
Abstract
Probiotics are critical for resisting the colonization of foodborne pathogens in gut. Lactiplantibacillus plantarum ZDY2013 has been proven to have multiple biological activities, including antagonizing pathogens. In this study, we systematically evaluated the protective effects of live and heat-killed L. plantarum ZDY2013 on Salmonella enterica serotype Typhimurium infection and investigated the potential mechanisms using mice infection model, HT-29 epithelial cell model, and RAW264.7 macrophage model. The results showed that live and heat-killed L. plantarum ZDY2013 could effectively reduce the Salmonella load in mice, improve the intestinal barrier by upregulating the expression of tight junction proteins (Zonula occludens protein 1 and Claudin-1) and the number of goblet cells, and alleviate the inflammation by decreasing the level of proinflammatory cytokines (interleukin-1β, tumor necrosis factor-α, interferon gamma and IL-6). In addition, L. plantarum ZDY2013 could significantly reduce the adhesion and invasion of S. Typhimurium to HT-29 epithelial cells, activate macrophages and enhance its bactericidal ability. These results indicated that live and heat-killed L. plantarum ZDY2013 has the potential to be developed as a functional health food against S. Typhimurium infection.
Introduction
Salmonella, a Gram-negative bacterium, is an intracellular pathogen (Li, 2022). Salmonella enterica serotype Typhimurium (S. Typhimurium) is one of the main biological causes of foodborne illness, and the incidence rate ranks first among Salmonella infections (Vieira et al., 2022). Salmonella enterica causes approximately 100 million infections and over 200,000 deaths per year (Wang et al., 2023). Salmonella Typhimurium is a nonspecific zoonotic bacterium (Dos Santos et al., 2019), which is responsible for hospital infection and explosive food poisoning with a high case fatality rate (Wei et al., 2022). The severity of the disease and the increase in antibiotic resistance of S. Typhimurium are also of concern (Johnson et al., 2018). Therefore, effective early protection of S. Typhimurium is serious for controlling its infection and outbreak through the development of new treatment strategies.
Probiotics have received extensive attention due to their various beneficial effects, especially the antagonistic activity against pathogenic infections. Lactiplantibacillus plantarum is one of the extensively explored probiotic species with wide antagonistic effects against pathogens (Li et al., 2023; Youssef et al., 2021). Meanwhile, there are also safety concerns with live microbes, especially in susceptible populations. Evidence has shown that heat-killed bacteria, as the necessary method in the preparation postbiotics (Sun et al., 2023) which defined as “preparation of inanimate microorganisms and/or their components that confers a health benefit on the host” (Vinderola et al., 2022), have probiotic effects, with safety advantages versus live probiotics (Adams, 2010; Piqué et al., 2019). Moreover, increasing studies showed that heat-killed probiotics are able to antagonize pathogens with antimicrobial compounds and by competition with pathogens for adhesion and colonization (Hirano et al., 2017; Noh et al., 2022; Piqué et al., 2019).
Previously, we have isolated L. plantarum ZDY2013 and found that it exhibits multiple biological activities, including regulation of intestinal microbiota, anti-inflammatory activities, antagonism against pathogenic bacteria (Tian et al., 2023; Wan et al., 2021; Zhang et al., 2021; Zhao et al., 2018). In this study, we systematically investigated the potential effects and mechanisms of live and heat-killed L. plantarum ZDY2013 on S. Typhimurium infection, aiming to open a new horizon on the antipathogenic efficacy of live and heat-killed L. plantarum ZDY2013 and their potential role in functional health foods.
Materials and Methods
Bacterial strains and culture conditions
Strain L. plantarum ZDY2013 (CCTCC M2014170), previously isolated from Chinese pickle (Huang et al., 2015), was cultured in de Man Rogosa and Sharpe (MRS) broth (Solarbio, Beijing, China) anaerobically at 37°C. Salmonella Typhimurium ATCC13311 was obtained from the American Type Culture Collection (ATCC) and cultured in Luria–Bertani (LB) broth (Solarbio, Beijing, China) aerobically at 37°C. The preparation procedure of heat-killed bacterial strains refers to our previous study (Tian et al., 2023). For preparation of heat-killed L. plantarum ZDY2013, overnight cultured cells were centrifuged (×2000 g, 10 min, at 4°C), washed twice with phosphate-buffered saline (PBS), adjusted the concentration to 1.0 × 1010 CFU/mL, and incubated at 70°C for 30 min. Thereafter, the treated cells were spread on MRS agar and cultured to verify the absence of viable cells.
