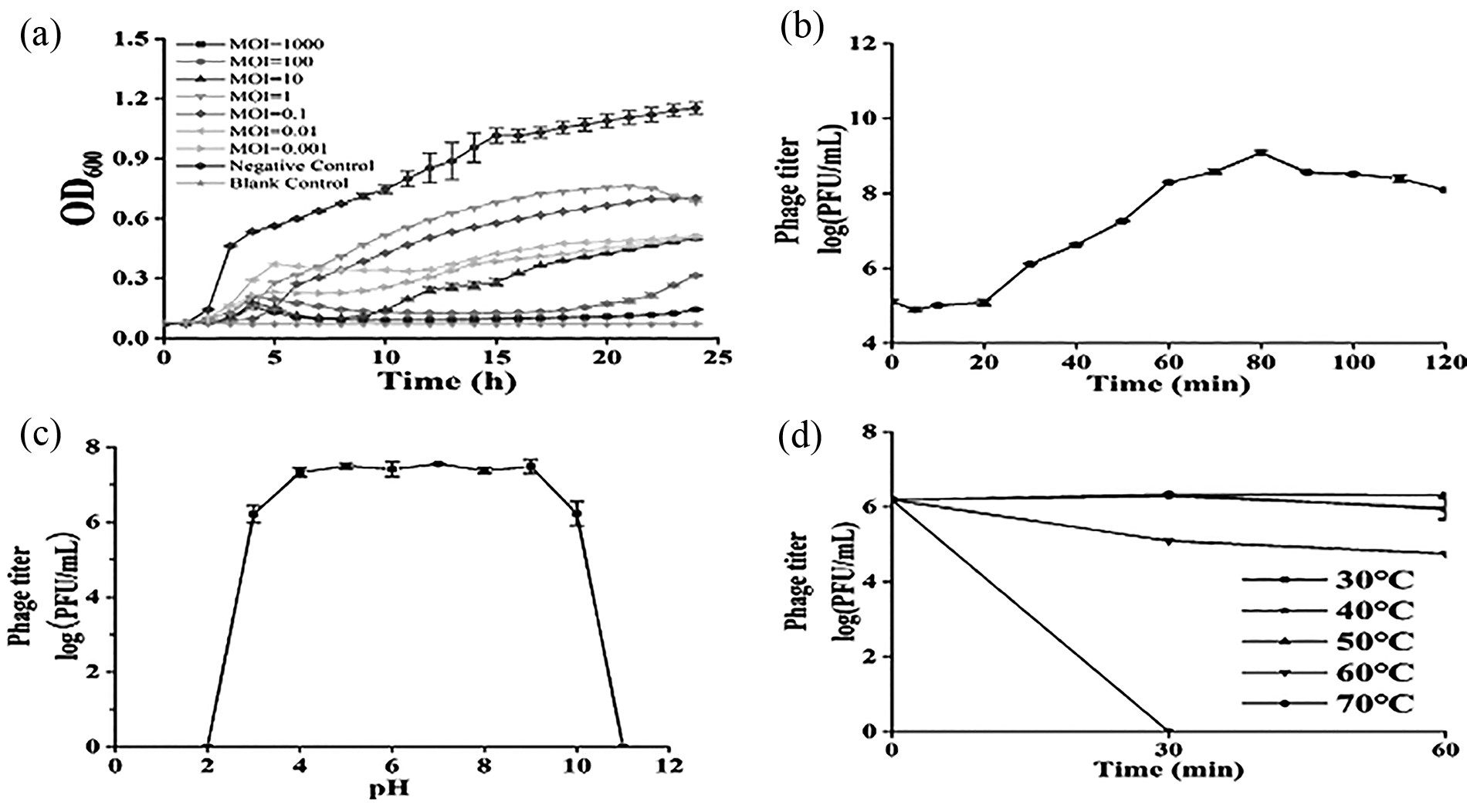
Biological characterization of phage 460s1.
Optimal Multiplicity of Infections for Phage 460s1

CFU, colony-forming unit; MOI, multiplicity of infection; PFU, plaque-forming unit.
