
Abstract
Multidrug resistance (MDR) has become a major impediment to a successful treatment for liver cancer patients, and one of the common reasons for MDR is the activation of ABCB1 gene, leading to the over-expression of P-glycoprotein (P-gp), which conferred cancer cells be resistant to a broad range of anticancer drugs. MicroRNAs (miRNAs) are a class of short, non-coding RNA moleculars that can regulate gene expression at the post-transcriptional level. In the current study, the aim is to explore whether miRNA participates in the regulation of MDR mediated by ABCB1. We found that the expression of ABCB1 was correlated with the doxorubicin IC50 dose in eight hepatocellular carcinoma (HCC) cell lines: Hep3B, HCC3, LM-6, SMMC7721, Huh-7, SK-Hep-1, HepG2 and BEL-7402. Using the bioinformatics, we discovered that there were several miRNAs that can bind to the 3’UTR of ABCB1 gene. Among these candidate miRNAs, miR-223 was chosen for further study. Then, EGFP reporter assay, real-time PCR and Western blot were performed to verify that miR-223 targeted ABCB1 3’UTR directly, and miR-223 downregulated ABCB1 at both mRNA and protein levels. Finally, we found that the over-expression of miR-223 increased the HCC cell sensitivity to anticancer drugs, and the inhibition of miR-223 had the opposite effect. Importantly, the over-expression or silencing of ABCB1 can rescue the cell response to the anticancer drugs mediated by miR-223 over-expression or inhibition, respectively. In conclusion, our findings indicated that miR-223 played an important role in the regulation of MDR mediated by ABCB1, and it suggests that miR-223 may be considered as a therapeutic biomarker for HCC patients who had MDR problems induced by high expression of ABCB1.
Introduction
Hepatocellular carcinoma (HCC) is the sixth most common cancer and the third cause of cancer-related death in the world. 1 The high mortality and the low-five-year survival rate, which is less than 7% in the United States, 2 seriously threaten the human health. Combination of chemotherapy is an important treatment method for HCC patients, besides surgery and other limited treatment. Recently, the HCC patients present high resistance to various chemotherapeutic reagents, such as doxorubicin, paclitaxel, 5-Fu and cisplatin,2–4 which are the first-line medicine for HCC treatment. Therefore, multidrug resistance (MDR) becomes a major obstacle to a successful therapy for HCC patients. One of the common forms for MDR is induced by the activation of ABCB1 gene, leading to the over-expression of its encoding protein, P-glycoprotein (P-gp), a 170–190 kD protein. It belongs to the ATP-binding cassette (ABC) transporter family and is an energy-dependent multidrug efflux pump, conferring to cancer cells resistant to a broad range of therapeutic reagents.5–7 Thus, it is crucial to discover a molecular biomarker for patients with MDR problems. It has been reported that miRNAs play an important role in the regulation of MDR.8,9
MicroRNAs (miRNAs) are a class of short, non-coding RNA molecules and regulate many genes through binding to the 3’UTR of target genes by complementary base pair, leading to the mRNA degradation or translation repression. 10 miRNAs are involved in various biological processes, such as cell death, cell proliferation, apoptosis and other processes. 11 It has been reported that miRNAs are frequently located in cancer-related genomic regions or in fragile sites, or some miRNAs are located in deletion sites leading to the miRNA downregulated in cancer cell samples, 12 all of which make miRNAs function like an oncogene or tumour suppressor in cancers. 13 miRNAs regulate about 60% of the protein-coding genes, 14 not only in cancer cell proliferation and metastasis,15–17 but also in the regulation of MDR. For example, miR-27 a and miR-451 regulate drug resistance mediated by P-gp in human cancer cells. 8 miR-222 plays an important role in the regulation of drug resistance in colorectal cancer by targeting ADAM-17. 18 miR-130 a is associated with P-gp-mediated drug resistance in ovarian cancer cells, 19 and downregulation of miR-200 c is related to poor drug response in human breast cancer cells. 20 Although these reports suggest that miRNAs exert an important role in drug resistance in some cancers, but there is little report about miRNAs involved in the regulation of drug resistance induced by ABCB1 gene (or P-gp) in HCC cells.
In the present study, we found that ABCB1 was correlated with doxorubicin IC50 in different HCC cells, and several miRNAs, including miR-223, targeted ABCB1 3’UTR directly. Further study showed that the over-expression of miR-223 can improve the cells chemotherapy response to doxorubicin and paclitaxel in cells with high-ABCB1 levels, and the inhibition of miR-223 can enhance the cells drug resistance in HCC cells with relatively low-ABCB1 expression. Thus, our data, at least partially, demonstrated the mechanism of miR-223 in the regulation of MDR mediated by ABCB1 in HCC cells.
Materials and methods
Cell culture and transfection
Hep3B cells were cultured in Eagle’s minimum essential medium. HCC3, LM-6 and SMMC7721 cells were cultured in Dulbecco’s modified Eagle’s medium. Huh-7, SK-Hep-1, HepG2 and BEL-7402 cells were cultured in modified Eagle’s medium. Cell-culture medium was supplemented with 10% fetal bovine serum and 1% PS (100 U/mL penicillin, 100 µg/mL streptomycin). All the cells were maintained in a humidified incubator with 5% CO2 at 37℃. All the transfection was performed by Lipofectamine™ 2000 (Invitrogen, USA) according to the manufacturer’s protocol.
Plasmid construct
All the miRNA mimics, the antisense oligonucleotides (ASO) and the controls were purchased from the Shanghai GenePharma Company in China. The 3’UTR of ABCB1 (containing the miRNA-binding sites) was synthesized and cloned into the downstream of pcDNA3-EGFP vector. The mutant 3’UTR of ABCB1 (four nucleotides within the binding sites were mutated) was also synthesized and cloned into the pcDNA3-EGFP vector. The gene-coding ABCB1 was amplified by PCR and cloned into the pcDNA3 vector.
