
Abstract
Multidrug resistance in foodborne pathogens poses a critical threat to food safety and public health. Enterobacter hormaechei is an emerging pathogen with wide environmental prevalence and is capable of causing severe infections. Bacteriophage-based intervention has gained significant recognition as a sustainable approach to combat foodborne pathogens and address antimicrobial resistance in food production systems. Despite this, research on E. hormaechei-specific phages is facing substantial challenges, primarily due to insufficient phage collections and inadequate genomic characterization of existing isolates. In this work, two novel virulent phages (Ehp-YZU-L3 and Ehp-YZU-L4) were isolated from wastewater samples in Yangzhou. Their morphological, biological, and genomic features were characterized. The two phages belonged to the Myoviridae family, with a latency period (10 and 40 min) and high burst size (192 and 292 plaque-forming unit [PFU]/host cell), and an optimal multiplicity of infection of 0.01. The complete genomic sequences of phages ranged from 163,779 to 170,652 bp and GC content of 39.8 − 40.2%, which consisted of 296 and 275 open reading frames of phage Ehp-YZU-L3 and Ehp-YZU-L4. The absence of both virulence-related genetic elements, antimicrobial resistance, and lysogeny-related genes in two genomes was confirmed. Two phages exhibited strong inhibitory effects against E. hormaechei in pork by a phage-dosage–dependent way, with a reduction range of bacterial counts by 1.73–2.87 Log CFU/g for Ehp-YZU-L3 and 1.96–3.20 log CFU/g for Ehp-YZU-L4 at 37°C for 4 h. These findings demonstrate considerable potential of these two phages for the biocontrol of E. hormaechei contamination in food production systems.
Introduction
Enterobacter hormaechei has emerged as a microbial risk owing to its recurrent contamination of food products, with primary sources including infant formula, animal-derived foods, and ready-to-eat fruits (Leighton et al., 2024; El-Zamkan and Mohamed, 2018). This foodborne pathogen usually causes infections that are particularly dangerous for immunocompromised individuals, neonates, and the elderly (Liu et al., 2022).
At this stage, the application of antibiotics remains the conventional approach for controlling E. hormaechei. However, inappropriate and excessive utilization of antibiotics causes the emergence of multidrug-resistant E. hormaechei and raises consumer concerns (Yeh et al., 2022). Studies have revealed the emergence of E. hormaechei carrying resistance genes, notably blaNDM-1, blaCTX- M -15, blaACT-25, blaKPC-2, blaVIM-1, and mcr-9. This phenomenon facilitates emergence of carbapenem-resistant E. hormaechei and intensifies global antimicrobial resistance crisis (Sadek et al., 2020; Leighton et al., 2024). In the global priority list of antibiotic-resistant bacteria, World Health Organization has identified carbapenem-resistant Enterobacter species as critical-priority pathogens, emphasizing immediate requirement for novel antimicrobial development. These limitations highlight the urgent need for alternative and effective biocontrol strategies to mitigate its presence in food products and food processing environment.
Phages demonstrate safety profiles and host specificity without interference with human microbiota and are generally recognized as safe (Chen et al., 2022). Developing phage-based antimicrobial agents demonstrates significant advantages in both temporal and financial aspects. Phages have been widely used to reduce the risk of antibiotic-resistant pathogens like Salmonella enterica, Staphylococcus aureus, Listeria monocytogenes, and Escherichia coli (Kosznik-Kwasnicka et al., 2022; Assad-Garcia et al., 2022; Abdullah et al., 2025; Shin et al., 2024), and spoilage bacteria like Pseudomonas fluorescens, Bacillus methylotrophicus, and Brochothrix thermophacta (Liu et al., 2025; Yuan et al., 2025; Lv et al., 2023). However, there is limited research published on phages targeting E. hormaechei, and this gap is particularly concerning given the increasing prevalence of multidrug-resistant E. hormaechei in the food industry.
This study aims to expand knowledge of E. hormaechei phages and highlight their critical role in advancing phage-based biocontrol strategies in the food industry. Specifically, two novel lytic phages exhibiting high specificity toward E. hormaechei were obtained from wastewater samples in Yangzhou, China. We then systematically characterized their biological traits and genomic features, followed by an assessment of their antimicrobial potential in a pork model.
Materials and Methods
Bacterial strains
The host strain E. hormaechei Eh-YZU5-3, originally isolated and identified from a pork sample collected in Yangzhou, was stored at −80°C in glycerol-supplemented (20%, v/v) tryptic soy broth (TSB). According to the disc diffusion method of Clinical Laboratory Standards Institute guideline (2022), E. hormaechei Eh-YZU5-3 is resistant to penicillin, amoxicillin, oxacillin, lincomycin, azithromycin, erythromycin, and rifampicin with a diameter of inhibition zone higher than 18 mm. Before use, E. hormaechei YZU5-3 was revived by streaking onto tryptic soy agar (TSA) plates and subsequent cultivation at 37°C for 12 h. All experiments were performed using biosafety level-2 facilities.
Isolation and purification of phages
Phages were obtained from untreated domestic sewage samples in Yangzhou from November to December 2023. Phage enrichment, isolation, and purification were conducted according to a previously reported method (Tang et al., 2023). Each 10 mL wastewater sample was subjected to centrifugation at 8000 × g and 4°C for 10 min for impurity removal, and resulting supernatant was filtered with 0.22 μm filters. Then, 100 μL culture of Eh-YZU5-3 at 37°C for 12 h was combined with 5 mL of each filtrate in semisolid TSB plates (0.7% agar), followed by incubation at 37°C for 18 h for clear plaque observation. Individual transparent plaque was suspended in SM buffer (100 mmol/L NaCl, 8 mmol/L MgSO4, 50 mmol/L Tris-HCl at pH 7.5) and combined with 100 μL of Eh-YZU5-3 overnight culture to form a double-layered plate. Following a 12 h incubation at 37°C, phage titer was calculated as described by Liu et al. (2025).
Phage morphology
Phage morphology was visualized using transmission electron microscopy (TEM) as described by Tang et al. (2023). In brief, 10 µL amplified phage suspension was deposited onto copper grids for 10 min adsorption, and then stained with 2% (w/v) phosphotungstic acid for 2 min and air-dried for 2 h. TEM analysis was performed at 80 kV accelerating voltage for phage morphological observation. Phage classification was conducted in accordance with taxonomic guidelines established in the Ninth Report of the International Committee on Taxonomy of Viruses (ICTV).
Phage host range
Eleven strains (Bacillus licheniformis 2–7, Bacillus cereus 38B-1, Staphylococcus aureus YZUstau1, Enterobacter hormaechei EH23487, E. hormaechei EH10432, E. hormaechei EH2-5, Escherichia coli CICC10664, Cronobacter sakazakii CICC 22919, Salmonella Typhimurium N26, Salmonella Enteritidis N23, and Pseudomonas fluorescens PF2) were selected to analyze host spectrum by spot assay (Zhang et al., 2023). Ten microliters of each strain (107 CFU/mL) were dropped onto TSA plates. Then, 10 μL of phage suspension (108 plaque-forming unit [PFU]/mL) was overlaid on the bacterial lawn. Plates were incubated for 6 h at 37°C. Host spectrum was evaluated by the presence of clear lytic zones.
Optimal multiplicity of infection (MOI) of phages
To evaluate the MOI (10, 1, 0.1, 0.01, 0.001) of phages, 100 μL of phage suspension was mixed with an equal volume of Eh-YZU5-3 (106 CFU/mL) in 5 mL TSB to achieve various MOI values. After incubation at 37°C for 12 h, the culture was centrifuged for 10 min at 8000 × g, and subsequent filtration was done by a 0.22-μm filter. Phage titers were quantified using a double-layer plating method, and maximum titer was the optimal MOI (Tang et al., 2023).
