
Abstract
Soy consumption is associated with thyroid disorders such as hypothyroidism, goiter, and autoimmune thyroid disease (ATD) as well as increased iodine requirement in certain cases. However, the anti-thyroid component(s) in soy are yet to be identified and the molecular mechanism(s) involved remain unclear. This study examined the effects of soy isoflavones (ISF) on iodide uptake and expression of thyroglobulin (Tg) and sodium/iodide symporter (NIS) in thyrocytes. Fischer rat thyroid cells (FRTL) were treated with Novasoy (a soy alcohol extract containing 30% ISF) or major ISF aglycones or glycosides for 24 h. Iodide uptake was measured by a colorimetric assay. The protein level of Tg and NIS was measured by Western blotting. Cytotoxicity of tested compounds was determined by the MTT cell proliferation assay. Iodide uptake in FRTL cells was dose-dependently suppressed by Novasoy added into the cell culture (10, 25, or 50 µg/mL, P < 0.05). However, neither the major ISF aglycones nor glycosides alone or in combination had similar effects. Novasoy (up to 200 µg/mL) had no cytotoxic effect. Novasoy (1, 10, and 50 µg/mL) and genistein (1 and 10 µM) markedly increased the protein content of a 40 kDa Tg fragment (P40, a known autoimmunogen) and non-glycosylated NIS in the FRTL cells (P < 0.05). Overall, this study demonstrated that the alcohol soluble component(s) other than the major ISF in soy remarkably inhibited iodide uptake in the FRTL cells. Soy ISF, particularly genistein, induced the production of P40, which might be responsible for the higher incidence of ATD reported in soy infant formula-fed children.
Introduction
Soy is a staple component of the Asian diets, and its global demand has increased dramatically in the last decade due to its potential health benefits. 1 These include lower prevalence of coronary heart disease, atherosclerosis, osteoporosis, type 2 diabetes, and even certain types of cancers. 1 Many health benefits of soy have been linked to its isoflavones (ISF) due to their hormonal, non-hormonal, and biological properties.1,2 However, excessive consumption of soy has also been associated with anti-thyroid effects such as hypothyroidism, goiters, increased iodine requirement, and autoimmune thyroid disease (ATD) notably in infants and individuals with iodine deficiency or compromised thyroid functioning.1,2 ATD represents more than 30% of organ-specific autoimmunity. Hashimoto’s disease or Hashimoto’s thyroiditis is the most common one, affecting approximately 3% of the population, and is also the most frequent cause of acquired hypothyroidism with or without goiter.3–5 Graves’ disease, on the other hand, ultimately leads to hyperthyroidism.4,5
Iodine is an essential element of thyroid hormones, triiodothyronine (T3) and thyroxine (T4), and its uptake into the thyrocytes is mediated by sodium-iodide symporter (NIS).6–9 Iodide is eventually integrated into the tyrosine residues of thyroglobulin (Tg) which is then cleaved by lysosomal proteases to produce T3 and T4. 10 In addition, the action of lysosomal proteases also produces other Tg fragments, notably a 40-kDa immunoreactive fragment known as peptide 40 (P40). P40 bears two hormonogenic sites and is shown to be associated with the onset of ATD.3,11,12
The thyroid disorders brought on from the excessive consumption of soy manifest primarily in infants fed soy formula, individuals deficient in iodide, or with compromised thyroid functioning. 2 ISF, one of the most studied soy components, however, are also believed to be linked to the anti-thyroid actions of soy. Genistein, daidzein, and glycitein are the major components of soy ISF. In soybean and the majority of Western soy products, ISF exist in their glucoside form (genistin, daidzin, and glycitin) which differ from their aglycone counterparts by the presence of a conjugated sugar molecule. Ingested ISF glucosides are converted to their bioactive aglycone forms through hydrolysis by intestinal microflora. 13 It was reported that in vivo, genistein and daidzein irreversibly inactivated thyroid peroxidase, the enzyme responsible for integrating iodine into tyrosine residues of Tg. However, addition of iodide restored its functionality, illustrating how soy raises the iodide intake requirement. 2 In vitro, genistein and daidzein were also shown to inhibit the iodination of tyrosine by competing as a substrate 2 in concentrations as low as those presented in the serum of individuals who consume soy products. 7 In CMT93 rectal and EMT6 breast cancer cells, genistein was a potent tyrosine kinase inhibitor, stimulating NIS expression and iodide uptake at 10 µM, but reducing the functional activity of NIS at high concentrations. 9
Although the anti-thyroid actions of soy have been known for decades, the bioactive components in the soy which interfere with the thyroid functions and the molecular mechanism(s) involved remain unclear. The objective of this study was, using rat thyrocytes as a model, to examine the effect of soy ISF on iodide uptake and expression of NIS and Tg.
