
Abstract
The desire to develop and evaluate drugs as potential countermeasures for biological and chemical threats requires test systems that can also substitute for the clinical trials normally crucial for drug development. Current animal models have limited predictivity for drug efficacy in humans as the large majority of drugs fails in clinical trials. We have limited understanding of the function of the central nervous system and the complexity of the brain, especially during development and neuronal plasticity. Simple in vitro systems do not represent physiology and function of the brain. Moreover, the difficulty of studying interactions between human genetics and environmental factors leads to lack of knowledge about the events that induce neurological diseases. Microphysiological systems (MPS) promise to generate more complex in vitro human models that better simulate the organ’s biology and function. MPS combine different cell types in a specific three-dimensional (3D) configuration to simulate organs with a concrete function. The final aim of these MPS is to combine different “organoids” to generate a human-on-a-chip, an approach that would allow studies of complex physiological organ interactions. The recent discovery of induced pluripotent stem cells (iPSCs) gives a range of possibilities allowing cellular studies of individuals with different genetic backgrounds (e.g., human disease models). Application of iPSCs from different donors in MPS gives the opportunity to better understand mechanisms of the disease and can be a novel tool in drug development, toxicology, and medicine. In order to generate a brain-on-a-chip, we have established a 3D model from human iPSCs based on our experience with a 3D rat primary aggregating brain model. After four weeks of differentiation, human 3D aggregates stain positive for different neuronal markers and show higher gene expression of various neuronal differentiation markers compared to 2D cultures. Here we present the applications and challenges of this emerging technology.
Introduction
A key strategy of the US Department of Defense to prepare for chemical and biological threat agents, both in warfare and terrorism, is the development of medical countermeasures (MCM). The development and evaluation of such drugs require test systems that can also substitute for the clinical trials normally crucial for drug development, as there are normally no patients for clinical development. This makes a traditional product registration with Food and Drug Administration (FDA) impossible. FDA aimed to accommodate MCM developments by issuing the Animal Rule, i.e., the suggestion to use appropriate animal models instead1,2 in May 2002, which allows substituting for evidence of efficacy (not safety!) in humans with qualifying animal studies. However, animal models have limited predictivity for drug efficacy,3,4 as is well known from many disappointments in clinical trials. 5 Currently, 95–97% of drugs fail the clinical part of traditional drug development. There is no reason why MCM would be more successful; on the contrary, these are diseases to treat, which we hardly know, which are very acute and the threat agents are designed to harm. Traditional in vitro and in silico approaches are not satisfying this demand.6,7 The US Department of Defense sponsored a National Academy of Sciences report, Animal Models for Assessing Countermeasures to Bioterrorism Agents, published in December 2011. 8 The key findings of the report state that neither animal nor alternative methods are available for this purpose, but the committee discouraged the development of further animal models while proposing the exploitation of new alternative approaches. This consensus of the National Academies panel extends in some sense the earlier report, Toxicity Testing for the 21st Century: A Vision and a Strategy, (Tox-21 c) 9 to drug development. Especially, after taking stock of the state of the art of animal-based evaluations, a call is made for novel approaches based on today’s biotechnology. This contributed to the substantial investment into new tools now underway to develop human-on-a-chip approaches, which might bring about a second generation of alternative approaches. 10 The avenue pursued focuses primarily on the combination of different three-dimensional (3D) (stem) cell-based organ equivalents, also known as microphysiological systems (MPS), combined with microfluidics. Over the last two years, three funding opportunities for human-on-a-chip approaches in the US totaled almost $200 million. An alliance of NIH, FDA, and the Department of Defense (DoD) agency DARPA (Defense Advanced Research Agency) is tackling the problem of evaluating drugs for which there are no patients and, hopefully, never will be patients. In parallel, DTRA (Defense Threat Reduction Agency) has started a program also aiming for producing a number of 3D human organ equivalents based on stem cell technology and combining them with microfluidics-on-a-chip.
It is important to note that in yet another field the limitation of animal models was realized and at the same time there is need to regulate these new products. This might open doors for new alternative approaches. FDA’s new interest in predictive in vitro tools for MCM might be an avenue for a broader acceptance of novel tools for the evaluation of drugs in general.