The phage 460s1 remained stable at pH 3–10, and titers of phage decreased at pH <3 and >10 (Fig. 2C). After incubation for 60 min at 30–50°C, the phage potency remained basically unchanged; under 60°C, phage titers decreased by 1.44 log plaque-forming unit/mL after 60 min; when the temperature reached 70°C, the phage was not detected after 30 min incubation. The results showed that the phage 460s1 was thermally stable at 30–50°C (Fig. 2D).
Inhibition of V. parahaemolyticus 460 in shrimp meat by phage 460s1
Ability of phage 460s1 to inhibit host bacteria in shrimp was measured at temperature for sale in retail markets (15°C) and under room temperature (25°C) (Fig. 3). Under 15°C, the concentration of V. parahaemolyticus in the experimental group decreased by 1.68 log CFU/g and 1.07 log CFU/g at 6 h and 12 h, respectively, compared with negative control. The bacteriostatic effect of phage was more significant at 24 h, and bacterial count decreased by 3.46 log CFU/g (p < 0.001) (Fig. 3A). Under 25°C, reductions of bacterial count in the experimental group at 6 h, 12 h, and 24 h were 1.00 log CFU/g, 1.67 log CFU/g, and 0.72 log CFU/g, respectively (Fig. 3B).
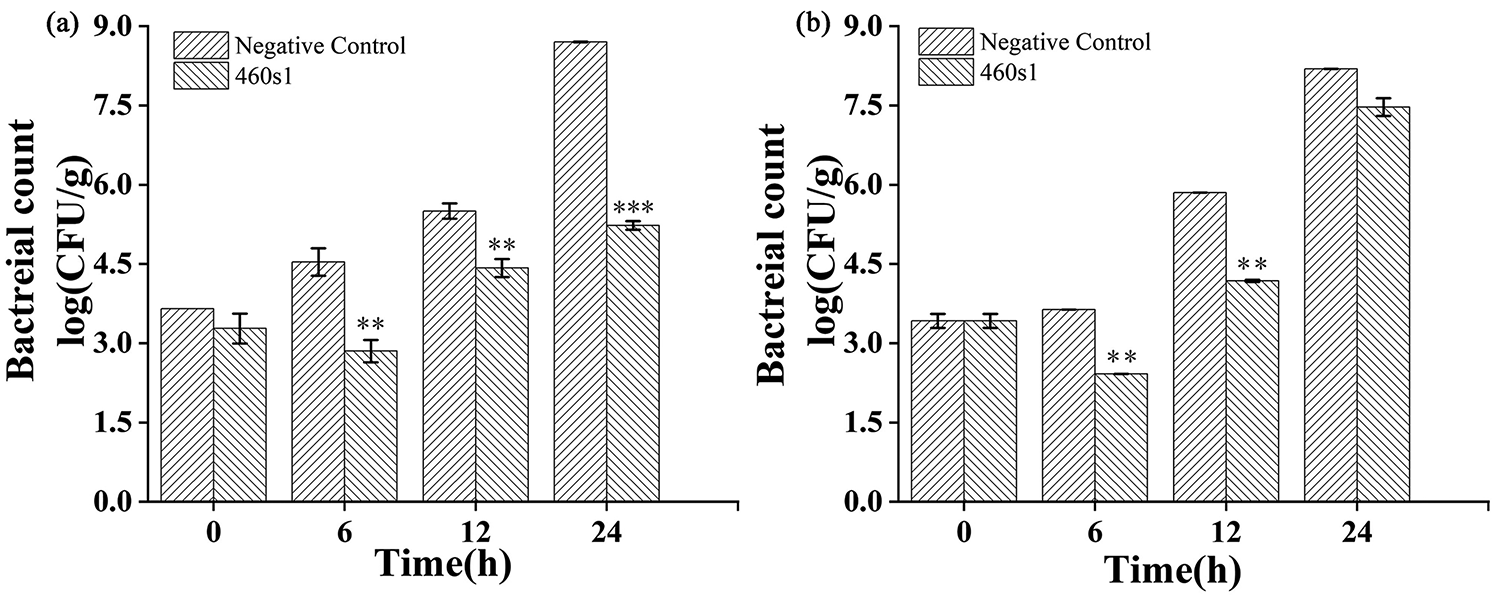
Inhibition of Vibrio parahaemolyticus 460 in shrimp by phage 460s1 at 15°C
Characterization of the genome of phage 460s1
Phage 460s1 is a double-stranded DNA with a length of 78135 bp and a GC content of 49%. Using the RAST tool, the genome was predicted to contain 105 ORFs, with the average length being 739 bp. A total of 34 ORFs were predicted to be functionally known, and the entire genome did not contain virulence and antibiotic resistance genes. They can be categorized into five main groups based