RNA isolation and real-time PCR
Total RNA was isolated using the TRIZOL reagent (Invitrogen, USA) according to the manufacturer’s instruction. The concentration and quality of RNA were measured using Nanodrop 1000 Spectrophotometer (Thermo Scientific, USA). For the miRNA reverse transcription (RT) reaction, special primers were used, and RNU6B (U6 small nuclear B non-coding RNA) was used as an internal control. For RT of mRNA, Oligo (dT) was used as the RT primer. The real-time PCR was performed by the SYBR Green PCR master Mix (Applied Biosystems, USA) according to the following conditions: 95℃ for 5 min followed by 40 cycles of amplification at 95℃ for 30 s, 56℃ for 30 s and 72℃ for 30 s. GAPDH was used as the internal control to normalize the expression of ABCB1.
Western blot analysis
HCC cells were plated into six-well plates at a density of 30 × 104 cells/well and transfected with miRNA mimic or ASO control when the cells reached 80% confluence in the second day. The cells were harvested at 48 h after transfection and lysed by RIPA buffer (50 mM Tris–Cl, PH 8.8, 150 mM Nacl, 1% NP−40, 1% sodium deoxycholate, 0.1% SDS) for 30 min at 4℃. The protein concentration was measured using BCA method, and 30 µg protein was loaded into 8% SDS–PAGE gels for analysis. The first antibody was mouse monoclonal anti-ABCB1 antibody (Calbiochem, San Diego, CA, 1:500 dilution) or anti-GAPDH antibody (Abcam, USA, 1:1000 dilution). The secondary antibody was the goat anti-rabbit or anti-mouse IgG conjugated with HRP (horseradish peroxidase) at a dilution of 1:1000. The bound antibodies were detected with the use of ECL-Plus Western Blotting Detection system (GE Healthcare), and the chemiluminescent signals were detected with the use of high-performance chemiluminescence film (GE Healthcare). GAPDH was used as an internal control.
EGFP reporter assay
The cells were planted into 48-well plates at a density of 3 × 104 cells/well before transfection, and when cells reached 80% confluence, the cells were co-transfected with miR-223 mimics or miR-223 ASO and pcDNA3/EGFP-ABCB1 3’UTR or mutant 3’UTR or control construct. The pDsRed2-N1 (Clontech) construct was transfected with the above plasmid as the spiked-in control. The cells were harvested at 48 h after transfection and lysed by RIPA buffer, and then the fluorescence intensity was measured by the Fluorescence Spectrophotometer F-4500 (HITACHI).
Cell viability assay (WST-1 assay)
The cell viability was measured by WST-1 assay according to the manufacturer’s protocol. The transfected cells were plated into 96-well plates at a density of 5000 cells/well. Then the cells were exposed to different chemotherapy reagents at a series of concentrations for 24 h and were incubated with WST-1 reagent for 1 h at 37℃ to measure the absorbance on A450nm with a spectrophotometer. On the basis of cell viability, the survival rate was calculated as follows: (cell viability without chemotherapy reagent − cell viability under different concentration)/(cell viability without chemotherapy reagent) × 100%. IC50 was calculated with software using the cell viability under different concentrations.
Silencing ABCB1 by siRNA transfection
The cells were plated onto six-well plates at a density of 30 × 104 cells/well and transfected with ABCB1 siRNA or control. At 48 h after transfection, the cells were harvested for protein analysis using ABCB1 antibody (Calbiochem, San Diego, CA, 1:500 dilution), and GAPDH was used as an internal control.
Statistical analysis
Differences between groups were determined by two-tailed Student’s t-test, and P < 0.05 was considered statistically significant. The relationship between ABCB1 and Doxorubicin IC50 was measured by Spearman correlation analysis. Each experiment was repeated at least twice, and each group was in triplicate.
Results
ABCB1 expression is associated with the response of HCC cells to doxorubicin reagent
It is well known that ABCB1-encoded protein, P-gp, plays an important role in the process of MDR. To explore the potential mechanism of ABCB1 in the regulation of cell response to doxorubicin in HCC cells, the expression of ABCB1 was detected by Western blot in eight different HCC cell lines: Hep3B, HCC3, LM-6, SMMC7721, Huh-7, SK-Hep-1, HepG2 and BEL-7402 cells (Figure 1a). Given that ABCB1 was differentially expressed in these HCC cell lines, which differed in their doxorubicin IC50 (Figure 1b), we concluded that ABCB1 was upregulated in doxorubicin-resistant HCC cell lines, except in SK-Hep-1 cells. As shown in Figure 1c, the expression of ABCB1 had a positive relationship with doxorubicin IC50 in eight HCC cell lines (P = 0.0155, R2 = 0.6508). Thus, these results suggest that ABCB1 may exert an important role in the process of HCC cell resistance to doxorubicin.