One-step growth curve
One-step growth curve was determined following the methodology reported by Tang et al. (2023). Briefly, phage-host mixtures were prepared at optimal MOI values, followed by 10 min incubation at 37°C. Cultures were centrifuged at 4°C and 12,000 × g for 1 min, and obtained pellets were reconstituted in 5 mL TSB and subjected to shaking at 37°C for 180 min. Samples were taken out with 10 min intervals during the first hour and 20 min intervals for following 2 h. Infection time (min) was plotted on horizontal axis, and logarithmic phage titer value (Lg PFU/mL) was plotted on vertical axis. Latent period, burst time, and burst size were determined from one-step growth curve.
Evaluation of phage stability
Thermal stability was evaluated by mixing 100 μL phage with 900 μL Phosphate Buffered Saline (PBS) and incubating at 50°C, 60°C, and 70°C for 60 min. Phage pH stability was assessed across a pH range of 2–12. PBS solutions were pH-adjusted using HCl or NaOH. Subsequently, 100 μL phage suspensions were mixed with 900 μL pH-adjusted PBS and incubated at 37°C for 2 h. Phage titer was determined using double-layer plating method to evaluate phage stability (Tang et al., 2023).
Lytic ability of phages at different MOIs
Overnight culture of Eh-YZU5-3 (106 CFU/mL) was mixed with phage suspension to achieve different MOIs (0.01, 0.1, 1, and 10), while a phage-free group was set as the control. After incubation for 12 h at 37°C, phage lytic activity was determined by measuring OD600nm values at 2 h intervals.
Complete genome sequencing of phages
Genomic DNA of each phage was extracted and purified by ABigen Lambda Phage Genomic DNA Kit (AB1141, ABigen Corporation, Beijing, China). Each genome was sequenced using Illumina NovaSeq 6000 platform (Illumina, San Diego, USA). Sequence assembly was conducted using ABySS (http://www.bcgsc.ca/platform/bioinfo/software/abyss) and GapCloser (https://sourceforge.net/projects/soapdenovo2/files/GapCloser/). Gene prediction and annotation were performed using GeneMarkS (http://topaz.gatech.edu/) and NCBI database (https://www.ncbi.nlm.nih.gov/). Virulence factors were assessed through VFDB online database (http://www.mgc.ac.cn/cgi-bin/VFs/v5/main.cgi), and resistance genes in genomes were analyzed using ResFinder (http://genepi.food.dtu.dk/resfinder). Phylogenetic tree analysis of 25 phage genomes was performed using the maximum likelihood algorithm in MEGA 11 software, with subsequent tree visualization and annotation conducted using iTOL (https://itol.embl.de/) (Tang et al., 2023).
Biocontrol of E. hormaechei in pork
Two hundred grams of fresh pork were exposed to 5% sodium hypochlorite for 20 min and rinsed with sterilized water to remove indigenous microbiota. Each prepared pork sample was artificially inoculated with Eh-YZU5-3 (104 CFU/mL) and treated with 100 μL phage suspension at different MOIs (0.01, 0.1, 1, and 10), with a phage-free group set as the control. Following 10 h incubation at 37°C, viable E. hormaechei was counted by plate counting on TSA plates.
Statistical analysis
All experiments were independently performed in triplicate, with results presented as mean ± standard deviation analyzed by SPSS 22.0. A one-way analysis of variance with Tukey’s test was performed to determine the differences, and significant differences were determined at a significance level of p < 0.05.
Results and Discussion
Isolation, morphology, and host range of phages
Previous results have demonstrated the isolation and application of phages to control E. hormaechei in the food industry (Torabi et al., 2022; Chen et al., 2023). However, research gaps indicated that remarkable phenotypic and genetic heterogeneity observed in natural populations limited comprehensive understanding of phages to inhibit multidrug-resistant E. hormaechei.
In this study, two novel lytic E. hormaechei phages, namely Ehp-YZU-L3 and Ehp-YZU-L4, were successfully isolated from domestic wastewater samples. As shown in Figure 1, Ehp-YZU-L3 and Ehp-YZU-L4 displayed regular icosahedral heads with a diameter of 75.00 and 74.35 nm and long and noncontractile tail measuring approximately 101.25 and 52.15 nm, respectively. Both Ehp-YZU-L3 and Ehp-YZU-L4 belonged to Myoviridae family within Caudovirales order. In contrast, previously identified E. hormaechei phages (Ehp-YZU9-2, Ehp-YZU9-3, Ehp-YZU10) belonged to the Podoviridae family (Chen et al., 2022), phage IME278 belonged to the Autographiviridae family (Chen et al., 2023), and phage PɸEn-HO belonged to Inovirus (Torabi et al., 2022). Results indicated a diverse morphology and high variability of E. hormaechei phages in nature and highlighted the importance of discovering diversity within E. hormaechei phages.

Transmission electron microscopy (TEM) micrographs of two Enterobacter hormaechei bacteriophages, Ehp-YZU-L3
As shown in Figure 2, lytic capacity of individual phages was determined against B. licheniformis 2–7, B. cereus 38B-1, S. aureus YZUstau1, E. hormaechei EH23487, E. hormaechei EH10432, E. hormaechei EH2-5, E. coli CICC10664, C. sakazakii CICC 22919, S. Typhimurium N26, S. Enteritidis N23, and P. fluorescens PF2. Two phages exhibited specifically lytic activity against E. hormaechei, which is mediated by highly specialized tail-spike proteins that facilitate targeted host recognition. Narrow host ranges of Enterobacter phages like EspYZU14, Ehp-YZU08, and Ehp-YZU10 were previously reported (Chen et al., 2022). Narrow host range enhances phage suitability for specific detection of bacteria in different food matrices with minimal false-positive results (Zhang et al., 2022; Zhang et al., 2023). Phage cocktail can improve host range and infect different bacterial species and minimize the emergence of phage-resistant bacteria (Choi et al., 2025).
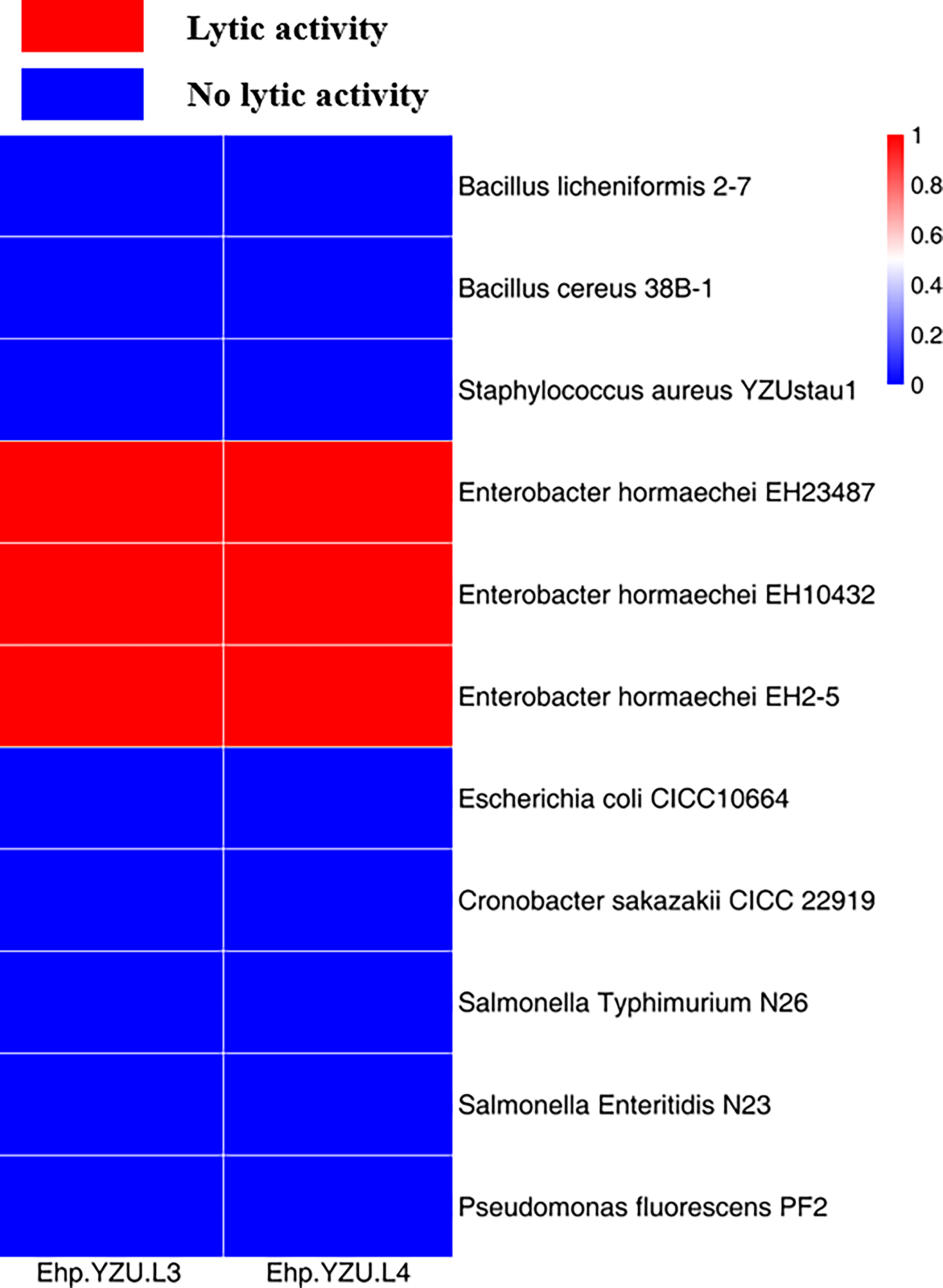
Determination of host range of phages Ehp-YZU-L3 and Ehp-YZU-L4.