Animals and experimental design
B Albino female mice (6 weeks) were acclimated for 2 weeks and randomly divided into 4 groups (n = 6): C (Control), ST (S. Typhimurium), ZDY2013 (L. plantarum ZDY2013), and ZDY2013-HK (heat-killed L. plantarum ZDY2013). The experimental design was shown as in Figure 1A,C group was administered with PBS by oral gavage for 11 days; ST group was infected with 0.2 mL (1.0 × 109 CFU/mL) of S. Typhimurium on the 8th day, and gavage with PBS on the other days; ZDY2013 and ZDY2013-HK groups were given 0.2 mL (1.0 × 1010 CFU/mL) of live or heat-killed L. plantarum ZDY2013 daily, except for the 8th day of infection with 0.2 mL (1.0 × 109 CFU/mL) of S. Typhimurium. The time points for fecal collection and the sample collection were referenced to previous study (Tian et al., 2023) with minor modification. The body weight of mice was recorded daily, and the feces were collected at 6, 12, 24, 48, and 72 h after S. Typhimurium infection. All mice were sacrificed on day 4 after S. Typhimurium infection. The ileum, cecum, colon, mesenteric lymph nodes (MLN), liver, and spleen were collected under sterile conditions. Serum was collected as previously reported (Kim et al., 2014). The organ index of liver and spleen was determined and calculated as: Index of organs = organ weight (g)/body weight (g) × 100. The protocol (no. SYXK (gan) 2021-0004) was approved by the Ethics Committee of Nanchang University, Jiangxi, P. R. China. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
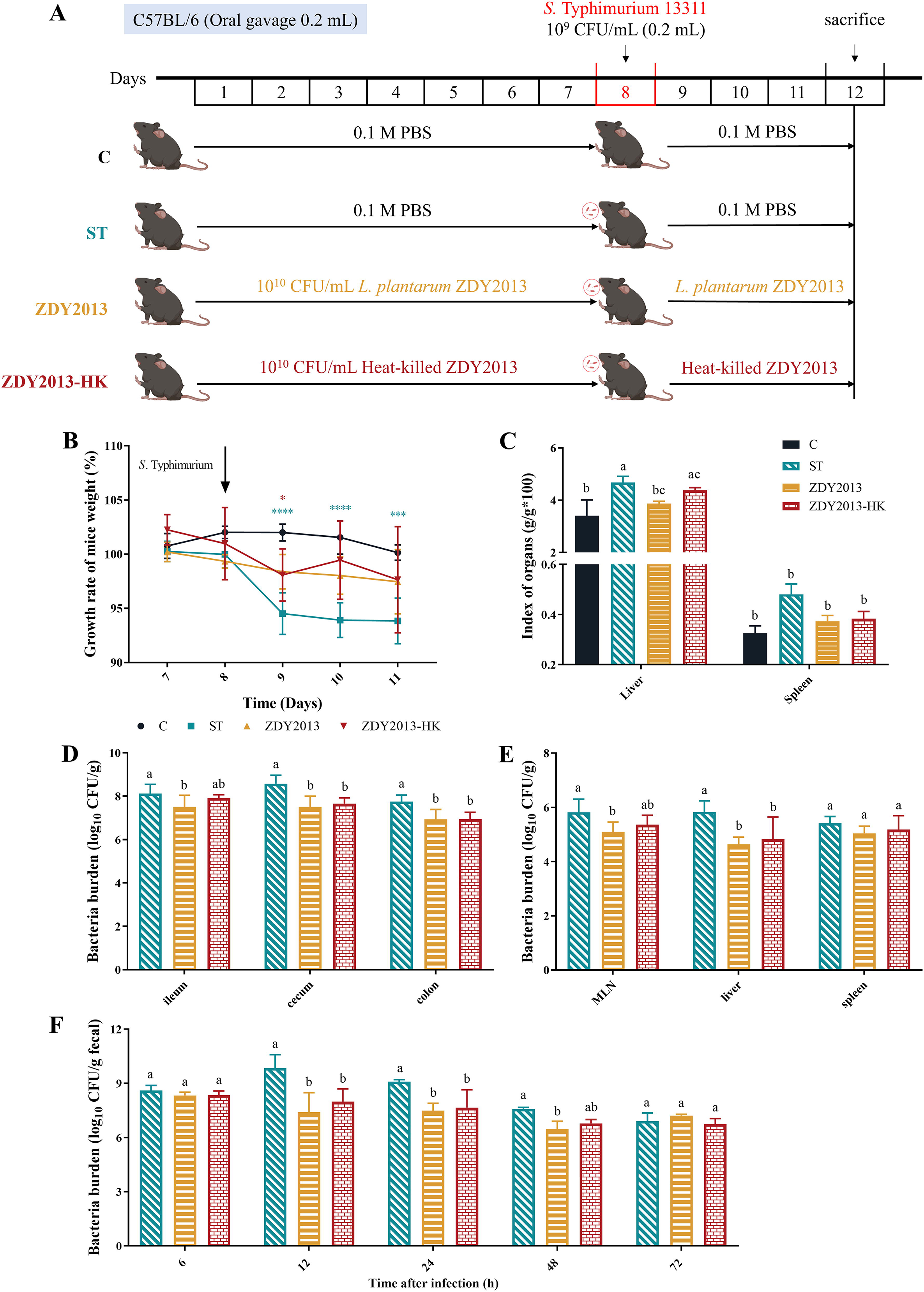
Animal experimental design and effects of live and heat-killed Lactiplantibacillus plantarum ZDY2013 against Salmonella Typhimurium infection. Animal experimental design
S. Typhimurium colonization
The burden of S. Typhimurium in the ileum, cecum, colon, MLN, liver, and spleen tissues and feces were detected using the agar plate method. Briefly, 0.01 g of tissue samples and feces were added to 1 mL sterile 0.01 mM PBS buffer (pH 7.2), followed by fully ground in a tissue grinder. The suspension was serially diluted (10−1, 10−2, 10−3) with 0.1 mL of each dilution and spread on Salmonella chromogenic agar (SCA; Hopebio, Qingdao, China). The CFU value of S. Typhimurium was counted after incubation at 37°C for 24 h according to our previous study (Tian et al., 2023).
Histological analysis
Histological examination and observation were performed as previously reported (Tian et al., 2023). In brief, liver, spleen, ileum, and colon tissues were fixed with 4% (v/v) paraformaldehyde phosphate buffer and embedded in paraffin. Sections were prepared and stained with Hematoxylin and Eosin. In addition, ileum tissues were fixed with Carnoy Fixative, sectioned, and stained with Periodic Acid–Schiff (PAS) to analyze the purple-red dyed goblet cells. Above samples were observed under a light microscope (Nikon Eclipse Ci, Nikon digital sight DS-Fi2; Nikon, Japan).
