
Abstract
This study aimed to investigate the role of CD147 in the progression of gastric cancer and the signalling pathway involved in CD147-mediated gastric cancer cell line SGC7901 proliferation and invasion. Short hairpin RNA (shRNA) expression vectors targeting CD147 were constructed to silence CD147, and the expression of CD147 was monitored by quantitative realtime reverse transcriptase polymerase chain reaction and Western blot and further confirmed by immunohistochemistry in vivo. Cell proliferation was determined by Cell Counting Kit-8 assay, the activities of matrix metalloproteinase (MMP)-2 and MMP-9 were determined by gelatin zymography, and the invasion of SGC7901 was determined by invasion assay. The phosphorylation and non-phosphorylation of the mitogen-activated protein kinases, extracellular signal-regulated kinase1/2 (ERK1/2), P38 and c-Jun NH2-terminal kinase were examined by Western blot. Additionally, the ERK1/2 inhibitor U0126 were used to confirm the signalling pathway involved in CD147-mediated SGC7901 progression. The BALB/c nude mice were used to study tumour progression in vivo. The results revealed that CD147 silencing inhibited the proliferation and invasion of SGC7901 cells, and down-regulated the activities of MMP-2 and MMP-9 and the phosphorylation of the ERK1/2 in SGC7901 cells. ERK1/2 inhibitor U0126 decreased the proliferation, and invasion of SGC7901 cells, and down-regulated the MMP-2 and MMP-9 activities. In a nude mouse model of subcutaneous xenografts, the tumour volume was significantly smaller in the SGC7901/shRNA group compared to the SGC7901 and SGC7901/snc-RNA group. Immunohistochemistry analysis showed that CD147 and p-ERK1/2 protein expressions were down-regulated in the SGC7901/shRNA2 group compared to the SGC7901 and SGC7901/snc-RNA group. These results suggest that ERK1/2 pathway involves in CD147-mediated gastric cancer growth and invasion. These findings further highlight the importance of CD147 in cancer progression, indicating that CD147 would be an attractive therapeutic target for gastric cancer.
Keywords
Introduction
Gastric cancer is the fourth most common cancer worldwide, with approximately 21,320 new cases of gastric cancer are diagnosed each year in the USA 1 and ∼990,000 worldwide, and it is the second most common cause of cancer death, with 738,069 deaths annually worldwide. 2 Despite much research, we do not yet have drugs adequate for treating gastric cancer. A better understanding of the molecular mechanisms driving gastric tumourigenesis is crucial and will provide enormous benefits in developing new therapeutic treatments for gastric cancer.
Extracellular matrix metalloproteinase inducer, also known as CD147, is a heavily glycosylated type I transmembrane glycoprotein and belonging to the immunoglobulin (Ig) superfamily. It contains two C2-like immunoglobulin extracellular domains, a transmembrane domain and a cytoplasmic domain. 3 The presence of a highly conserved acidic residue (glutamic acid) and leucine zipper-like sequences within the hydrophobic sequence of the transmembrane domain suggests that intramembrane associations are likely to occur with other membrane proteins. 4 CD147 is expressed in multifarious forms, including high glycosylated (HG 45–65 kDa) and low glycosylated (LG 32–44 kDa) forms as well as the native 27-kDa protein. It is usually expressed at low levels in most normal tissues, but is highly upregulated during dynamic cellular events, such as tissue remodelling and cancer progression.4–6 CD147 is highly expressed on the surface of various tumours, including gastric cancer.4,7–11
Matrix metalloproteinases (MMPs), a family of zinc-dependent proteolytic enzymes, play a central role in the degradation of the extracellular matrix. MMPs expression allows degradation of the basement membrane that are believed to affect tumour angiogenesis, lymphangiogenesis, tumour growth, local invasion and subsequent distant metastasis. 12 CD147 induces several MMPs, especially MMP-2, MMP-9 in stromal fibroblasts and endothelial cells as well as in tumour cells themselves.13,14 Evidence indicates that the N-terminal Ig domain is required for stimulation of fibroblast MMPs production by CD147.14,15
Mitogen-activated protein kinases (MAPKs) are serine/threonine kinases activated in response to a variety of external signals. Three major subclasses of MAPK superfamily, namely, extracellular signal-regulated kinase1/2 (ERK1/2), c-Jun NH2-terminal kinase (JNK) and P38 have been identified. ERK is one of a crucial mediator in Raf/MEK/ERK signalling axis maintaining diverse cellular function including cell proliferation, apoptosis as well as migration. 16 Constitutive activation of ERK signalling is usually linked with neoplastic transformation such as uncontrolled cell growth by stimulating cell survival pathway and increasing cell motility.17–20 Many studies have shown that upregulation of MAPK signal transduction pathways may play an important role in tumourigenesis and metastatic potential of gastric cancer.21–24 In addition, Both Pathway Studio and STRING analysis showed phosphoproteins can be functionally categorized into EGFR-MAPK1/3 (ERK1/2) cascades in gastric cancer cells. 25
Until recently, signalling events downstream of CD147-mediated tumourigenesis and metastasis have been studied in several cancers, but have not yet been fully elucidated in gastric cancer. Evidence showed that CD147 stimulate cell survival pathway signalling, including phosphorylation of ERK, SPAK/JNK in hepatocellular carcinoma cells.26,27 We hypothesized that CD147 regulated MAPK signalling pathways in gastric cancer, the aberrant activation of which has been implicated in cancer growth, metastasis and invasion. In the present study, we demonstrated that CD147 induced gastric cancer growth and invasion through the ERK1/2 pathway.
Materials and methods
Reagents and cells
The vector pSilencer 3.1-H1 neo was purchased from Ambion Inc. (Austin, TX, USA). G418 was purchased from Gibco BRL (Grand Island, NY, USA). U0126 (specific ERK1/2 inhibitor), p-P38, P38, p-ERK1/2, ERK1/2, p-JNK and JNK-specific antibodies were purchased from Cell Signaling Technology (Beverley, MA, USA). CD147- and β-actin-specific antibodies were purchased from Santa Cruz (CA, USA). Human gastric cancer cell line SGC7901 was provided by Digestive Department of Jiangsu Province Hospital, China.