MOI of phages
As shown in Figure 3, optimal MOI of Ehp-YZU-L3 and Ehp-YZU-L4 was 0.01, with not significantly (p > 0.05) different titers of 1.01 × 1010 and 1.02 × 1010 PFU/mL. In a previous work, the optimal MOI of phage IME278 was also 0.01 (Chen et al., 2023). Low optimal MOIs of phages suggested that a minimal number of phages can yield a substantial quantity of progeny, which is beneficial in food industry applications.
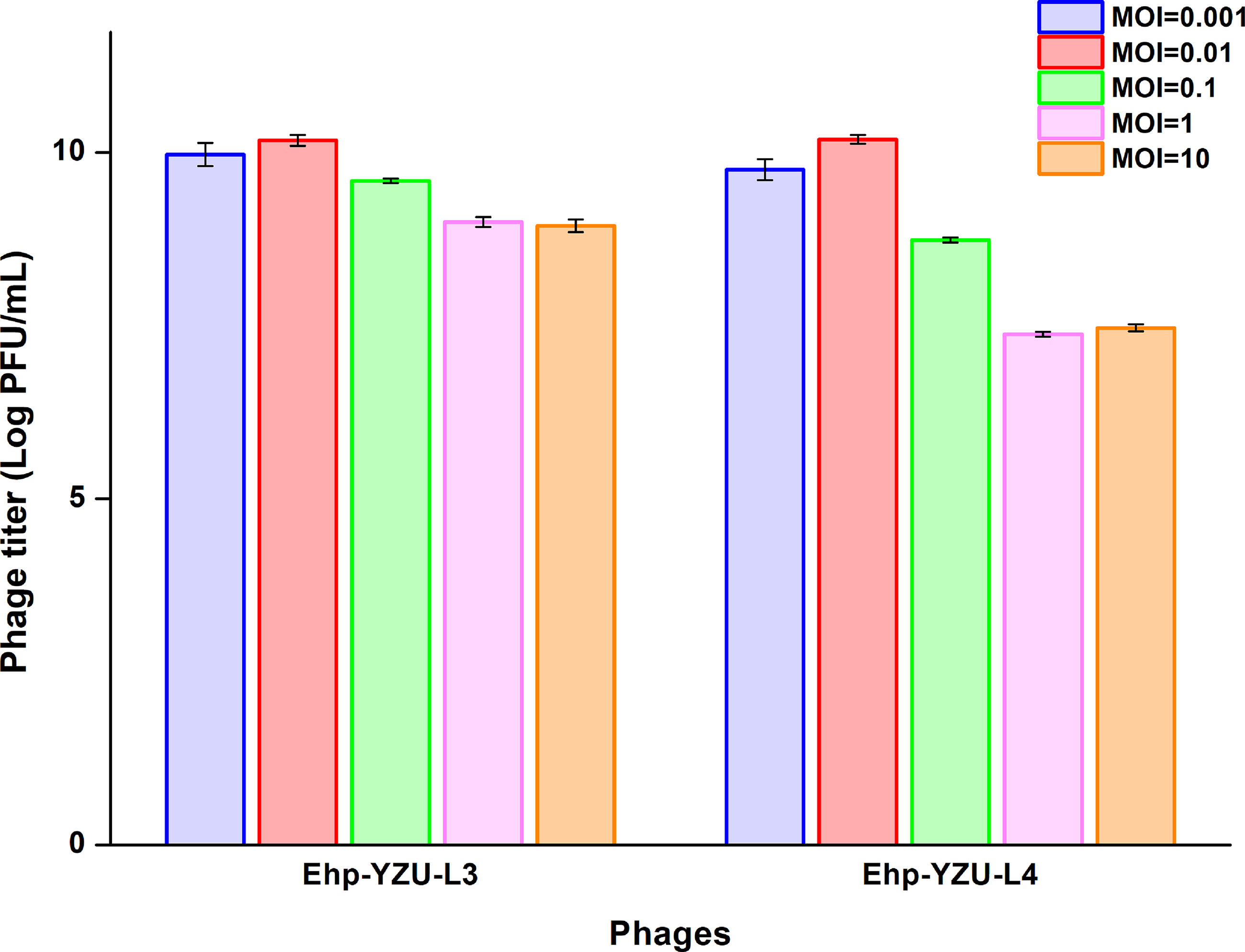
Determination of the optimal multiplicity of infection (MOI) of phages Ehp-YZU-L3 and Ehp-YZU-L4. All values are expressed as the mean ± standard deviation from three independent biological replicates (n = 3). Different letters indicate the significant differences (p < 0.05) in the phage titer of each phage.
One-step growth curve of phages
As presented in Figure 4, Ehp-YZU-L3 and Ehp-YZU-L4 exhibited significantly different (p < 0.05) burst sizes of 192 and 292 PFU/host cell, latent periods of 10 and 40 min, and burst periods of 120 and 140 min. A shorter latent period indicates that phages can efficiently inject their nucleic acids and proliferate, assemble, and release. Phages with high lysis yields achieve rapid concentration increases by 100-fold in a short duration (Amarillas et al., 2017). Phages in this study demonstrated shorter latent times and greater burst sizes compared with other E. hormaechei and Enterobacteriaceae phages (Chen et al., 2022; Torabi et al., 2022).
