
Abstract
The urogenital microbial infection in pregnancy is an important cause of maternal and neonatal morbidity and mortality. Uropathogenic Escherichia coli strains which express Dr fimbriae (Dr+) are associated with unique gestational virulence and they utilize cell surface decay accelerating factor (DAF or CD55) as one of the cellular receptor before invading the epithelial cells. Previous studies in our laboratory established that nitric oxide reduces the rate of E. coli invasion by delocalizing the DAF protein from cell surface lipid rafts and down-regulating its expression. The phosphoinositide 3-kinase/protein kinase B (PI3K/Akt) cell signal pathway plays an important role in host-microbe interaction because many bacteria including E. coli activate this pathway in order to establish infection. In the present study, we showed that the PI3K/Akt pathway negatively regulated the expression of DAF on the epithelial cell surface and thus inhibited the adhesion of Dr+ E. coli to epithelial cells. Initially, using two human cell lines Ishikawa and HeLa which differ in constitutive activity of PI3K/Akt, we showed that DAF levels were associated with the PI3K/Akt pathway. We then showed that the DAF gene expression was up-regulated and the Dr+ E. coli adhesion increased after the suppression of PI3K/Akt pathway in Ishikawa cells using inhibitor LY294002, and a plasmid which allowed the expression of PI3K/Akt regulatory protein PTEN. The down-regulation of PTEN protein using PTEN-specific siRNA activated the PI3K/Akt pathway, down-regulated the DAF, and decreased the adhesion of Dr+ E. coli. We conclude that the PI3K/Akt pathway regulated the DAF expression in a nitric oxide independent manner.
Introduction
The urogenital microbial infection (urinary tract infection or UTI) in pregnancy is an important cause of maternal and neonatal morbidity and mortality.1–3 The most common bacteria that colonize the urinary tract and cause UTI are the uropathogenic Escherichia coli. 4 The uropathogenic E. coli bearing Dr fimbriae (Dr+) have been associated with various pathologies such as UTIs in young adult women, chronic diarrhea in children, and chronic pyelonephritis in pregnant women.5–7 The epithelial cell adhesion and invasion of Dr+ E. coli has been shown to be facilitated by glycosylphosphatidylinositol (GPI)-anchored cell surface proteins of the host such as carcinoembryonic antigen (CEA)-related cell adhesion molecules (CEACAMs or CD66e) and decay accelerating factor (DAF or CD55).8–10 Dr adhesins are a conserved family of Gram-negative bacterial adhesion proteins and they are assembled via chaperone-usher pathway. 11 The biogenesis of Dr adhesins at the structural and molecular level is well known.12–15 However, the mechanisms of interactions between Dr+ E. coli and host cells that are crucial for bacterial pathogenesis are much less known. Previous studies in our laboratory have shown that nitric oxide (NO) down-regulates the DAF expression in a dose- and time-dependent manner in an endometrial epithelial cell line. The reduced DAF expression level was associated with reduced in vitro invasion of Dr+ E. coli suggesting that the down-regulation of DAF expression may be one of the mechanisms by which NO exhibits protective role against Dr+ E. coli infection during pregnancy. 16
DAF is a complement regulatory protein and is widely expressed on the surface of all the serum exposed cells including epithelial cells. DAF is bound to the plasma membrane by lipid raft associated GPI anchor. 17 Apart from its complement regulatory role, DAF has been implicated in many physiological and pathophysiological conditions. Expression of DAF in endometrium was found to be up-regulated during secretory phase and implantation, suggesting a role in immune tolerance of the implanting conceptus.18,19 The DAF deficiency has been associated with various diseases such as paroxysmal nocturnal hemoglobinurea, luteal phase defect, malarial anemia, psoriatic lesions, and apoptotic neutrophils.20–24 In contrast, higher levels of DAF expression are associated with tumorigenesis, decreased tumor cell lysis, and metastasis of many types of cancers.25,26
DAF expression levels may vary between tissues and under different physiological and pathophysiological conditions. 27 The DAF gene expression is regulated by transcription factors such as specificity protein 1 (Sp1) and cAMP response element-binding protein (CREB).17,28 One of the major cellular signaling cascades that activate these transcription factors is the phosphoinositide 3-kinase (PI3K) pathway.29,30 Therefore, we hypothesized that PI3K pathway may has a role in the regulation of DAF expression as well as adhesion of Dr+ E. coli to cells which express DAF.
The PI3K pathway is a major signaling cascade downstream of growth factor receptor tyrosine kinases. PI3K catalyzes the production of lipid second messengers such as phosphatidylinositol-3,4,5-triphosphate (PIP3) in the cell membrane. These phosphoinositides then activate phosphoinositide-dependent kinases (PDK) that allows the membrane recruitment of protein kinase B (PKB or Akt), a serine-threonine kinase which is the central molecule in the PI3K signaling pathway. PKB is then activated by PDK-dependent phosphorylation at two phosphorylation sites, amino acid residue threonine-308, and serine-473. 31 A lipid phosphatase, Tensin Homology Deleted on Chromosome Ten (PTEN) regulates the activity of PI3K/Akt by dephosphorylating phosphoinositides. 32 In this report, we examined the role of PI3K/Akt pathway in the regulation of DAF expression and the cellular adhesion of Dr+ E. coli using two different human genital tract epithelial cell line models, endometrial Ishikawa and cervical HeLa cells. We have chosen these two cell lines for this study because of their expression of DAF and the difference in the constitutive activity of the PI3K/Akt pathway. We show here that PI3K/Akt pathway negatively regulates the DAF expression and cellular adhesion of Dr+ E. coli.
Materials and methods
Cell culture
Ishikawa cells are a human endometrial cell line derived from a well-differentiated endometrial adenocarcinoma that has been shown to mimic endometrial epithelial cells.
33
Ishikawa cells were routinely cultured in Eagle's minimum essential medium (MEM) containing 2 mM
Antibodies and reagents
The DAF antibody (monoclonal antibody, cloneIH4) was a gift from D. M. Lublin, (Washington University School of Medicine, Seattle, WA, USA). The Akt (9272), pAkt-Thr-308 (9275), pAkt-Ser-473 (9271), PDK1(3062), pPDK1 (3485), PTEN (9552), and β-actin (4970) were purchased from Cell Signaling Technology (Danvers, MA, USA). The secondary anti-mouse antibody raised in goat was purchased from Molecular Probes (Grand Island, NY, USA). The horseradish peroxidase-conjugated anti-rabbit secondary antibody raised in donkey was obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The LY294002 was purchased from Cayman Chemicals (Ann arbor, MI, USA). The L-N-G-Nitroarginine methyl ester (L-NAME, N5751) was purchased from Sigma-Aldrich (St. Louis, MO, USA).
Transient transfections of cells
The PTEN plasmid was purchased from addgene (28298, Cambridge, MA, USA). Ishikawa cells were grown in 12-well cell culture plates for 24 h and the media were replaced by Opti-MEM reduced serum medium (31985070, Invitrogen, Carlsbad, CA, USA). The PTEN plasmid (7.8 µL) was diluted by mixing with 92.2 µL of Opti-MEM reduced serum medium to get a final concentration of 1 µg/mL. Similarly 2 µL of lipofectamine 2000 (11668019, Invitrogen) was diluted by mixing with 98 µL of Opti-MEM reduced serum medium and incubated at room temperature for 5 min. Diluted PTEN plasmid (50 µL) was then mixed with diluted lipofectamine 2000 (50 µL) and incubated at room temperature for 20 min. Lipofectamine-PTEN plasmid mixture (100 µL) was added per well and the cells were incubated at 37℃ for 48 h. For the controll cells, diluted lipofectamine (100 µL) prepared as described above was used.