The expression of ABCB1 is related with the IC50 of doxorubicin in HCC cells. (a) The expression of ABCB1 was analysed by Western blot analysis in eight HCC cell lines, and GAPDH was considered as an internal control. (b) The IC50 of doxorubicin in HCC cell lines. (c) The relationship between doxorubicin IC50 and the relative ABCB1 expression level in eight HCC cell lines (R2 = 0.6508, P = 0.0155)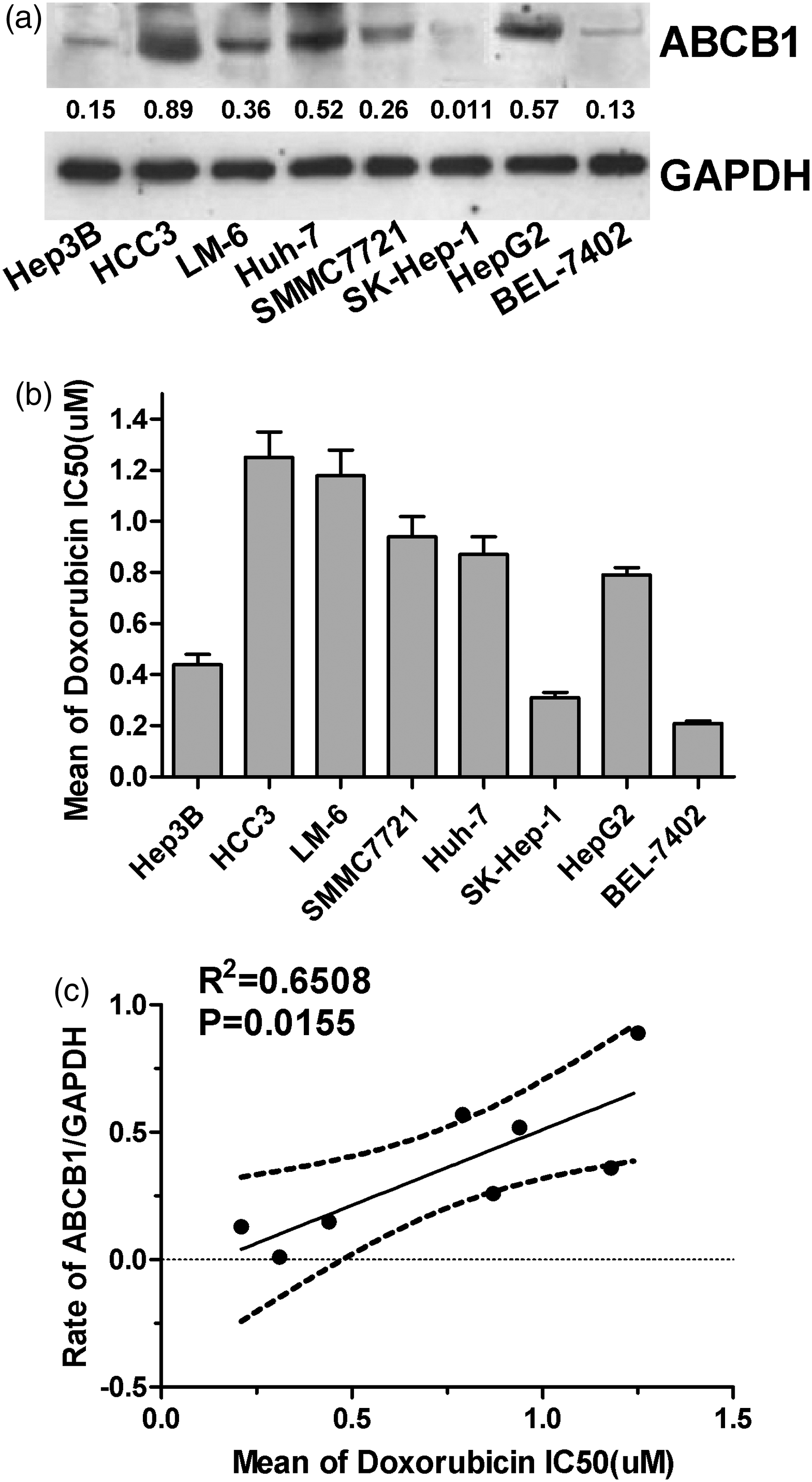
Systematic validation of miRNAs targeting ABCB1 in HCC cells
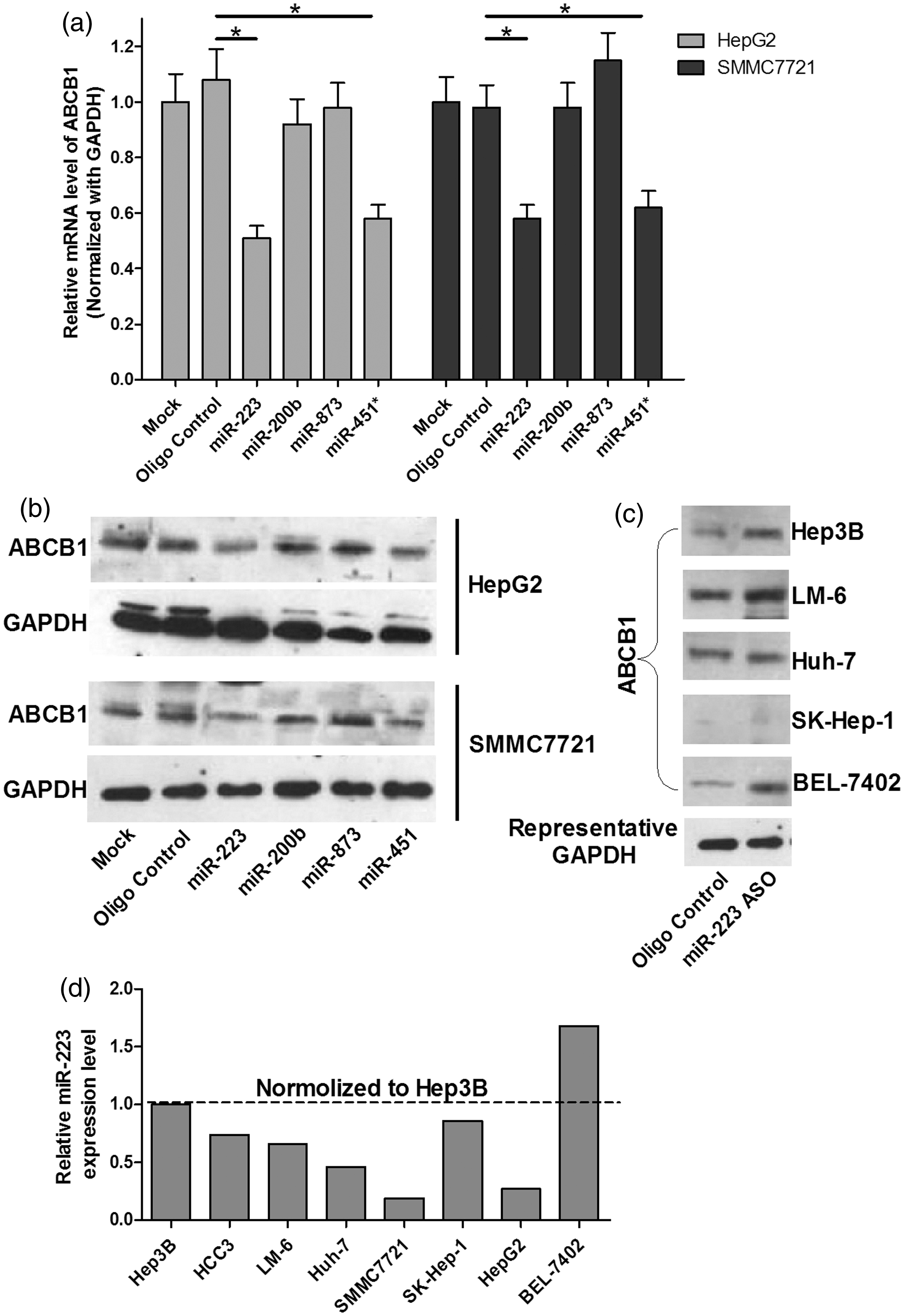
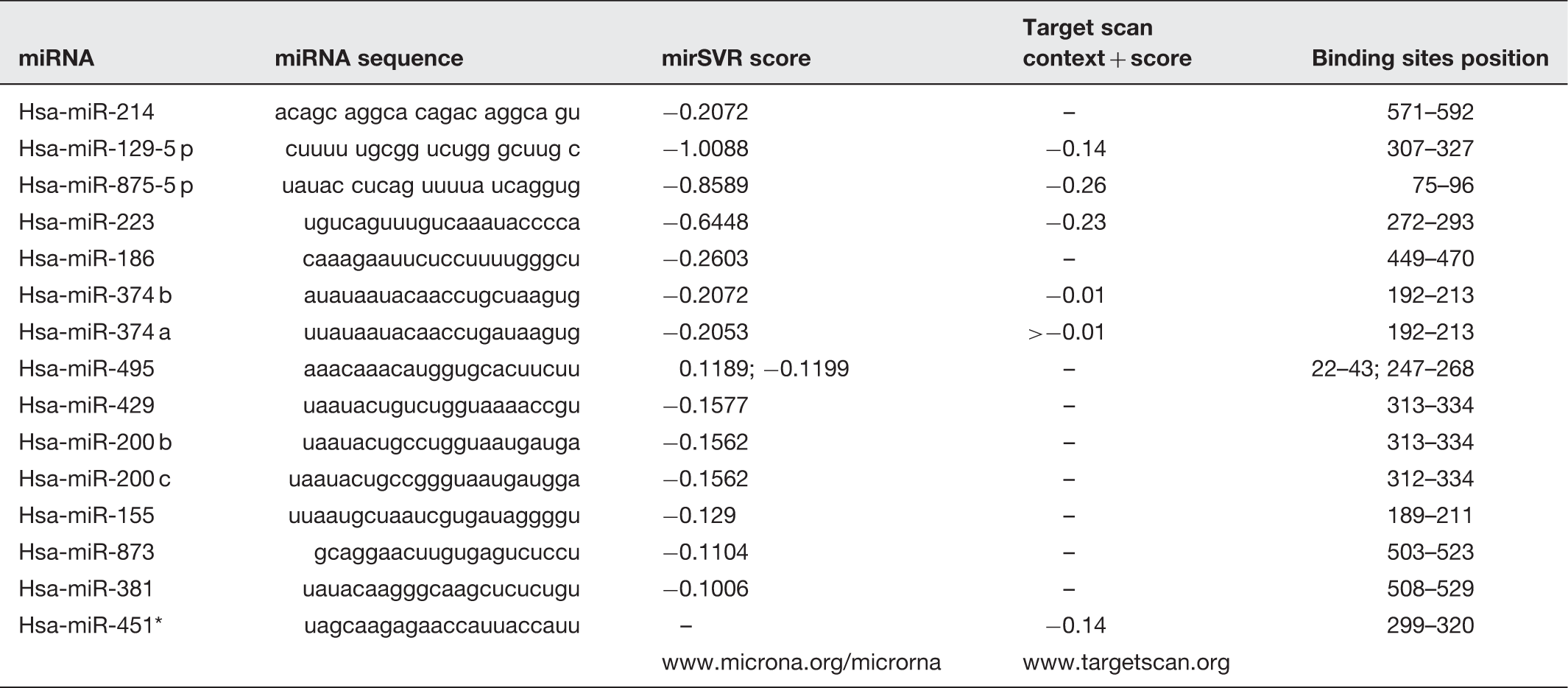
To explore the role of miRNA in ABCB1-mediated MDR in HCC cells, TargetScan and microCosm were used to predict the miRNAs that had putative binding sites on ABCB1 3’UTR. Table 1 displayed the candidate miRNAs for ABCB1. Then, we used the EGFP reporter assay to test the effect of these miRNAs on the EGFP intensity controlled by ABCB1 3’UTR. First, we transfected the cells with all miRNA mimics or the mimic control in both Hep3B and SMMC7721 cell lines. As shown in Figures 2a and b, real-time PCR analysis indicated that the expression of most of the miRNAs increased by more than 10-fold in miRNA mimics-treated cells. Next, as shown in Figure 2c, only the over-expression of miR-223, miR-200 b, miR-873 and miR-451* significantly reduced the EGFP intensity controlled by ABCB1 3’UTR. Furthermore, to validate the miR-223, miR-200 b, miR-873 and miR-451*, truly suppressed endogenous ABCB1, we performed real-time PCR and Western blot to detect ABCB1 mRNA and protein levels. As shown in Figures 3a and b, the over-expression of miR-223 or miR-451* repressed ABCB1 on both mRNA and protein levels, but miR-873 and miR-200 b had no significant effect on ABCB1 expression. We also discovered that inhibition of miR-223 increased the ABCB1 expression in most of the HCC cell lines except in SK-Hep-1 cells by Western blot analysis (Figure 3c). Finally, we detected the endogenous miR-223 expression in HCC cells and found that miR-223 is differently expressed in eight HCC cell lines, which was inversely correlated with ABCB1 levels in these cells, indicating a concordant inverse correlation between miR-223 and its target gene (Figure 3d). Taken together, these data indicate that miR-223 may target ABCB1 and downregulate ABCB1 expression.