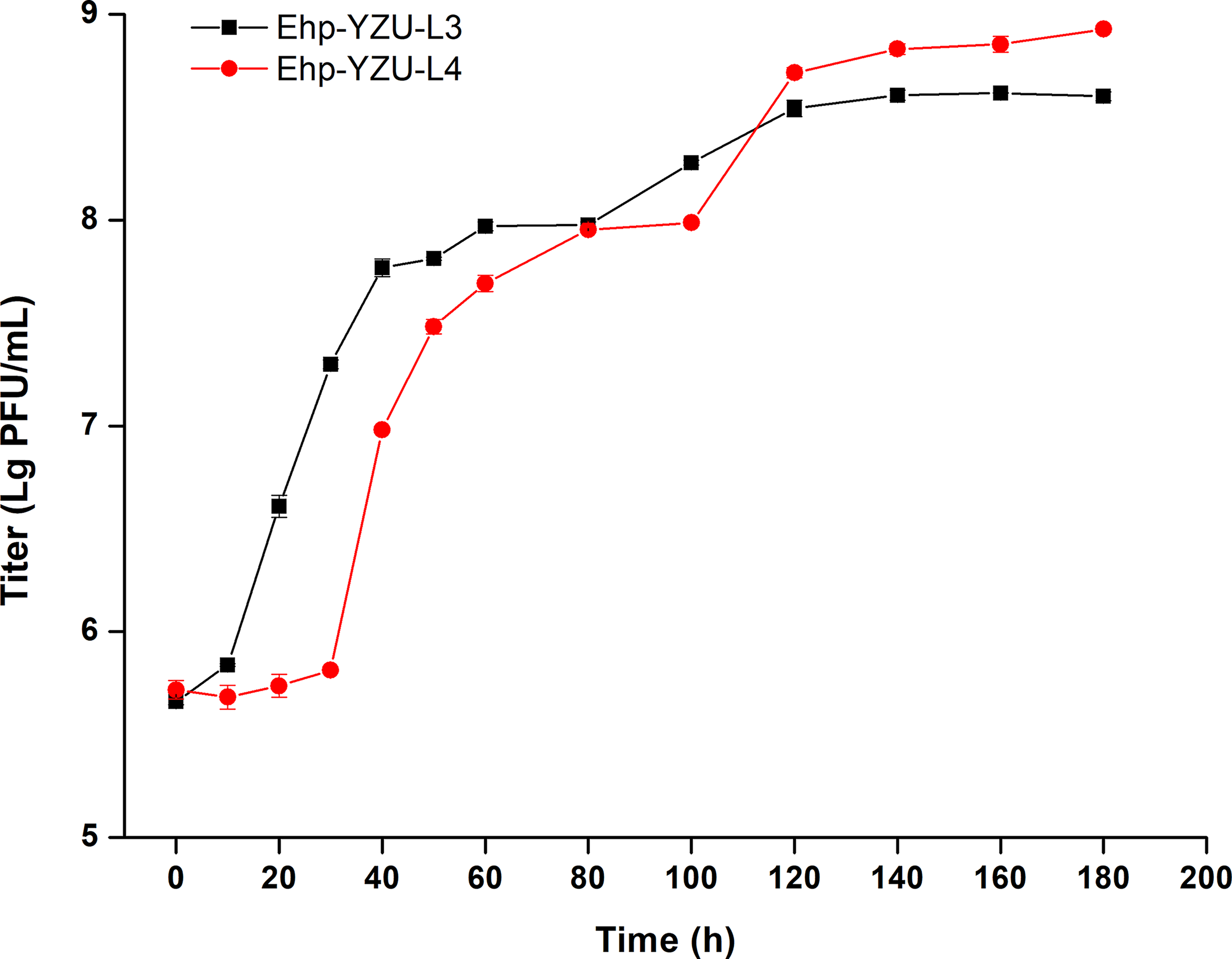
One-step growth curve of the two phages. Phage suspensions were collected every 10 min for 60 min and every 20 min for another 120 min. All values are obtained from three independent biological replicates (n = 3).
Thermal and pH stability of phages
As shown in Figure 5, lytic activity of phages had an initial titer of 107 PFU/mL after 2 h across a broad pH spectrum of 2–12. Ehp-YZU-L3 demonstrated good lytic activity against its host with a titer around 107 PFU/mL across a pH range of 3–10. However, significantly (p < 0.05) less pH stability of Ehp-YZU-L4 was observed, since Ehp-YZU-L4 maintained a titer around 105 PFU/mL within pH spectrum of 3–11. Titer of phages rapidly decreased under extremely low (pH < 3) or high (pH > 11) conditions. Low pH can cause denaturation of capsid proteins, while high pH may disrupt the integrity of phage genomes (Jończyk et al., 2011).
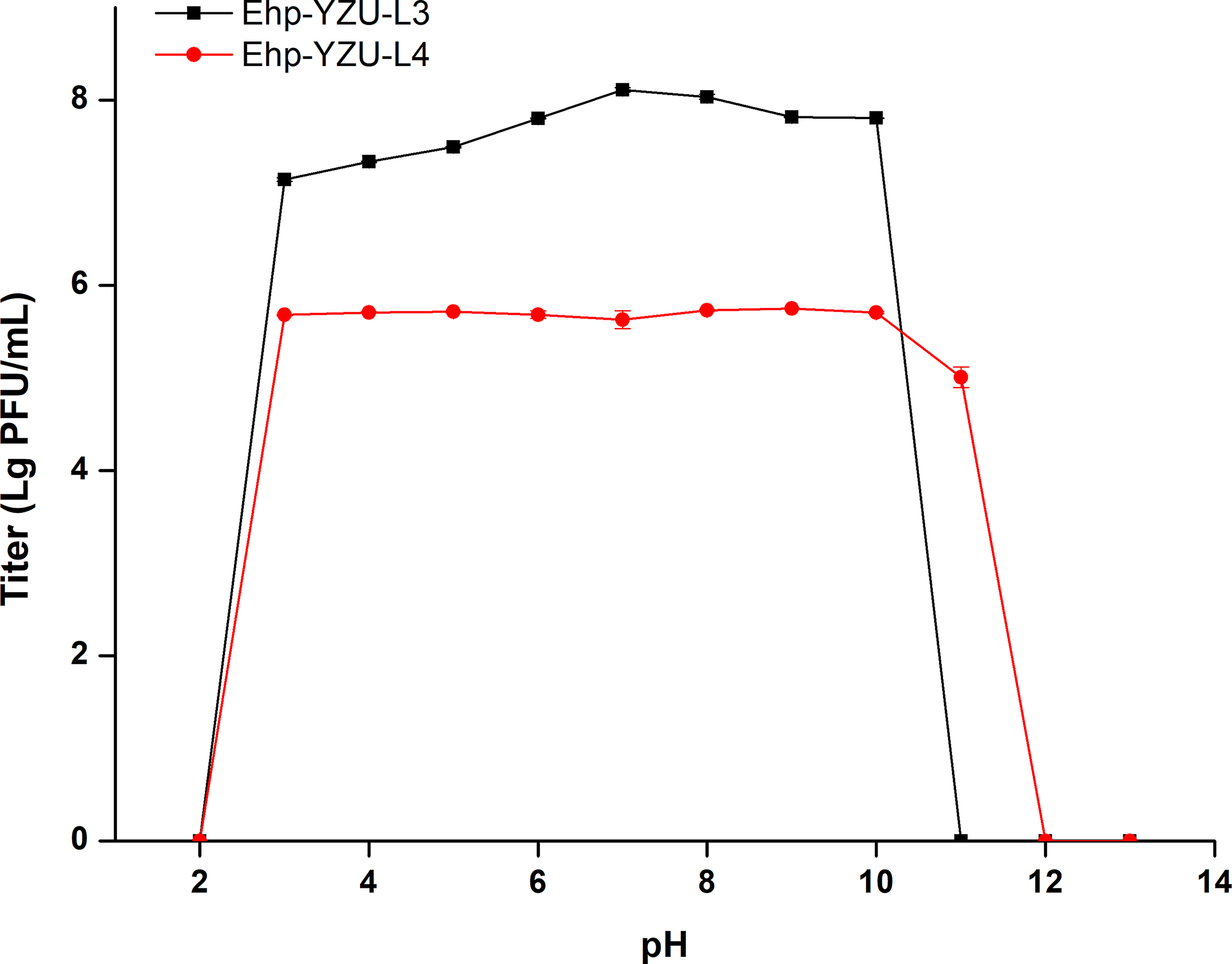
Stability of phages Ehp-YZU-L3 and Ehp-YZU-L4 at various pH values (2–14). All values are obtained from three independent biological replicates (n = 3).
This study assessed phage activity with an initial titer of 1010 PFU/mL after 60 min treatment at 50°C, 60°C, and 70°C. As shown in Figure 6A, activity of Ehp-YZU-L3 significantly (p < 0.05) decreased to 6.45 and 5.96 Log PFU/mL after treatment for 60 min at 50°C and 60°C but completely lost lytic activity at 70°C for 40 min. Ehp-YZU-L4 showed significantly (p < 0.05) less resistance to temperature and was totally inactive at 60°C for 20 min (Fig. 6B). High temperatures induce protein denaturation, nucleic acid degradation, and capsid disassembly, leading to irreversible loss of function. Our results align with previous studies, confirming that Enterobacter phages are highly sensitive to elevated temperatures while remaining stable at moderate and low temperatures (Chen et al., 2022). For this reason, their sensitivity to high temperatures ensures easy inactivation during thermal processing, eliminating residual concerns and enhancing food safety. In addition, this property allows phages to be used in targeted, pre-thermal treatment stages for pathogen control without interfering with subsequent processing steps (Tang et al., 2023; Yuan et al., 2025).