Intestinal permeability analysis
Intestinal permeability was detected by the fluorescein isothiocyanate dextran (FITC-dextran) method as previously reported (Napolitano et al., 1996; Tian et al., 2023). Briefly, after 4 h fasting, mice were gavage with 4-kDa FITC-dextran (Sigma-Aldrich, USA) at a dose of 60 mg/100 g body weight. After 4 h administration of FITC-dextran, blood samples were collected and centrifuged at 600 g for 5 min to obtain serum. And then the serum was measured under 490 nm excitation and 525 nm emission by fluorescence spectrometer (Thermo Fisher Scientific, Wilmington, USA).
Reverse Transcription Quantitative Polymerase Chain Reaction
The colon tissue was ground in a tissue grinder to analyze the mRNA level of tight junction proteins (Zonula occludens protein 1 [ZO-1] and Claudin-1), cytokines (interferon gamma [IFN-γ], interleukin-1β [IL-1β], IL-10, IL-6, and tumor necrosis factor-α [TNF-α]), and inducible nitric oxidesynthase (iNOS) according to the literature (Tian et al., 2023). Total RNA extraction, cDNA synthesis, and quantitative polymerase chain reaction was performed as previously described (He et al., 2021; Tian et al., 2023). The 2-△△Ct method was used to analyze the relative mRNA levels. The β-actin gene served as a reference gene. The primers used are listed in Supplementary Table S1.
Immunofluorescence assay
Tissue sections (5 µm) of colon were incubated with 3% bovine serum albumin (Servicebio, Wuhan, China) for 30 min to block nonspecific binding. Then sections were incubated overnight at 4°C with the mixed primary antibody, that is, ZO-1 and Claudin-1 (Servicebio, Wuhan, China). Followed by incubation with the corresponding secondary antibody at room temperature for 50 min in dark conditions. Nuclear staining and section observation were performed according to the method previously reported (Tian et al., 2023). Diamidinophenylindole (Servicebio, Wuhan, China) was used for nuclear staining. The slides were visualized and photographed using a Nikon Eclipse C1-U inverted fluorescence microscope equipped with a Nikon DS cooled camera (Nikon, Japan).
Enzyme-Linked Immunosorbent Assay
Serum IFN-γ, IL-1β, IL-10, IL-6, and TNF-α were measured by ELISA kits (Neobioscience, Shenzhen, China) according to the instructions (Tian et al., 2023).
Antiadhesion and invasion assay
The antiadhesion and invasion assay was performed according to previous study (Tian et al., 2023). HT-29 cells in 6-well plates (5 × 105 cells/well) were incubated with live or heat-killed L. plantarum ZDY2013 (109 CFU/well) for 1 h, and then S. Typhimurium (109 CFU/well) was inoculated and incubated for another 1 h. After the nonadherent S. Typhimurium was removed, the adherent S. Typhimurium cells were detached with Ethylenediaminetetraacetic acid–trypsin solution and counted by plating on the SCA plates. For invasion assay, after incubation, gentamicin (100 µg/mL) was added to eliminate the extracellular S. Typhimurium, and then HT-29 cells were lysed with 0.1% Triton X-100. The invasive bacteria were analyzed by serial dilution of the suspension applied to the SCA plates. Adhesion (invasion) rate = A1/A0 × 100%, where A1 is the number of adherent (invasive) S. Typhimurium cells in ZDY2013 or ZDY2013-HK group, A0 is the number of adherent (invasive) S. Typhimurium cells in C group (ST group).
Phagocytosis assay
The phagocytosis assay was conducted following the methodology outlined in our prior research (Tian et al., 2023). RAW264.7 macrophages in 96-well plates (1 × 105 cells/well) were incubated with live or heat-killed L. plantarum ZDY2013 (106 CFU/well) for 1 h, washed twice with Hanks, and then incubated with Dulbecco's modified eagle medium with 10% fetal bovine serum and neutral red solution for another 1 h. The extracellular neutral red was removed by washing, and the cells were lysed with lysis buffer to determine OD540. The phagocytosis rate was calculated as: Phagocytosis rate = A1/A0 × 100%, where A1 is the absorbance of ZDY2013 or ZDY2013-HK group, A0 is the absorbance of C group.
S. Typhimurium survival in RAW264.7 macrophages
The operational procedures for cocultivation and the experimental method for Salmonella counting were cited from our previous study (Tian et al., 2023), except that the cultivation time was modified. In brief, the cultured RAW264.7 was seeded in 24-well plates (1 × 106 cells/well), live or heat-killed L. plantarum ZDY2013 (106 CFU/well) was added and incubated for 1 h. And then S. Typhimurium (106 CFU/well) was added and co-incubated for another 1 h. After washing, 50 μg/mL gentamicin was added and incubated for 1 h to remove the extracellular Salmonella. Thereafter, medium with 5 μg/mL gentamicin was added, which was established as 0 min, and incubated for 30, 60, 90, and 120 min. After washing twice with Hanks, 1% Triton X-100 was added for lysis, and S. Typhimurium were counted by plating the serial suspension dilutions on SCA plates.
Statistical analysis
All data presented are expressed as mean ± standard deviation. Statistical analyses were carried out by GraphPad Prism Version 7.0. All data were analyzed using two-way analysis of variance for multiple comparisons followed by Tukey’s multiple comparison test. The differences were considered statistically significant when the p value was less than 0.05.
Results
Effect of L. plantarum ZDY2013 on the body weight, organ index, and Salmonella typhimurium load of S. Typhimurium infected mice
As shown in Figure 1B, after S. Typhimurium challenge, body weight of mice in the ST group was much less than that of the Control group (p < 0.0001). Administration of live and heat-killed L. plantarum ZDY2013 could alleviate this weight loss, which showed comparable weights to the Control group on the third and fourth day after infection. Meanwhile, liver index in the ST group was significantly (p < 0.05) increased compared with the Control group, which was effectively alleviated by the live L. plantarum ZDY2013 (p < 0.05) (Fig. 1C). However, there was no significant difference in spleen index among all groups.