The effect of the predicted miRNAs on the EGFP intensity of ABCB1 3’UTR. (a,b) The expression of miRNAs was analysed by real-time PCR in the cells transfected with miRNA mimics or the mimic control. (c) EGFP reporter analysis was used to study the effect of the miRNAs on the ABCB1 3’UTR in the cells co-transfected with miRNA mimics and ABCB1 3’UTR, or the mimic control and ABCB1 3’UTR. *P < 0.05 The effect of the four candidate miRNAs on ABCB1 expression in HCC cells. (a) The effect of miRNAs on ABCB1 mRNA level was analysed by real-time PCR analysis. GAPDH was considered as an internal control. *P < 0.05. (b) The effect of the miRNAs on the ABCB1 protein level by Western blot. GAPDH was considered as an internal control. (c) The effect of inhibition of miR-223 on the ABCB1 protein level was analysed by Western blot in different HCC cell lines. GAPDH was considered as an internal control. (d) Quantification of miR-223 in eight HCC cells was carried out using real-time PCR. The expression level of endogenous miRNA-223 was indicated as relative CT value and normalized by its level in Hep3B cells Predicted miRNAs for ABCB1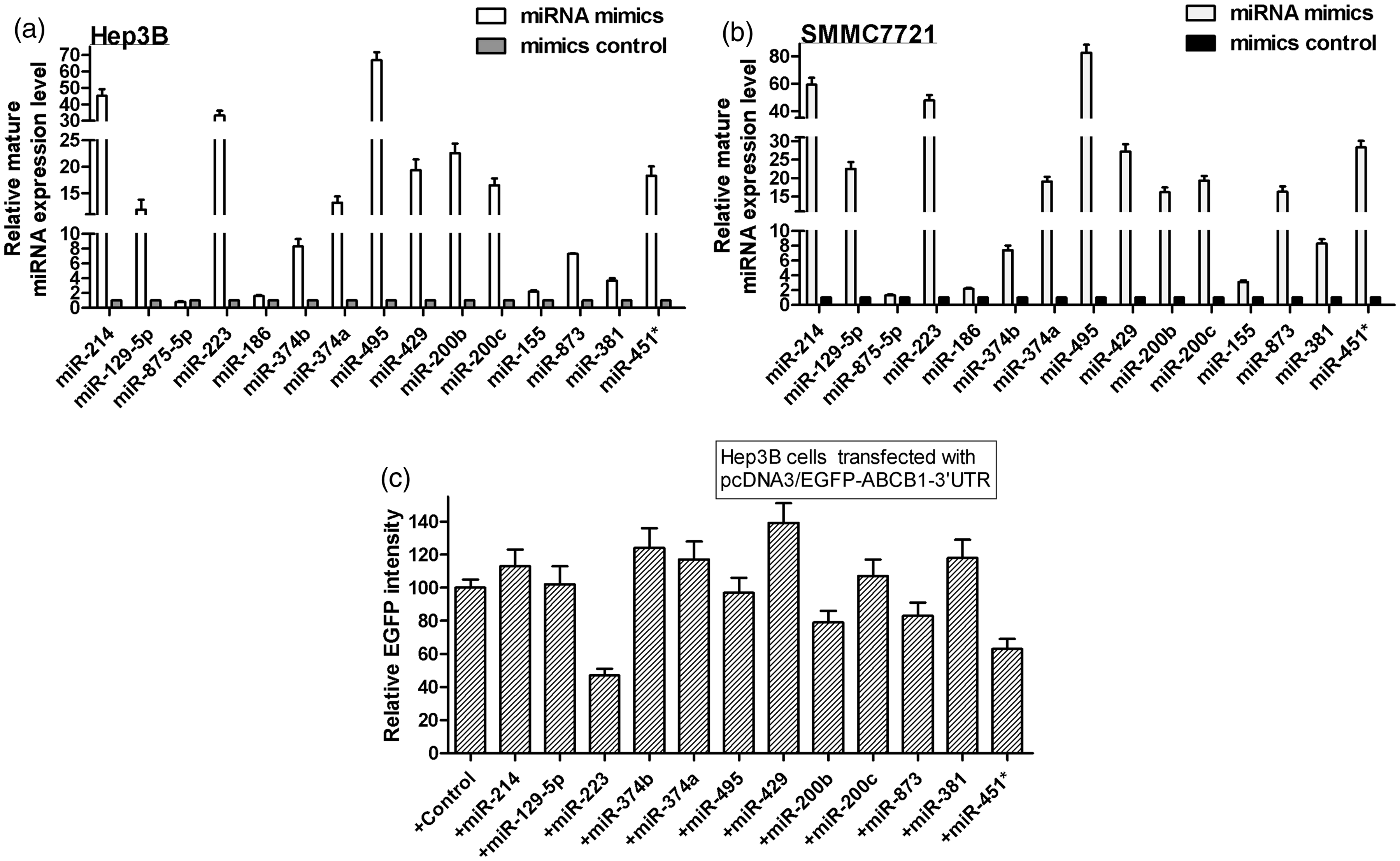
MiR-223 downregulates ABCB1 directly in HCC cells
To validate miR-223 that specifically suppresses ABCB1 expression through targeting its 3’UTR, we made a four-point mutation in the miR-223-binding site of ABCB1 3’UTR. Figure 4a displayed the algorithms of miR-223 with ABCB1 3’UTR and the mutant-binding sites. The over-expression or inhibition of miR-223 was again verified by real-time PCR analysis in both LM-6 and BEL-7402 cells, and we found that miR-223 expression was increased by about 150 and 100% in miR-223 mimics-transfected LM-6 and BEL-7402 cells, respectively, and reduced by about 70% in miR-223 ASO-transfected cells (Figure 4b). Then we performed the EGFP reporter assay by co-transfecting the cells with miR-223 mimics or the ASO of miR-223, together with the pcDNA3/EGFP-ABCB1 3’UTR or the mutant 3’UTR, or the control constructs in both LM-6 and BEL-7402 cell lines. Figure 4c indicated that over-expression of miR-223 reduced the EGFP intensity controlled by ABCB1 3’UTR by about 40% in both cell lines. In contrast, inhibition of miR-223 increased the EGFP intensity by about 100 and 70% in LM-6 and BEL-7402 cell lines, respectively. However, when using a mutant vector containing a four-point mutation in the miR-223-binding site, the EGFP intensity was not significantly changed by either over-expression or blocking of miR-223 (Figures 4c and d). In combination with the above results that miR-223 suppressed ABCB1 at mRNA and protein levels, our data suggested that miR-223 targeted the 3’UTR of ABCB1 directly in HCC cells.