The PTEN-specific siRNA (signal silence PTENsiRNA I) was purchased from Cell Signaling Technology (6251). The HeLa cells were grown in 12-well cell culture plates for 24 h and the media were replaced by Opti-MEM reduced serum medium. The PTEN siRNA (11.16 µL) was diluted by mixing with 88.84 µL of Opti-MEM reduced serum medium to get a final concentration of 1 µg/mL. Similarly, 4 µL of lipofectamine 2000 was diluted by mixing with 94 µL of Opti-MEM reduced serum medium and incubated at room temperature for 5 min. Diluted PTEN siRNA (50 µL) was then mixed with diluted lipofectamine 2000 (50 µL) and incubated at room temperature for 20 min. Lipofectamine-PTEN siRNA mixture (100 µL) was added per well and the cells were incubated at 37℃ for 48 h. For the controll cells, PTEN siRNA was replaced by a scrambled siRNA. After the transfection, cells were harvested by detergent lysis followed by scraping using a cell scraper for Western blot analysis.
Western blotting
Harvested cells were lysed on ice for 30 min with 200 µL lysis buffer (9803, Cell Signalling Technology) containing 5 µL each of Phosphatase Inhibitor Coctail 1, Phosphatase Inhibitor Coctail 2 and Protease Inhibitor (Sigma-Aldrich) supplemented with Pefablock SC to 1 mM. Lysates were collected by centrifugation (14,000 g) at 4℃ for 10 min. Protein concentrations in the cell lysates were determined by Pierce BCA protein assay kit (22660, Thermo Scientific, Rockford, IL, USA), against BSA standards. Proteins were seperated by polyacrylamide gel electrophoresis using NuPAGE 4-12% Bis-Tris mini gels (Invitrogen) with MOPS SDS as running buffer at 200 V for 50 min. Electrophoresis was carried out under reducing conditions except for DAF, for which non-reducing conditions were employed because the IH4 antibody used recognizes its epitope under non-reducing condition. The protein bands of interest were identified by Western blotting and visualized on a blue sensitive autoradiography film using supersignal west pico chemiluminiscence substrate (32132, Thermo Scientific) according to the instructions provided by the manufacturer. The films were scanned and the protein band densitometric analysis was performed using the AlphaEase Flurochem 8000 software (Alpha Innotech, Santa Clara, CA, USA).
RNA isolation and real-time quantitative polymerase chain reaction
Total RNA was isolated from the cells using RNeasy mini kit (74104, Qiagen, Valencia, CA, USA) according to the procedure recommended by the manufacturer. RNA extraction was followed by DNase I (Qiagen) treatment to remove DNA contamination. Total RNA of 1 µg was reverse transcribed into complementary DNA using avian myeloblastosis virus reverse transcriptase (M5101, Promega, Madison, WI, USA) and random oligonucleotide hexamers (N8080127, Invitrogen). Quantitative real-time polymerase chain reaction (PCR) was carried out using CFX96 system and SYBR green master mix (Biorad, Hercules, CA, USA). A comparative cycle of threshold fluorescence (CT) method was used with housekeeping gene as internal control.
Proximity ligation assay
The proximity ligation assay (PLA) is a PCR-based immuno-technique for the visualization of in situ protein expression, protein–protein interactions, and posttranslational protein modifications. The in situ expression of DAF proein on the cell surface was visualized and quantified by PLA. The Duolink II kit used for the PLA was purchased from Olink Bioscience (Uppsala, Sweden). The HeLa or Ishikawa cells sparsley grown in 16-well lab-Tek chamber slides (Electron Microscopy Sciences, Hatfield, PA, USA) were transiently transfected with PTEN-specific siRNA and PTEN plasmid, respectively, for 24 h. A scrambled siRNA or tranfection medium served as controls. The cells were washed twice with phosphate-buffered saline (PBS) and then fixed using 4% paraformaldehyde. The PLA was then performed according to the instructions provided by the duolink II kit manufacturer.
Bacterial adhesion to the cells
For E. coli adhesion study, we used a recombinant strain (the laboratory E. coli K-12 strain carrying plasmid pBJN406). 34 The Ishikawa and HeLa cells were grown in 8-well lab-Tek chamber slides (Electron Microscopy Sceinces) and transiently transfected with PTEN plasmid and PTEN siRNA, respectively, for 24 h. The media were removed and the cells were rinsed with PBS. Then, bacterial suspensions in 500 µL of PBS, prepared as described previously35,36 for the adherence assay, were added to the monolayers of Ishikawa and HeLa cells, incubated at 37℃ for 3 h. The cells were then rinsed with PBS three times, and the bacteria adhered to the cells were visualized by gram staining.
Statistical analysis
GraphPad Prism (version6) software was used for statistical analysis. Comparisons between the groups were made by t-test. Statistical significance (p < 0.05) between the groups was determined by Bonferroni's multiple comparison method.
Results
Adhesion of E. coli expressing Dr+ adhesin to epithelial cells was associated with constitutive activity of PI3K-Akt pathway
The PI3K/Akt pathway is constitutively active in endometrial epithelial cell line Ishikawa because of a truncated mutation in PTEN gene.
37
In cervical epithelial cell line HeLa, functional PTEN is expressed and therefore, the PI3K/Akt pathway is regulated.
38
Therefore, we used these two cell lines to evaluate the role of PI3K/Akt pathway in the expression of DAF protein and the adhesion of E. coli that express Dr+ adhesin. Initially, we confirmed the expression levels of PTEN in HeLa and Ishikawa cells and then analyzed the expression levels of DAF, PDK1, Akt, phosphor PDK1 (pPDK1), and phospho-Akt [pAkt (Ser473) and pAkt (Thr308)] proteins by Western blotting. The PTEN was expressed only in HeLa cells whereas the total PDK1 and total Akt were expressed in both the Ishikawa and the HeLa cells (Figure 1a). However, Western blotting followed by densitometric analysis revealed that pPDK1 levels were significantly higher in the Ishikawa cells compared to that of HeLa cells (Figure 1b and c). The pAkt (Ser473) and pAkt (Thr308) were detected only in the Ishikawa cells (Figure 1d). Taken together, these results confirmed that the PI3K/Akt pathway was regulated in HeLa cells and constitutively active in Ishikawa cells. The Western blot analysis of DAF protein levels revealed that its expression was dramatically higher in the HeLa cells compared to that of Ishikawa cells, suggesting an association between the PI3K/Akt pathway and the expression of DAF protein (Figure 1e). This result led us to analyze the difference in adhesion of Dr+ E. coli to the HeLa and the Ishikawa cells. When Dr+ E. coli were allowed to bind to the monolayers of HeLa and Ishikawa cells in vitro, we found that the number of bacteria adhered to individual HeLa cells was significantly higher compared to that of Ishikawa cells (Figure 1f). These data suggested a possible association between PI3K/Akt regulated DAF expression and Dr+ E. coli adhesion to the epithelial cells.
