
Abstract
Lung cancer, which can be divided into two major clinical–pathological categories, small cell lung cancer and non-small cell lung cancer, is the leading cause of cancer-related death worldwide. MicroRNAs (miRNAs), small non-coding RNAs approximately 22 nucleotides in length, have been reported to be upregulated or downregulated in disease states and specific cell types. Recently, miRNAs have gained recognition as major regulators of human gene expression. MiRNAs can control highly complex signal transduction pathways and other biological pathways by targeting and controlling gene expression, accounting for their important role in lung cancer. Findings from recent studies on the roles of miRNAs in lung cancer are summarized in this review. Understanding miRNA functions in lung cancer will bring molecular-level insight leading to better prognosis, diagnosis, and therapeutic approaches.
Introduction
MicroRNAs (miRNAs) are highly conserved noncoding RNAs about 18–24 nucleotides (nt) long involved in gene regulation.
1
Initially generated as long primary transcripts, miRNAs are then processed by the RNase III-like enzyme Drosha
2
and the double-stranded RNA-binding domain protein DGCR8 (Pasha in Caenorhabditis elegans)
2
into shorter precursors (pre-miRNAs) that are about 60–80 nt long and have a hairpin structure. The pre-miRNAs are exported from the nucleus to the cytoplasm by exportin 5,
3
a member of the nuclear transport receptor family.4–6 In the cytoplasm, double-stranded miRNAs that are approximately 22 nt in length are generated by another RNase III enzyme, Dicer.7–12 The RNA duplex is loaded onto an Argonaute (Ago) protein to generate the effector complex, called the RNA-induced silencing complex (RISC). An individual miRNA is capable of regulating dozens of distinct transcripts and together numerous miRNAs are believed to modulate more than one-third of the mRNA species encoded in the genome.
13
miRNAs are involved in critical biological processes, including development, differentiation, apoptosis, and proliferation,1,14 through imperfect pairing with target messenger RNAs (mRNAs) of protein-coding genes and the transcriptional or post-transcriptional regulation of their expression.15–18 Dysregulation of miRNAs is often related to cancer inhibition or progression, suggesting that miRNAs may act as tumor suppressor genes or oncogenes (Figure 1).