ABCB1 is a direct target of miR-223. (a) Showed the algorithms of miR-223 with the potential-binding sites on the 3’UTR of ABCB1 and the four-mutated nucleotides within the binding sites. (b) The expression of miR-223 was analysed by real-time analysis in the cells transfected with miR-223 mimic or ASO of miR-223 in LM-6 and BEL-7402 cells. (c,d) The effect of miR-223 on the EGFP intensity with ABCB1 3’UTR by EGFP reporter assay in the cells co-transfected with miR-223 mimics or ASO and pcDNA3/EGFP-ABCB1 3’UTR or the mutant construct or with the controls. RFP intensity was considered as an internal control. *P < 0.05.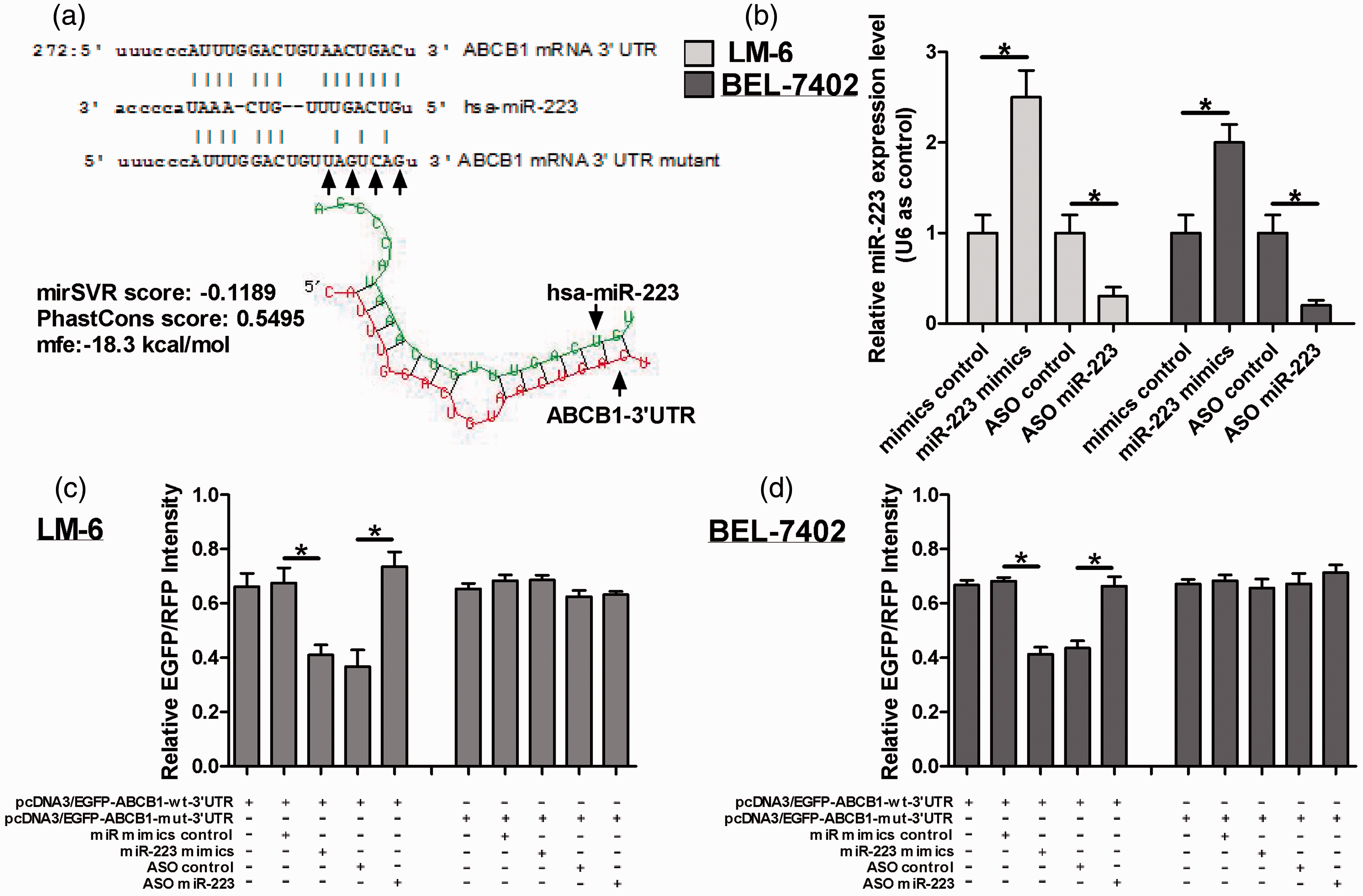
MiR-223 over-expression increases the HCC cell sensitivity to doxorubicin and paclitaxel
Given that ABCB1 is involved in MDR, we explored whether miR-223 can regulate the MDR mediated by ABCB1. The rescue experiment was preformed to validate that miR-223 regulates the MDR through ABCB1 rather than other genes. First of all, as shown in Figure 5a, we discovered that the over-expression of ABCB1 lacking the 3’UTR can reverse the endogenous ABCB1 protein level in HCC cell lines, SMMC7721 and HepG2, in which ABCB1 is highly expressed . Then, we detected the effect of miR-223 on the cell response to paclitaxel and doxorubicin in HCC cells by measuring the survival rate and IC50 dose of the chemotherapeutic reagents. As shown in Figures 5b and c, over-expression of miR-223 reduced the cell-survival rate under paclitaxel and doxorubicin treatment, compared to the mimic control. Importantly, over-expression of ABCB1 can restore the cell-survival rate reduced by miR-223. Accordingly, the over-expression of miR-223 had the same effect on IC50 of paclitaxel and doxorubicin in both cell lines. In addition, we found that miR-223 over-expression increased the cisplatin IC50, but over-expression of ABCB1 cannot rescue the cell response to cisplatin mediated by miR-223, indicating that other gene might be involved in this process. Moreover, miR-223 had no effect on the cell response to 5-Fu treatment (Figure 5c). Thus, these results suggest that miR-223 can regulate the cell response to doxorubicin and paclitaxel mediated by ABCB1, but not cisplatin and 5-Fu.