The association between constitutive activity of PI3K/Akt pathway and adhesion of Dr+ E. coli to epithelial cells. The expression of PI3K/Akt pathway proteins in HeLa and Ishikawa cell grown as monolayers for 24 h was examined by Western blot. The E. coli K-12 strain carrying the plasmid pBJN406, encoding the production of Dr fimbriae was allowed to bind to HeLa and Ishikawa cells for 3 h and then the number of bacteria adhered to individual cells was examined by gram staining. (a) Western blot for PTEN, total PDK1, and total Akt proteins in HeLa and Ishikawa cells. (b) Western blot for pPDK1 protein in HeLa and Ishikawa cells. (c) Densitometric analysis of pPDK1 protein levels. *p < 0.0001 versus HeLa. (d) Western blot for activated forms of Akt, pAkt(ser473) and pAkt(Thr308). (e) Western blot for DAF. (f) Gram staining to visualize E. coli K-12 strain carrying plasmid pBJN406 adhered to HeLa and Ishikawa cells. Data are representative images or expressed as mean values ± SEM of each group from three separate experiments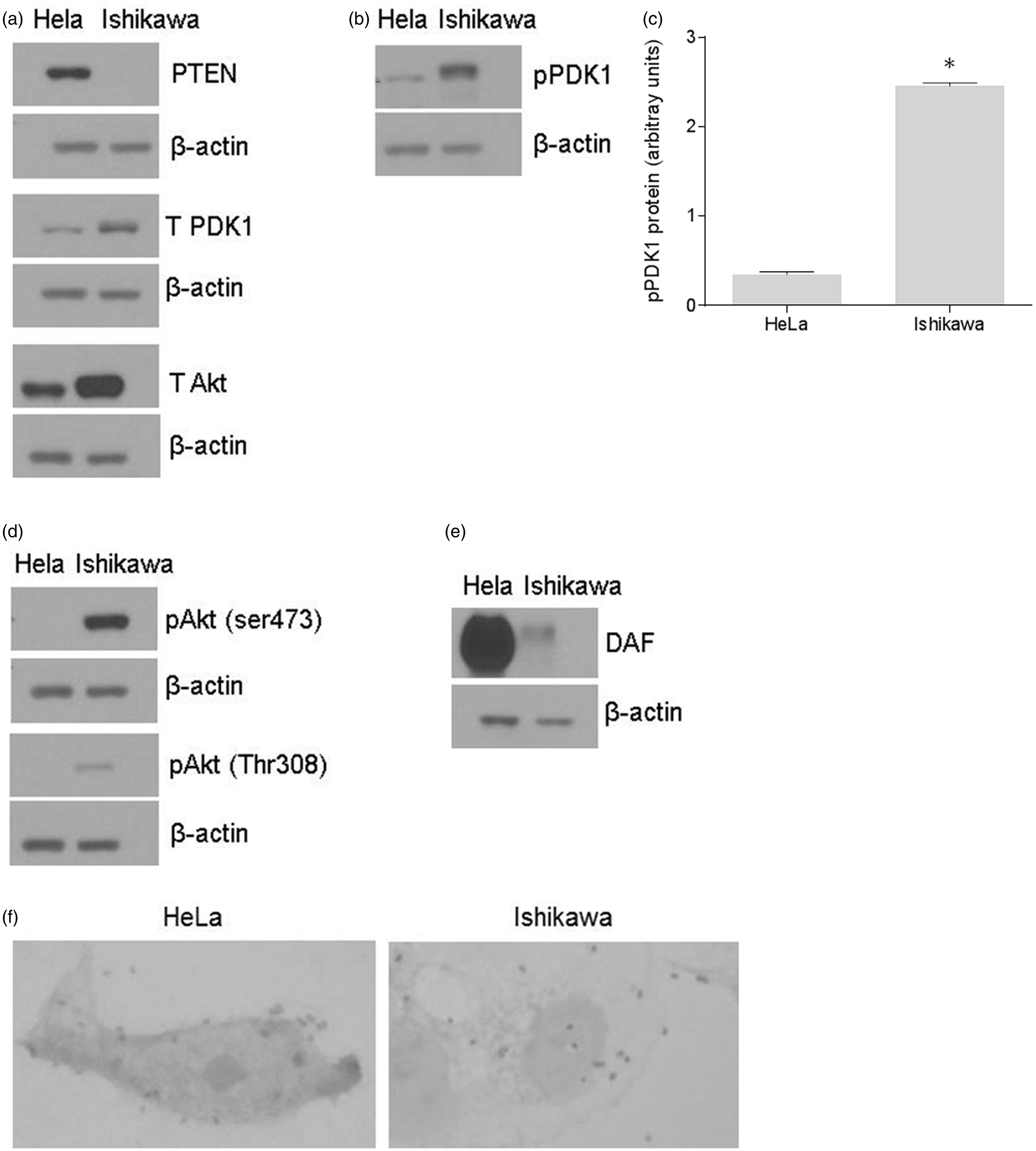
Pharmacological inhibitor of PI3K/Akt pathway up-regulated the expression of DAF mRNA and protein
In order to estbliash a direct evidence for the link between PI3K/Akt pathway and the DAF gene expression, we inhibited PI3K/Akt pathway in Ishikawa cells using pharmacological inhibitor LY294002. The Western blot followed by densitometric analysis of Akt and its phosporylation after treating Ishikawa cells with LY294002 for 24 h revealed that level of phosphorylation at both Ser473 and Thr308 reduced siginficantly compared to that of untreated cells (Figure 2a–c). Then we analyzed the effect of LY294002 on the expression of DAF gene in Ishikawa cells. As shown in Figure 2(d), the real-time quantitative PCR analysis revealed that DAF mRNA expression increased significantly in LY294002 treated cells compared to that of control cells. Furthermore, Western blot and densitometric analysis showed an increase in DAF protein expression after LY294002 treatment compared to controll cells (Figure 2e and f). These data suggested a direct inverse relation between PI3K/Akt pathway and the DAF gene expression.