on their functions: DNA replication modules (ORF 19, 24, 25, 26, 27, 28, 31, 35, 36, 37, 38, 42, 43, 44, 45, 46, 77, 82, 90), DNA modification modules (ORF 21, 30, 87), structural modules (ORF 1, 48, 55, 59, 63, 68, 72, 73, 74), lysis module (ORF 53), and packaging module (ORF 74, 76). Among the well-defined ORFs, the ORF36 encodes putative thymidylate synthase, which is required for DNA synthesis and repair, and the ORF31 encodes putative thymidylate kinase, which is involved in the regulation of DNA synthesis.
Phage 460s1 was compared with other phage genomes in the GenBank database by BLAST. It was found that phage 460s1 had the highest identity (89.18%) with V. parahaemolyticus phage vB_ VpS_ C2 (MZ592922.1) (Fig. 4). The phylogenetic tree was constructed based on the sequences of phage terminase large subunit using MEGA, and phage 460s1 was observed to form a cluster with V. parahaemolyticus phage R01 and V. parahaemolyticus phage VVP001, indicating close relationships (Fig. 5).
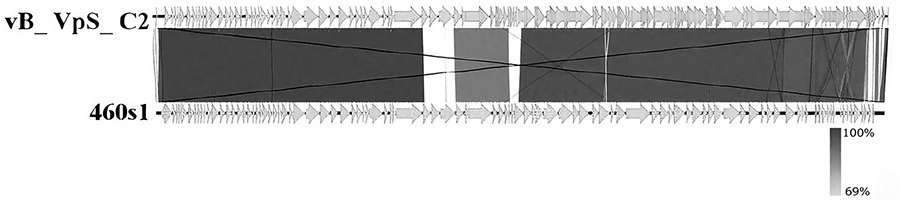
Genome-wide comparative mapping of phage 460s1.

Phylogenetic tree constructed based on terminase large subunit.
Expression and purification of ORF53 protein
The ORF53 was predicted to encode endolysin according to the genome annotation. To verify its lytic activity, ORF53 was cloned into an expression vector, and recombinant plasmids were introduced into E. coli BL21 and T7E, respectively. The expression of ORF53 protein was induced by IPTG at 16°C and 37°C, respectively (Fig. 6), and it can be seen that there was a significant difference in the induction effect under the two temperatures, and the appropriate temperature for the expression of the target protein was 37°C.