19
In this review, we summarize recent work in molecular genetics concerning the diagnostic and therapeutic potential of miRNAs in human lung cancer.
MicroRNA biogenesis and function in cancer. After canonical animal microRNA (miRNA) genes are transcribed by RNA polymerase II (Pol II) to generate the primary transcripts (pri-miRNAs), the Drosha/DGCR8 enzyme complex generates precursor miRNAs (pre-miRNAs) in the nucleus. The pre-miRNAs are exported into the cytoplasm in concert with Exprotin 5, after which the cytoplasmic RNase III Dicer cleaves them to produce miRNA duplexes. Finally, one strand of the duplex is incorporated into the RNA-induced silencing complex to enable inhibition of translation of the target mRNA. The reduction or deletion of an miRNA that functions as a tumor suppressor leads to tumor formation. A reduction of mature miRNA levels leads to the inappropriate expression of the target oncoprotein. The overall outcome might involve increased proliferation, invasiveness, or angiogenesis, or decreased levels of apoptosis, finally leading to tumor formation. The amplification or overexpression of a miRNA with an oncogenic role may also result in tumor formation. Increased amounts of an oncogenic miRNA can eliminate the expression of the target tumor suppressor gene and lead to cancer progression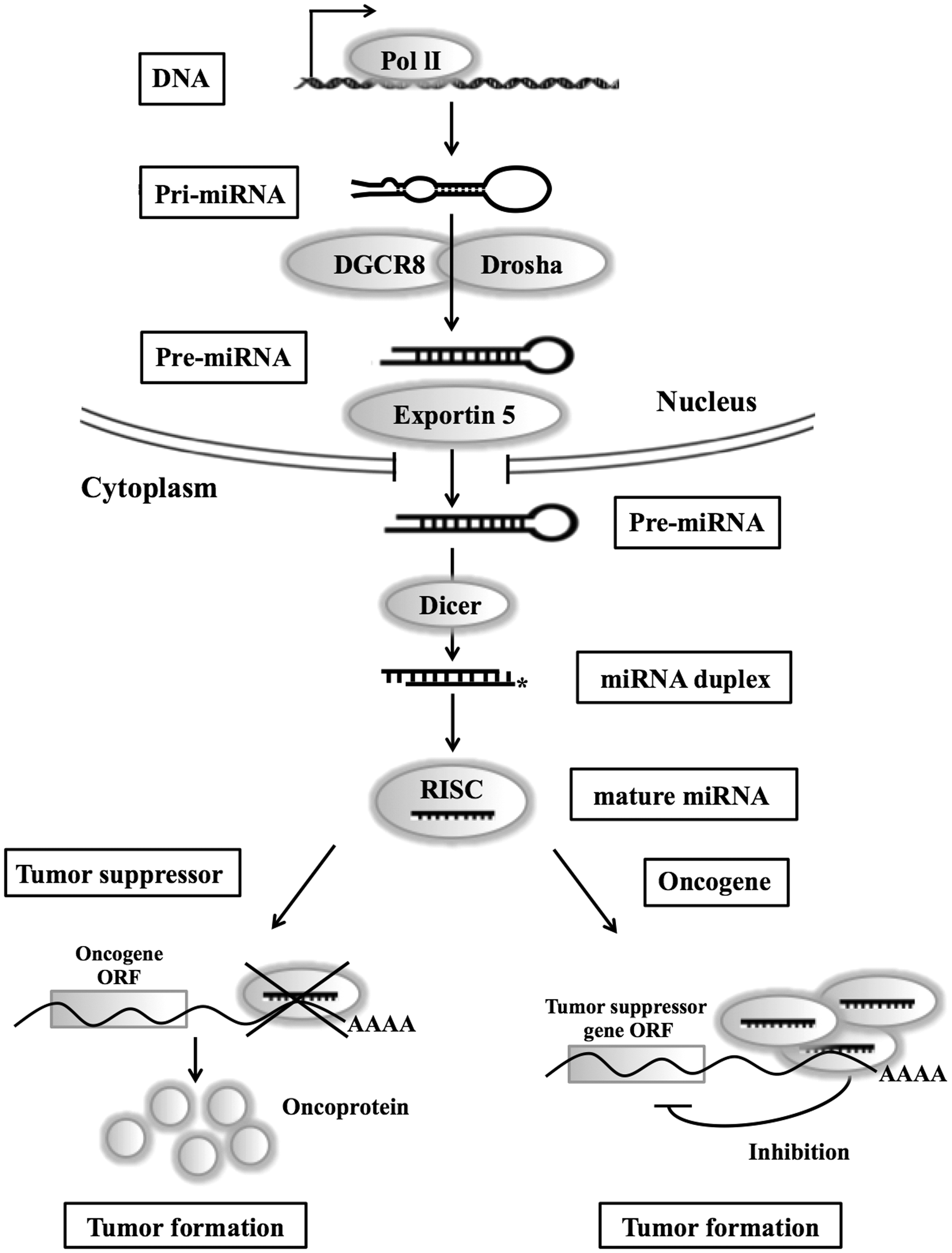
Pathogenesis of lung cancer
Lung cancer is a disease of uncontrolled cell growth in the tissues of the lung. Lung cancer still has the highest mortality and the lowest survival rate of all types of solid cancers. 20 It is the leading cause of cancer death in males and the second leading cause of cancer death in females. 20 According to cancer statistics reported in 2012, the five-year survival rate of patients diagnosed with lung cancer showed the least increase, along with liver, pancreatic, and stomach cancers. 21 Lung cancer pathogenesis is closely associated with tobacco smoking.