Materials and methods
Chemicals and reagents
Fischer rat thyroid cell line (FRTL, ATCC Catalog No. CRL-1468), culture media, and MTT cell proliferation assay kit were obtained from the American Type Cell Culture collection. Newborn calf serum, trypsin-EDTA as well as penicillin, streptomycin, and glutamine mixture were purchased from GIBCO Invitrogen Corp (Grand Island, NY, USA). Bovine insulin was from Gemini Bio-Products (West Sacramento, CA, USA). Novasoy ISF concentrate was from Archer Daniels Midland Company; genistein, glycitein, and daidzein were purchased from LC Laboratories (Woburn, MA, USA). Tris, bovine TSH, insulin and transferrin, hydrocortisone, somatostatin, glycyl-
Cell culture
FRTL cells were cultured in Kaighn’s modified Ham’s F12K medium with 2 mM
Iodide uptake assay
Cells were seeded into 96-well tissue culture plates at a density of 2.5 × 105 cells/mL. After attachment, the cells were either treated with soy ISF and sodium iodide (NaI) simultaneously for 1 h, or pre-treated with ISF for 24 h prior to 1 h incubation in NaI. After incubation, plates were washed for three times to remove any extracellular NaI and ISF, and cells were lysed. NaI levels were assayed using the Sandell-Kolthoff reaction. 15 Briefly, ammonium cerium sulphate (10.5 mM) and sodium arsenite (24 mM) were added to the cell lysate and incubated for 30 min. The colour change caused by the iodide-catalysed reduction of yellow cerium (IV) to the colourless cerium (III) in the presence of sodium arsenite (III) was measured at 420 nm using a spectrophotometer (Infinite® M200 Pro, Tecan). Iodide levels were calculated against standards ranging from 0 to 300 nM NaI.
ISF treatments were carried out at varying concentrations using Novasoy, a soy alcohol extract containing 30% ISF, the aglycone form of the main ISF components found in soy (genistein, daidzein, glycitein), and their glucosides (genistin, daidzin, glycitin). Novasoy contains genistin, daidzin, and glycitin in a ratio of 1:0.95:0.13, respectively. Uptake buffer alone without NaI was used as negative control in the experiment. NaClO4, an inhibitor of iodide uptake through competitively binding to the NIS, 15 was used as a positive control. DMSO, the solvent in which the ISF were prepared, was included in the controls and its concentration was matched to the highest amount contained in the ISF treatment. Cell proliferation was measured using the MTT cell proliferation assay kit.
Western blot analysis
Cells seeded at 2 × 106 cells/60 mm dish were treated with genistein (1, 10, and 100 µM) or Novasoy (1, 10, and 50 µg/mL) for 24 h, and then lysed in Frack’s buffer (20% glycerol, 20 mM Tris, pH 7.5, 150 mM NaCl, 2 mM EDTA and 1% Triton X-100) containing a Complete Mini protease inhibitor cocktail (Roche). Cell lysates were sonicated using a Branson Sonifier 250 and then centrifuged at 15,000 × g, 4℃ for 20 min. The supernatant was collected and the protein concentration was determined using the Bio-Rad DC protein assay kit in accordance with the manufacturer’s instructions. Western blotting was performed as described previously. 16 Total protein samples (20–80 µg) were mixed with loading buffer, resolved by 12% SDS-PAGE (110 V, 2 h), and then electrotransferred (50 V, 1 h) onto nitrocellulose membranes. After blocking with skim milk powder (5%) in Tris-buffered saline (10 mM Tris pH 8.0, 150 mM NaCl; TBS) and Tween-20 (0.05%; TBST-T), membranes were incubated overnight with primary antibody (1:2000) and subsequently with HRP-conjugated secondary antibody (1:10,000). Immunoreactivity was detected by chemiluminescence autoradiography using the SuperSignal West Dura kit according to the manufacturer’s instructions and the images were scanned. All membranes were reprobed with β-tubulin antibody (1:5000). The intensities of the protein bands of interest and the β-tubulin band were determined densitometrically using Scion Image. The intensity of the target proteins was normalized by the respective β-tubulin intensity.
Statistical analysis
Results are expressed as means ± SEM. A probability of P < 0.05 was considered to be significant. Effects of ISF treatments on FRTL cells were analysed by one-way ANOVA. Differences between individual means were determined by Fisher least significant difference post-hoc test. Data were analysed using Statistica Version 8.1 (StatSoft).
Results
Iodide uptake in FRTL cells exposed to ISF
Incubation of the FRTL cells with 10 µM NaI significantly increased iodide uptake compared to control (UB) (P < 0.01). NaClO4 (50 µM), a known competitive inhibitor of iodide uptake, completely blocked the NaI-induced increase in the iodide uptake (P < 0.01). Addition of Novasoy at higher levels (50–200 µg/mL) into the FRTL cells during the incubation with 10 µM NaI dose-dependently suppressed iodide accumulation compared to DMSO control (P < 0.05, Figure 1a). Genistein slightly reduced the accumulation of iodide in the cells at certain levels (Figure 1b), whereas low level of daidzein and glycitein (1 µM) increased iodide contents in the cells (P < 0.05, Figure 1c and d).