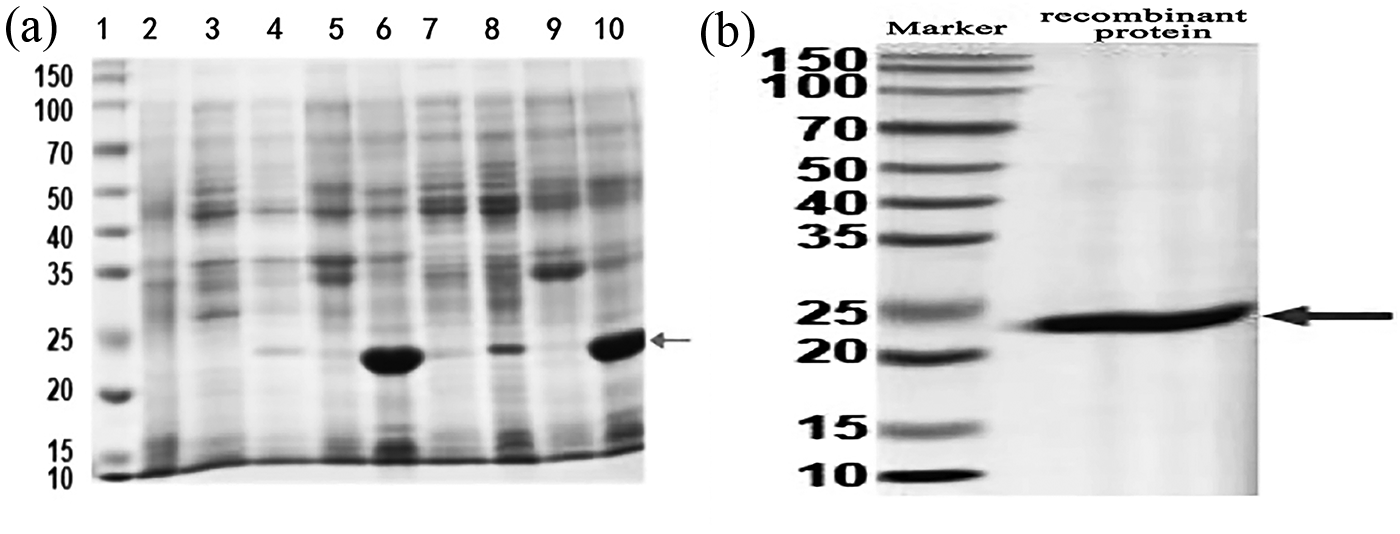
Expression and purification of ORF53 protein.
The protein in the inclusion body was renatured, denatured, and finally purified. The gel electrophoresis of purified protein showed that the protein was around 22 KDa, which was the same size as that of the target protein after the addition of His tag, indicating that the target protein was successfully obtained.
Lytic activity and lytic spectrum of Lys460s1
To determine the lytic activity of ORF53 protein, V. parahaemolyticus during the exponential growth (pretreated with chloroform) was incubated with different concentrations of ORF53 protein, and the turbidity was observed. It was found that the turbidity of V. parahaemolyticus decreased significantly, and the OD600 decreased from the initial value of 0.82 to 0.58 within 60 min when the concentration of ORF53 protein was 10 µmol/L (Fig. 7). Thus, ORF53 protein was considered as endolysin, which was named as Lys460s1.
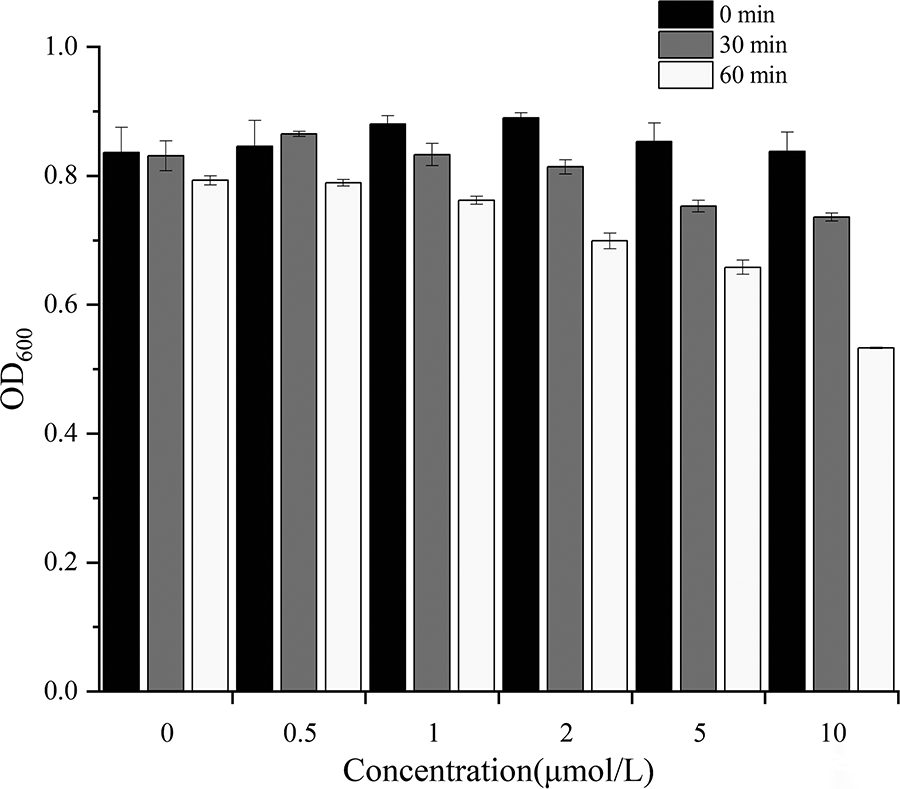
Lytic capacity determination of ORF53 protein. ORF, open reading frame.