As part of the larger efforts to create the human-on-a-chip, we report here on the efforts to develop a brain-on-a-chip and its biological and medical applications. Neurotoxicity is a major hazard and the main target for chemical and biological threat agents. Because of the complexity of the human brain, this organ represents an enormous challenge to model in animals or traditional cell culture systems. The complexity of brain development, with many additional windows of vulnerability, amplifies this if neurodevelopmental effects are addressed. Noteworthy, traditional animal regulatory tests for developmental neurotoxicity (DNT) use 1400 animals and cost $1.4 million per substance and still allow only a superficial test of a few central nervous system (CNS) functions. At the same time, advances of human relevant cellular models of the CNS will support the study and development of neurodegenerative diseases and their treatment, a major area of basic and pharmaceutical research. Neurodevelopmental disorders, such as learning disabilities, dyslexia, attention deficits hyperactivity disorder (ADHD), and autism, have increased over the years.11–13 These disorders affect not only the individual but also families and the society. For example, autism spectrum disorders (ASDs) that affect 1.1% of children in US 12 lead to morbidity and functional limitations that account for direct care and indirect costs of $126 billion per year. 14 However, we have limited knowledge about the genetic basis, environmental threats, and/or gene/environment interactions that increases the risk to develop these disorders.15,16 The same holds true for adult neurological disorders such as, e.g., Parkinson’s, Alzheimer, and Schizophrenia. Animal models cannot reflect human interindividual genetic differences that might contribute to these diseases and also limit the development of new personalized drugs that could enhance treatment. Consequently, human models reflecting different genetic backgrounds would likely improve the understanding of CNS toxicity and disease.
Modeling the complexity of the human brain
We have a limited understanding of the function of the CNS, especially during development and neuronal plasticity, which is difficult to reproduce with traditional in vitro methods. Consequently, the use of more complex models such as MPS is pivotal to reproduce the function and architecture of the brain. The CNS consists of numerous different cell types such as neurons, oligodendrocytes, astrocytes, and microglia, all with crucial roles for the organs biology and function. 17 There are several kinds of neurons which release different neurotransmitters that will induce diverse responses. The precise communication and regulation between these cells (neurons–neurons and neurons–glial cells) are of high importance for the function of the CNS. 18 Oligodendrocytes are key for the myelination that provides electrical insulation and make transmission along the axon more rapid. 19 This reduces ion leakage and decreases the capacitance of the cell membrane that is crucial for the neuronal impulse. In brain development, astrocytes guide the neurons to the right position during the migration process and play an important role in the assembly of synapses. While in the adult brain, astrocytes’ main function is to protect neurons and maintain the ionic and trophic balance of the extracellular milieu. Moreover, astrocytes have shown to regulate neuronal calcium levels by release of glutamate and might have important roles in regulating neuronal physiology and pathology. 20 Contrary to the protective role, microglia and astrocytes can also become activated in response to neuronal damage, which can make them secrete a variety of proinflammatory and neurotoxic factors, e.g., cytokines and free radicals, 21 which can enhance the neuronal damage. Hence, an in vitro model that aims to simulate the in vivo situation of the brain needs to have several of these cell types present 22 and mimic some of the functional process that are essential in the CNS, such as electrical firing, neuronal and astrocytic calcium waves, pruning, myelination, cell–cell interaction, migration, and neurogenesis.
Another important factor of the brain is the extracellular matrix (ECM). Over many years, it was thought that the brain has limited stromal space with an insignificant ECM in comparison to other organs. ECM commonly found in other organs such as fibronectin and collagen is not present in the brain, but studies have shown that a variety of other components are filling significant amounts of space between neurons and glial cells.23,24 Many of these are proteoglycans secreted by neurons or glia cells. 25 Apart from being a homeostasis regulator, the ECM has the capacity to bind growth factors that can enhance their interaction with the cells. 26 Furthermore, in the nervous system, the ECM has been related to several specific processes such as mitogenesis, angiogenesis, neurite and glial outgrowths, and synapse morphology. 27
The use of 2D co-cultures (e.g., astrocytes and neurons) has clearly enhanced the mimicking of the in vivo brain compared to pure neuronal cultures, especially in a functional perspective. For example, the NT2 cell line can be differentiated into different cell types such as neurons, astrocytes, and oligodendrocytes and has shown neuron–glia interaction with present glutamatergic and GABAergic functional synapses. 28 Other processes such as myelination have also been reproduced in vitro. 29 These 2D cultures present some advantages due to the fact that they are easy to obtain, easy to handle, and have optimized readout systems. However, even with a mixed cellular system, the complex cell–cell interactions and the neurite networks within the brain remain a challenge to model with traditional monolayer cultures. In vivo, astrocytes often interact with numerous neurons, which is not achievable in 2D systems as they restrict the cellular shape. Three-dimensional models are therefore more likely to reproduce the complexity of directional growth and cell–cell connections and resembles more closely the in vivo situation. 30 Three-dimensional cultures have shown improved cell survival and enhanced neuronal differentiation compared to traditional 2D cultures.31–33 This could be as the 3D structure leads to an improved cell–cell and cell–matrix interaction that provides better cell signaling and gap junction connections.