Up-regulation of DAF by pharmacological inhibitor of PI3K/Akt pathway. The monolayer of Ishikawa cells was treated with LY294002 (50 μM) dissolved in DMSO or DMSO alone (control) for 24 h. (a) Western blot for total Akt and its activated forms, pAkt(ser473) and pAkt(Thr308). (b) Density analysis of pAkt(ser473) Western blot expressed as ratio between total Akt and pAkt(ser473). *p< 0.0001 versus control. (c) Density analysis of pAkt(Thr308) Western blot expressed as ratio between total Akt and pAkt(Thr308). *p < 0.01 versus control. (d) Fold increase in the expression of DAF mRNA as assessed by real-time qPCR after treating the Ishikawa cells with the LY294002. *p < 0.001 versus control. (e) Western blot for DAF protein. (f) Density analysis of DAF Western blot expressed as fold increase in the expression after treating Ishikawa cells with LY294002. p < 0.03 versus control. Data are representative images or expressed as mean values ± SEM of each group from three separate experiments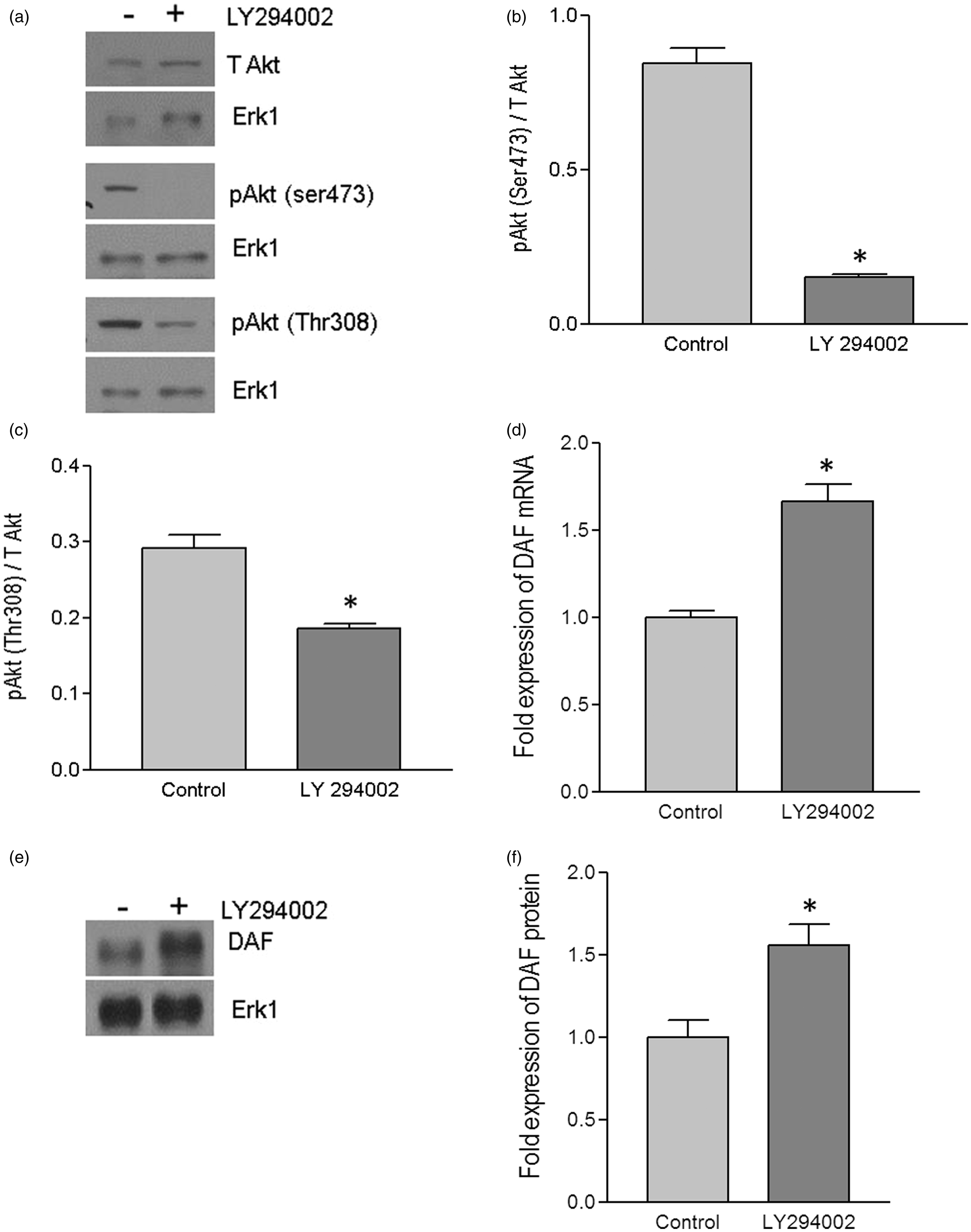
Introduction of a plasmid expressing PTEN into Ishikawa cells up-regulated DAF expression and increased Dr+ E. coli adhesion
To further confirm the role of PI3K/Akt in DAF gene expression and Dr+ E. coli adhesion, we sought to manipulate the PI3K/Akt pathway by altering the expression of PTEN in Ishikawa and HeLa cells. Initially, we introduced a mammalian expression plasmid for PTEN into the Ishikawa cells. The introduction of PTEN plasmid allowed the Ishikawa cells to overexpress PTEN protein (Figure 3a and b). Overexpression of PTEN protein significantly reduced the level of phosphorylation in PDK1 and Akt (Figure 3c–f). To assess whether this negative regulation of Akt activity due to the overexpression of PTEN protein had an effect on DAF gene expression, we performed real-time quantitative PCR and Western blot followed by densitometric analysis. As shown in Figure 3(g)–(i), the expression of DAF mRNA and protein increased significantly after the overexpression of PTEN protein in Ishikawa cells.
Up-regulation of DAF and increase in Dr+ E. coli adhesion after the expression of PTEN in Ishikawa cells. Ishikawa cells were transiently transfected with plasmid encoding functional PTEN gene. For control, cells were treated with equivalent amount of transfection regent lipofectamine 2000. After 24 h of transfection, cells were either lysed for Western blot and real-time qPCR or E coli. K-12 strain carrying plasmid pBJN406, encoding the production of Dr fimbriae was allowed to bind to the cells for 3 h. The number of bacteria adhered per cell was counted after the gram staining. Similarly, Ishikawa cells sparsely grown on 16-well lab-Tek chamber slides were transfected as mentioned above, fixed using formalin and proximity ligation assay was performed. (a) Western blot for PTEN expression. (b) Density analysis of Western blot indicating the fold increase in the PTEN protein expression. *p < 0.05 versus control. (c) Western blot for total PDK1, total Akt, and their activated forms, pPDK1, pAkt(ser473), and pAkt(Thr308). (d) Western blot density analysis of pPDK1 protein levels expressed as ratio between pPDK1 and total PDK1. *p < 0.05 versus control. (e) Western blot density analysis of pAkt(ser473) expressed