Over-expression of miR-223 contributes to cells sensitivity to doxorubicin and paclitaxel. The cells were transfected with miR-223 mimics, or co-transfected miR-223 mimic and pcDNA3/ABCB1, or with the control. Then, (a) Western blot showed that over-expression of ABCB1 can reverse the endogenous ABCB1 level reduced by miR-223 over-expression. GAPDH was considered as an internal control. (b) The effect of miR-223 over-expression on the cell-survival rate and the effect of ABCB1 over-expression on the survival rate mediated by miR-223 were analysed by WST-1 assay under paclitaxel and doxorubicin treatment. (c) WST-1 assay showing the effect of the miR-223 over-expression and ABCB1 on the IC50 of different chemotherapeutic reagents, 5-Fu, cisplatin, doxorubicin and paclitaxel
Inhibition of miR-223 resulted in the HCC cell resistance to doxorubicin and paclitaxel
To validate the effect of miR-223 on cell response to doxorubicin and paclitaxel, inhibition of miR-223 was performed in Hep3B and BEL-7402 cell lines, which had a relatively low-ABCB1 expression. In contrast, as shown in Figure 6a, we discovered that silencing ABCB1 can reduce the endogenous ABCB1 protein level that is upregulated by miR-223 blocking. Furthermore, inhibition of miR-223 increased the cell-survival rate and IC50 under paclitaxel and doxorubicin treatment, and silencing ABCB1 abolished the cell-survival rate and IC50 increased by miR-223 inhibition (Figures 6b–c). In addition, inhibition of miR-223 only reduced the IC50 of cisplatin and had no effect on the 5-Fu treatment (Figure 6c).
Inhibition of miR-223 increases the HCC cell resistance to doxorubicin and paclitaxel. The cells were transfected with ASO of miR-223, or co-transfected miR-223 ASO and ABCB1 siRNA, or with the control. Then, (a) Western blot shows that silencing ABCB1 can rescue the endogenous ABCB1 expression increased by the inhibition of miR-223. GAPDH was considered as an internal control. (b) The effect of inhibition of miR-223 on the cell-survival rate and the effect of silencing ABCB1 on the cell survival rate mediated by miR-223 inhibition were analysed by WST-1 assay under paclitaxel and doxorubicin treatment. (c) WST-1 assay showed the effect of miR-223 and ABCB1 on the IC50 of different reagents after the cells were transfected with ASO of miR-223 and ABCB1 siRNA, or with the control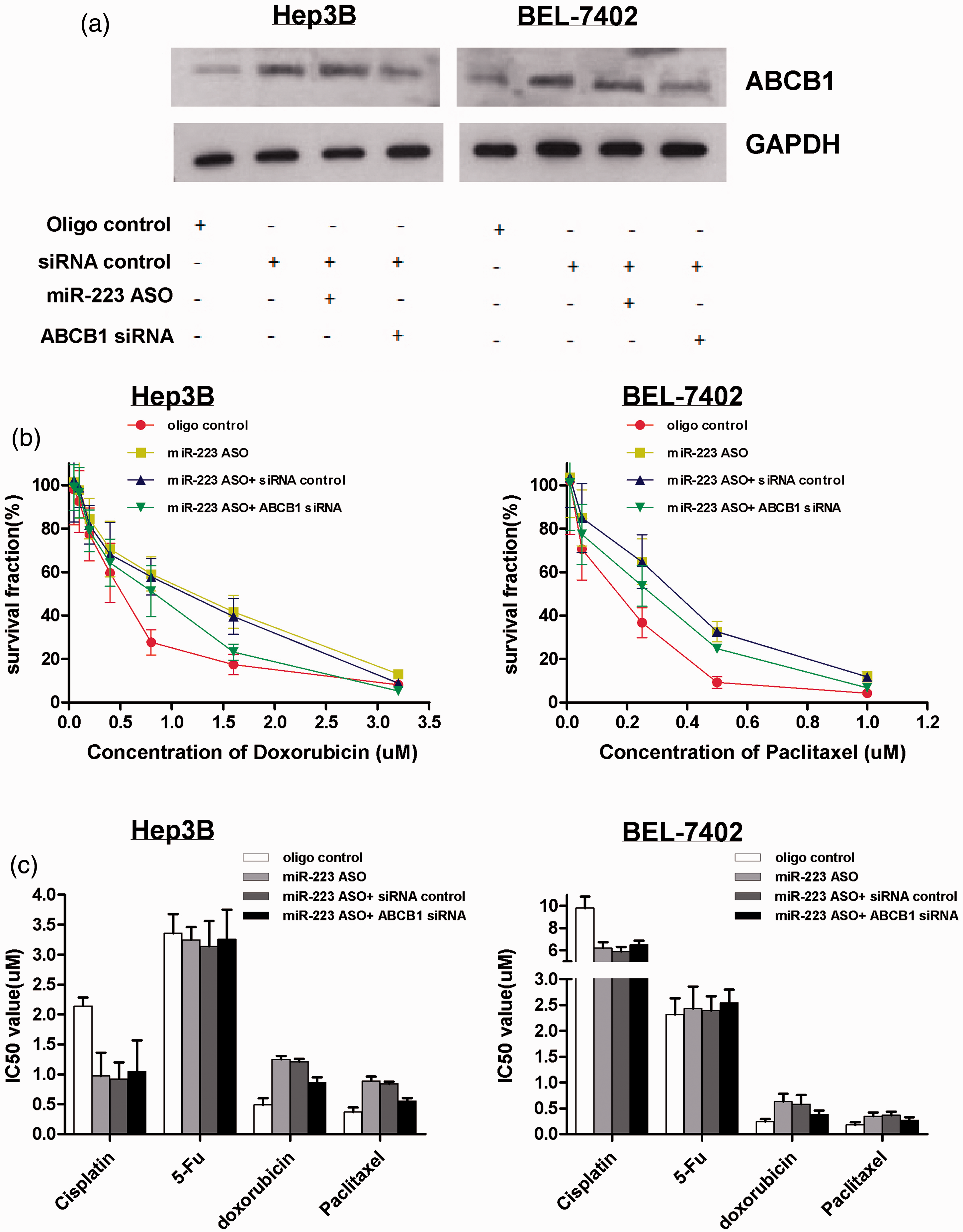
Therefore, these results indicate that miR-223 plays an important role in the regulation of ABCB1-MDR to doxorubicin and paclitaxel, rather than 5-Fu and cisplatin resistance.