Cell culture
Cells were cultured in Dulbecco's modified eagle medium (DMEM) medium (Gibco BRL) supplemented with 10% newborn calf serum (Gibco BRL) in a 5% CO2 humidified atmosphere at 37℃.
Construction of shRNA expression vectors
The sequences of the designed CD147-specific shRNAs

Transfection of cells and generation of stable cell clones
SGC7901 cells were plated in six-well plates at a density of 3 × 105 cells per well and incubated overnight. Cells were transfected with pSilencer-CD147-shRNA1, pSilencer-CD147-shRNA2 and pSilencer-sncRNA, respectively, using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. Forty-eight hours after transfection, SGC7901 cells were diluted to 1:10 for passage and neomycin resistance clones were selected in the medium containing 400 µg/mL G418 for two weeks. The positive clones were picked and expanded to establish cell lines. The stable transfection cell clones, designated as SGC7901/shRNA1, SGC7901/shRNA2 and SGC7901/snc-RNA, were verified by quantitative realtime reverse transcriptase polymerase chain reaction (RT-PCR) and Western blot analysis.
Quantitative realtime RT-PCR assays
Total cellular RNA was extracted using TRIzol reagent (Invitrogen). RNA quantification was performed using spectrophotometry. The primer sequences used were for CD147: (sense) 5'-CCATGCTGGTCTGCAAGTCAG-3' and (antisense) 5'-CCGTTCATGAGGGCCTTGTC-3'; β-actin (sense) 5'-CTGGAACGGTGAAGGTGACA-3' and (antisense) 5'-AAGGGACTTCCTGTAACAACGCA-3'. The mRNA level for CD147 was analyzed by one-step realtime RT-PCR with RNA-direct™ SYBR Green Realtime PCR MasterMix (Toyobo Co., Ltd., Osaka, Japan) according to the manufacturer's instructions. The cycling conditions were as follows: 90℃ for 30 s, 61℃ for 20 min, 95℃ for 60 s, then 40 cycles at 95℃ for 15 s, 60℃ for 1 min. Crossing threshold (Ct) values of the CD147 gene were normalized to β-actin, and then the data were calculated and analyzed by a comparative 2−ΔΔ Ct method (ΔCt = Cttarget − Ctβ-actin). 29 The amplification was monitored on an ABI prism 7500 realtime PCR apparatus (Applied Biosystems, USA). Each sample was prepared in triplicate, and all reactions were triplicated independently to ensure the reproducibility of the results.
Western blot analysis of CD147, p-P38, P38, p-ERK1/2, ERK1/2, p-JNK, JNK MAP kinases
Cells were lysed with ice-cold lysis buffer (50 mmol/L Tris-HCl, pH 7.4, 150 mmol/L NaCl, 1 mmol/L MgCl2, 100 µg/mL phenylmethanesulphonyl fluoride and 1% Triton X-100) for 30 min on ice. Protein concentrations were determined by bicinchoninic acid (BCA) protein assay. Proteins were denatured in 5x sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) sample buffer for 5 min at 100℃. Equal amounts (30 µg) of lysate proteins were separated on 10% SDS-PAGE gels, and transblotted onto polyvinylidene difluoride membranes (Pall Corporation, USA). After blocking with 5% non-fat dry milk in TBST buffer (10 mmol/L Tris, pH 7.5, 150 mmol/L NaCl, and 0.05% Tween 20) for 2 h at room temperature, membranes were incubated with specific antibodies against CD147(1:500), β-actin (1:1000), p-P38 and P38 (1:1000), p-ERK and ERK (1:1000), p-JNK and JNK (1:1000) in dilution buffer (5% BSA in TBS) overnight at 4℃. Followed by incubation in a 1:3000 dilution of secondary antibodies conjugated to horseradish peroxidase (Santa Cruz) for 2 h at room temperature. Protein bands were detected using ECL detection system (Boster, Wuhan, China). All of the Western blots were performed at least three times.
Gelatin zymography
Protein concentrations in conditioned medium were determined using the BCA method (BCA kit; Pierce, Rockford, IL, USA). The gelatinolytic activities of mmP-2 and mmP-9 in the conditioned medium were assayed by electrophoresis on 10% polyacrylamide gels containing 1 mg/mL of gelatin (type A, Sigma, St Louis, MO, USA) at 4℃. PAGE gels were run at 120 V, washed in 2.5% Triton X-100 for 1 h, and then incubated for 20 h at 37℃ in activation buffer (50 mmol/L Tris-HCl, pH 7.5, 5 mm/L CaCl2, 0.02% Brij-35). After staining with Coomassie Blue (10% glacial acetic acid, 30% methanol and 0.5%Coomassie Blue) for 3 h, the gel was destained with a solution of 10% glacial acetic acid, and 50% methanol without Coomassie Blue for 30 min. White lysis zones indicating gelatin degradation were revealed by staining with Coomassie blue R-250. The mmP-2 and mmP-9 activities were also investigated after serum-starved SGC7901 cells were pretreated with U0126 (5 µmol/L) for 2 h by gelatin zymograph.