Effects of Novasoy (a soy alcohol extract containing 30% isoflavones, ISF) and major isoflavone aglycones on iodide uptake in FRTL cells. Iodide uptake assay was conducted in the FRTL cells (5 × 104/well of 96-well plate) after incubated in the absence (uptake buffer, UB) or presence of 10 µM NaI with 50 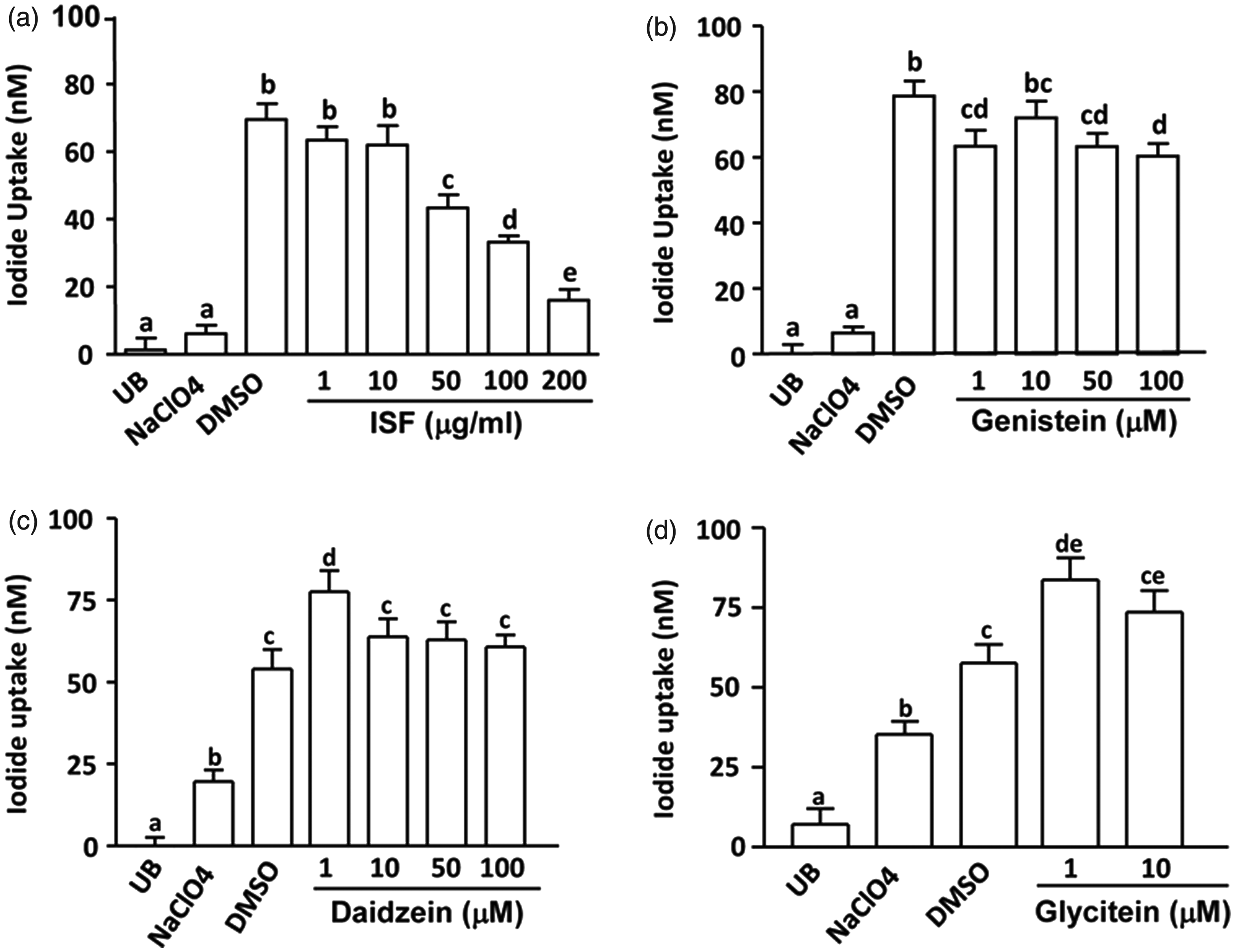
Pre-treatment of FRTL cells with increasing amounts of Novasoy (10, 25, and 50 µg/mL) for 24 h markedly reduced the iodide uptake compared to DMSO control (P < 0.01, Figure 2a). Genistein or daidzein (10, 50 and 100 µM) alone (Figure 2b and c) or a combination of genistein (25 µM), daidzein (25 µM), and glycitein (2.5 µM) (Figure 2e) increased iodide accumulation (P < 0.05); however, glycitein alone (Figure 2d) or a combination of genistin (25 µM), daidzin (25 µM), and glycitin (2.5 µM) (Figure. 2e) had no effects on iodide uptake.