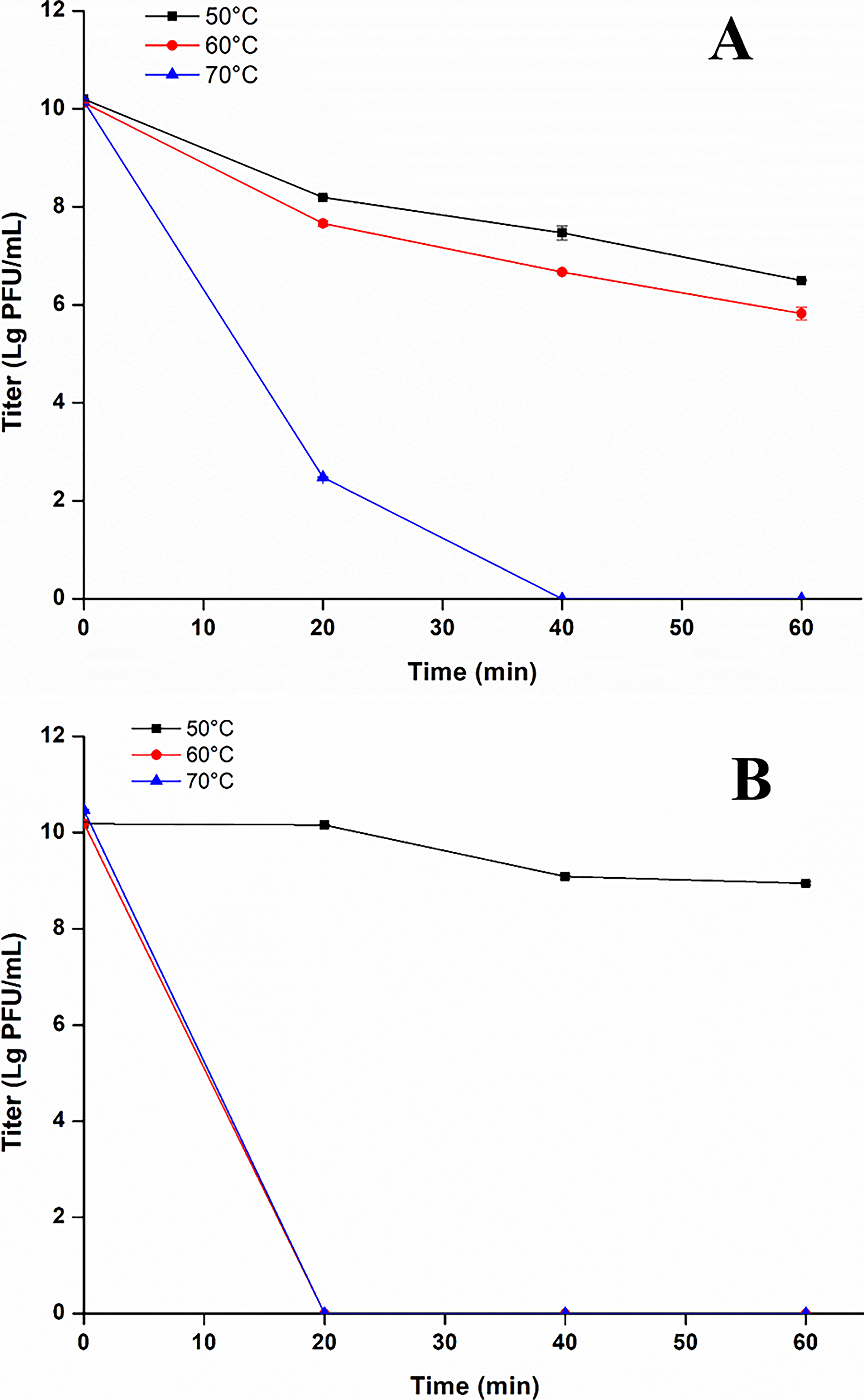
Stability of phages Ehp-YZU-L3
Lytic ability of phages
As shown in Figure 7, both phages inhibited E. hormaechei growth within 12 h and significantly (p < 0.05) reduced OD600nm values in phage-treated groups when tested in TSB. Furthermore, the inhibitory effect was MOI-dependent, supporting the hypothesis that endolysin activity is a primary mechanism for host inhibition (Zhang et al., 2023).
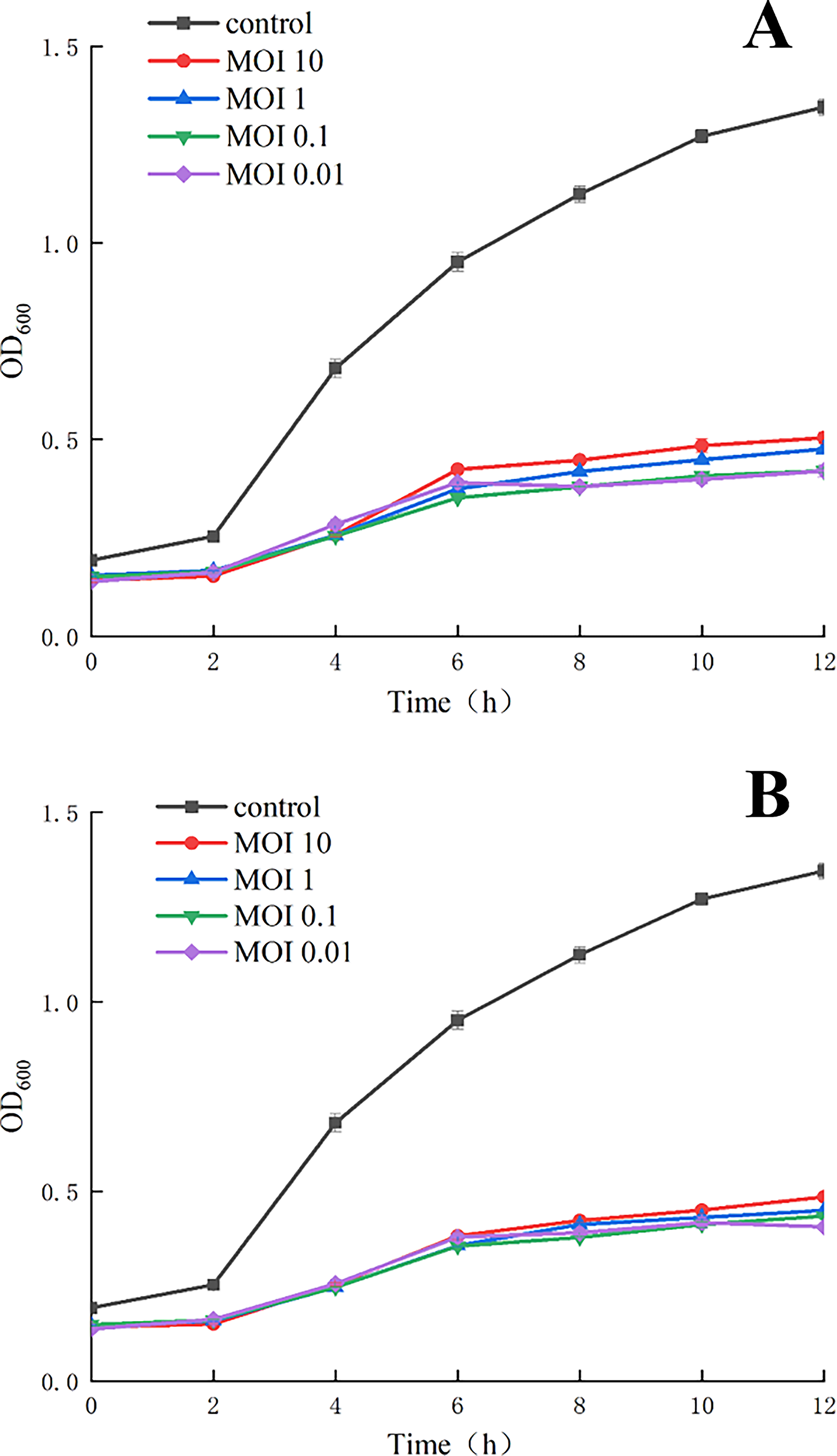
Lytic ability of Ehp-YZU-L3
Genome sequencing of phages
The type, GC content, full length, total predicted open reading frames (ORFs), and number of functionally annotated ORFs of phages are provided in Table 1. The DNA type of phages was double-stranded, with a genome length of 163,779–170,652 bp, and GC content of 39.8 − 40.2%. The genome size of both phages was higher than previously reported E. hormaechei phages IME278, Ehp-YZU08, Ehp-YZU10, Ehp-YZU9-2, and Ehp-YZU9-3 (Chen et al., 2022; Chen et al., 2023). Meanwhile, the GC contents of Ehp-YZU-L3 and Ehp-YZU-L4 were lower than those of the reported E. hormaechei phages in the same studies. Moreover, genome‐wide analysis of phages revealed a total of 296 and 275 ORFs of Ehp-YZU-L3 and Ehp-YZU-L4, respectively, and 146 and 87 ORFs of which were functionally annotated (Fig. 8A and B). High number of hypothetical proteins suggests a scarcity of homologous phage genes in existing GenBank database.