In addition, cell–cell signaling can promote proliferation of glial and neural stem cells (NSC) as well as can enhance neurogenesis and synapse formation, a crucial process for brain functionality. For example, the development of 3D cell cultures from a human umbilical cord blood-derived neural stem cells line (HUCB-NSC) showed functional properties observed by spontaneous electrical activity, a physiology that could not be obtained from the same cell line cultured in 2D. 34 Several neuronal cultures have also shown spontaneous electrical activity in 2D.35–37 However, interestingly cells that do not display electrical activity in 2D can become active when cultured in 3D, indicating an enhanced cell differentiation and maturation. Functional synapses have also been observed in other 3D neuronal cultures at similar or earlier time points compared to 2D models.35,38,39 Furthermore, neural stem/progenitor cells in 3D cultures showed an increased differentiation into neurons with 70% versus 14% in 2D cultures. 32 Not only neurons obtain enhanced phenotype in 3D, a study using a microglial cell line showed that microglial cells placed in an artificial 3D matrix developed bi or multipolar phenotypes, whereas cells cultured under traditional 2D conditions only showed one amoeboid phenotype. 40
The third dimension in cell cultures has shown to be especially important for improvement of drug discovery and toxicity testing, as these models are much closer to the in vivo situation compared to monolayer cultures.41–44 The EU-funded integrated project “ACuteTox” aimed to use in vitro systems to predict oral acute systemic toxicity using approximately 50 endpoints and different cell models such as neuronal cell lines, primary neuronal cell cultures, brain slices, and aggregated brain cell cultures. 45 The use of multiendpoint assays in 3D re-aggregating brain cultures seemed to be the most complete test, with optimum prediction for neurotoxicants. The same rat primary aggregating brain cell model was identified by the DNT consensus process with so far three conferences and two workshops among the most representative models for DNT studies.46–48 The model includes all relevant cell types (several different types of neurons, astrocytes, oligodendrocytes, and microglia) in a 3D structure thereby supporting the cell–cell interactions that are crucial for mechanistic studies. Rat primary aggregating brain cultures are prepared from embryonic day 16 fetal rat brains as previously described. 38 Briefly, the dissected tissue is mechanically dissociated and cells re-suspended in a modified serum-free media. Cells are re-aggregated and maintained under constant gyratory shaking at 37℃ in an atmosphere of 10% CO2. After approximately three days in vitro, all cells are aggregated into 3D aggregates, which are robust in terms of size and morphology. Our laboratory and others have characterized this model with regard to immunohistochemistry, electrophysiology, myelination, pharmacological behavior and gene markers of neurodevelopment, and DNT effects.38,44,49–53 We have used the model in a project granted by the US Food and Drug Administration (#U01FD004230), in order to map pathways of DNT. 54
Three-dimensional brain cell cultures can be obtained from different sources such as cell lines,40,55 primary cells,32,56 and stem cells.34,57,58 Depending of the cells used, the 3D culturing techniques can vary, e.g., stem cells are commonly forming 3D structures spontaneously, while cell lines and primary cells often need support by, e.g., ECM, hanging drop, or rotation-mediated culture techniques.
As previously discussed, the brain consists of numerous different interacting cell types making it a major challenge to model in vitro. Consequently, more sophisticated 3D cell systems are necessary to provide a better image of the in vivo situation. In addition, the brain is closely connected to the blood–brain barrier (BBB) and the brain vasculature that are especially important to model in development of drugs for the CNS. The following section will therefore describe the efforts to develop cerebrovascular systems in vitro.