Lung cancers are classified histologically according to the size and malignancy of the cells involved in tumor development. Generally, lung cancer can be divided into two main categories: small cell lung cancer (SCLC) and non-small cell lung cancer (NSCLC). NSCLC, which accounts for approximately 80% of all lung cancers, comprises squamous cell carcinoma (20–25% of lung cancers), large cell carcinoma (15–20% of lung cancers), and adenocarcinoma (30–40% of lung cancers).22,23 Adenocarcinoma is the predominant histological subtype of lung carcinoma. Furthermore, adenocarcinomas most often originate at peripheral sites in the lungs and are the most common subtype in individuals who have never smoked.24,25 Squamous cell lung cancers usually start from the central bronchus and are most closely correlated with smoking history. Small cell carcinomas comprise a smaller portion of lung cancers (approximately 16%) and have a tendency to arise from cells in the large airways. 26 Given the heterogeneous origin and nature of lung cancers, biomarker development should be tailored to the different lung cancer types and stages.
Many miRNAs are also extensively involved in cancer pathogenesis. Studies have identified an important role of miRNAs in the regulation of the cell cycle and carcinogenesis in different organs, including the lung.27,28 The first discovery of miRNA dysregulation in lung cancer was reported by Volinial et al., who identified a group of miRNAs frequently aberrantly expressed in tumor tissues with respect to normal tissue counterparts. 29
miRNA expression and genetics in lung cancer
Cancer is a complex genetic disease involving structural and expression abnormalities of both coding and noncoding genes. The alteration of protein-coding oncogenes and/or tumor suppressor genes have been thought to be the causes of tumorigenesis.30,31 Therefore, many studies have been trying to elucidate the molecular mechanisms that cause cancer development and cancer prevention. Although several oncogenes and tumor suppressor genes have been identified in human and/or other model animal genomes, the mechanism of cancer formation is yet to be identified. A recently identified class of miRNAs may provide new insight in cancer research. More than 50% of miRNA-encoding loci reside in chromosomal regions altered during tumorigenesis, 32 and expression profiling reveals characteristic miRNA signatures for many tumor types that predict disease status and clinical outcome. 33
The expression level of miRNAs changes have subsequently been detected by different groups in many types of human tumors.19,34–37 miRNAs have been contributed to tumorigenesis because they can function either as tumor suppressors or oncogenes. The effect of identification of genomic abnormalities is the activity of miRNAs, they are worked the same as previously described for protein-coding genes, such as chromosomal rearrangements, genomic amplifications or deletions and mutations. The abnormalities both in protein-coding genes and miRNAs in cancer status are identified. 38
The biological consequences of disrupted miRNA expression are revealed by a wide range of cellular or organismal phenotypes, including cancers.29,39 In the lungs, miRNAs play critical roles in both development and tumorigenesis.40–43 Hundreds of miRNAs are dynamically regulated during normal lung organogenesis in both mice and humans.44,45 The overall expression profile of miRNAs is similar in both mouse and human lungs,45,46 suggesting evolutionary conservation of miRNA expression. Twenty-three of the 30 most highly expressed miRNAs have been detected in the adult lungs of both mice and humans, miR-26a, miR-99, members of the let-7 family, the miR-29 family, and the miR-30 family being expressed highly in both species. miR-134, miR-154, miR-214, miR-296, miR-299, miR-323, miR-337, and miR-370 are highly expressed in both the neonatal mouse and the fetal human lungs, and subsequently decline in adult lung. 45 In contrast, expression of miR-26b, miR-29a/b, miR-146-3p, and miR-187 is increased in the adult lungs of both mice and humans. These results may contribute to the development of a novel therapeutic approach for treating diseases that result in dysregulation of normal development or homeostasis, such as lung cancer.
It has been shown that miRNAs may mutate or change in expression (over- or underexpression) in human cancer. 29 The report showed that 35 miRNAs are upregulated and only three miRNAs are downregulated in lung tumors compared with normal tissues. 29 In a comparison of miRNA expression in human lung cancer patients’ tissues and noncancerous lung tissues, 43 miRNAs showed statistical differences, with 15 miRNAs upregulated in the lung cancer tissues. 47 The differences in expression patterns between lung cancer samples and normal control did not show the same trend in these two studies, possibly due to differences in the samples, technologies used, or even the analytical methods used. Taken together, miR-21, miR-24, and miR-210 were upregulated, whereas miR-30a-5p, miR-30d, miR-126*, and miR-143 were downregulated in lung cancer tissues compared with healthy controls.