Cell proliferation assay
Before the cell proliferation assay, trypan blue exclusion test of cell viability was performed and the viability of the three groups of cells (SGC7901, SGC7901/snc-RNA and SGC7901/shRNA2) was >98%. Cell proliferation in vitro was analyzed using Cell Counting Kit-8 (CCK-8; Sigma) according to the manufacturer’s protocol. Briefly, 1 × 104 cells of each group were plated per well in three 96-well microplates in 100 µL of medium. After 24, 48 and 72 h of culture, respectively, the CCK-8 solution (10 µL) was added to each well and the plates were returned to standard tissue incubator conditions for an additional 1.5 h. Absorbance at 450 nm was measured using a microplate reader. The inhibition rate was calculated as (1 − [optical density (OD) value of the transfectant/OD value of untreated SGC7901]) × 100%. The proliferation assay was also performed to examine the inhibitory effects of U0126 on cell proliferation. Cells were trypsinized and seeded at a density of approximately 1 × 104 cells/well in 96-well cell culture plates in 100 µL culture medium with 10% fetal bovine serum (FBS). After 24 h, the medium was refreshed with 1% FBS containing U0126 with a series of concentration for 24, 48 and 72 h, respectively. Each experiment was done in triplicate.
Invasion assay
Appropriate Matrigel (BD Biosciences, Bedford, MA, USA) was added to the upper chamber of the transwell apparatus with 8-µm pore size membrane (Costar, Cambridge, MA, USA). After the Matrigel solidified at 37℃, serum-free DMEM containing 1 × 105 cells in 200 µL was added into the upper chamber; the lower chamber received 600 µL of 10% FBS-containing medium. After incubated at 37℃ for 24 h, membranes coated with Matrigel were swabbed with a cotton swab and fixed with 100% methanol for 20 min. The membranes with cells were soaked in 0.1% crystal violet for 20 min and then washed with distilled water. The number of cells attached to the lower surface of the polycarbonate filter was counted at × 200 magnification under a light microscope. The invasion assay was also performed after SGC7901 cells were treated with U0126 in a concentration of 72 h half maximal inhibitory concentration (IC50) of CCK-8 assay. After trypsinization, 1 × 105 cells in 200 µL were incubated with U0126 (4 µmol/L). There are no significant inhibition on cell proliferation within 24 h in the concentration (Figure 2b). Results were expressed as mean of triplicate experiments.
In vivo tumour progression assay
All protocols involving animals were reviewed and approved by the ethics review committee of the Animal Experimental Center of Nanjing First Hospital. We used 4- to 6-week-old BALB/c nude mice (Center for Comparative Medical Research of Yangzhou University, Yangzhou, Jiangsu, China). Tumour cells harvested with trypsin, washed with DMEM, and resuspended in serum-free DMEM were subcutaneously inoculated (1.5 × 106/0.15 mL) into the left axillary fossa. The size of the transplanted tumours was measured every seven days and the tumour volume was calculated using the formula V = 1/2 × (L × W2). The mice were sacrificed at 42 days post-inoculation. Harvested tissues were fixed in 4% buffered formalin, embedded in paraffin, sectioned at 4 µm, and stained with H&E and immunohistochemistry analysis was used to detect CD147 and p-ERK1/2 protein expressions. CD147 (1:50 dilution, Santa Cruz Biotechnology) and p-ERK1/2 polyclonal antibody (1:200 dilution, Cell Signaling Technology) were used in standard indirect immunoperoxidase procedures.
Statistical analysis
SPSS13.0 software was used. Each assay was performed at least three times. The data were expressed as mean ± SD, and Student's t-test was used to determine the significance of differences in multiple comparisons. P < 0.05 was considered to be statistically significant.
Results
Specific siRNA inhibits the expression of CD147 in SGC7901 cells
The knock-down efficiencies of different CD147-specific shRNAs in SGC7901 cells were evaluated using realtime qPCR and Western blot. As show in Figure 1, pSilencer-CD147-shRNA2 effectively inhibited expression of CD147 in tumour cells, while snc-RNA showed no effects (P < 0.05). Western blot analysis confirmed the down-regulation of CD147 protein by pSilencer-CD147-shRNA2 (P < 0.05; Figure 1b). Thus, SGC7901/siRNA2 cell clone was chosen for further experiments.
CD147-specific shRNAs results in the reduction of CD147 mRNA and protein levels in SGC7901 cells. (a) Relative mRNA levels were analysed by quantitative RT-PCR. β-actin was used as normalization control. *P < 0.05 compared with SGC7901. (b) Western blot analysis of CD147 protein expressions. β-actin was used as loading control. HG: high glycosylated form; LG: low glycosylated form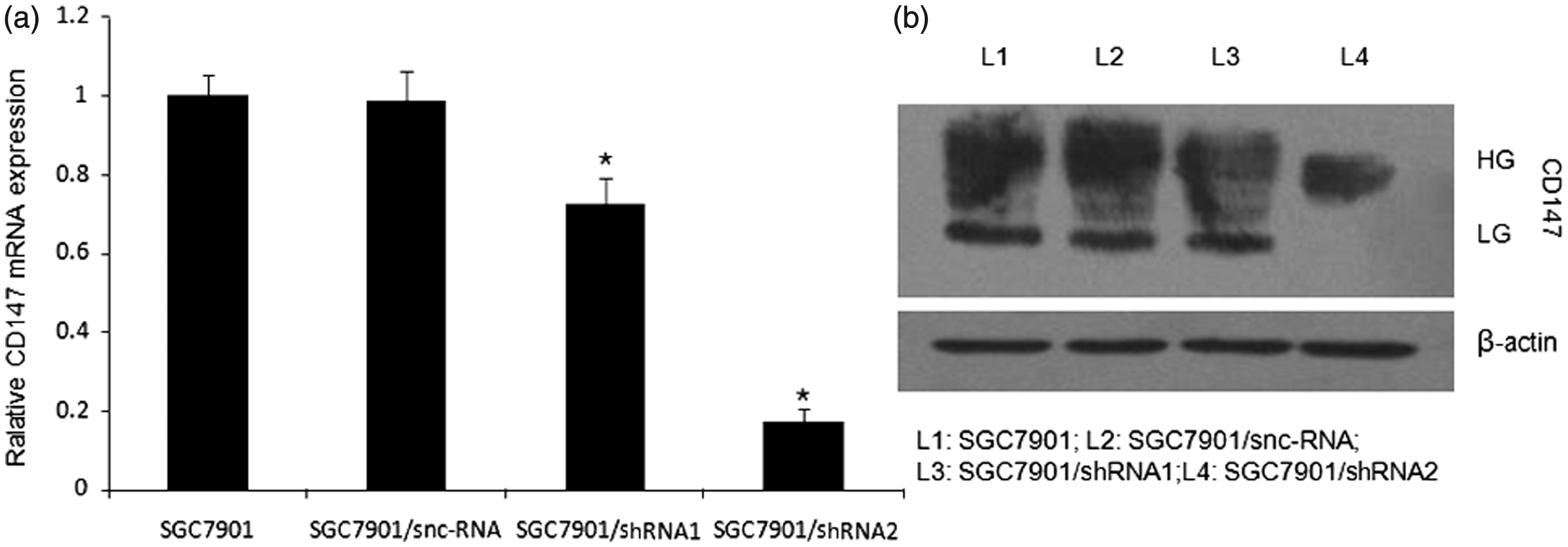
CD147 silencing reduces the proliferation of SGC7901 cells
We determined the proliferation of SGC7901, SGC7901/snc-RNA and SGC7901/shRNA2, respectively. As shown in Figure 2(a), compared with SGC7901, the proliferation of SGC7901/shRNA2 was inhibited to 74.85% (P < 0.05), 77.86% (P < 0.05) and 74.79% (P < 0.05) at 24, 48 and 72 h, respectively. There was no significant difference between SGC7901/snc-RNA and SGC7901 (P > 0.05).