as ratio between pAkt(ser473) and total Akt. *p < 0.05 versus control. (f) Western blot density analysis of pAkt(Thr308) expressed as ratio between pAkt(Thr308) and total Akt. *p < 0.001 versus control. (g) Fold increase in DAF mRNA as assessed by real-time qPCR. *p < 0.0001 versus control. (h) Western blot for DAF protein expression. (i) Density analysis of Western blot indicating the fold increase in the DAF protein expression. *p < 0.001 versus control. (j) PLA assay for the expression of DAF protein on the surface of Ishikawa cells. (k) The DAF protein levels on the surface of Ishikawa cells as assessed by PLA assay, expressed as number of signals per nucleus. *p < 0.01 versus control. (l) The adherence of E. coli K-12 strain carrying the plasmid pBJN406 to the Ishikawa cells as revealed by gram staining. (m) The average number of bacteria adhered per cell. *p < 0.01 versus control. Data are representative images or expressed as mean values ± SEM of each group from three separate experiments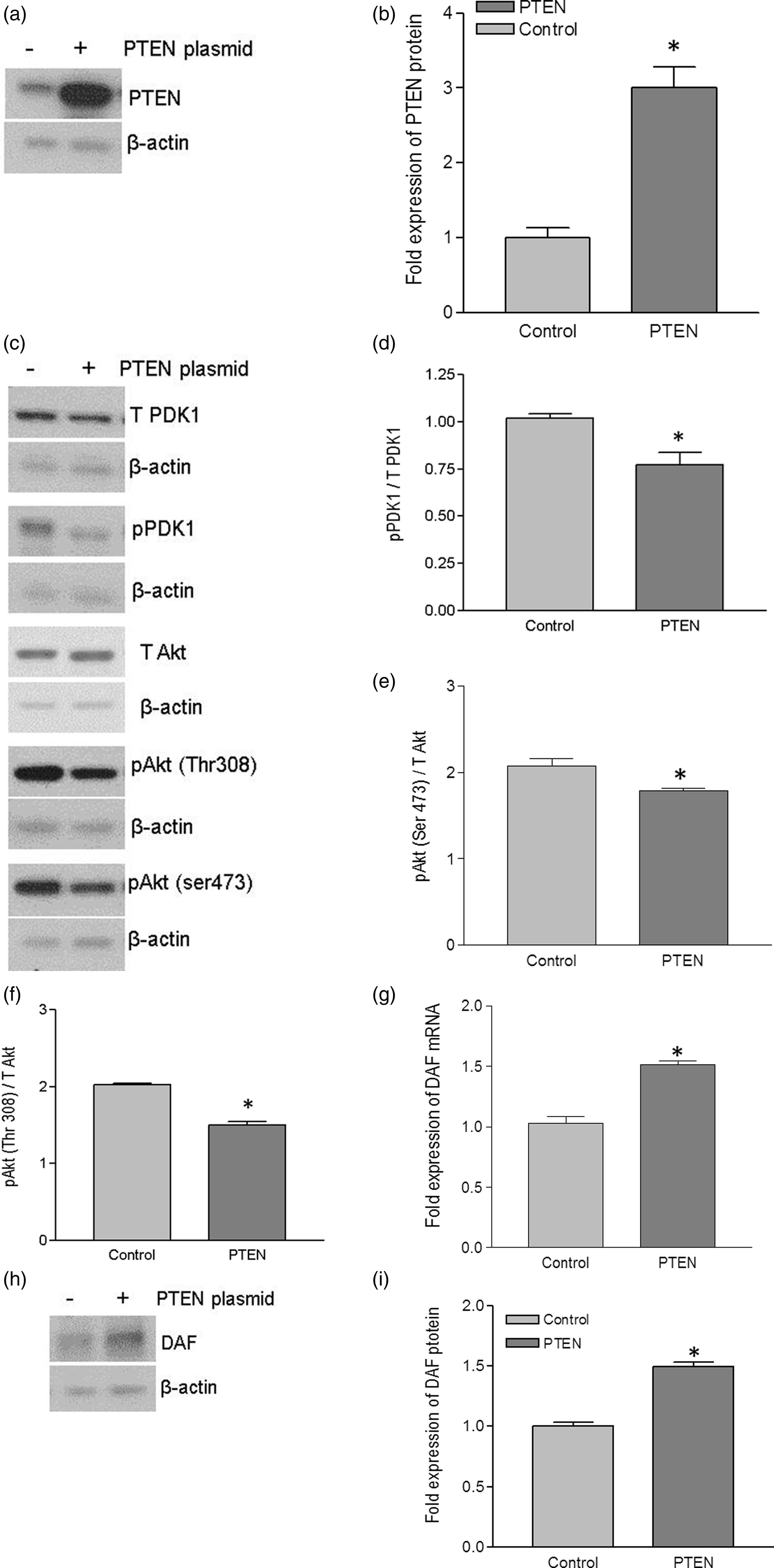
Although the qPCR and Western blot provided the information on expression levels of DAF, we wanted to know the changes in cellular localization of DAF protein. The DAF protein is a membrane bound protein which is anchored to the cell surface through GPI molecule and thus it is available on the cell surface for the bacterial ligand to bind before establishing the infection. Therefore, we sought to determine whether up-regulation of DAF protein increased DAF protein levels on the cell surface by performing PLA. The PLA revealed that the cell surface expression of DAF significantly increased in the Ishikawa cells transfected with PTEN plasmid compared to that of control cells (Figure 3j and k). Furthermore, when the Dr+ E. coli were allowed to bind to Ishikawa cells for 3 h, their adhesion to individual Ishikawa cells significantly increased after the introduction of PTEN plasmid (Figure 3l and m). Overall, theses data indicated that introduction of a plasmid vector for the expression of PTEN into Ishikawa cells up-regulated DAF expression and increased Dr+ E. coli adhesion.
Down-regulation of PTEN using siRNA in HeLa cells down-regulated DAF expression and decreased Dr+ E. coli adhesion
We further confirmed the inverse association between PI3K/Akt pathway and the DAF expression by using a PTEN-specific siRNA. When HeLa cells were transfected with PTEN-specific siRNA, expression of PTEN protein was reduced significantly (Figure 4a and b). The suppression of PTEN expression led to an increase in the phosphorylation of PDK1 and Akt (Figure 4c–f) compared to the control cells. As shown in Figure 4(g)–(i), PTEN down-regulation also induced a decreased expression of DAF mRNA and protein compared to the control cells. Furthermore, the PLA showed a decrease in the DAF protein levels on the cell surface after the siRNA mediated PTEN suppression in the HeLa cells (Figure 4j and k). The adhesion of Dr+ E. coli to HeLa cells also decreased significantly after the transfection with PTEN-specific siRNA (Figure 4i and m). Taken together, these results confirmed that PI3K/Akt pathway negatively controls the expression of DAF on the cell surface and thus plays an important role in the adhesion of Dr+ E. coli.