MPS to mimic the cerebrovascular system
The BBB is a unique biological barrier, formed by capillary endothelial cells between blood and brain interstitial fluid (ISF), the choroid plexus epithelium between blood and ventricular CSF, and the arachnoid epithelium between blood and subarachnoid CSF, 59 that regulate the interchange between blood and brain extracellular fluid. Individual neurons are rarely more than 8–20 µm from a brain capillary. 60 Complex tight junctions between adjacent endothelial cells force molecules to go through transcellular routes by passive diffusion (water, some gases, lipid soluble molecules) or by selective transportation (glucose, amino acids). 61 The BBB and brain vasculature have shown to be crucial for the development and function of the CNS. Moreover, perturbation of the cerebrovascular system can contribute to disease, e.g., hemodynamic alterations and pro-inflammatory responses have shown to adversely affect vascular integrity that can lead to diverse neurological disorders such as Alzheimer’s disease, multiple sclerosis, HIV-associated encephalopathy, and epilepsy. 62 A functional in vitro model of the brain circulation system could therefore contribute to better understand the response mechanisms of the cerebrovascular system. Moreover, such a model could be useful in the development of drugs targeting the CNS that need to penetrate the BBB. Processes such as interendothelial tight junctions, carrier-mediated transport systems, astrocyte interaction, vascular integrity, and permeability are some relevant elements in the BBB system.63–65 Various groups have combined different cell types in co-cultures to simulate the BBB in vitro, however, mostly from primary animal sources. 66 Brain microvessel endothelial cells have been isolated from the cortex (grey matter) using different methods (mechanical dispersion, enzymatic, differential seeding, selective outgrowth, selective isolation, etc.), with the possibilities to reach 95% purity. 66 Some cryopreserved brain capillary endothelial cell lines are also commercially available.66,67 The endothelial cells are often co-cultured with pericytes, astrocytes, microglia, and/or neurons to form the BBB model.68,69 For example, co-cultures of rat brain endothelial cells (RBE4) and rat E-18 cortical cells formed an endothelial barrier with tight junctions and neuroinflammation responses. 70 Non-cerebral endothelial cell lines have been used as a BBB model as they re-induce their BBB properties when co-cultured with astrocytes. In many studies, BBB in vitro systems use incubation chambers (transwell plates) where endothelial cells are plated on the porous membrane of the insert and the astrocytes at the bottom surface of the well.71,72 However, other techniques such as a co-culture in gel matrix have also been used lately. 73
As mentioned before, most of the in vitro models of the BBB utilize murine or porcine brain endothelium and astrocytes. Very few BBB models have been developed from human cells. However, in recent years, a humanized in vitro BBB model was developed using multiculture human cell systems. 74 Hatherell et al. reconstructed the human-derived BBB components in vitro in a 3D configuration using human astrocytes, human brain vascular pericytes, and human cerebral microvascular endothelial cells. More recently, induced pluripotent stem cells (iPSCs) have been successfully differentiated into BBB endothelial cells, which open new opportunities for MPS. 75 However, development of 3D structures of the vasculature is still necessary to mimic the cell organization in the physiological way.76,77
The simplest 3D models can be established using multilayer cultures. As an example, endothelial cells derived from human pluripotent stem cells (hPSCs) were co-cultured with primary rat astrocytes in membrane pore transwell plates resulting in a model with many BBB attributes similar to the molecular permeability in vivo (e.g., well-organized tight junctions, appropriate expression of nutrient transporters and polarized efflux transporter activity, barrier properties). 75
Even though structural similarities between in vitro and in vivo BBB have been obtained, the challenge to incorporate crucial aspects such as flow or physiological shear stress still remains. These features play a critical role in modulating the differentiation of vascular endothelial cells into a BBB phenotype.65,78 A human model of the capillary-venue system was recently developed using co-cultures of adult human brain microvascular endothelial cells, human adult astrocytes, and human brain vascular smooth muscle cells using a hollow fiber technology. 76 A pump generates a flow through two segments (capillaries and venules) creating a hemodynamic microenvironment. The perfusate of the capillary module achieves levels that simulate in vivo shear stress, pressure, and flow rate. Microfluidics is a multidisciplinary engineering field that deals with behavior, precise control, and manipulation of fluids on a submillimeter scale. However, in the latest years, new technologies have been established to develop microfluidic BBB systems, which obtain better control of the shear stress and flow of the model.79–82
Combining human stem cell technologies, bioengineering and MPS, with incorporation of microfluidic systems, will likely generate more representative models of the BBB. One of the most promising sources to obtain human MPS is iPSCs. The use of these cells gives a range of possibilities allowing cellular studies of individuals with different genetic backgrounds (e.g., human disease models) (Figure 1). The following section will focus on iPSCs in the context of the development of MPS.