To investigate the effect of live and heat-killed L. plantarum ZDY2013 administration on the Salmonella Typhimurium loads of infected mice, the viable counts of Salmonella Typhimurium were detected. The results showed that there was a large amount of Salmonella Typhimurium in ileum, cecum, colon, MLN, liver, and spleen of infected mice, with 8.12, 8.57, 7.75, 5.82, 5.83, and 5.41 log10 CFU/g, respectively. Mice pretreated with live and heat-killed L. plantarum ZDY2013 harbored markedly (p < 0.05) lower viable S. Typhimurium loads in intestinal tract, MLN and liver, except that in ileum and MLN of ZDY2013-HK group (Fig. 1D and 1E). Moreover, L. plantarum ZDY2013 could significantly reduce the fecal loads of S. Typhimurium at 12, 24, 48 h, except 72 h postinfection (Fig. 1F), which confirmed that administration of ZDY2013 could reduce the S. Typhimurium burden in the intestinal tract of mice, thereby reducing its amount in feces during early-stage post infection.
Effect of L. plantarum ZDY2013 on liver, spleen, and intestinal barrier of S. Typhimurium infected mice
As shown in Figure 2A and 2B, after S. Typhimurium infection, the liver exhibited disordered arrangement of hepatic cords and congestion of hepatic sinuses, while the spleen showed architectural destruction and blurring of the marginal zone and periarteriolar lymphatic sheath. Following administration of live and heat-killed L. plantarum ZDY2013, the hepatic cords were neatly restored, and hepatic sinus congestion were greatly reduced, especially in the live L. plantarum ZDY2013 group; the liver and spleen damage were also effectively improved.
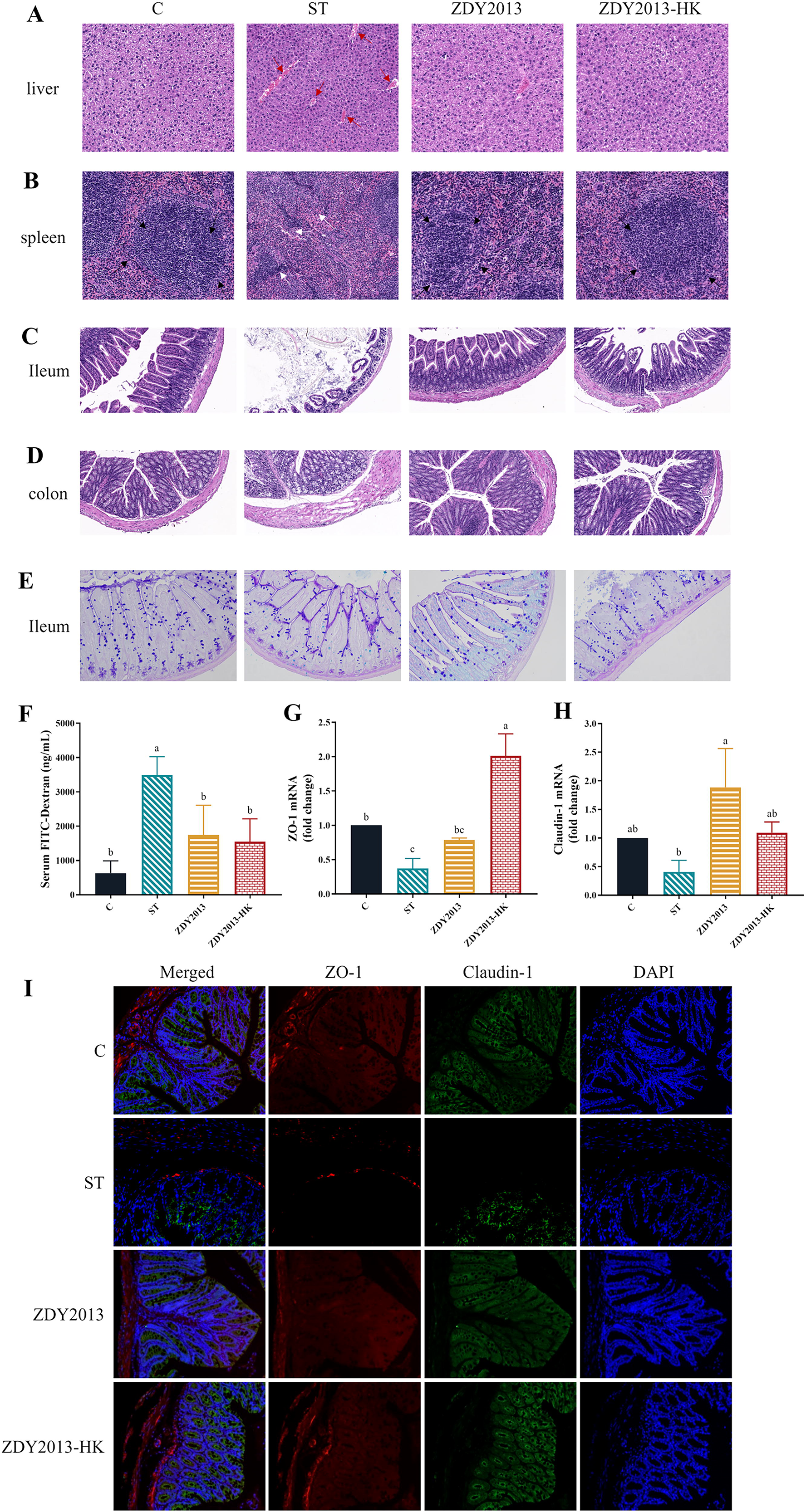
Effects of live and heat-killed Lactiplantibacillus plantarum ZDY2013 on tissue injury and intestinal barrier failure under Salmonella Typhimurium challenge. H&E staining (20×) of liver tissue sections
As shown in Figure 2C and 2E, the morphological analysis of ileum and colon showed that after S. Typhimurium infection, the length of intestinal villi was shortened, distribution was sparse, crypt abscesses and inflammatory infiltration occurred, and the number of goblet cells in ileum tissue decreased. Following administration of live and heat-killed L. plantarum ZDY2013, the overall structure of the small intestine was improved, similar with that of C group (Fig. 2C), and the edema of colonic submucosa was improved (Fig. 2D). The number of goblet cells increased, especially in ZDY2013 group (Fig. 2E). Moreover, after S. Typhimurium infection, the serum FITC-dextran of mice was significantly (p < 0.05) increased, the transcription level (p < 0.05) and expression level of ZO-1 and Claudin-1 were reduced, which were effectively improved by administration of live and heat-killed L. plantarum ZDY2013 (Fig. 2F–I).