Discussion
miRNAs have been known as regulators of various genes involved in cancer initiation, development and prognosis. Even more miRNAs target genes participating in the chemoresistance to different anticancer drugs. 21 Wong et al. 22 showed that miR-223 is highly downregulated in HCC cells and inhibits the cell viability. Previous reports suggested that miR-223 plays an important role in human granulopoiesis, 23 and miR-223 can serve as an important biomarker in liver injury, including HCC. 24 The role of miR-223 in chemoresistance of HCC cells is not yet clear. In this study, we showed that miR-223 may exert a significant role on the HCC cell chemoresistance mediated by ABCB1.
So far, there are few studies about the regulation of ABCB1 by miRNAs.8,20 For instance, miR-27 a and miR-451 are upregulated in ovarian cell lines resistant to multidrug and activate ABCB1 expression in the process of chemoresistance. 8 The over-expression of miR-200 c can downregulate P-gp and enhance the chemosensitivity to doxorubicin in human breast cancer cell MCF. 20 Additionally, miR-222 modulates the ADAM-17, a new MDR-related gene, in human colorectal carcinoma cells. 18 In the current study, we identified that ABCB1 was a direct target gene for miR-223, and miR-223 regulated the HCC cells’ response to doxorubicin and paclitaxel mediated by ABCB1.
ABCB1 is located on chromosome 7, and it belongs to the family of ABC transporter. It is responsible for reducing the accumulation of drugs in chemoresistant cancer cell lines and involved in the development of MDR. Previous studies showed that P-gp can restore the HCC cells, QGY-7703, resistant to a high concentration of therapeutic reagents. 25 Renal-cell carcinoma poorly responds to the traditional chemotherapy, and the high level of ABCB1 in renal carcinoma cells may be the contributor to the chemoresistance. 26 Accordingly, our study demonstrated that there existed a positive relationship between ABCB1 expression and IC50 dose of doxorubicin in different HCC cell lines, which suggested that ABCB1 may be related with the cell response to doxorubicin. Then, WST-1 assay indicated that miR-223 affected the cell-survival rate under doxorubicin and paclitaxel treatment, and ABCB1 can restore the cell-survival rate mediated by miR-223, which further validated that miR-223 participated in the regulation of chemoresistance to doxorubicin and paclitaxel mediated by ABCB1.
In our study, we pointed out that ABCB1 was regulated by several miRNAs, such as miR-451*, miR-200 b, miR-223 and miR-873. These candidate miRNAs were first validated by EGFP reporter assay. But miR-873 and miR-200 b had no effect on ABCB1 mRNA expression by real-time PCR analysis. For miR-223 and miR-451*, we found that these two miRNAs influenced ABCB1 mRNA and protein levels. Considering that there were few reports about miR-451* in cancers, miR-223 was chosen for further study. Moreover, we found that inhibition of miR-223 by ASO had no effect on the ABCB1 expression in SK-Hep-1 cell line. It may be due to that ABCB1 expression was very low, compared to other cell lines, even if miR-223 inhibition can increase ABCB1 expression, the endogenous level was relatively low. Although ABCB1 may be regulated by other miRNAs and miR-223 may modulate other genes besides ABCB1, our results, at least partially, indicate that miR-223 is involved in the regulation of chemoresistance mediated by ABCB1.
In addition, our study also explores the effect of miR-223 on IC50 of 5-Fu and cisplatin. We discovered that miR-223 affected the cisplatin IC50, but ABCB1 cannot restore the cisplatin IC50 mediated by miR-223. The reason may be that miR-223 regulated the cell response to cisplatin through other target genes except ABCB1. Both miR-223 and ABCB1 had no effect on 5-Fu IC50, suggesting that cell resistance to 5-Fu was regulated by other miRNAs and genes.
In conclusion, our study suggested that miR-223 played an important role in the regulation of MDR mediated by ABCB1. Importantly, miR-223 may serve as an important therapeutic biomarker for MDR in HCC patients.
Footnotes
Author contributions
Tao Yang wrote the manuscript, Zhi-min Zheng, Xiao-na Li, Zhen-fu Li, Yan Wang, Yun-feng Geng and Li Bai summarized and read the manuscript; Xian-bo Zhang and Tao Yang performed the experiments.
ACKNOWLEDGEMENTS
The work was supported by the Key Technology R&D Program of Hebei Province (No. 12277737).