Decrease in the proliferation potential of SGC7901 cells transfected with CD147-specific shRNA and effect of U0126 on SGC7901 cells growth. (a) Gastric cancer cells (SGC7901, SGC7901/snc-RNA and SGC7901/siRNA2) seeded in 96-well microplates were cultured for 24, 48 and 72 h and their numbers were determined by absorbance. *P < 0.05 compared with SGC7901. (b) SGC7901 cells were treated with indicated concentrations of U0126 for 24, 48 and 72 h, respectively. U0126 inhibited SGC7901 cells growth dose and time dependently
CD147 silencing reduces MMP-2 and MMP-9 activities in SGC7901 cells
Since MMP-2 and MMP-9 play a critical role in tumour cell invasion, we examined the effects of CD147 silencing on the enzyme activities of MMP-2 and MMP-9 by using gelatin zymography. The gelatinolytic activities of both MMP-2 and MMP-9 were found to be reduced markedly in SGC7901/shRNA2 compared with SGC7901 and SGC7901/snc-RNA (P < 0.05; Figure 3). There was no significant difference between SGC7901/snc-RNA and SGC7901 (P > 0.05).
Analysis of MMP-2 and MMP-9 activities in SGC7901 cells gelatin zymography. Cells were incubated for 24 h and conditioned media were used for the measurement of MMP-2 and MMP-9 protein levels by gelatin zymography. (a) Photographs of the MMP-2 and MMP-9 bands, which are representative of three independent experiments, are shown. (b) Quantitative analysis of the bands. *P < 0.05 compared with SGC7901 and SGC7901/snc-RNA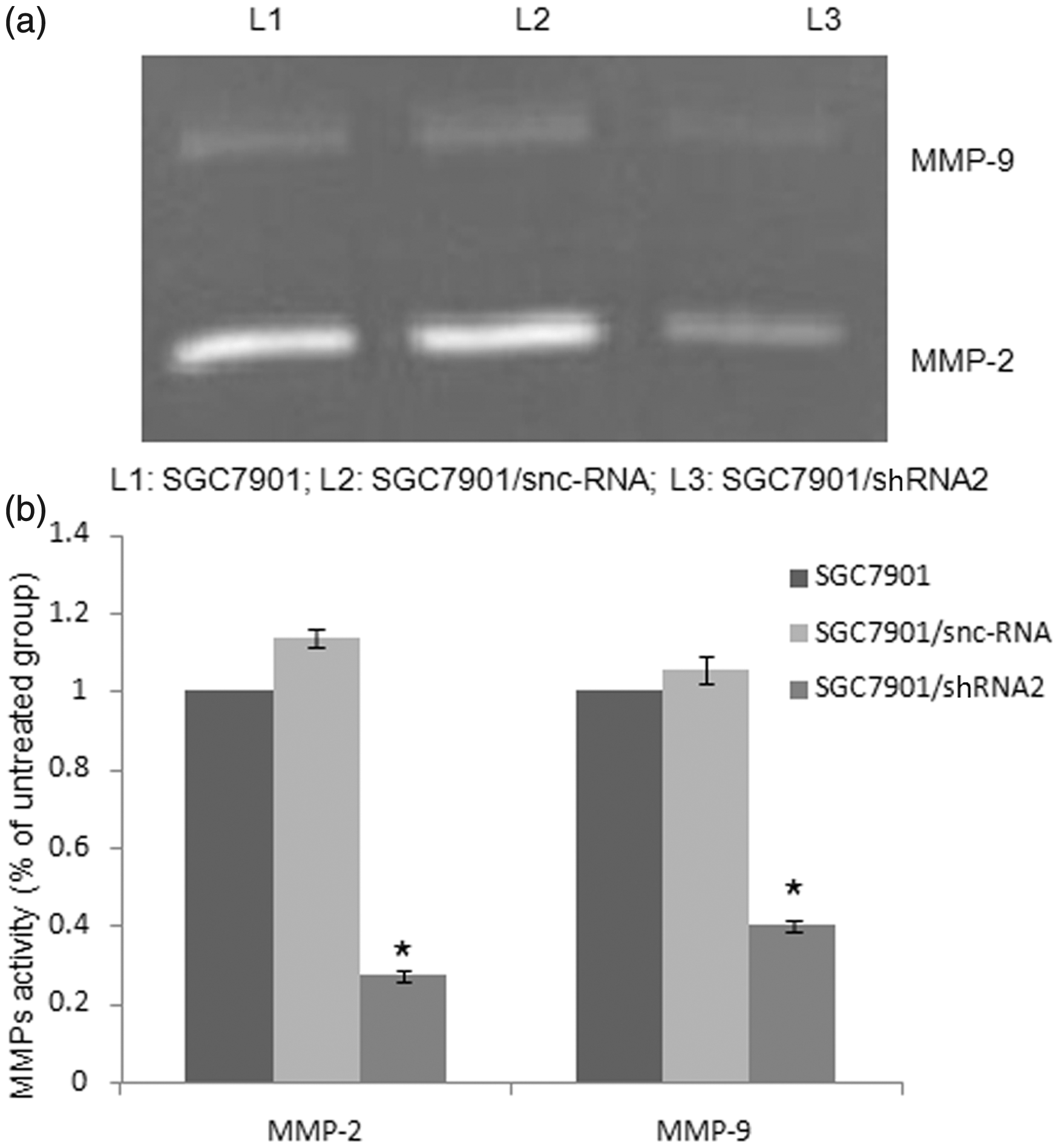
CD147 silencing reduces the invasive ability of SGC7901 cells in vitro
To examine whether the down-regulation of CD147 in SGC7901 affected its invasive ability, we performed an in vitro Matrigel Transwell analysis. The results showed that SGC7901 and SGC7901/snc-RNA cells had a similar ability to pass through the Matrigel coated filter, because the numbers of invading cells were roughly equal (Figure 4). The number of SGC7901/shRNA2 cells passing through the Matrigel was markedly lower than the numbers of SGC7901 and SGC7901/snc-RNA cells (P < 0.05). There was no significant difference between SGC7901/snc-RNA and SGC7901 (P > 0.05).