Effects of pretreatment with Novasoy (a soy alcohol extract containing 30% isoflavones, ISF) and major isoflavone aglycones or glucosides on iodide uptake in FRTL cells. FRTL cells (5 × 104/well of 96-well plate) were pretreated with DMSO (vehicle), NaClO4 (50 µM) or increasing amounts of (a) ISF (1, 10, 25, 50 µg/mL), (b) genistein (1, 10, 50, 100 µM), (c) daidzein (1, 10, 50, 100 µM), (d) glycitein (1, 2.5, 5, 10 µM), (e) ISF (50 µg/mL), GDG-a (25 µM genistein, 25 µM daidzein, and 2.5 µM glycitein) or GDG-g (25 µM genistin, 25 µM daidzin, and 2.5 µM glycitin) for 24 h before the iodide uptake assay was conducted. The values represent means ± SEM of four independent experiments performed in five replicates. Means with different letters differ, P < 0.05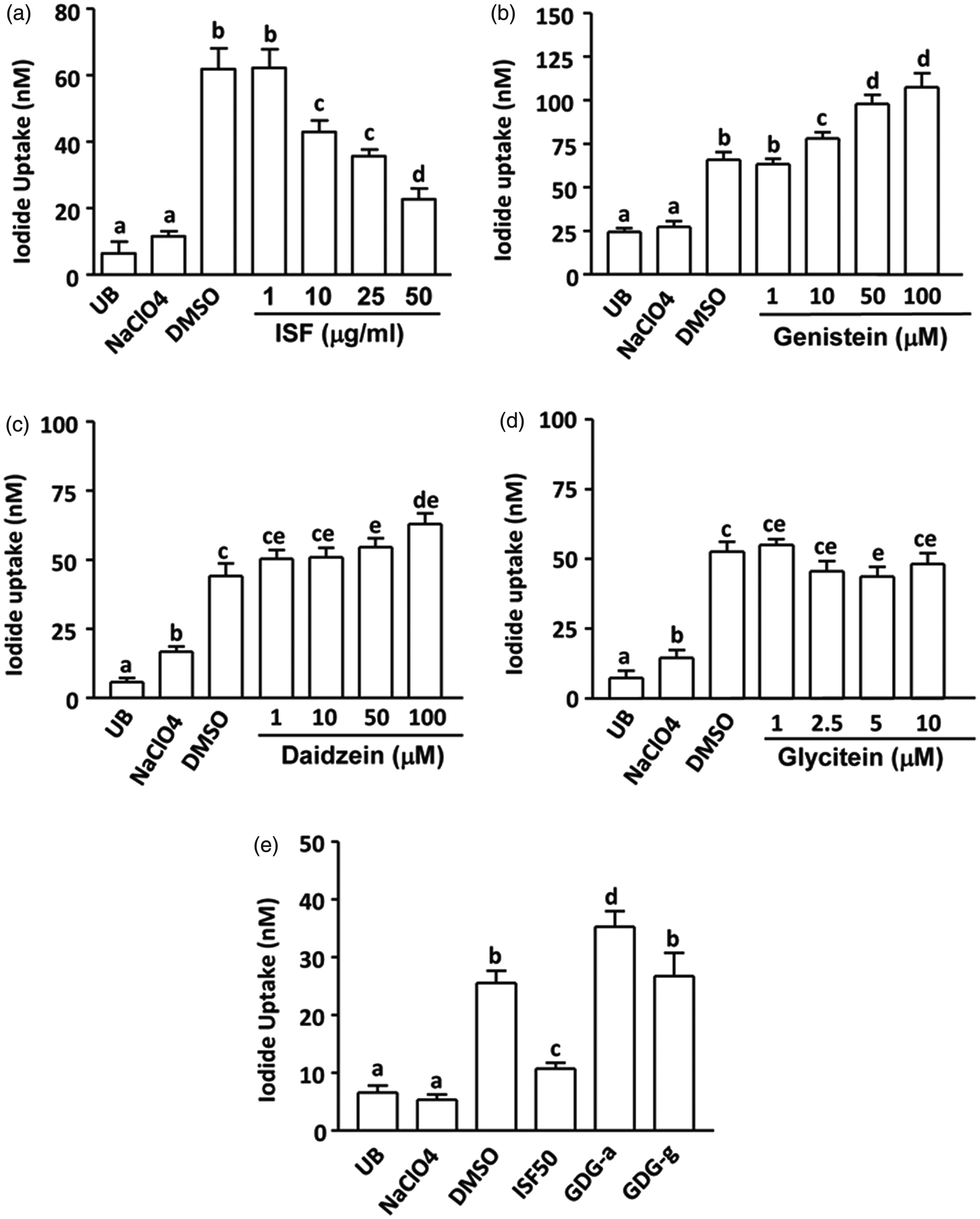
Effects of NaI levels on inhibition of iodide uptake by Novasoy
Soy consumption increased iodine requirement in order to maintain normal thyroid function as previously reported in both animals and humans. This experiment examined whether the levels of iodine added affect the effect of Novasoy. The results showed that percentages of inhibition of iodide uptake by Novasoy (50 µg/mL) were significantly lower in the thyrocytes supplied with 20 and 40 µM NaI than in those with 10 µM NaI (P < 0.05, Figure 3).