To estimate the lytic spectrum of Lys460s1, the same tested strains, including 23 strains of V. parahaemolyticus, 1 strain of E. coli, 1 strain of Salmonella, and 1 strain of P. fluorescens, were used, and the results are shown in Table 2. The endolysin Lys460s1 was active against 20 out of 23 V. parahaemolyticus strains.
Environmental resistance of endolysin Lys460s1
To study its thermal stability, Lys460s1 was incubated from 30°C to 90°C, and its lytic ability was tested. The activity of Lys460s1 decreased with increasing temperature in the range of 30–60°C. At 60°C, the activity of endolysin decreased by 45.5% (Fig. 8A).
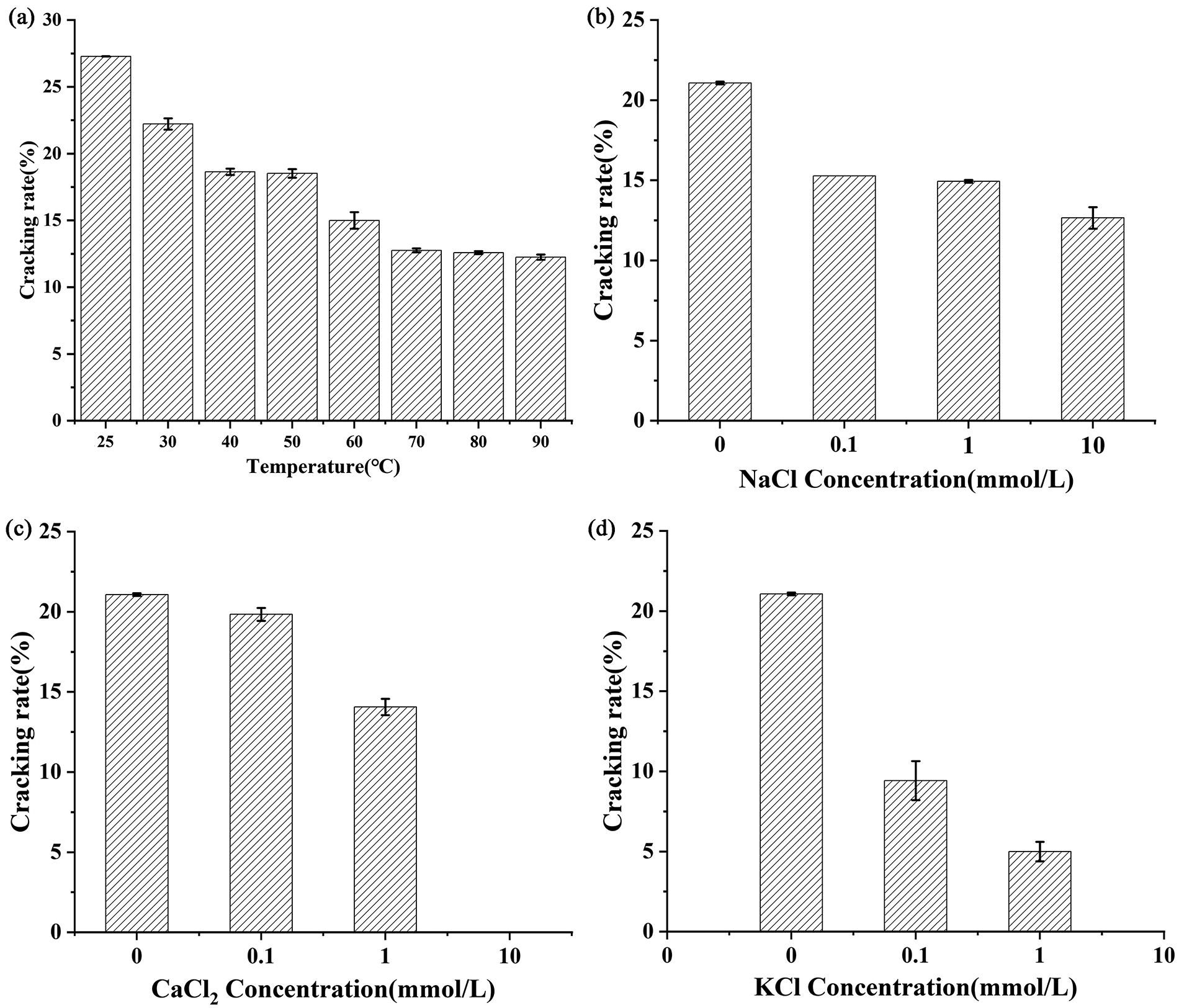
Environmental resistance of endolysin Lys460s1.
The activity of Lys460s1 were significantly affected by concentration of CaCl2 and KCl 10 mmol/L NaCl reduced the lytic activity of Lys460s1 by nearly 10% (Fig. 8B). Low concentration of CaCl2 (0.1 mmol/L) reduced its activity to 19.84%, while high concentration of CaCl2 (10 mmol/L) caused a complete loss of its enzyme activity (Fig. 8C).
Discussion
In this study, V. parahaemolyticus 460 was used as the host bacterium for the isolation of phage 460s1. Host range is a key determinant of how phages affect microbial diversity in the natural environment, including within the microbiome (Duarte et al., 2018). Phage 460s1 lysed 16 out of 23 antimicrobial-resistant V. parahaemolyticus strains, suggesting that 460s1 has a wide host spectrum and has the potential to prevent and control food contamination caused by antimicrobial-resistant V. parahaemolyticus. In addition, no lytic activity was found against E. coli, Salmonella, and P. fluorescens, suggesting that the phage specifically lysed V. parahaemolyticus. This narrow specificity may be an