Effects of CD147-specific shRNA on invasion of SGC7901 cells. (a) Crystal violet staining results of the lower surface filters showed that the cells passed through the filter and attached to the lower side of the filter (×200). (b) The average number of cells that invaded through the filter was counted. The data were obtained from three independent experiments. *P < 0.05 compared with SGC7901 and SGC7901/snc-RNA. (A color version of this figure is available in the online journal)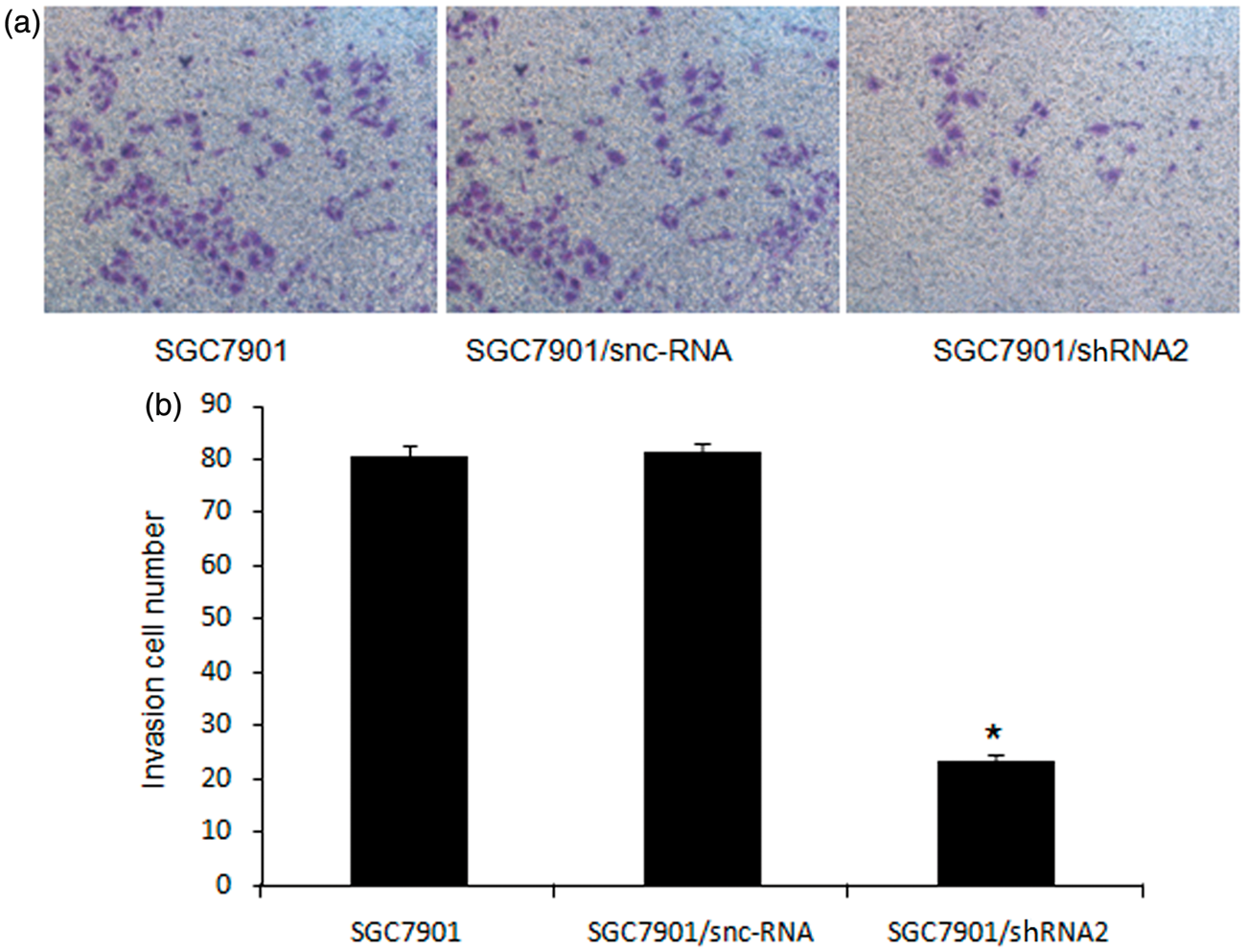
CD147 silencing inhibits phosphorylation of ERK1/2 but not other MAP kinases
MAPK pathway plays an important role in tumourigenesis and metastatic potential of gastric cancer, so we detect the effects of CD147 silencing on MAPKs expression. The results showed that CD147 silencing inhibited phosphorylation of ERK1/2 (P < 0.05) without inhibiting phosphorylation of P38 and JNK (P > 0.05; Figure 5). In addition, CD147 silencing had no effects on non-phosphorylation of MAPKs (P > 0.05).