Effects of NaI levels added on the inhibitory action of Novasoy on iodide uptake in FRTL cells. FRTL cells were treated with or without ISF (50 µg/mL) for 24 h and then added with increasing amounts of NaI (10, 20, 40 µM) for iodide uptake. The percentage of iodide uptake inhibition by ISF was calculated. The values represent means ± SEM of three independent experiments conducted in five replicates. Means with different letters differ, P < 0.05
Cell proliferation
No ISF treatments were shown to be cytotoxic except for 10 μM glycitein, which showed slight suppression of cell proliferation (Figure 4b). Genistein (50 and 100 μM) and 200 µg/mL Novasoy even increased cell proliferation (Figure 4a, b). The combination of ISF aglycones (GDG-a) significantly increased cell proliferation (Figure 4a).
Effects of Novasoy (a soy alcohol extract containing 30% isoflavones, ISF), major soy isoflavone aglycones and glucosides on FRTL cell proliferation. FRTL cells (5 × 104/well of 96-well plate) were pretreated with (a) DMSO (vehicle), or genistein (10, 50, 100 μM)), Daidzein (10, 50, 100 µM) or GDG (50 µM genistein, 50 µM daidzein, and 10 µM glycitein), or (b) DMSO, glycitein (2.5, 5, 10 µM), ISF (25, 50, 100, 200 μg/mL) for 24 h. Cell proliferation was expressed as percentage of DMSO (vehicle). The values represent means ± SEM of three independent experiments conducted in five replicates. Means with different letters differ, P < 0.05
Effect of Novasoy and genistein on NIS expression
Two forms of NIS, non-glycosylated (∼50 kDa) and glycosylated (∼100 kDa), were detected in the cultured cells (Figure 5). While the content of glycosylated NIS was unchanged by genistein and Novasoy treatments, the non-glycosylated NIS content showed significant increase at 10 µM and then returned to control level at 100 µM. Cells exposed to Novasoy showed an elevated non-glycosylated NIS content at 1 µg/mL but gradually decreased to control level at 50 µg/mL.