Schematic figure of the generation and application of a brain-on-a-chip. Somatic cells can be obtained from donors with different genetic backgrounds. After prior reprograming to human-induced pluripotent stem cell (hiPSC), they can be differentiated into neuronal and glial cells and be combined to form a microphysiological system (MPS). MPS can be incorporated into chips in order to measure different parameters and can be a useful tool for toxicity testing of chemicals and drugs. (A color version of this figure is available in the online journal.)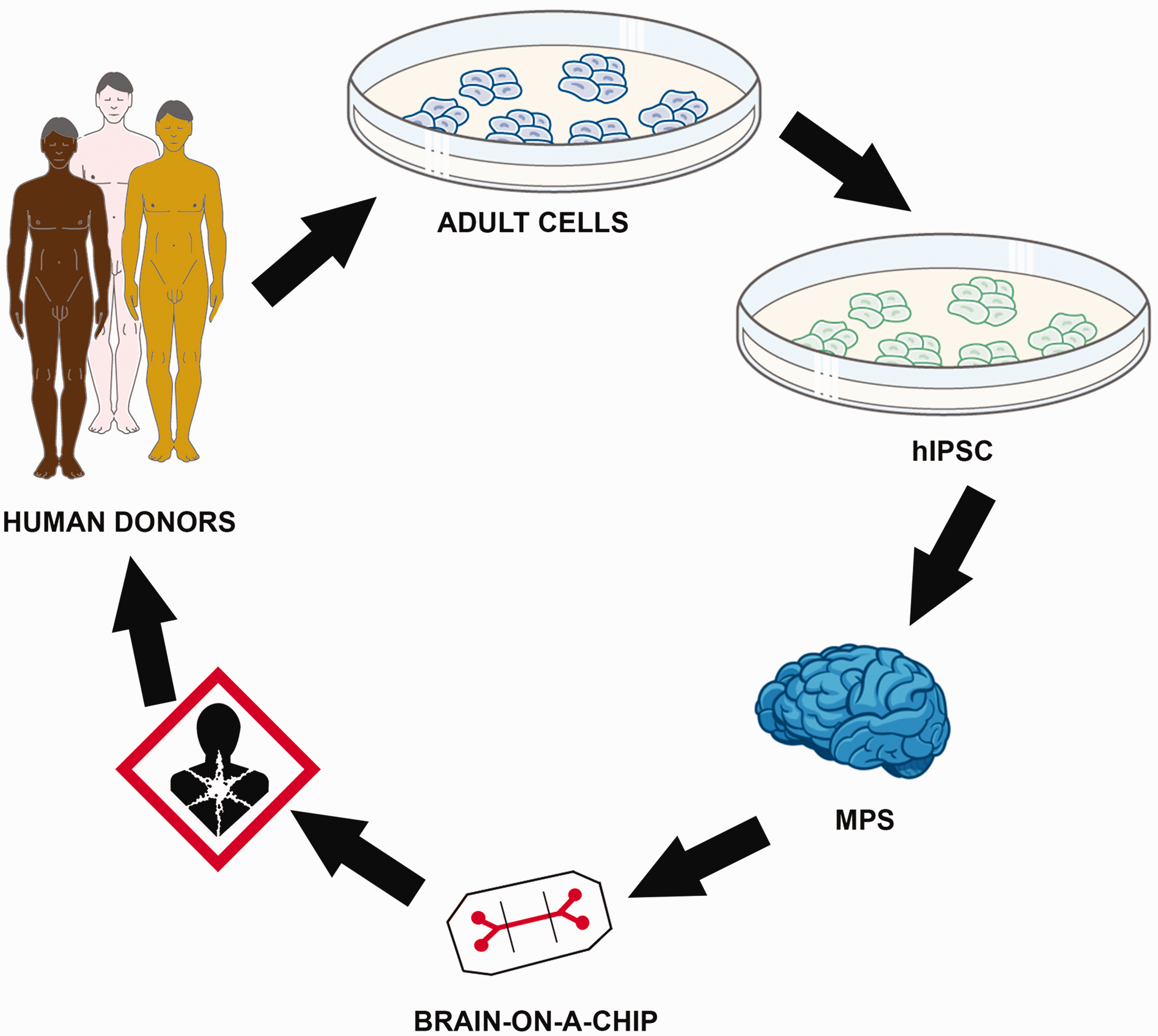
The potential of iPSCs for MPS
Human embryonic stem cells (ESCs) have changed the scientific view of regenerative medicine and disease therapies, and it has become a powerful tool to study human biology. However, many ethical issues have appeared related with the precedence and isolation of these cells from human embryos. In 2006, Yamanaka’s group discovered the possibility to obtain pluripotent cells by reprograming mouse fibroblast cells by introducing exogenous factors. Four transcription factors related with ESCs potency maintenance (Oct3/4, Sox2, Klf4, and c-Myc) were introduced by retroviral infection to enable generation of pluripotent cells (Figure 1). 83 Later, these iPSCs, were also generated from human adult cells 84 and showed the capability to generate all three germ layers. 85 The expression of these transcription factors led to the global reversion of the somatic epigenome into pluripotent ESCs state. 86 Since then, different combinations of transcription factors and transfection techniques of the cells have been developed.85,87
The capability to obtain iPSCs from adults with different genetic backgrounds opens up a new realm of possibilities for stem cell research and medicine. Three major areas where the application of iPSCs can make a huge difference are in regenerative medicine, study of diseases, and in toxicity testing of chemicals and drugs. 88 Nevertheless, the reprograming process introduces some challenges. Small dissimilarities between ESCs and iPSCs have been observed, 89 such as differences in methylated regions. 90 These differences are likely due to the cell epigenetic memory conserved in the reprogramed cells. 90 The impact of these changes on the cell development and function is still unknown.