Effects of CD147-specific shRNA on MAPK expression in SGC7901 cells. (a) The phosphorylation and non-phosphorylation of ERK1/2, P38 and JNK were examined by Western blot. Western blot analysis showed that si-CD147 inhibited the phosphorylation level of ERK1/2 but not other MAP kinases. (b) Quantitative analysis of the bands *P < 0.05 compared with SGC7901 and SGC7901/snc-RNA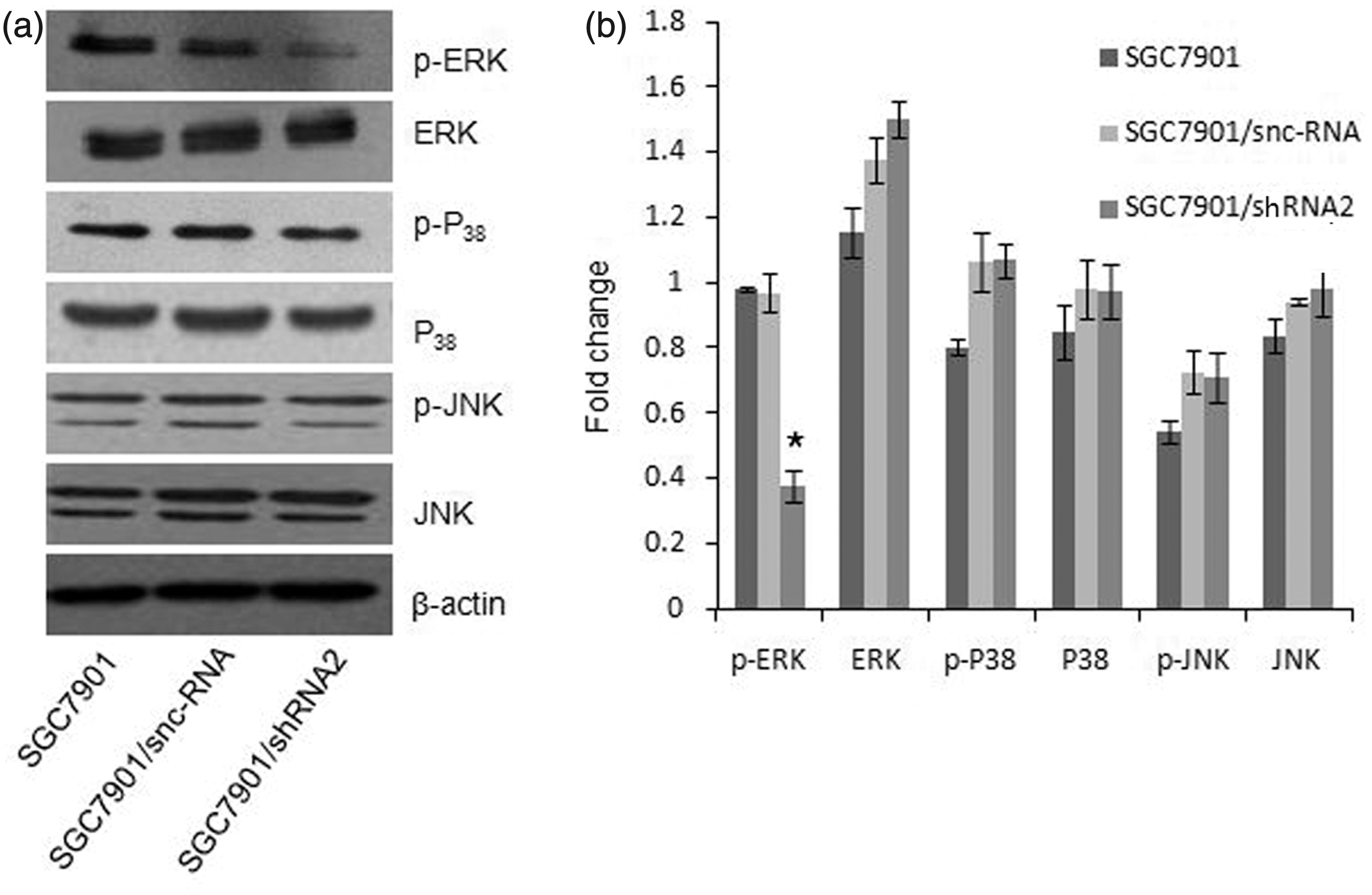
ERK1/2 inhibitor U0126 reduces MMP-2 and MMP-9 activities in SGC7901 cells
We first tested the inhibitory effects of the ERK1/2 inhibitor U0126 on the phosphorylation of ERK1/2. When SGC7901 cells were treated with U0126 (1–15 µmol/L) for 24 h, ERK1/2 phosphorylation was dose-dependently inhibited. In addition, when these cells were exposed to the inhibitor at 5 µmol/L for varying times, time-dependent suppression of ERK1/2 phosphorylation was observed (data not shown). In order to identify whether the activities of MMP-2, MMP-9 are related to ERK1/2, we examined the effects of ERK1/2 inhibitor U0126 on the activities of MMP-2, MMP-9 by gelatin zymography. The results showed that MMP-2, MMP-9 activities were significantly reduced in SGC7901 cells treated by U0126 in a concentration of 5 µmol/L (P < 0.05; Figure 6a, c).