intrinsic feature of phages and a result of the long-term evolution of phage–bacteria interactions in nature and may help to strengthen the relationship between the two (Antonovics et al., 2013; Koskella and Meaden, 2013). Due to this specificity, phages can be used as precise antimicrobial agents.
It has been shown that phages achieved rapid population growth by infecting bacteria and replicating themselves, so their titers increased accordingly with increasing bacterial concentration (Arisaka et al., 2003; Lopes et al., 2018). Despite this, high MOI was not necessarily more favorable for phage replication. This was because at high MOI, bacteria may be inactivated “externally lysed” by simultaneous attack of multiple phages before the phage completed its replication cycle, and therefore the phage replication process may not be fully carried out, resulting in overall phage production not being as high as expected (Zhang et al., 2018; Larpin et al., 2018). Similarly, this also explained why the potency of phage 460s1 did not always increase when the MOI was increased from 0.001 to 1000 in this study. On the contrary, phage 460s1 reached the highest potency at an MOI of 0.01.
The lysis curves demonstrated the phage’s ability to lyse host bacteria. Liu et al. (2024) found that the number of bacteria increased after 11 h when bacteriophage VP41s was used for bacteriostatic assay (MOI = 1000). After the addition of phage LPST144 (MOI = 1000), the host bacteria still began to enter a stable period after 9 h (Yang et al., 2020). In this study, the bacteriostatic effect of MOI = 1000 group was much better than the other groups, and the OD600 value was always <0.2 during 3–23 h. It indicated that phage 460s1 has a high lysis efficiency against host bacteria.