iPSCs can be differentiated into neuronal and glial specific phenotypes. Several protocols have been established with various degrees of efficiency for dopaminergic neurons,91–93 astrocytes, 94 and oligodedrocytes.95,96 A special challenge to achieve organo-typic culture is the representation of different cell types in a co-culture. Traditionally, these had to be separately isolated or differentiated and mixed. However, some protocols allow direct-derived mixed cultures from stem cells, similarly to the normal embryonic development of an organ system.
ESCs and iPSCs are mainly cultured as 2D models, however, as previously discussed 3D models are essential to mimic the complexity of the human brain. Recently, different groups have developed 3D polarized neuroepithelial structures similar to the structure formed in the embryonic cortex from both ESCs97–99 and iPSCs.99,100 These 3D organoid culture systems have shown to mimic various discrete brain regions with expression of specific markers of forebrain, midbrain, and hindbrain as well as other brain structures such as choroid plexus and even immature retina.99,101 The 3D iPSCs model developed by Lancaster et al. 99 showed the typical organization of the ventricular zone and the same type of migration between brain regions as observed in vivo. Even though the organoid model does not fully recapture the organization of the brain, 102 such a model will likely be particularly valuable for studies of neurodevelopmental and disease mechanisms.
Our group (funded by the NIH #1U18TR000547) has in the last year established a humanized model of the 3D rat primary aggregating brain model (previous described) using human iPSCs.
54
iPSCs are derived and characterized from healthy and diseased donors and are differentiated in a step-wise neural differentiation protocol through embryoid body formation as previously described.
103
Embryoid bodies are then cultured in suspension in neuronal precursor (hNPC) medium (DMEM/F12, Neurobasal, heparin, N2). After two weeks, embryoid bodies are dissociated using Accutase® (incubation 45 min at 37℃) and seeded as single cell cultures on poly-L-ornithine/laminin coated six-well plates and passaged mechanically as adherent cultures (Figure 2(a)). For neuronal differentiation of NPCs, early passage neuronal-precursor cells are detached and dissociated into single cell suspension using Accutase®, distributed onto non-treated six-well plates and cultured under constant gyratory shaking at 37℃ in an atmosphere of 5% CO2 (Figure 2(b) and (c)). After four days in hNPC medium, the medium is changed to differentiation medium (neurobasal medium supplemented with 2 mM L-glutamine, B27, GDNF, and BDNF) and aggregates are kept under differentiation for up to eight weeks.