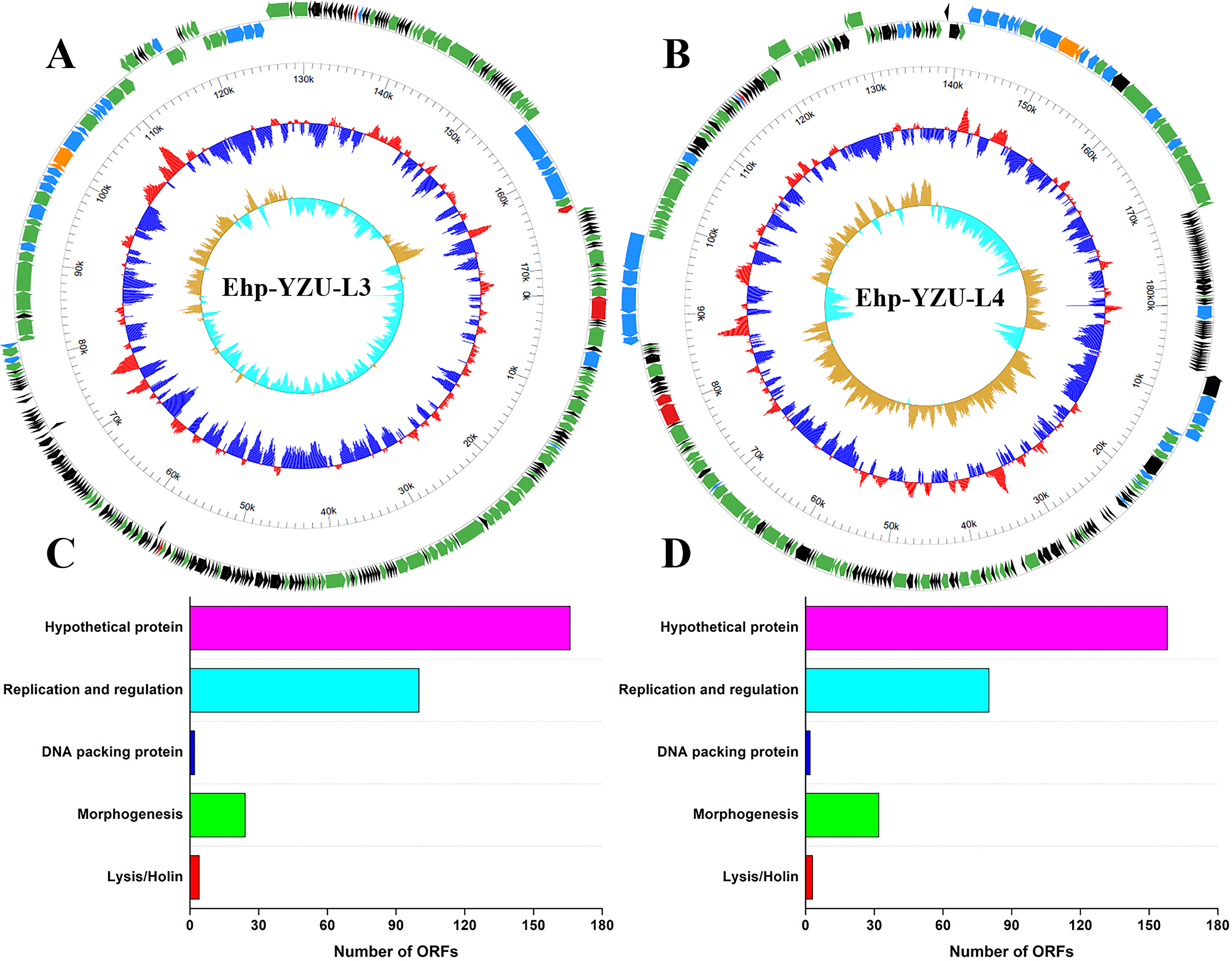
Genome map of phages Ehp-YZU-L3
Genome Features of Two Phages, Ehp-YZU-L3 and Ehp-YZU-L4

ORFs, open reading frames.
As shown in Figure 8C and D, all predicted ORFs were categorized into five distinct functional modules: lysis/holing (4 and 3 proteins for Ehp-YZU-L3 and Ehp-YZU-L4), morphogenesis (24 and 32 proteins for Ehp-YZU-L3 and Ehp-YZU-L4), DNA packing protein (2 proteins for Ehp-YZU-L3 and 2 proteins for Ehp-YZU-L4), replication and regulation (100 and 80 proteins for Ehp-YZU-L3 and Ehp-YZU-L4), and hypothetical proteins. Detailed functional characterization of individual ORFs is presented in Supplementary Table S1. The lysis module, encoding endolysin and associated proteins, facilitates host cell lysis during the terminal phase of the phage life cycle, enabling the release of progeny virions. Lysins play an indispensable role in phage-mediated bacterial lysis, while endolysins target peptidoglycan bonds within cell wall structures. ORFs for holin (ORF131 in Ehp-YZU-L3, ORF131 in Ehp-YZU-L4) and endolysin (ORF276 in Ehp-YZU-L3, ORF53 in Ehp-YZU-L4) were found in phages. Expression and application of lytic proteins offer promising potential for antimicrobial interventions.
In replication and regulation module, genes encoding key replication machinery components, including DNA polymerase, primase/helicase, single-stranded DNA-binding proteins, and ATP-dependent DNA ligase, indicate a significant level of autonomy in phage DNA replication and transcription processes relative to host cellular systems (Cheng et al., 2024). Predicted DNA polymerase and helicase genes exhibited functional activation that was presumably mediated by the single-stranded DNA-binding protein, which plays a crucial role in DNA repair mechanisms and genomic reorganization processes. For packaging model, Ehp-YZU-L3 and Ehp-YZU-L4 have DNA packaging proteins that facilitate the precise encapsulation of viral genome within the capsid. Two ORFs for terminase small subunit (ORF185 in Ehp-YZU-L3, ORF226 in Ehp-YZU-L4) specifically facilitate viral genome recognition during pre-virion formation, while terminase large subunit (ORF186 in Ehp-YZU-L3, ORF225 in Ehp-YZU-L4) mediates genome cleavage and subsequent translocation into procapsid structure. In morphogenesis module, notable disparities in structural proteins were identified, revealing distinct compositional patterns that align with morphological distinctions captured by TEM imaging. Moreover, infection process initiates through the interaction between phage tail fiber proteins and host-specific surface receptors. These structural components are important for bacterial recognition and binding specificity. The abundance of tail fiber-encoding genes in phages indicates their enhanced capacity for host infection and suggests evolutionary adaptation for efficient host recognition.