The mechanisms regulating miRNA expression and maturation are largely unknown, but studies have suggested the involvement of several mechanisms, including genetic and epigenetic alterations.48,49 The role of miRNA single nucleotide polymorphism (SNP) in disease has been only recently identified. SNPs have been identified in miRNAs important in cancer. The expression of miR-125a is altered in breast cancer, 50 this miRNA having a variant allele due to an SNP in the mature miRNA sequence that decreases expression. 49 Patients with NSCLC who carry a variant homozygote of rs11614913 located in the 3p mature miRNA region of miR-196a2 have poor survival, possibly due to a mechanism of elevated expression of mature miR-196a and weakened target binding of miR-196a2-3p. 51 In addition, polymorphisms in the rs2910164 of miR-146a and the rs11614913 of miR-196a2 are positively associated with a better recurrence-free survival in stage II and stage III patients with completely resected NSCLC. 52 SNPs in miRNA binding sites may also be associated with disease: a point mutation identified in the 3′ untranslated region (UTR) of SLITRK1 in several patients with Tourette’s syndrome disrupted the binding of miR-189. 53 Furthermore, the let-7 miRNA binding site SNP in KRAS is significantly associated with increased risk for NSCLC among moderate smokers and represents a new paradigm for let-7 miRNAs in lung cancer susceptibility. 54
Recent studies have also demonstrated the importance of epigenetic modulation of miRNAs. In lung carcinogenesis through the p53 tumor suppressive pathway, expression of the miR-34 family has been reported to be regulated by DNA methylation. miR-34b/c promoter hypermethylation was associated with a high probability of recurrence and correlated with poor overall survival and low disease-free survival in stage I NSCLC. 55 In addition, DNA methylation of miR-34b can serve as a biomarker for an invasive phenotype of NSCLC. 56 The frequency of miR-34b/c methylation was significantly higher in SCLC than in NSCLC. Consequently, miR-34b/c expression was decreased in SCLC, causing tumor cell growth, migration, and invasion. 57 Patients with stages I-III NSCLS had tumor-specific miR-9-3 and miR-193a methylation. The patients with miR-9-3 methylated lung squamous cell carcinoma (LSCC) had a statistically significant shorter disease-free survival as well as shorter overall survival than patients with miR-9-3 unmethylated LSCC. miR-9-3 methylation is a prognostic parameter in patients with LSCC. 58 miR-212 was highly downregulated in NSCLC patients. The transcriptional silencing of miR-212 in NSCLC involves H3K27me3/H3K9Ac or H3K9me3/H3K9Ac-associated histone modification. 59 miR-29 can universally alter methylation status through targeting of DNA methyltransferases 3A and B (DNMT3A and 3B), leading to dysregulation of the critical tumor suppressors fragile histidine triad protein (FHIT) and WW domain-containing oxidoreductase (WWOX). 60 High expression of HDACs has been reported in several cancer tissues, including stomach, colorectal, kidney, breast, and lung.61–65 Though the mechanism of highly expressed HDAC1 in lung cancer remains undetermined, the down expression of miR-449a/b, which regulates HDAC1 by directly binding with its 3′ UTR, might be one such mechanism. Considering that miR-449a/b inhibits cell growth and anchorage-independent growth, it may have a tumor suppressor function, making it potentially valuable in lung cancer therapy. 66
Many recent miRNA studies discussed here involving the polymorphism and epigenetics of miRNAs offer fresh insights into early diagnostics and potential therapies for human lung cancer.