Down-regulation of PTEN using PTEN-specific siRNA down-regulated DAF expression and decreased Dr+ E. coli adhesion in HeLa cells. The HeLa cells were transiently transfected with PTEN-specific siRNA. For control, cells were treated with a scrambled siRNA. After 24 h of transfection, cells were either lysed for Western blot and real-time qPCR or E coli. K-12 strain carrying plasmid pBJN406, encoding the production of Dr fimbriae was allowed to bind to the cells for 3 h. The number of bacteria adhered per cell was counted after the gram staining. Similarly, HeLa cells sparsely grown on 16-well lab-Tek chamber slides were transfected as mentioned above, fixed using formalin and proximity ligation assay was performed. (a) Western blot for PTEN expression. (b) Density analysis of Western blot indicating the fold decrease in the PTEN protein expression. *p < 0.01 versus control. (c) Western blot for total PDK1, total Akt, and their activated forms, pPDK1, pAkt(ser473) and pAkt(Thr308). (d) Western blot density analysis of pPDK1 protein levels expressed as ratio between pPDK1 and total PDK1. *p < 0.05 versus control. (e) Western blot density analysis of pAkt(ser473) expressed as ratio between pAkt(ser473) and total Akt. *p < 0.001 versus control. (f) Western blot density analysis of pAkt(Thr308) expressed as ratio between pAkt(Thr308) and total Akt. *p < 0.001 versus control. (g) Fold decrease in DAF mRNA as assessed by real-time qPCR. *p < 0.01 versus control. (h) Western blot for DAF protein expression. (i) Density analysis of Western blot indicating the fold decrease in the DAF protein expression. *p < 0.05 versus control. (j) PLA assay for the expression of DAF protein on the surface of HeLa cells. (k) The DAF protein levels on the surface of HeLa cells as assessed by PLA assay, expressed as number of signals per nucleus. *p < 0.01 versus control. (l) The adherence of E. coli K-12 strain carrying the plasmid pBJN406 to the HeLa cells as revealed by gram staining. (m) The average number of bacteria adhered per cell. *p < 0.05 versus control. Data are representative images or expressed as mean values ± SEM of each group from three separate experiments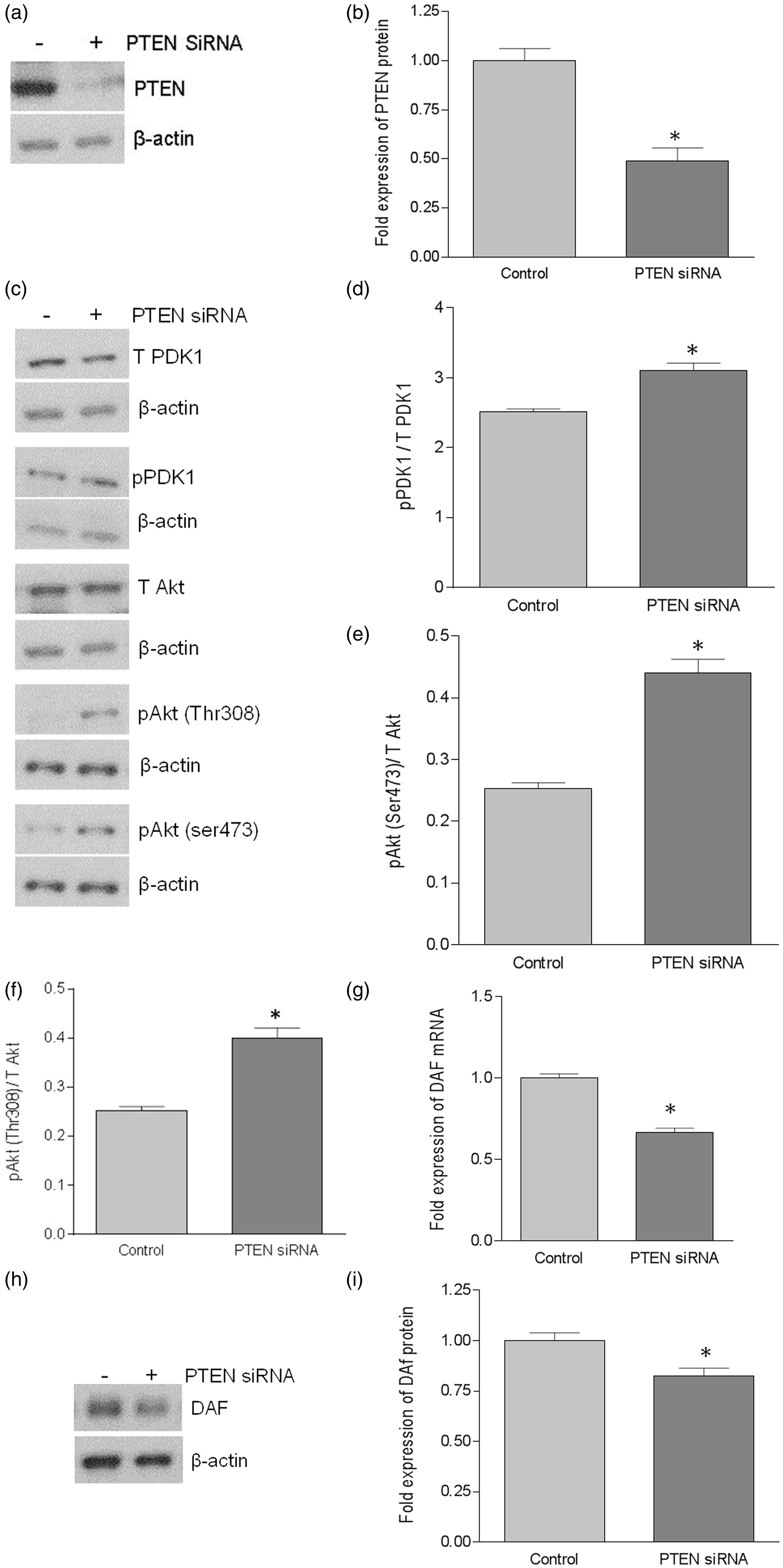
Effect of PI3K/Akt pathway on DAF expression was not mediated by NO
Earlier studies in our laboratory demonstrated that NO down-regulates the expression of DAF. Therefore, we sought to examine whether negative regulation of DAF expression by PI3K/Akt pathway was mediated through NO. In order to study the possible role of NO, we inhibited the endogenous synthesis of NO in PTEN knock-down HeLa cells by treating the cells with NOS inhibitor L-N-G-Nitroarginine methyl ester (L-NAME) for 24 h. The transfection with PTEN-specific siRNA reduced the expression of DAF mRNA in HeLa cells compared to that of control cells as revealed by real-time quantitative PCR. However, inhibition of NO syntehesis using L-NAME did not reverse the effect of PI3K/Akt activation (Figure 5a) suggesting that NO was not mediating the effect of PI3K/Akt on DAF expression.