Effect of L. plantarum ZDY2013 on the immunity of S. Typhimurium infected mice
As shown in Figure 3A–J, administration of live and heat-killed L. plantarum ZDY2013 could significantly reduce the elevated levels of serum IFN-γ, IL-1β, and IL-6 caused by Salmonella Typhimurium, as well as the elevated transcription levels of colon IFN-γ, IL-1β, IL-6 and TNF-α (p < 0.05), and significantly increase the reduced transcription level of colon IL-10 (p < 0.05). In addition, administration of heat-killed L. plantarum ZDY2013 could significantly enhance the increased transcription level of colon iNOS (p < 0.05) (Fig. 3K).
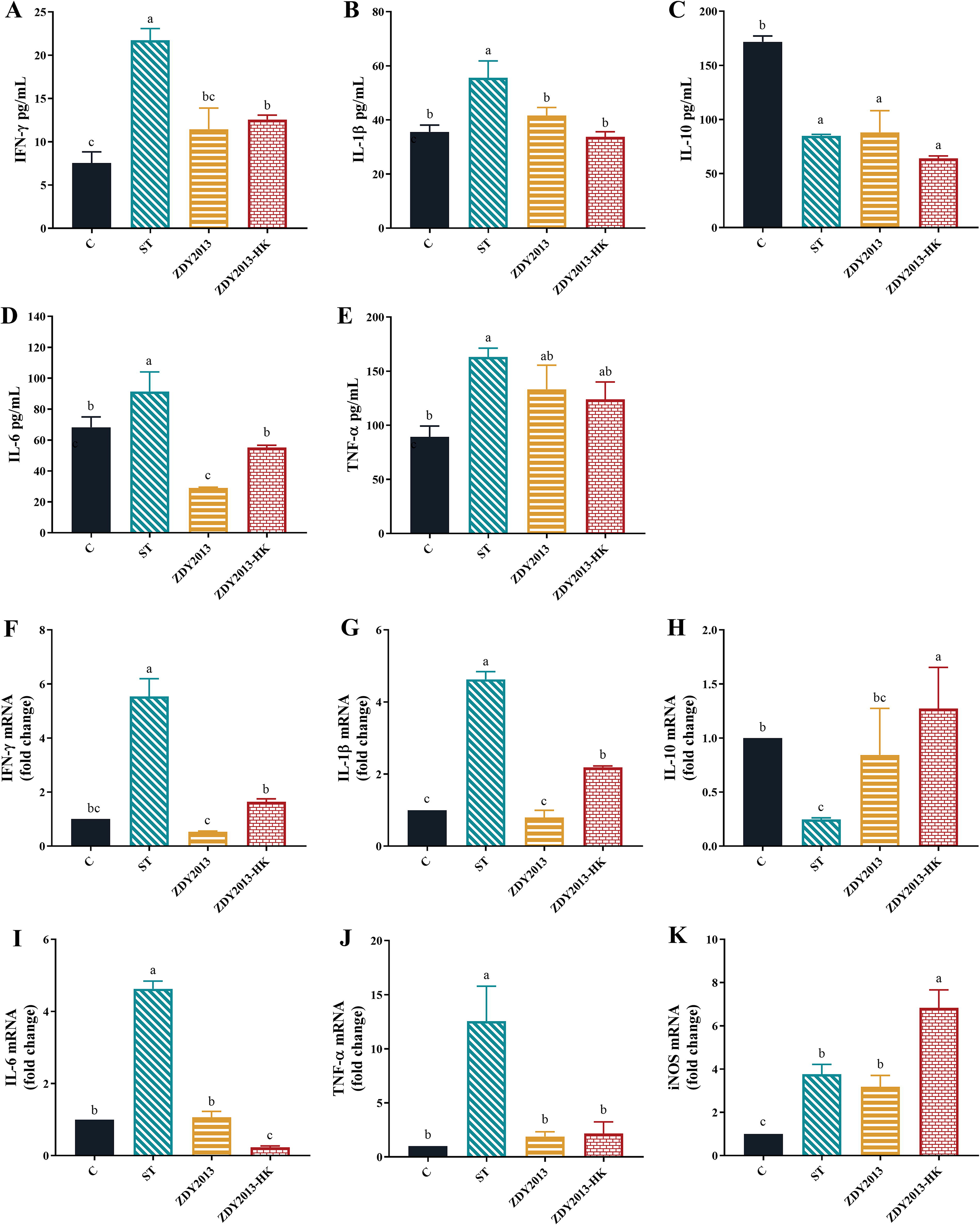
Effect of live and heat-killed L. plantarum ZDY2013 on cytokine and iNOS levels of mice. The contents of IFN-γ
Antiadhesion and invasion of L. plantarum ZDY2013 against S. Typhimurium in HT-29 cells
As shown in Figure 4, heat-killed L. plantarum ZDY2013 could significantly (p < 0.05) reduce the adhesion of S. Typhimurium to HT-29 cells (Fig. 4A). Both live and heat-killed L. plantarum ZDY2013 could significantly (p < 0.05) reduce the invasion of S. Typhimurium, with invasion rates of 2.92% and 22.94%, respectively (Fig. 4B).