As shown in Figures 9 and 10, Ehp-YZU-L3 and Ehp-YZU-L4 have evolutionary homology with Enterobacteria phage CC31 (Accession Number NC_014662.1) from wastewater in Asia and Cronobacter phage LPCS28 (Accession Number NC_070937.1) from an unknown sample in Asia. According to ICTV, genome sequence identity of <95% suggests that Ehp-YZU-L3 and Ehp-YZU-L4 are newly identified phages.
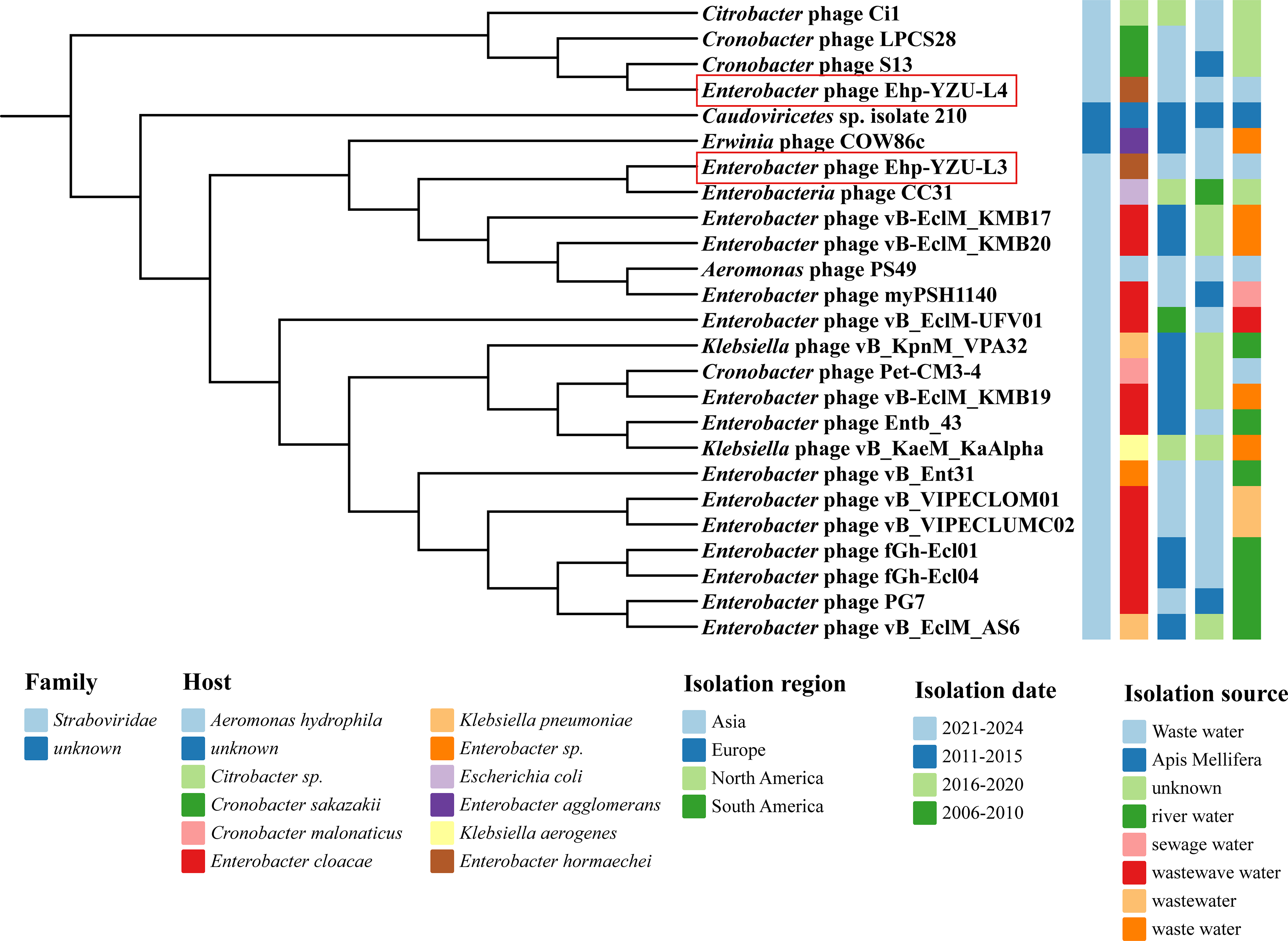
Whole-genome phylogenetic tree of phage Ehp-YZU-L3 and Ehp-YZU-L4 with other 23 Enterobacter phages in the NCBI database.
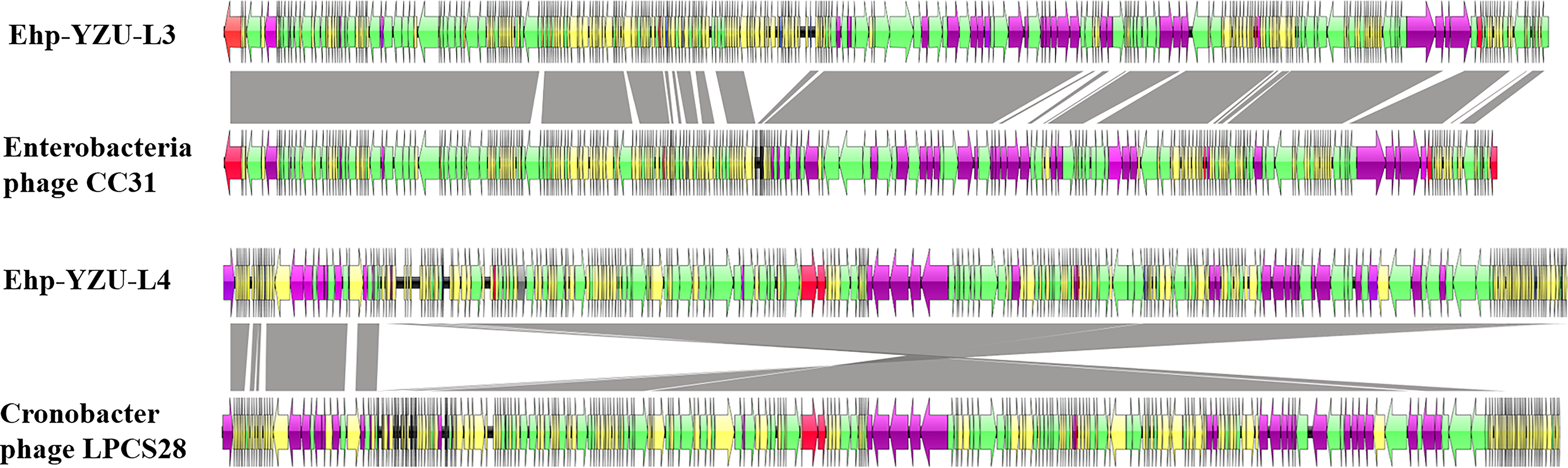
Comparative analysis of the whole genome sequence between Ehp-YZU-L3 and Enterobacteria phage CC31, and Ehp-YZU-L4 and Cronobacter phage LPCS28.
By phylogenetic tree analysis, Ehp-YZU-L3 and Ehp-YZU-L4 and other 23 phages are divided into two branches. Clade 1 included 4 phage genomes, including Ehp-YZU-L4, while Clade 2 contained 21 phage genomes, including Ehp-YZU-L3. Phages with high homology may have originated from different regions at different times, reflecting wide distribution of E. hormaechei phages and significant genetic evolutionary distance. Given limited records of E. hormaechei phages in current genomic databases, this investigation addresses a significant research gap by elucidating characteristics of E. hormaechei phages.
Both phage genomes encode no virulence genes, antibiotic resistance genes, lysogeny-related genes, or tRNA genes through VFDB and ResFinder databases, further supporting that Ehp-YZU-L3, Ehp-YZU-L4, and their derivatives are natural biocontrol agents without potential safety issues. Further analysis of unknown functional genes in phage genomes will facilitate the development of safer, more efficient phage agents exhibiting high solubility and a broad host range.