Effects of Novasoy (a soy alcohol extract containing 30% isoflavones, ISF) and genistein on glycosylated (a) and unglycosylated (b) sodium iodide symporter (NIS) protein content in FRTL cells. FRTL cells (2 × 106 cells/60 mm dish) were treated with DMSO (vehicle), genistein (1, 10, or 100 μM) or ISF (1, 10, or 50 µg/mL) for 24 h. The values represent means ± SEM of three independent experiments conducted in triplicate. Means with different letters differ, P < 0.05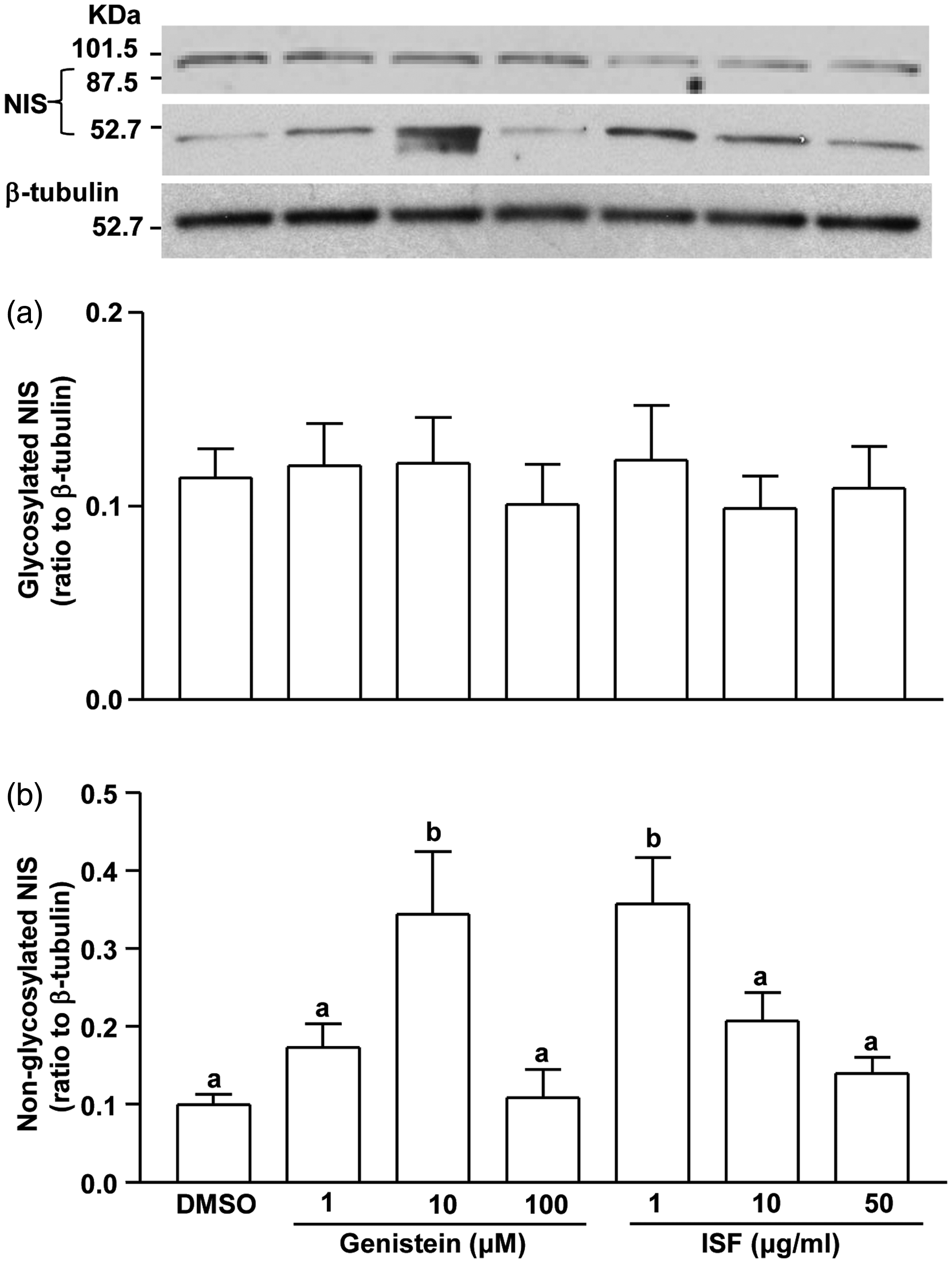
Effect of Novasoy and genistein on Tg expression
Two forms of Tg proteins, intact Tg (∼330 kDa, P330) and cleaved Tg fragment (∼40 kDa, P40), were detected in the FRTL cells. Treatment with genistein (10 µM) and Novasoy (10 µg/mL) significantly increased P330 content (P < 0.05, Figure 6a). P40 protein was markedly increased in the cells treated with genistein (1, 10 µM) or Novasoy (10, 50 µg/mL) (P < 0.05, Figure 6b). The contents of both P330 and P40 in the cells treated with 100 µM genistein were not different from those of control (P > 0.05, Figure 6a and b).
Effects of Novasoy (a soy alcohol extract containing 30% isoflavones, ISF) and genistein on thyroglobulin (Tg, 330 kDa) (a) and Tg fragment (40 kDa) (b) content in FRTL cells. FRTL cells (2 × 106 cells/60 mm dish) were treated with DMSO (vehicle), genistein (1, 10, or 100 μM), or ISF (1, 10, or 50 µg/mL) for 24 h. The values represent means ± SEM of three independent experiments conducted in triplicate. Means with different letters differ, P < 0.05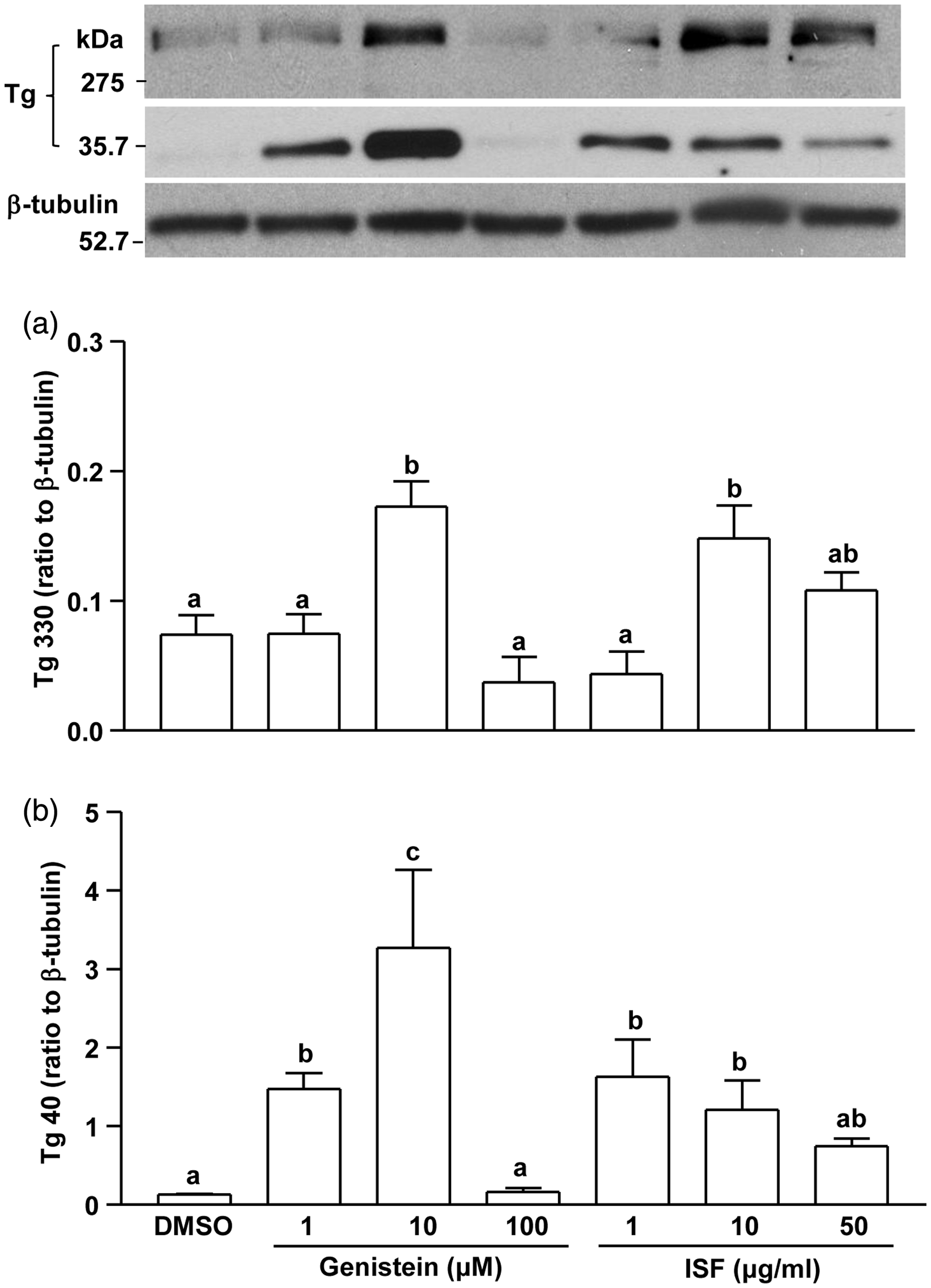
Discussion
Addition of Novasoy into iodide uptake assay dose-dependently decreased iodide content in the FRTL cells. However, the major soy ISF aglycones, genistein, daidzein, and glycitein did not have similar effects. Although genistein alone caused a minor inhibition on iodide uptake, treatments with daidzein and glycitein had no inhibitory effects, which could not explain the uptake inhibition observed with Novasoy. Novasoy is a soy alcohol extract and contains about 30% ISF. This indicated that other alcohol soluble component(s) rather than the major ISF may be responsible for the anti-thyroid actions. Many minor soy phytoestrogens such as coumestrol, formononetin, biochanin