The effect of PI3K/Akt pathway on DAF expression was not mediated by NO. The HeLa cells were transiently transfected with PTEN-specific siRNA or a scrambled siRNA (control) for 24 h and then treated with either nitric oxide synthase inhibitor L-NAME or NO donor DETA NONOate for 24 h followed by cell lysis for real-time qPCR and Western blot, respectively. (a) Fold change in the expression of DAF mRNA in HeLa cells. *p < 0.0001. (b) Western blot for total PDK1, total Akt, and their activated forms, pPDK1, pAkt(ser473), and pAkt(Thr308) in Ishikawa cells. (c) Western blot density analysis for total PDK1, total Akt, and their activated forms, pPDK1, pAkt(ser473) and pAkt(Thr308) in Ishikawa cells. *p < 0.05 versus control, **p < 0.001 versus control, †p < 0.05 versus control, ††p < 0.05 versus control, ‡ p < 0.05 versus control. Data are representative images or expressed as mean values ± SEM of each group from three separate experiments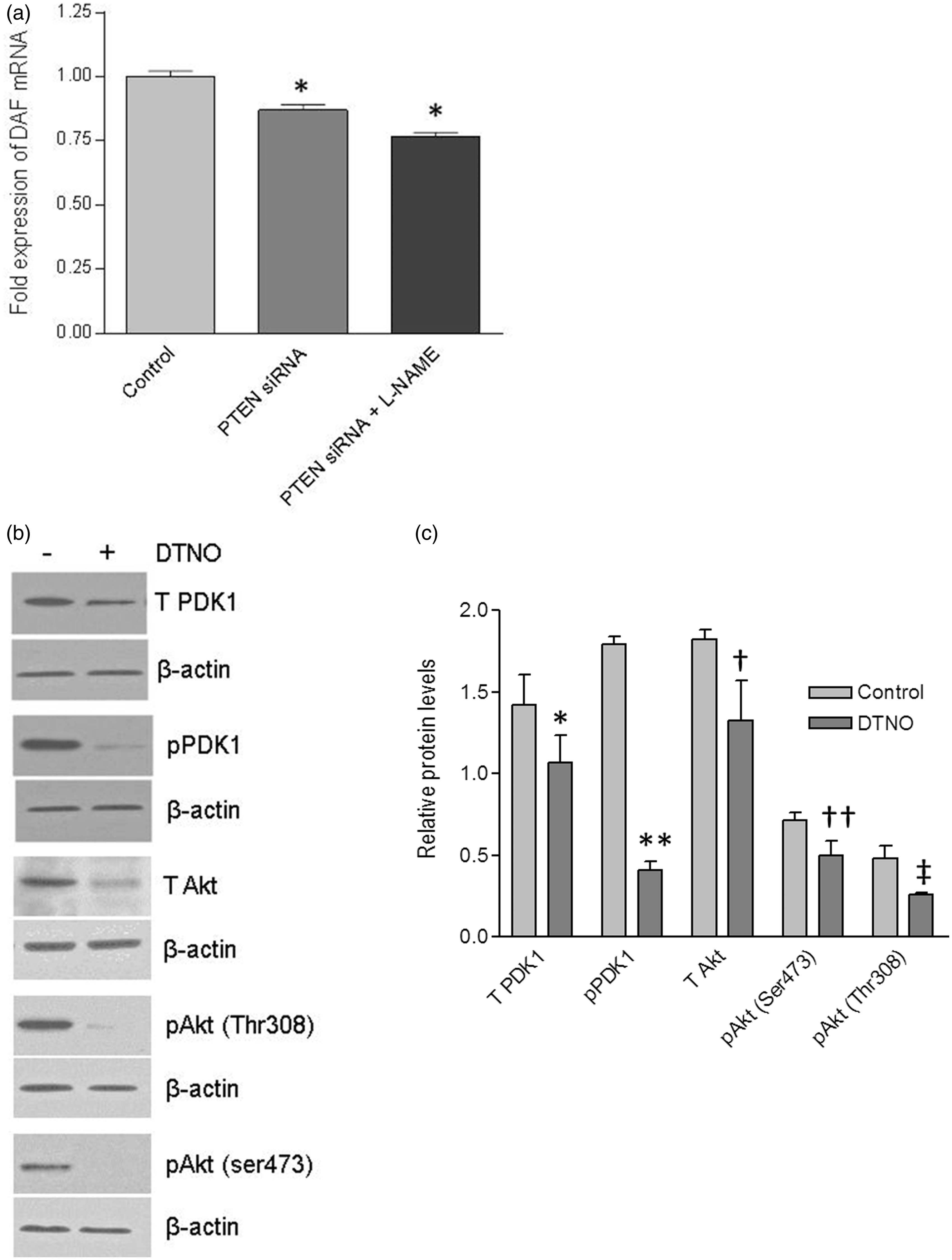
Then we asked whether the NO can trigger the PI3K/Akt pathway in Ishikawa cells to down-regulate the DAF, because NO activates PI3K/Akt pathway in bovine and human endothelial cells. 39 Previously we observed that in Ishikawa cells, NO donor diethylenetriamine (DETA) NONOate (DTNO) down-regulated the expression of DAF at a concentration of 0.1 mM or higher. 16 Therefore, in order to examine the effect of NO on the PI3K/Akt pathway, we treated the Ishikawa cells with 1 mM of DTNO for 24 h. As shown in the Figure 5(b) and (c), the Western blot followed by densitometric analysis revealed that total PDK1 and total Akt protein expressions decreased significantly after DTNO treatment. Correspondingly, the levels of pPDK1 and pAkt also decreased significantly in DTNO treated cells compared to that of control cells. Together, these data demonstrated that NO reduced the PI3K/Akt signaling by down-regulating the pathway proteins, PDK1 and Akt.
Inhibition of NO synthesis using NOS inhibitor, L-NAME failed to reverse the effect of PI3K/Akt activation in HeLa cells, prompting us to accept that NO was not mediating the effect of PI3K/Akt pathway on the DAF protein expression. Instead, we observed down-regulation of PDK1 and Akt by NO, which could reduce the signaling and consequently ought to up-regulate the DAF expression. However, our previous in vitro and in vivo experiments showed a significant down-regulation of DAF by NO.16,40 This contradiction could be due to a more direct effect of NO on DAF gene expression because, NO decreased the DAF mRNA transcription and half-life by inhibiting the binding of transcription factor Sp1 to 5′-untranslated region of DAF gene and HuR protein to 3′-UTR of DAF mRNA, respectively. 41
Discussion
The adherence of bacteria to mucosal surfaces is an important step in the pathogenesis of most infections in humans and animals. 42 The pathogenic E. coli strains express on their surfaces adhesins and invasins which are responsible for the recognition and binding of specific membrane-bound host molecules acting as receptors. Among pathogenic E. coli strains, uropathogenic and diarrheagenic E. coli express Dr-family of adhesins such as Afa, Dr, and F1845. 43 The major receptors on the host cells recognized by these adhesins are type IV collagen, DAF, and CEACAMs.43,44 In this report, we have shown that PI3K/Akt pathway negatively regulated the expression of DAF in the epithelial cells and thus restricted the attachment of E. coli that expressed Dr adhesin.
Although, bacterial adhesins can bind to a number of receptors on the host cell, DAF-specific pathogens are known to cause chronic recurrent infections. The Dr/Afa family of adhesins includes at least 13 E. coli adhesins involved in human diseases. 43 Some of these adhesins exhibit narrow specificity toward the host receptors. Except AfaE-VII and AfaE-VIII, all other Afa/Dr adhesins bind to DAF.45–47 However, collagen binding phenotype is unique to the Dr adhesins whereas only a subfamily of Afa/Dr adhesins designated Afa/Dr-I (Afa/DrCEA) bind to the CEACAM family of proteins.48,49 Although binding of Dr adhesin to type IV collagen is critical for the development of persistent renal infection, role of type IV collagen in the development of intestinal infection is not clear. The DAF rather than type IV collagen is present at the apical surface of the polarized intestinal epithelial cells where E. coli tend to colonize.49,50 DAF is expressed in various parts of digestive, urogenital, and respiratory tracts.49,51 In the digestive tract, Helicobacter pylori, which causes chronic gastritis attaches to the DAF protein on gastric epithelial cells.52,53 Coxsackieviruses and Enteroviruses also utilize DAF as receptor on the host cell surface.49,54–56 Therefore, pathogenicity due to microbial or viral ligands and DAF interactions appear to be involved in the development of protracted often subclinical bacterial and viral diseases.
Bacterial pathogens manipulate several host cell kinase signaling such as nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB), mitogen-activated protein kinase (MAPK), and PI3K/Akt to exploit down-stream effector functions in order to establish intracellular infection.
57
PI3K/Akt pathway is an essential signaling pathway in the process of phagocytosis and therefore, that allows bacterial pathogens to hijack PI3K/Akt signaling either to induce their uptake by host cells through a process similar to phagocytosis or to inhibit their phagocytosis by immune cells such as macrophages. Yersinia species inject a phosphatase to disrupt focal adhesion complex-mediated activation of PI3K/Akt pathway in macrophages and thus inhibit their phagocytosis.
58
Shigella flexneri and Pseudomonas aeruginosa activate the host cell PI3K/Akt pathway to induce their epithelial uptake by inducing actin reorganization in host cells.