Functions of miRNAs in lung cancer
Lung cancer-related miRNAs and their potential target genes and functions (details in the text)
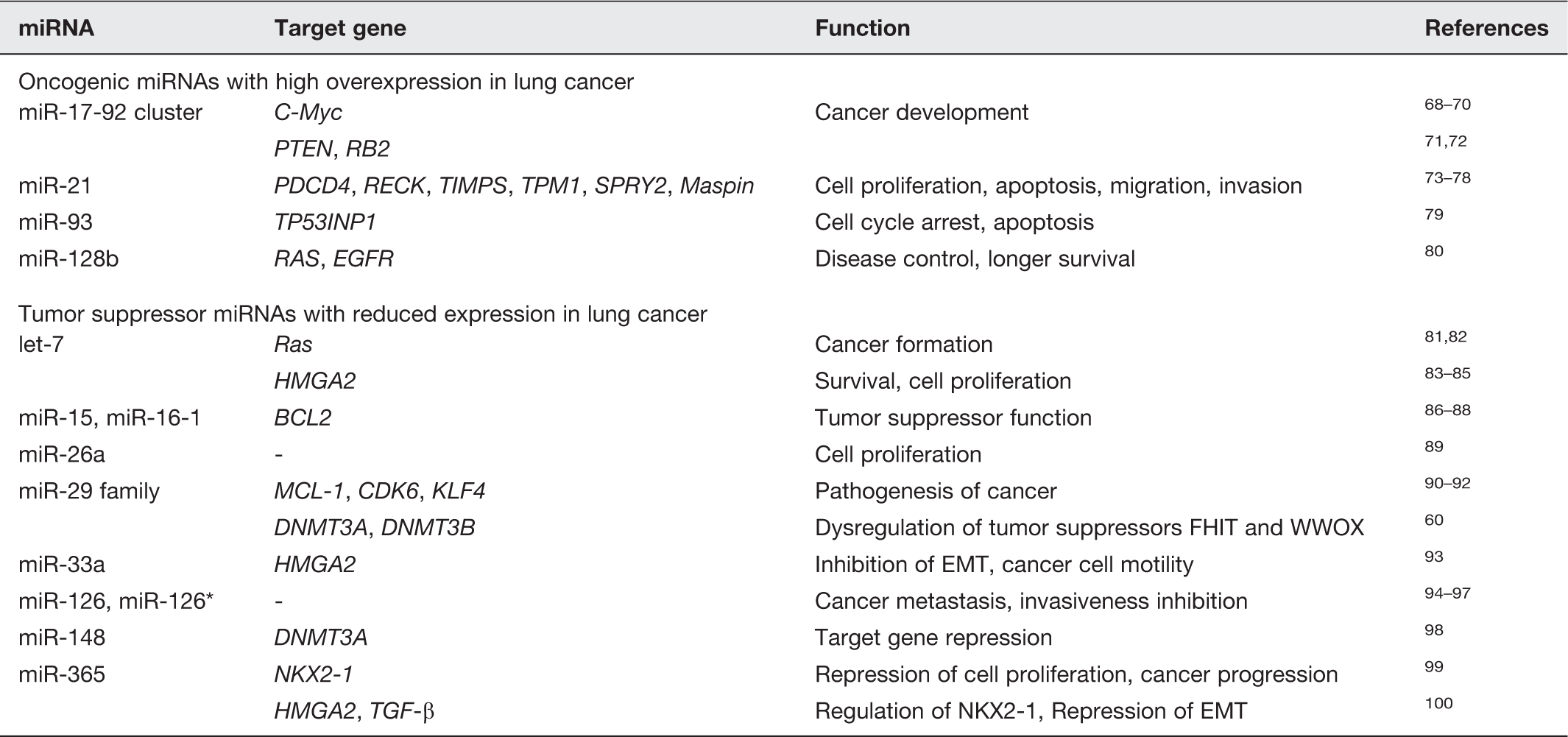
miR-21 is specifically overexpressed in a variety of cancers: glioblastoma, lung cancer, stomach cancer, prostate cancer, colon cancer, and breast cancer.29,47,101–104 Especially in NSCLC, miR-21 has been causally linked to cellular proliferation, apoptosis, migration, and invasion, with prognostic significance.73,74 Knockdown of miR-21 in MDA-MB-231 (metastatic breast cancer cells) significantly reduces invasion and lung metastasis. 73 miR-21 is known to target several genes, including programmed cell death 4 (PDCD4), RECK, metallopeptidase inhibitors (TIMPs), tropomyosin 1 (TPM1), Sprouty homolog 2 (SPRY2), and mammary serine protease inhibitor (Maspin).73–78 Ectopic expression of miR-93 in a lung carcinoma cell line (H1299) reduces tumor protein p53-inducible nuclear protein 1 (TP53INP1), which could lead to cell cycle arrest and apoptosis. 79 miR-128b was recently found to be associated with the overexpression of RAS and epidermal growth factor receptor (EGFR). 80 miR-128b was demonstrated to directly regulate EGFR, its loss having a favorable impact on sensitivity to EGFR tyrosine kinase inhibitors (TKIs) that was comparable to EGFR copy number gain and associated with significantly improved disease control and longer survival.