54
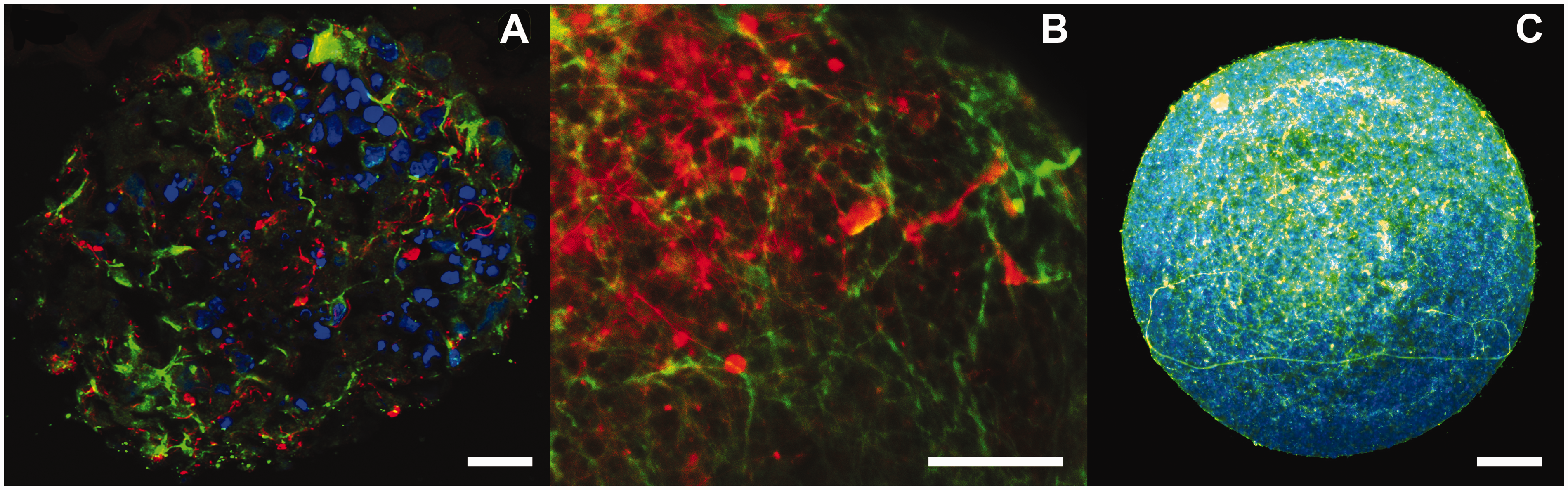
After four weeks of differentiation, the human 3D aggregates stain positive for different neuronal markers such as Map-2 (dendrites), NF200 (axons) (Figure 3(a)), and the astrocytic marker GFAP (Figure 3(b)). NF200 shows further increased expression after eight weeks of differentiation (Figure 3(c)). Interestingly, different neuronal markers showed higher mRNA expression in 3D cultures compared to 2D cultures.
54
Initially, several different approaches were developed in parallel before the protocol described above was selected as it showed robust and reproducible neural differentiation. This project aims to use the human 3D brain model to study gene/environmental interactions; however, the model also has the capacity to study brain development, neuronal disorders, and drug therapies, especially as iPSCs can be derived from individuals with different genetic backgrounds, e.g., disease models. The cell model is exposed to reference chemicals with well-described neurotoxic/DNT effects and the predictivity is assessed by gene expression, immunohistchemistry, and calcium signaling (functional endpoint) measurements. Effects are compared to human data from the literature and previously obtained data from the rat 3D model. In addition, we determine whether iPSCs derived from donors with neurological disorders (e.g., Down’s syndrome) retain dysfunctional signaling pathways expected by the genetic modification. Finally, we evaluate if the genetic modification increases sensitivity to chemical exposure as, e.g., Down’s syndrome patients have shown increased sensitivity to oxidative stress. This approach gives broader information on gene/environmental interactions and can also provide information on the actual disease. iPSCs have already been successfully derived from patients with different neuronal disorders (Table 1) and have shown to mimic the relevant cellular and/or molecular phenotype of the respective disorder. For example, iPSCs derived from Huntington’s disease (HD) were able to reproduce the disease phenotype.