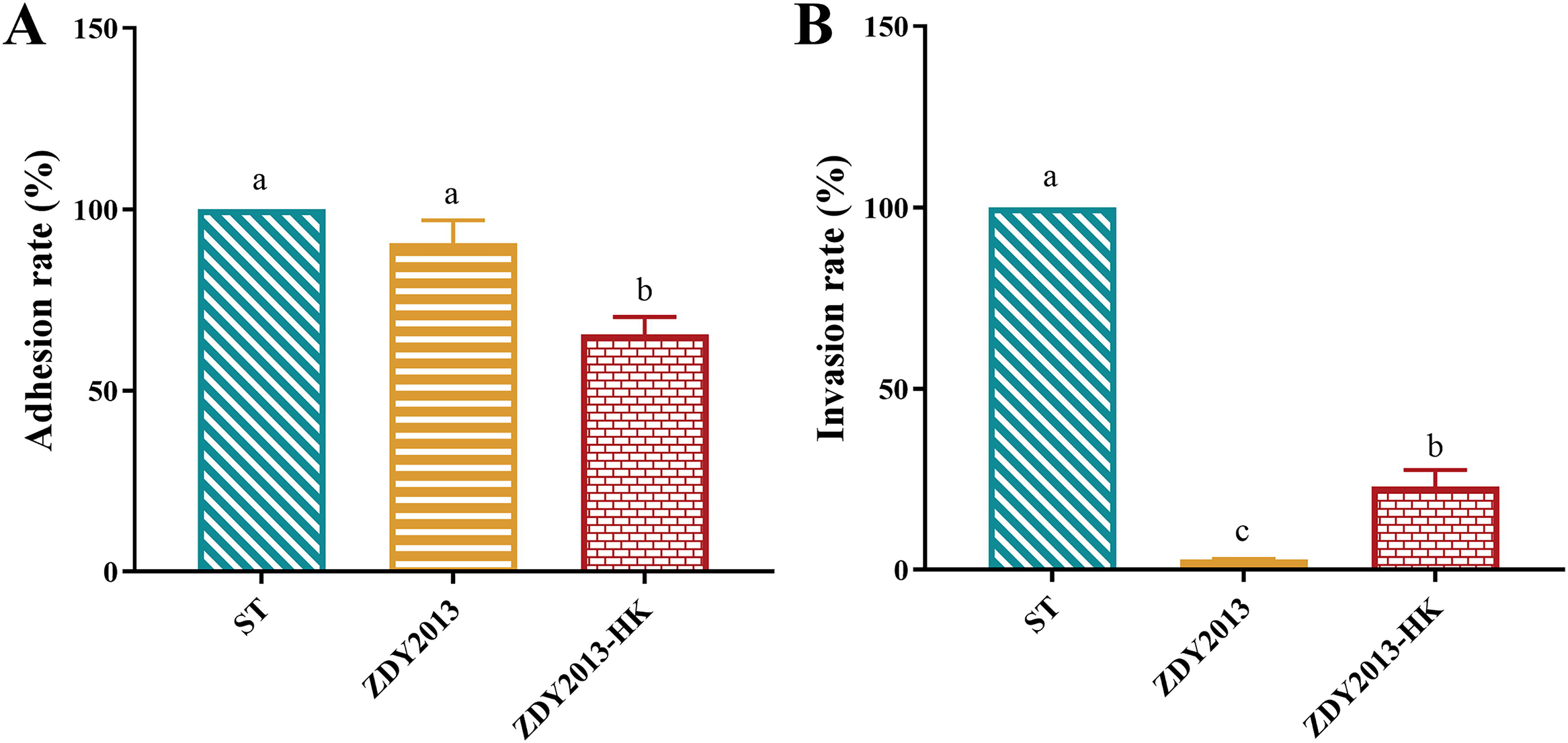
Effect of live and heat-killed Lactiplantibacillus plantarum ZDY2013 on adhesion
Effect of L. plantarum ZDY2013 on activating macrophage
The results showed that live and heat-killed L. plantarum ZDY2013 could enhance the phagocytosis rate of RAW264.7 cells (Fig. 5A). Furthermore, the survival assay in RAW264.7 cells showed that the viable number of S. Typhimurium in ZDY2013 and ZDY2013-HK groups was significantly (p < 0.05) higher than that in the ST group at 0 min and 30 min, but significantly (p < 0.05) lower after 60 min (Fig. 5B). These results indicate that both forms of ZDY2013 can enhance the phagocytic and bactericidal capacities of the macrophages.