Antibacterial effect of phages on E. hormaechei in pork
As shown in Figure 11A and B, the bacterial count of control group without phages increased from 4.15 to 8.42 Log CFU/g after being cultured at 37°C for 10 h, while Ehp-YZU-L3 and Ehp-YZU-L4 significantly (p < 0.05) inhibited E. hormaechei growth in a dose-dependent way. The highest inhibitory effect was observed at 4 h, and Ehp-YZU-L4 showed a better inhibitory effect than Ehp-YZU-L3, with a reduction range of viable counts by 1.73–2.87 Log CFU/g for Ehp-YZU-L3 and 1.96 to 3.20 Log CFU/g for Ehp-YZU-L4. After 4 h of phage treatment, bacterial count of each group increased due to the formation of phage resistance but was still lower than the control group, suggesting their potential serving as ideal biological control agents. Compared with previously reported phages, the two phages exhibit a better antibacterial effect (Chen et al., 2022; Chen et al., 2023). Results aligned with previous results, indicating that inhibition effect has a positive correlation with phage dosage, and high phage treatment was more favorable to inactivate bacteria (Cheng et al., 2024; Yuan et al., 2025). Their lytic activity is in line with genomic features, since ORFs for holin, endolysin, and tail fiber proteins were observed in both Ehp-YZU-L3 and Ehp-YZU-L4. Phage tail fiber proteins are critical for host recognition and adsorption, and holin and endolysin can degrade bacterial peptidoglycan and form membrane pores. In the future, lytic capacity of the phage against target E. hormaechei strains in a meat matrix under practical cold-chain storage will be measured.
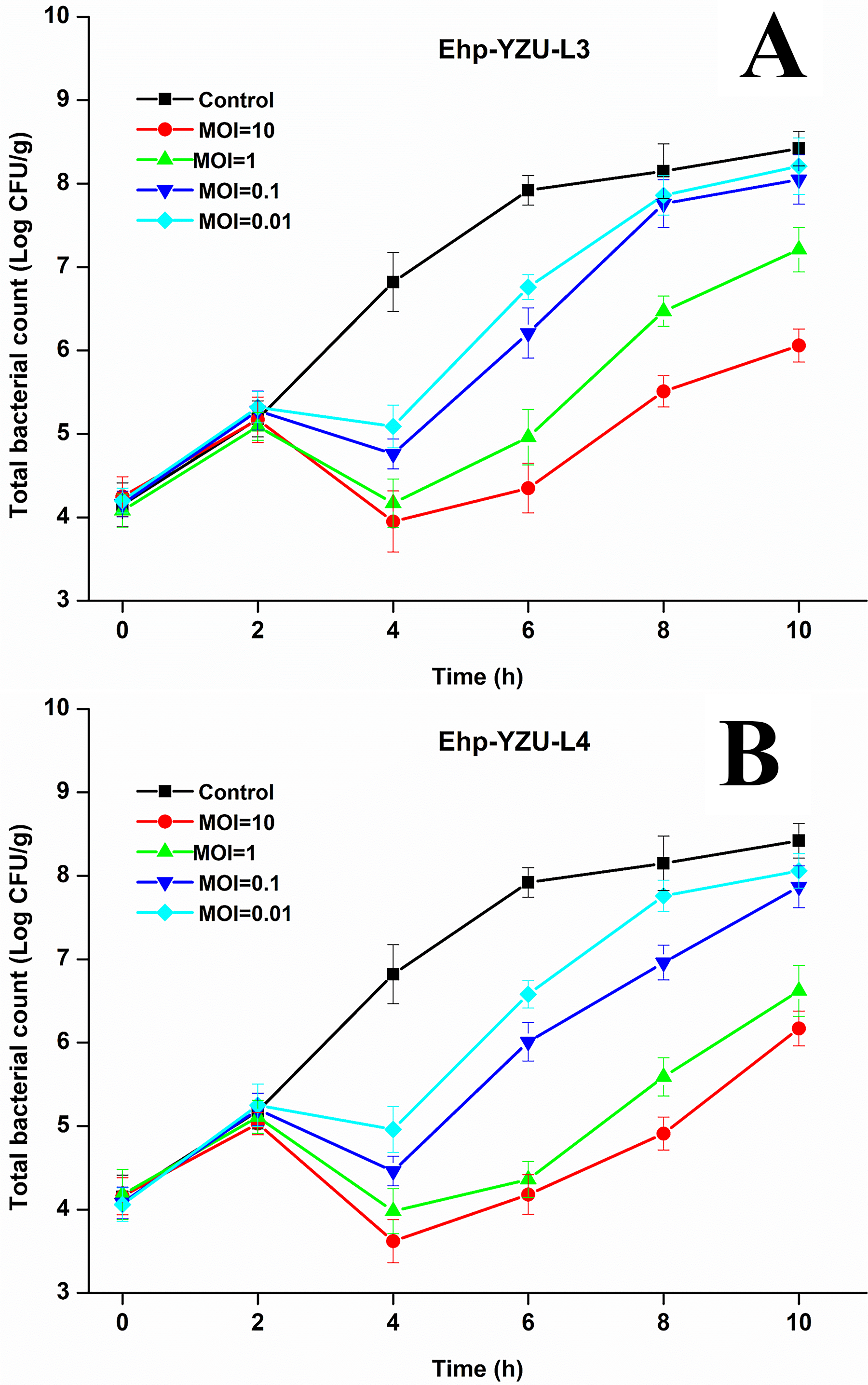
Biocontrol of E. hormaechei in pork with different MOIs by phages Ehp-YZU-L3
Despite this, inhibition on E. hormaechei in other meat products still needs to be tested. Moreover, phage treatment will not alter the visual or tactile quality of food matrices (Guo et al., 2021). However, bacterial regrowth observed in this study is likely driven by the development of phage-resistant variants (Yuan et al., 2025). To overcome this, phage cocktails are effective in enhancing antibacterial efficacy. More importantly, they possess a distinct advantage over single-phage preparations by mitigating the emergence of phage-resistant strains (Luo et al., 2025). This benefit stems from the fact that different phages within the cocktail typically target diverse bacterial receptors, making it difficult for bacteria to develop simultaneous resistance against multiple phages. Thus, further validation of the efficacy and resistance-suppressing capacity of the EHp-YZU-L3/EHp-YZU-L4 cocktail would provide valuable insights and constitutes a priority for subsequent investigations.
Conclusion
To conclude, two novel lytic E. hormaechei phages belonging to the Myoviridae family were isolated. Both phages demonstrated biological characteristics, including a short latent period, large burst size, environmental tolerance across pH 3–11 and thermal conditions up to 70°C for 20 min, and significant antibacterial efficacy in meat products. Although genomic analysis revealed a substantial proteins annotated as hypothetical proteins, the genome simultaneously contained conserved genetic elements crucial for phage life cycle and biochemical processes, but without virulence and drug resistance genes. These results suggest that phages Ehp-YZU-L3 and Ehp-YZU-L4 can be developed as novel agents for the biological control of multidrug-resistant E. hormaechei contamination and transmission in the food chain. In the future, isolation and evaluation of the phage’s broad-spectrum efficacy across diverse E. hormaechei strains strengthen the practical application value of the findings. In addition, the Ehp-YZU-L3/Ehp-YZU-L4 cocktail will be employed to recognize different bacterial receptors and expand the overall host coverage and to mitigate the emergence of phage-resistant strains in food biocontrol scenarios.
Authors’ Contributions
Conceptualization by B.L. and L.Y. Data curation by B.L., C.C., and Y.S. Formal analysis by B.L., C.C., and L.Y. Funding acquisition by L.Y. Methodology by C.C. and L.Y. Project administration by X.J. and L.Y. Software by B.L., C.C., and Y.S. Resources by X.-a.J. and L.Y. Supervision by X.-a.J. and L.Y. Validation by C.C. and Y.S. Visualization by C.C. and Y.S. Writing—original draft preparation by B.L. Writing—review and editing by L.Y.
Footnotes
Funding Information
This work was supported by the National Natural Science Foundation of China (Grants No. 32302953), China Postdoctoral Science Foundation (Grants No. 2021TQ0274, 2022M720120), the Open Project Program of Jiangsu Key Laboratory of Zoonosis (Grants No. R2504), and “QingLan” Talent Support Program of Yangzhou University.
Disclosure Statement
All authors declare that there is no conflict of interest.