Several miRNAs, including let-7, miR-15, miR-26, miR-29, miR-30, miR-126, and miR-142, show decreased expression in lung cancer compared with normal lung. Their expression is also reduced in the serum of lung cancer patients. Interestingly, the expression levels of these miRNAs increase during embryonic and/or neonatal lung development. The let-7 was the first miRNA found to be downregulated in lung cancer, 81 in which the 3′ UTR of the human RAS gene contains various let-7 complementary sites and the expression of let-7 related genes is 50% lower than in healthy tissue.81,82 Loss of the let-7 control of RAS expression could lead to RAS overexpression and contribute to the formation of a human cancer. Kumar et al. showed that let-7g-expressing NSCLCs have reduced expression of RAS and high mobility group AT-hook 2 (HMGA2) proteins. 105 HMGA2 is targeted by let-7 and its expression is inversely associated with survival in patients with lung and gastric cancers.83,84 In a lung cancer cell, ectopic expression of let-7 decreases HMGA2 and cell proliferation. 85 It has been shown that the 3′ UTR of human HMGA2 harbors multiple let-7 complementary sites. 85 Silencing of HMGA2 by let-7 enhances differentiation, but produces no effect on self-renewal in tumor-initiating cells. 106 miR-15a and miR-16-1, which are encoded by a region on chromosome 13q14.3, are lost or downregulated in cancer and show a tumor suppressor function by targeting the B-cell lymphoma 2 (BCL2) oncogene.86–88 miR-26a has the ability to diminish the proliferation of MYC-dependent cells. 89 The miR-29 family (miR-29a, miR-29b, and miR-29c) has also been implicated in the pathogenesis of cancer. Expression of the miR-29 family is reduced in tumors and has been shown to target several key components of cellular survival: MCL-1, an induced myeloid leukemia cell differentiation protein; CDK6, a cell cycle regulator protein; and the dedifferentiation modulator Kruppel-like factor 4 (KLF4).90–92 miR-148 represses DNMT3A expression through a region in its open reading frame, 98 and miR-126 and miR-126* can inhibit cancer metastasis and invasiveness in lung cancer as well as breast and prostate cancer patients.94–97
Recent work has focused on identifying recurrent genetic alterations associated with the development and progression of malignancy. Previous studies in NSCLC revealed frequent focal DNA amplifications at chromosome 14q13.3,
107
an area containing genes with potential roles in lung cancer, including NK2 homeobox 1 (NKX2-1).107–109 Within the lung, NKX2-1 expression is confined mainly to the peripheral airways and small bronchioles.110,111 The oncogenic and suppressive function of NKX2-1 in the same tumor type substantiates its role as a dual function lineage factor. Interestingly, miRNAs have been found that regulate NKX2-1. Qi et al. and our previous study found that miR-365 can regulate NKX2-1 in human lung cancer.99,100 Another study found that NKX2-1 directly upregulates miR-33a by binding to the promoter of SREBF2, the host gene of miR-33a, which then represses the HMGA2 (Figure 2).
93
Loss of heterozygosity (LOH) of chromosome 3p is one of the most frequent genetic events in lung carcinogenesis. The loss of the gene encoding miR-128b (located on chromosome 3p) is correlated with the response to targeted inhibition of EGFR.
80
LOH of miR-128b can be considered equivalent to losing a tumor suppressor gene because it permits increased production of EGFR. miR-128b is a regulator of EGFR in NSCLC cell lines, studies having determined that miR-128b directly regulates EGFR.
80
miR-128b LOH is common in tumor samples and is significantly correlated with the clinical response to and survival after gefitinib (EGFR inhibitor) therapy. Yu et al. identified a signature of five miRNAs (let-7a, miR-221, miR-137, miR-372, and miR-182) that predict treatment outcome in NSCLC patients.