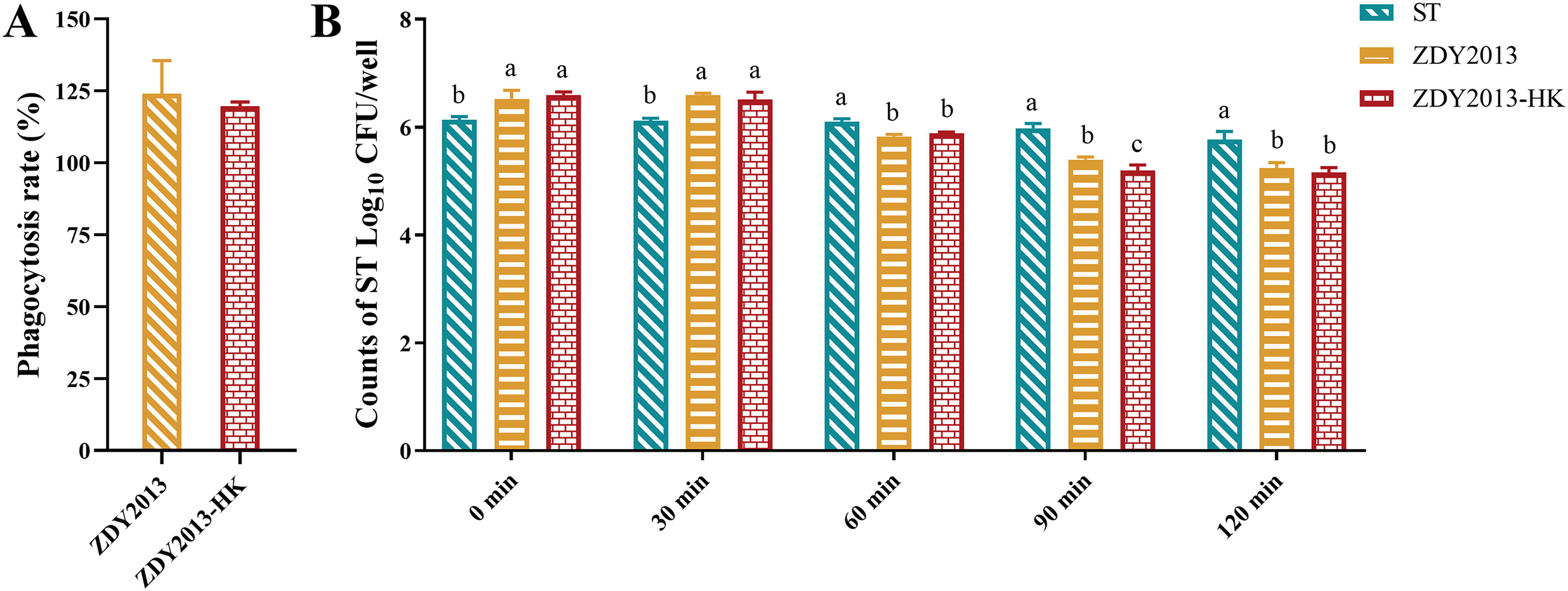
Effect of live and heat-killed Lactiplantibacillus plantarum ZDY2013 on phagocytosis of macrophage RAW264.7 cells and the survival of Salmonella Typhimurium in RAW264.7 cells. Phagocytosis ratio of live and heat-killed L. plantarum ZDY2013 treated RAW264.7 cells compared with untreated cells
Discussion
Probiotics play important roles in immunomodulation and enhancing epithelial barrier function to suppress intestinal inflammation caused by pathogens (Mousavi Khaneghah et al., 2020; Zhang et al., 2023). We established a mice infection model and found that live and heat-killed L. plantarum ZDY2013 could effectively alleviate weight loss, liver and spleen injury caused by S. Typhimurium infection, and reduce Salmonella loads. Similarly, live and heat-killed Lactobacillus reuteri GMNL-263 was reported to show antagonistic effects against Salmonella with similar fold reduction in Salmonella weight loss and bioburden in rats (Hsieh et al., 2016).
The intestinal epithelium, as the largest mucosal surface, provides interface between external environment and the host, and governs their interaction through intestinal permeability and mucosal defense (Fasano and Shea-Donohue, 2005). Abnormal expression of tight junction proteins causes barrier dysfunction and increased epithelium permeability (Li et al., 2018; Lucke et al., 2018). Goblet cells secrete mucus to prevent bacterial contact with intestinal epithelium (Birchenough et al., 2015). S. Typhimurium, as an enteric pathogen, mainly adheres and invades the distal small intestine and colon, uses its flagella and virulence island to open M cells into the lamina propria, destroys the intestinal barrier and further shifts to extraintestinal organs (Broz et al., 2012; Bula-Rudas et al., 2015). The in vitro HT-29 cell model showed that live and heat-killed L. plantarum ZDY2013 could effectively inhibit the adhesion and invasion of S. Typhimurium to cells, especially the latter. Inhibition of pathogen invasion by Lactiplantibacillus may be due to multiple factors: competitive exclusion of pathogens, enhancement of intestinal barrier function, regulation of the immune response (Li et al., 2012; Peng et al., 2022; Xu et al., 2009; Yu. et al., 2011). Previously, the anti-adhesion capacity of L. plantarum ZDY2013 against Helicobacter pylori was shown to be mainly due to its effective competitive effect (Zhao et al., 2018). However, the underlying mechanisms by which L. plantarum ZDY2013 inhibits the adhesion and invasion of S. Typhimurium need to be further studied.
Meanwhile, our results showed that live and heat-killed L. plantarum ZDY2013 could significantly restore the increased intestinal permeability caused by Salmonella via up-regulating the transcription and expression of intestinal tight junction proteins (Claudin-1 and ZO-1) and the number of goblet cells. The level of inflammation is also closely related to intestinal permeability. We found that live and heat-killed L. plantarum ZDY2013 could effectively reduce the level of proinflammatory cytokines (IFN-γ, IL-1β, IL-6, and TNF-α), but increase anti-inflammatory cytokine (IL-10). It has been reported that TNF-α induces reorganization of ZO-1 and Claudin-1 (Zolotarevsky et al., 2002), IFN-γ decreases the expression of ZO-1 and Occludin (Mankertz, 2000), and IL-10 blocks IFN-γ-indued epithelial permeability (Madsen et al., 1997). Therefore, these results verified the protective role of live and heat-killed L. plantarum ZDY2013 in the intestine of mice.