Effects of ERK1/2 inhibitor U0126 on MMP-2, MMP-9 activities and invasion potential of SGC7901 cells. (a) Analyses of MMP-2 and MMP-9 activities. Before gelatin zymograph, serum-starved SGC7901 cells were pretreated with U0126 (5 µm/L) for 2 h. Photographs of the MMP-2 and MMP-9 bands, which are representative of three independent experiments, are shown. (b) Before transwell assay, we blocked the signalling pathway using ERK1/2 inhibitor U0126 in a concentration of 4 µmol/L which had no significant inhibition of cell proliferation (×200). (c) Quantitative analysis of the bands. *P < 0.05 compared with SGC7901. (d) The average number of cells that invaded through the filter was counted. The data were obtained from three independent experiments. *P < 0.05 compared with SGC7901 and SGC7901/snc-RNA. (A color version of this figure is available in the online journal)
Inhibitory effects of U0126 on cell proliferation and invasion
To determine the effect of U0126 on cell proliferation, SGC7901 cells were treated with indicated concentrations of U0126 for 24, 48 and 72 h, respectively. U0126 inhibited SGC7901 cells growth dose and time dependently (72 h U0126 IC50 = 4 µmol/L), as show in Figure 2(b). To examine whether ERK1/2 signalling pathway involved in SGC7901 invasion ability, we blocked the signalling pathway using ERK1/2 inhibitor U0126 in a concentration of 4 µmol/L which had no significant inhibition of cell proliferation. The results showed that the invasion ability was inhibited by U0126 relative to SGC7901 group (P < 0.05; Figure 6b, d).
CD147 silencing inhibited tumourigenesis of gastric cancer cells in vivo
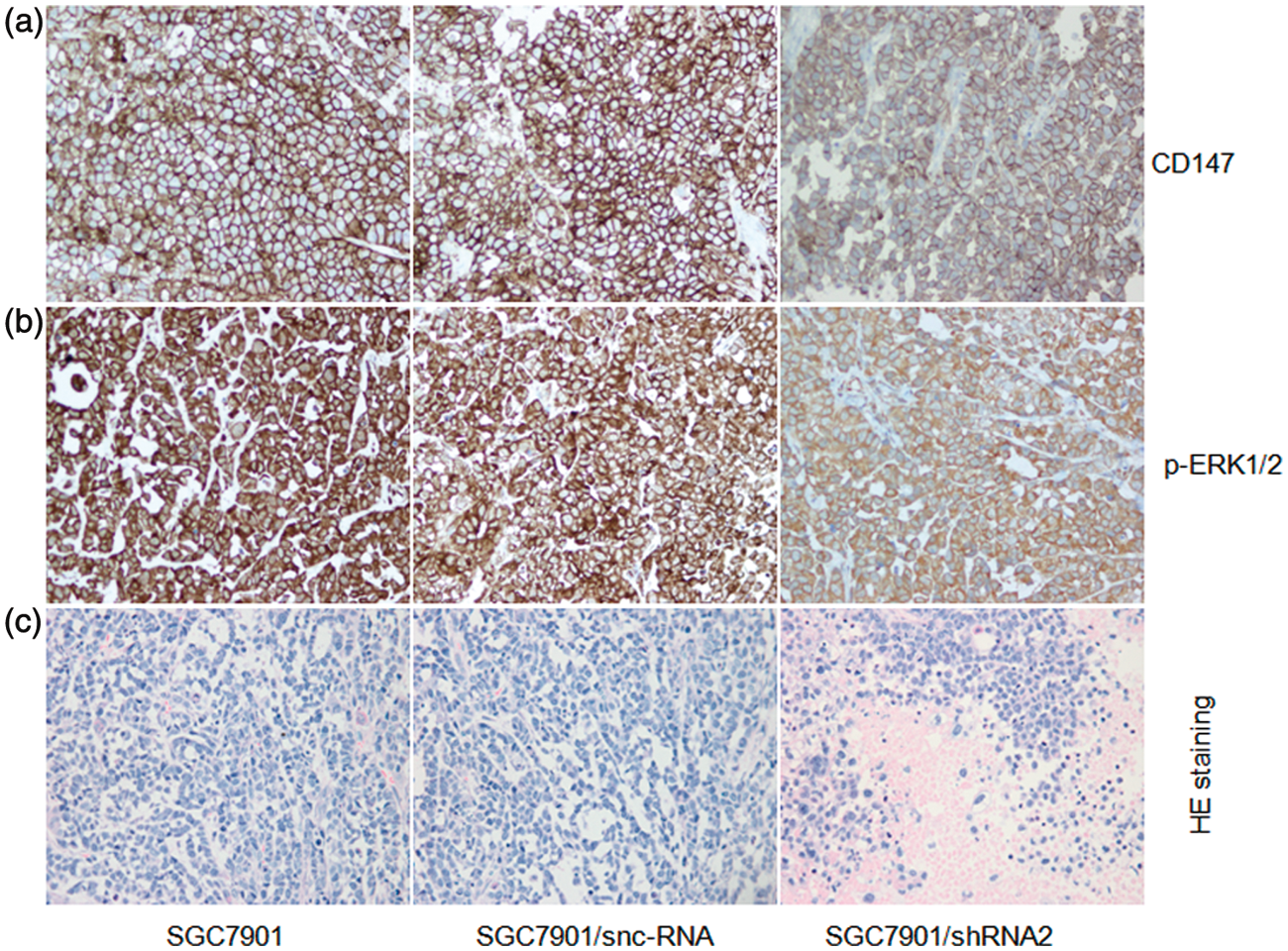
To further confirm the effects of CD147 on tumourigenesis of gastric cancer, tumour formation assay was performed in nude mice. After cell inoculation into nude mice for 40 days, the transplanted tumours were excised and their volume was calculated. In mice inoculated with SGC7901/shRNA2, the tumour volume was markedly smaller than the volume of SGC7901 and SGC7901/snc-RNA cells (P < 0.05). There was no significant difference between SGC7901/snc-RNA and SGC7901 (P > 0.05; Figure 7). Immunohistochemistry analysis showed that CD147 and p-ERK1/2 protein expressions were significantly decreased in tumour derived from SGC7901/shRNA2 compared to tumours derived from SGC7901 and SGC7901/snc-RNA cells (P < 0.05; Figure 8a, b). Histological examination did not reveal obvious morphological changes between the tumours generated from the three groups, but there were larger areas of necrosis in tumours formed by injecting with SGC7901/shRNA2 cells (Figure 8c).