43
Patients with a high risk score for these five miRNAs had an increased relapse rate and shortened survival times.
Schematic summary of NKX2-1 regulation. A model for NKX2-1 regulation in lung cancer. miR-365 mediates the TGF-β-dependent suppression of NKX2-1. Note that a positive feedback loop exists between miR-365 and TGF-β signaling and that miR-365 directly regulates NKX2-1 as well as targeting an epithelial–mesenchymal transition (EMT)-promoting gene, HMGA2. NKX2-1 directly upregulates miR-33a expression, after which miR-33a represses HMGA2. NKX2-1 represses lung cancer progression by suppressing cancer cell motility, proliferation, and EMT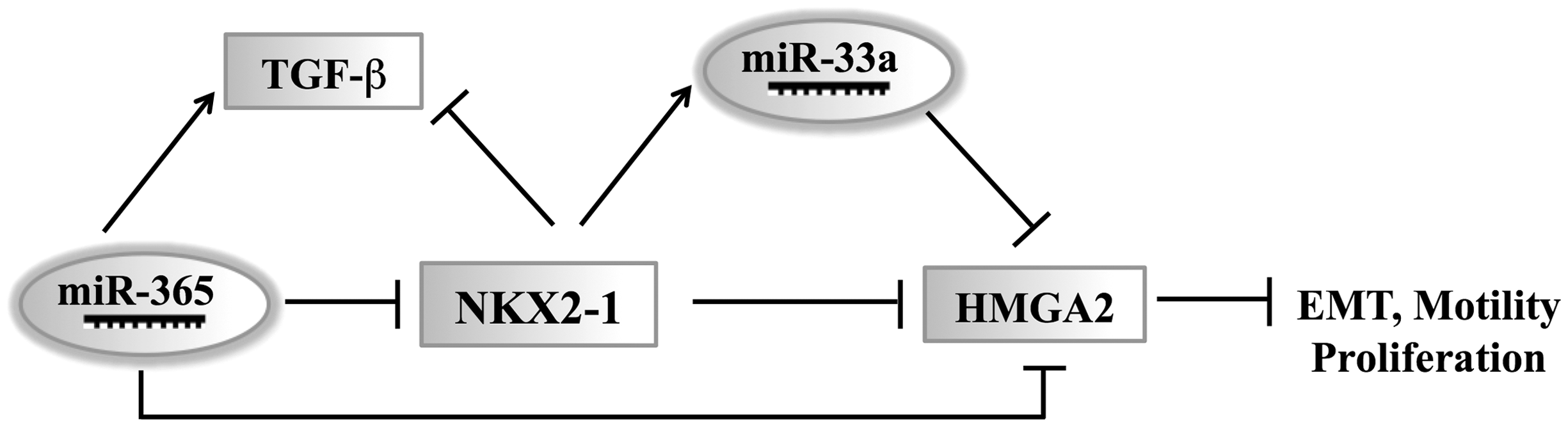
Many dysregulated miRNAs in lung cancer are located at the frequently deleted or amplified regions. 86 Both miR-21 and miR-205 are located at the region amplified in lung cancer, whereas miR-126 and miR-126* are located at 9q34.3, a region deleted in lung cancer. Rapid progress in functional studies of miRNAs together with these results will promote our understanding of lung cancer cause and cure.
Serum miRNAs as biomarkers of lung cancer
The low survival rate for lung cancer may be attributed to the low rate of diagnosis in early stages, making early diagnosis a great imperative. Compared with other cancers, lung cancer has few subjective symptoms and there are few screening methods for lung cancer diagnosis. At present, diagnosis of lung cancer still largely depends on imaging techniques such as X-rays, computed tomography (CT) scans, and magnetic resonance imaging (MRI). 112 Recent advances have improved both the sensitivity and accuracy of low-dose spiral CT. 113 However, despite such advances in imaging diagnostic technology, problems such as economic burden, exposure to radiation, high misdiagnosis rates, and poor early diagnosis rate remain.