Moreover, we found that the administration of heat-killed L. plantarum ZDY2013 significantly increased the transcription level of iNOS, a crucial NOS isoform that triggers the synthesis of effector molecule NO (Palmieri et al., 2020). In response to inflammatory signals, immune cells such as macrophages increase production of cytokines and NO, which are important for killing pathogens (Palmieri et al., 2020). Macrophages play a crucial role in the host’s first-line defense against various infections. Under proinflammatory circumstances, activated macrophages (M1) directly neutralize xenobiotics by phagocytosis and kill pathogens by secreting proinflammatory cytokines and cytotoxic molecules such as NO (Cheng et al., 2019; Rocha-Ramirez et al., 2017). However, on the other hand, macrophages were used by Salmonella as their host cells, promoting the spread of Salmonella in the body, leading to infection (Wang et al., 2019). The macrophage model showed that live and heat-killed L. plantarum ZDY2013 could strengthen the phagocytosis of RAW264.7 and the intracellular killing effect on S. Typhimurium.
Conclusions
In conclusion, live and heat-killed L. plantarum ZDY2013 could effectively reduce the Salmonella load, improve the intestinal barrier, suppressing the adhesion and invasion of Salmonella to intestinal epithelium, upregulating the level of tight junction proteins and the number of goblet cells, activate the macrophage by enhancing the phagocytic ability, reduce the intracellular survival rate of Salmonella, and improve the intestinal inflammation (Fig. 6). Taken together, these findings contribute to better understand the mechanism by which live and heat-killed L. plantarum ZDY2013 regulates the host immune responses against pathogen infection, which will lay the foundation for developing protection and treatment strategies against S. Typhimurium infections in the future.
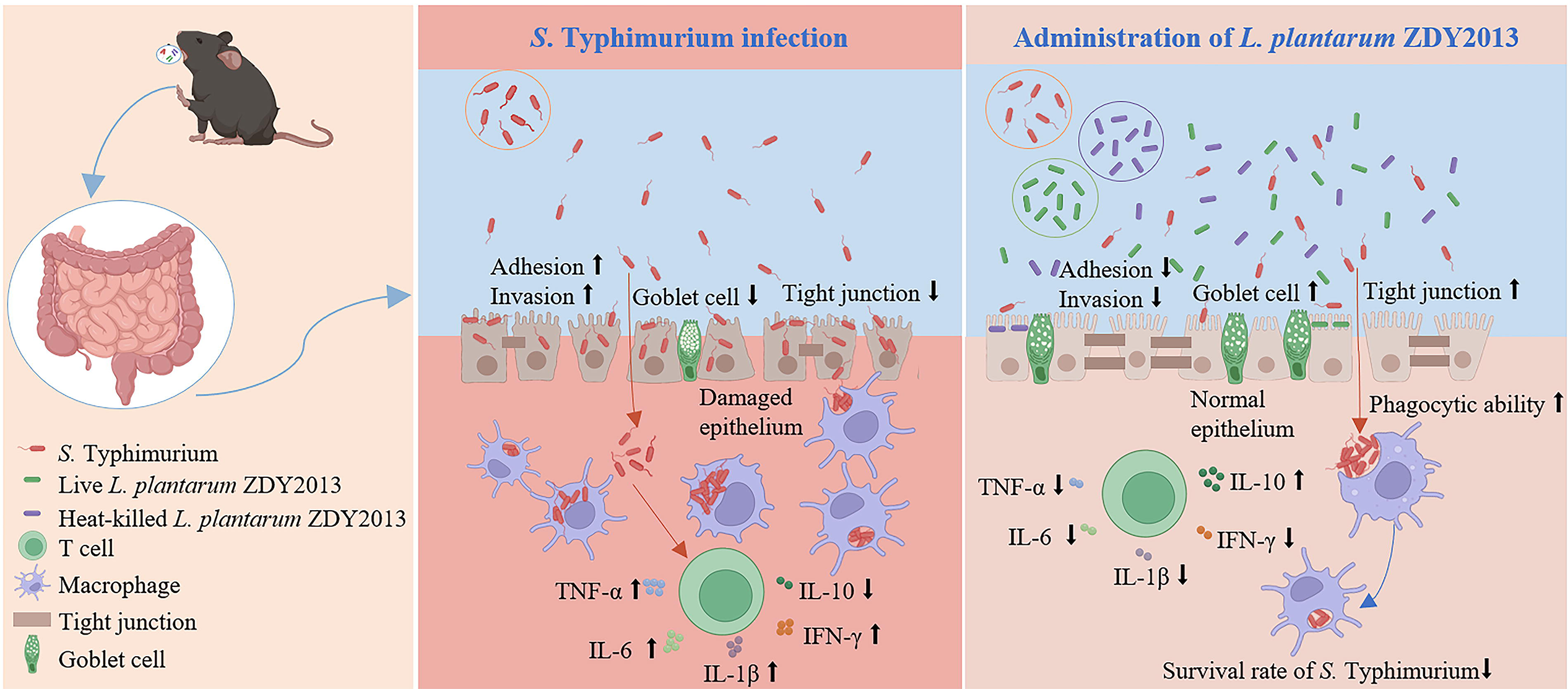
Proposed model of live and heat-killed Lactiplantibacillus plantarum ZDY2013 against Salmonella Typhimurium infection.
Authors’ Contributions
Y.W.: Methodology and writing—original draft. L.T.: Methodology, investigation, software, and validation. J.T.: Investigation, software, and validation. Q.L.: Investigation, software, and validation. Z.L.: Investigation, software, and validation. X.T.: Conceptualization, writing—review and editing, and data. H.W.: Conceptualization.
Footnotes
Author Disclosure Statement
The authors declare that they have no conflict of interest.
Funding Information
This work was supported by the National Natural Science Foundation of China (grant no. NSF 32060030, Beijing, China) and Jiangxi Provincial Natural Science Foundation (grant no. 20232BAB205007).
Ethical Approval
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.