58
Adherent E. coli strains also have been reported to manipulate PI3K/Akt pathway in order to establish infection. Peiffer et al.
59
reported that in an in vitro intestinal cell model, a clinically isolated strain of E. coli C 1845 that expressed F1845 adhesin induced F-actin reorganization in a PI3K/Akt-dependent manner. However, role of host cell PI3K/Akt pathway in the protection against bacterial invasion is not clear. In this report, we have provided the evidence to show that PI3K/Akt pathway could be involved in restricting the attachment of Dr+ E. coli to epithelial cells. PI3K/Akt pathway is essential for pathogenic E. coli to gain entry in to host cells whereas PI3K/Akt signaling activated by ligand-receptor interactions in host cells down-regulates DAF and restricts their attachment as proposed in Figure 6. The regulation of PI3K/Akt signaling in the physiological context is complex and therefore, further studies are required to establish overall role of PI3K/Akt pathway in Dr+ E. coli infection. However, we suggest that restriction of DAF expression could contribute to limit the level of virulence of Dr+ E. coli and therefore, may direct infection process toward chronic subclinical infection in contrast to acute inflammatory diseases caused by other pathogenic bacteria such as Shigella species.
Pictorial representations for the role of PI3K/Akt pathway in Dr+ E. coli infection. (a) Pathogenic bacteria such as Shigella, Pseudomonas, and Group B streptococcus bind to host cells and activate PI3K/Akt pathway that allows them to induce actin reorganization resulting in their internalization. Yersinia species inhibit PI3K/Akt pathway and thus restrict their phagocytosis by macrophages.
58
E. coli bearing F1845 adhesins bind to DAF on host cell surface and activate PI3K/Akt pathway by an unknown mechanism causing actin reorganization and their internalization.
59
The induced internalization of these pathogenic bacteria produces an acute infection process. (b) Activity of PI3K/Akt pathway is regulated by several ligands such as hormones through different transmembrane receptors. Expression of DAF gene is negatively regulated by PI3K/Akt pathway. NO also down-regulates DFA expression by inhibiting its transcription and destabilizing mRNA
41
. Thus, both PI3K/Akt pathway and NO reduce the expression of DAF on host cell surface resulting in restricted binding of Dr+ E. coli. Restricted binding of E. coli may help in establishing a symbiotic type of relation where both are protected from a lethal outcome thus leading to a chronic type of infection. (A color version of this figure is available in the online)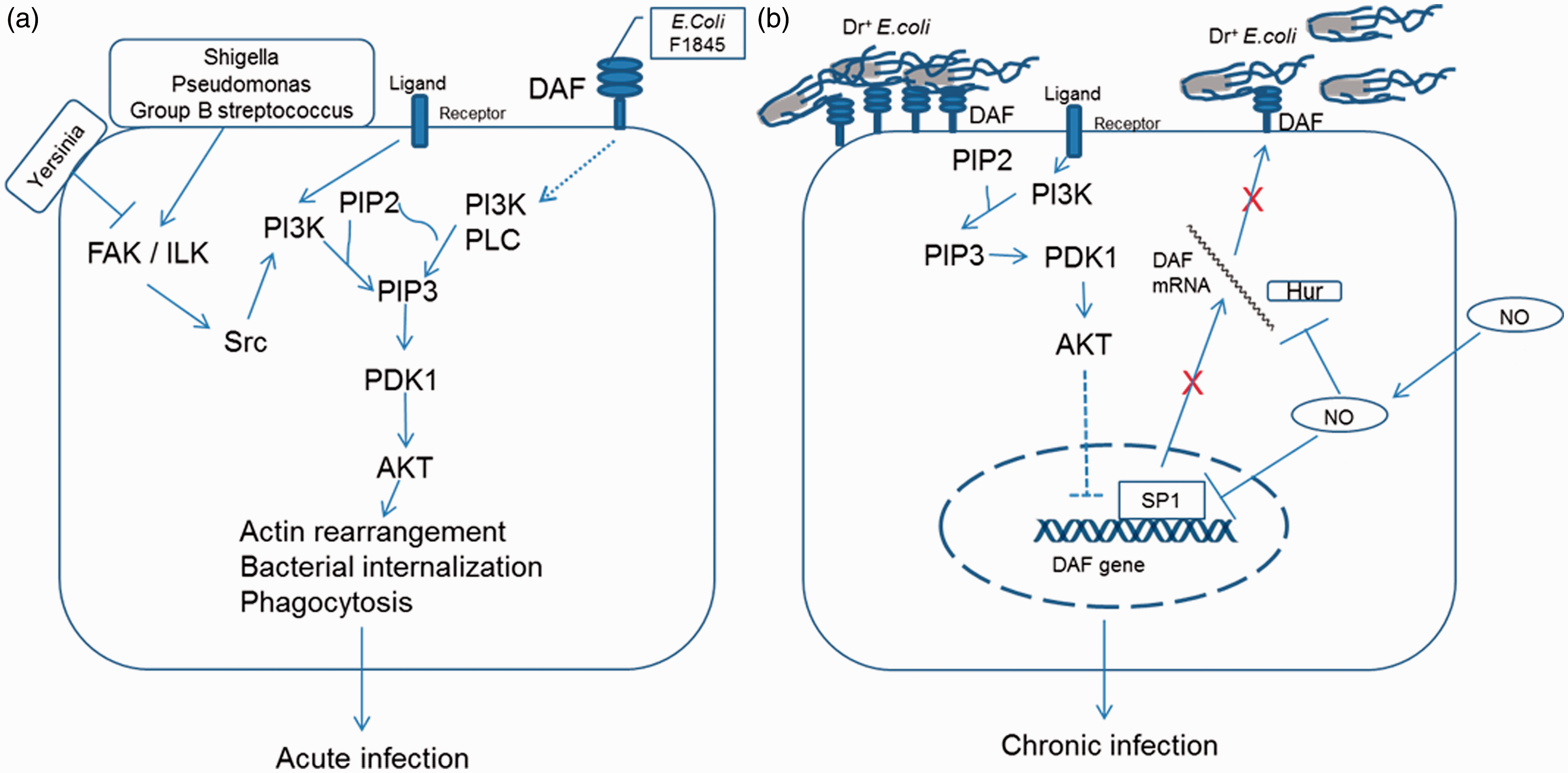
In summary, the present study provides evidence for the association between the PI3K/Akt-mediated variations in the DAF expression levels on the epithelial cells and the adhesion of Dr+ E. coli to epithelial cells. The involvement of two independent host systems, NO and PI3K/Akt in the down-regulation of DAF in the context of Dr+ E. coli pathogenesis appears to be consistent with the host-pathogen strategy to achieve well-controlled limited infection level. Restricted DAF expression may therefore, lead to chronic instead of acute infection thus leading to a symbiotic host-pathogen status, protecting both from a lethal outcome.
Footnotes
Author contributions
MB, PG, and CY participated in the design and interpretation of the data; MB and DL conducted the experiments; MB wrote and CY and BJN reviewed the manuscript; and BJN and SN provided the plasmids pBJN406 and protocol for the preparation of bacteria for adhesion assay.
ACKNOWLEDGEMENTS
This work was supported by the National Institute of Health (grant number HL72620, HD57013).