Recently identified cancer biomarkers are being used in attempts to improve the diagnosis, prognosis, and therapeutic monitoring of lung cancer using genomic and proteomic techniques. Biomarkers that could indicate the cancer status of individuals are varied and include physiological markers, visual characteristics, genes, and proteins. 114 Among the many kinds of biomarkers, miRNA biomarkers are currently being investigated in conjunction with the development of techniques such as miRNA microarrays, miRNA qRT-PCR, and deep sequencing.115–117
miRNAs are of particular importance in cancer biology because many have been shown to be subject to amplification or deletion, a hallmark of the cancer genome. 32 Furthermore, miRNAs are better classifiers of tissue origin for cancer cell lines or tumor tissues than are mRNA biomarkers, having been used to trace the tissue origin of cancers of unknown primary origin. 118 Moreover, miRNA signatures have been used to define molecular subsets of tumors29,119 and predict outcome.32,47,120 Many attributes of miRNAs, such as their tissue specificity,121–123 high stability, and ability to regulate multiple mRNA targets, make them attractive as a novel class of biomarkers in lung cancer. In particular, many reports have begun identifying circulating miRNAs as cancer biomarkers. 124
Blood samples are good sources of biomarkers because their use would allow the development of noninvasive biomarker assays; miRNAs in blood have important physiological functions and secreted miRNAs can be used to assess physiological conditions.124–126 The relative stability of serum miRNAs compared to mRNAs also supports their use as cancer biomarkers (serum miRNAs are resistant to RNase A digestion). In addition, miRNAs have proved stable in serum under harsh conditions such as boiling, low and high pH, extended storage, and freeze–thaw cycles.124–126 These independent studies, which have demonstrated that miRNAs are released into the body fluid and exist there in a sufficiently stable form, suggest a new approach utilizing circulating miRNAs for the noninvasive detection of human cancer. The first comprehensive analysis of miRNAs in serum from NSCLC patients was performed by Chen et al. using deep sequencing technology. 125 In the study, the miRNA profile in the pooled serum from patients with NSCLC was compared with the profile from healthy controls. Twenty-eight miRNAs were missing and 63 new miRNAs were detected in the serum from the lung cancer patients when compared with the healthy subjects, 8 serum miRNAs being uniquely detected in lung cancer patients. 125 Two NSCLC-specific miRNAs (miR-25 and miR-223) were further validated in an independent trial sample set consisting of serum from 152 lung cancer patients and 75 healthy controls; qRT-PCR confirmed significantly higher expression of both miRNAs in the sera of lung cancer patients compared to those of healthy controls.
Following these initial discoveries, numerous studies have been performed to identify miRNAs with diagnostic relevance in body fluids such as plasma, serum, sputum, and lung effusions. Many private companies involved in the development of miRNA-based diagnostic tests have paid great attention to blood-based noninvasive early detection methods due to some advantages of miRNAs over protein-based early detection systems, including their potential for detection in smaller quantities in a high throughput manner. Rosetta Genomics Ltd., together with U.S Genomics Inc., reported the development of a lung cancer detection technology that showed that miRNAs could be used as clinical biomarkers for a wide range of indications. 126 The successful use of these new biomarkers in clinical practice is expected in the near future.
Conclusion
Lung cancer is a spectrum of diseases with numerous alterations in expression patterns resulting from acquired genetic and epigenetic mechanisms. Despite many years of effort to identify biomarkers for human lung cancers, no discovery has generated the kind of interest garnered by the early studies exploring the potential of miRNAs. It is important from a pharmaceutical perspective that some miRNAs function as oncogenes while others act as tumor suppressors. For oncogenic miRNAs, miRNA inhibitors with “antagomirs” could be used to specifically inhibit each endogenous miRNA individually. For tumor suppressor miRNAs, a desirable therapeutic strategy is to enhance their function in cells. The feasibility of miRNA-based therapeutics is well supported by data from preclinical models and ongoing clinical trials. The potential of miRNAs in facilitating the diagnosis, prognosis, and survival outcomes for lung cancer should also be realized.
Footnotes
Author contributions
Both authors participated in writing this review and editing of the manuscript.
ACKNOWLEDGMENTS
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Korean Government (MSIP) (2008-0062282 and 2011-0014086).