Phage exists in various environments, and temperature and pH are important factors affecting their activity. For bacteriophages with high tolerance, their application prospects will be more extensive. In this study, the phage 460s1 was able to maintain high activity at temperatures up to 50°C and pH 3–10. It was better than phage F23s1 (pH tolerance range of 4–10) isolated by Xia et al. (2022). The stability of the phage under acidic and alkaline conditions allowed its application in food matrices with different pH values. The high tolerance of phage 460s1 suggested that it had great promise for use.
The feasibility of applying phage to inhibit V. parahaemolyticus has been reported. Zhang et al. (2018) used phage VPp1 to sterilize V. parahaemolyticus in oysters. The concentration of V. parahaemolyticus decreased by about 0.83 log CFU/g after phage addition at 6 h. Similar results were found in another study (Li et al., 2018). In the present study, the number of V. parahaemolyticus in the experimental group decreased by 1.68 log CFU/g compared with that of the negative control group at 6 h under 15°C, and the reduction of bacterial count at 24 h was 3.46 log CFU/g. This suggested that phage 460s1 was effective in the inhibition of V. parahaemolyticus in the preservation process of seafood.
Our study indicated that the endolysin Lys460s1 lysed 20 strains of 23 antimicrobial‐resistant V. parahaemolyticus and showed a wider lytic spectrum than the phage (16/23). These results are consistent with previous studies. Li et al. (2018) found that the antimicrobial spectrum of the hydrolyzed mixture of LysVPp1 (9/12) was expanded, compared with phage VPp1 (3/12). Oliveira et al. (2016) demonstrated that the endolysin ABgp46 exhibited a broader spectrum than its original phage in the presence of an outer membrane destabilizer. The reason might be that endolysins can hydrolyze some conserved peptidoglycan structures in bacterial cells. Hence, endolysin Lys460s1 was more advantageous than phage in controlling the growth of V. parahaemolyticus.
The thermal stability of phage endolysin depended on each individual phage isolate and represented the ability of endolysin in the application. A previous work showed (Larpin et al., 2018) that Staphylococcus aureus phage endolysin LysH5 was completely inactivated after incubation for 30 min at 63°C or 15 min at 75°C. In this study, Lys460s1 maintained 55.6% of its enzymatic activity at 60°C, indicating that most enzymes were intolerant to high temperatures. The thermal stability of enzymes was related to its structure, such as helix content, amino acid residues, and hydrogen bonding (Liu et al., 2019). The methods for improving thermal stability with enzyme modification included rational design, irrational design, and semi-rational design (Verma and Satyanara, 2012). It is a creative work to modify natural enzymes into industrial enzymes with high thermal stability.
Metal ions can activate or inhibit enzyme activity by interacting with the amino or carboxyl groups of amino acids (de Cassia Pereira Josiani et al., 2017). In the present study, NaCl, CaCl2, and KCl reduced the ability of Lys460s1 to lyse V. parahaemolyticus. It was reported that the endolysin OalV17 isolated from the marine bacterium Vibrio sp. increased its activity in the presence of divalent metal ions such as Cu2+, Zn2+, Mg2+, Ni2+, Ba2+, Mn2+, and Ca2+, whereas its activity was inhibited in the presence of trivalent metal ions such as Al3+ and Fe3+ (Wang et al., 2013). Therefore, the identification of metal ion-binding sites was essential for understanding the functional mechanisms of enzymes and the role of metal ions in enzyme catalysis.
Conclusions
In conclusion, bacteriophage technology is a method of inhibiting bacterial growth without the use of antibiotics. Bacteriophage 460s1 exhibited resistance to common extreme environments, and its endolysin can effectively inhibit the growth of antimicrobial-resistant strains. The bacteriophage and endolysin showed specificity for V. parahaemolyticus control and have the potential to be used as a food additive in seafood.
Authors’ Contributions
Y.L. and T.S.: Methodology, software, and writing—original draft. B.H.: Methodology. M.Z.: Writing—review and editing. H.W.: Conceptualization, writing—review and editing, and supervision. H.W.: Conceptualization and funding acquisition.
Footnotes
Disclosure Statement
All authors have no conflicts of interest to declare.