Effects of CD147-specific shRNAs on tumour formation in vivo. (a) SGC7901, SGC7901/snc-RNA, SGC7901/siRNA2 were inoculated subcutaneously into the nude mice. (b) Tumour volumes were calculated. *P <0.05, compared with SGC7901 or SGC7901/snc-7901. (A color version of this figure is available in the online journal) Immunohistochemistry staining of CD147 and p-ERK1/2 protein and H&E staining. (a) CD147 expression in subcutaneous tumours was detected by immunohistochemical analyses. *P < 0.05 compared with SGC7901 and SGC7901/snc-RNA (×400). (b) p-ERK1/2 expression in subcutaneous tumours was detected by immunohistochemical analyses. *P < 0.05 compared with SGC7901 and SGC7901/snc-RNA (400×). (c) Histological analysis was performed in implanted tumours with H&E staining (400×).
Discussion
The expression of CD147 and its role in regulating MMPs expression, tumour progression and early metastasis have been extensively studied in various cancer types.4,7–11,30 Our previous studies have shown that CD147 plays an important role in the proliferation, invasion, metastasis and chemosensitivity of the human gastric cancer cell line SGC7901. 31 To better understand the function of CD147 in tumour biology, we thoroughly characterized the effects of CD147 silencing in gastric cancer cell line SGC7901. In this study, CD147 silencing was found to significantly reduce cell proliferation, invasion and MMPs activities, thus confirming an essential role for CD147 in tumour biology. We then set out to identify the mechanism by which CD147 regulates such diverse functions.
In this study, we discovered that CD147 has a more fundamental role in tumour invasion by its ability to regulate ERK1/2 signalling in gastric cancer. CD147 silencing cells down-regulated ERK1/2 signalling in SGC7901/shRNA2, as shown in Figure 5. We examined the phosphorylation and non-phosphorylation ERK1/2, P38 and JNK by Western blot and we found that CD147 silencing inhibited the phosphorylation of ERK1/2, but not the non-phosphorylation ERK1/2 and other MAPKs in SGC7901/shRNA2. Previously, ERK signalling pathway involved in CD147-mediated cancer progression have been researched in several studies and several lines of evidence have shown that CD147 stimulate cell survival pathway signalling, including phosphorylation of ERK, SPAK in hepatocellular carcinoma cells.26,27 Previous studies have reported that purified recombinant CD147ECD has been shown to activate the MAPK pathway and stimulate MMPs expression in cultured fibroblasts. 32 In addition, study showed that dimerization is essential for HAb18G/CD147 promoting tumour invasion via MAPK pathway. 33 To examine whether ERK1/2 signalling pathway was involved in CD147-mediated gastric cancer invasion, we blocked the ERK1/2 signalling by ERK1/2 inhibitor U0126 in SGC7901 cells, and then tested the MMP-2 and MMP-9 activities by gelatin zymography and cell invasion potential by transwell assay. The results showed that MMP-2 and MMP-9 activities were significantly reduced, and the invasion ability was highly inhibited (Figure 6). The significant inhibition of MMPs activities and invasion by ERK inhibitor confirmed that ERK1/2 signalling pathway is necessary for gastric cancer invasion. In vivo nude mouse experiments suggested that inhibition of CD147 expression reduces tumour cell tumourigenicity. Immunohistochemical staining showed that CD147 and p-ERK1/2 proteins were significantly decreased in tumour derived from SGC7901/shRNA2 cells compared to tumours derived from SGC7901 and SGC7901/snc-RNA. Our results illustrate that CD147 has a more fundamental role in tumour invasion than earlier realized by its ability to regulate ERK1/2 signalling in gastric cancer. Nevertheless, other signalling pathways have also been studied in CD147-mediated tumourigenesis, angiogenesis, drug resistance and metastasis. CD147 was found to stimulate tumour angiogenesis by elevating vascular endothelial growth factor and MMP expression in neighbouring fibroblasts via the PI3K-AKT signalling pathway.34,35 CD147 is also involved in multidrug resistance of cancer cells via hyaluronan-mediated activating of ErbB2 signalling and cell survival pathway activities.36–38 CD147 regulates the canonical Wnt/b-catenin signalling pathway in accelerating lung tumourigenesis. 39 HAb18G/CD147 promotes epithelial–mesenchymal transition through TGF-b signalling and is transcriptionally regulated by Slug. 40 The disparity may be related to different cancer cell lines and different focus of study. Previously, CD147 have been studied in tumour progression through angiogenesis, drug resistance, metastasis and invasion. In this study, we focused on the signalling pathway involved in CD147-mediated tumour invasion in gastric cancer. Mechanism of CD147 in tumour development is complicated. Recently, CD147 has been suggested to interact with a wide variety of molecules at the cell surface, such as cyclophilins, integrins, caveolin-1 and monocarboxylate transporters.14,41–43 Determining which, if any, of these interactions are important for CD147’s role in gastric tumourigenesis and warrants further investigation.
In summary, we identify CD147 as a novel regulator of the ERK1/2 signalling pathway in gastric cancer growth and invasion. Our results strengthen the proposal that CD147 would make an attractive target for anticancer therapies of gastric cancer.
Footnotes
Author contributions
All authors participated in the interpretation and review of this manuscript. LC and YP designed and performed the research, they contributed equally to this work.
Acknowledgements
This work was supported by National Natural Science Foundation of China (No. 30873022) and Science and Nanjing Science and Technology Committee project (No. 201108025) and Nanjing Health Young Talent Project.