A, lignans, and polyphenolic compounds saponins as well as alcohol soluble carbohydrates are contained in Novasoy and their potential antithyroid effects remain to be examined. The possible cellular mechanism(s) involved in the suppression of iodide uptake could be (a) competition of the alcohol soluble component(s) with iodine for binding to NIS, a transmembrane protein that mediates the transport of iodide into the thyrocytes; (b) stimulation of post-translational modifications of the NIS thereby reducing iodide binding affinity or transporting efficiency; and (c) increased efflux of intracellular iodide.
Pre-treatment of the FRTL cells with Novasoy for 24 h significantly inhibited iodide uptake. However, this did not correlate with non-glycosylated NIS content initially increased above control levels at 1 µg/mL and then returned to control levels at 50 µg/mL. Since Novasoy also inhibited iodide uptake during short term exposures, its primary target appears to be NIS activity rather than the content. Interestingly, genistein showed a dose-dependent effect on NIS content. It remarkably increased non-glycolsylated NIS content at 10 µM, while higher level (100 µM) caused a return to control levels. The iodide uptake assay for the same exposure showed, however, dose-dependent increase in iodide abundance from 1 to 100 µM genistein. Although the molecular mechanism(s) involved in the regulation of NIS protein expression by genistein and Novasoy is unclear, it has been shown that genistein significantly increased the mRNA abundance of NIS in thyroid cancer cell line FTC-133, 17 murine rectal (CMT93), and breast (EMT6) cancer cells. 9 Therefore, the soy extract and genistein may be useful in sensitizing the thyroid tumors which is lack of NIS expression to radioiodide therapy. Thyroid expression of NIS and Tg is controlled by transcription factors Nkx2.1/TITF1, FoxE1/TITF2, and Pax8. Particularly Pax8 is believed to play an even more important role.18–20 Which transcription factor(s) is involved in the regulation of NIS and Tg by genistein and Novasoy remain to be investigated.
Dose-dependent effects of genistein have been previously reported in other cell types such as cardiomyocytes, adipocytes, breast cancer cells, osteoblasts, and osteoprogenitor cells.21–24At lower concentrations (1–10 µM), genistein exerted estrogenic effects while at higher concentrations (>10 µM), anti-estrogenic and tyrosine kinase inhibitory effects were observed.25,26 In some cases, the cell activity was stimulated at one concentration and comparatively depressed at the other. For example, in cardiomyocytes, low concentrations of genistein promoted the production of anti-apoptotic proteins while high concentrations induced apoptosis by up-regulating apoptotic proteins. 21 A similar phenomenon was observed in this study, where 1–10 µM genistein boosted thyrocyte functioning by stimulating the expression of NIS, and 100 µM caused a return to control levels. However, the dose-dependent accumulation of iodide in the cells responding to genistein treatment was inconsistent with NIS protein content, suggesting that the other cellular mechanism(s) such as the NIS activities including binding affinity and transporting efficiency may play a role in this regard. Conversely, addition of increasing amounts of genistein into the FRTL cells during the iodide uptake assay slightly attenuated the iodide uptake, indicating existence of potential competition of genistein with iodine molecule for NIS binding.
Treatment of the thyrocytes with the mixtures of ISF glucosides or aglycones at similar levels and ratio as contained in the Novasoy failed to suppress iodide uptake. Although minor decreases were observed at low dose of genistin or glycitin alone (0.1 µM, data not shown), neither could match the degree of iodide uptake down-regulation observed in exposures to Novasoy. In contrast, the combined aglycones were able to significantly up-regulated iodide uptake due to enhanced cell proliferation. A quadruple increase in NaI reduced the inhibition of Novasoy on iodide uptake by 25%, which is consistent with other studies showing that the anti-thyroid actions of soy can be reversed by increasing iodine intake or supplementation.27–30 This also might be the reason why excessive soy consumption doubled the iodine requirements to maintain the normal thyroid functions as reported in animal and human studies.27,31,32
Additionally, treatment of FRTL cells with genistein significantly increased the content of P40, an identified autoimmunogen that might be responsible for the onset of ATD.3,5,11,12,33 The content of the intact Tg (P330) was increased in parallel with P40 by the same genistein and Novasoy treatments. Novasoy also increased, although to a lesser extent, the content of P330 and P40 Tg fragments that may be attributed to the action of the genistein contained. Various studies showed that the amount of cleaved Tg peptides increased in proportion to the iodoamino acid content of Tg molecules.3,33 Moreover, higher thyroid hormone synthesis and iodination were associated with generation of excessive reactive oxygen species, resulting in the oxidative cleavage of Tg to produce P40 and expose its cryptic epitopes.11,12 Increased production of reactive oxygen species also induces cell necrosis which in turn causes an inflammatory response that triggers thyrocytes to internalize P40 and present them to T-cells. This initiates an autoimmune response against Tg damaging thyroid functioning and causing ATD.3,11,12 Conversely, high dose of genistein (100 μM) decreased Tg fragmentation; this could be attributed to the antioxidant properties of genistein including the reduction of free radical production and thus oxidative cleavage12,34 or inhibition of other cellular events such as tyrosine kinase activation because genistein is a potent tyrosine kinase inhibitor. 35
Increased content of Tg induced by genistein and Novasoy does not necessarily indicate higher thyroid hormone production since release and diffusion of thyroid hormones from Tg and out of the thyrocytes are determinant steps of the hormone synthesis. Furthermore, soy has been shown to increase thyroid hormone production in several studies, although not consistently and not to a high extent.16,36–38 At least four possible mechanisms have been proposed to explain the potential anti-thyroid actions of soy. First, the excessive amount of phytates present in soy could bind to T3 in the lumen and increase its loss in feces, and therefore reduced the efficacy of oral thyroid hormones by interfering with the uptake of exogenous thyroid hormones. 39 Additionally, soy ISF interfere with DNA binding to thyroid hormone receptors because of their structural similarity to T3.40–42 Third, ISF may affect T4 binding to plasma protein. 43 Finally, ISF can reduce the deiodination of T4 to produce T3 which is catalysed by 5’-deiodinase type I. 43
In conclusion, this study demonstrated for the first time, to the best of our knowledge, that alcohol soluble component(s) in soy other than the major ISF dose-dependently inhibited iodide uptake in FRTL cells. However, the bioactive component(s) remain to be identified. Soy alcohol extract, specifically genistein at concentrations comparable to physiological levels in infants fed soy formulas stimulated the production of Tg and a Tg fragment P40 (a known autoimmunogen), which might be responsible for the higher incidence of autoimmune thyroid diseases reported in soy infant formula-fed children.
Footnotes
Author contributions
LT and MH conducted the experiments, data analysis, and wrote the manuscript. CW provided technical assistance and review of the manuscript. CWX designed the experiments, interpreted the studies, and wrote the manuscript.
Acknowledgment
This research was funded by Health Canada.