115
Human 3D brain models using iPSCs derived from individuals with neuronal disorders will be a useful tool for research of the brains physiology and pathology.
Generation of a human 3D neuronal cell model. Phase contrast picture of human NPCs (a) that are cultured as single cell suspension during constant gyratory movement (b) to form 3D neural aggregates with (c) a diameter of approximately 300 µm. (A color version of this figure is available in the online journal.) Characterization of a 3D neural cell model by immunohistochemistry. After four weeks of differentiation, human 3D aggregates stain positive for the neuronal markers Neurofilament 200 (NF200) (green) and Microtubule associated protein 2 (Map-2) (red) (a) indicating the presence of neurons. After eight weeks of differentiation aggregates show a higher density of Map-2 (green) positive cells and in addition stain positive for the astrocytic marker glial fibrillary acidic protein (GFAP) (red) (b). Moreover, the eight weeks aggregates show increased staining of NF200 (green) (c). Nucleus is stained with Hoechst (blue) in panels a and c. Bars represent 50 µm. (A color version of this figure is available in the online journal.) iPSCs derived from patients with neuronal disorders

In conclusion, the possibility to generate MPS from donors with different genetic background provides a great opportunity for drug development, (developmental) neurotoxicity, and a useful tool to understand brain complexity and mechanisms of brain disease.
Future applications and assays of a human brain-on-a-chip
There is a critical lack of knowledge when it comes to toxicity of drugs and other xenobiotic chemicals on the adult and developing brain. The discovery of iPSCs has led to significant possibilities to study human toxicity and disease also from a genetic point of view. The use of iPSCs cells within MPS has the potential to build human “mini-brains” that better reveal the complex physiology and function of the brain, e.g., migration, neuronal differentiation, network elaboration cell–cell interactions (neuron–neuron and neuron–glial cells), myelination, and synapse formation.35,116 The Boyden chamber is often used for in vitro migration studies but is one of the assays that are less suitable for complex 3D models. 117 Migration in 3D neural models can instead be assessed by measuring radial migration away from adherent neurospheres;118,119 however, the migration within the MPS still resembles a challenge. The inclusion of cells transfected with fluorescent proteins combined with confocal microscopy offers new opportunities here. Neuronal differentiation is one of the most complex processes of the CNS and includes several events such as outgrowths of axons and dendrites, expression of neurotransmitters and receptors, synapse formation, and maturation of electrical excitability. Different morphological and biochemical endpoints have been developed to measure neuronal network elaboration, e.g., the neurite outgrowth assay or detection of neuronal cytoskeleton proteins or mRNAs.52,120 However, to achieve mature neuronal function, the neurons must form cell–cell connections through synapses. Most common assessment of synapse formation is immunocytochemical analysis for pre and postsynaptic markers.121,122 However, to fully assess the functionality, calcium and/or electrophysiological measurements are necessary. Several techniques for calcium imaging have been developed such as fluorescent assays and more complex reporter cell lines.99,123,124 One of the most recent and promising tools for electrical activity measurements is micro electrode array (MEA).35,36,38,125,126 This technique measures extracellular whole neuronal networks and provides more relevant physiological information than other conventional methods for electrophysiology assessment, e.g., patch clamp. The most commonly used in vitro systems for electrical recordings are 2D hippocampus slices and primary dissociated cultures, normally from spinal cord or cortex.35,36,127 However, many different in vitro systems can be used with MEA, including human stem cells and 3D brain cultures.37,128 MEAs have been used in several studies of pharmacological and toxicological responses and can be used to assess the functionality of a neuronal model.35,38,125,128 Finally, glial maturation and function, such as glutamate uptake and transformation into glutamine by astrocytes and myelination of oligodendrocytes are often assessed by cell specific markers53,129 or biochemical assays.130,131 Several of these assays are mainly developed and applied for traditional 2D cell models why many of them need to be optimized for the more complex 3D models and can represent a challenge for the more complex MPS.
Challenges associated with MPS and the human-on-a-chip concept
MPS have shown a great potential for the study of diseases and to generate new in vitro methods for toxicity testing and drug discovery. However, there are also several challenges associated with MPS and the human-on-a-chip approach. 10 The approach is extremely promising; still the question emerges whether the enormous efforts to create such complex systems are necessary to predict human effects. Some of the challenges include differentiation into various cell types to achieve true organotypic cultures. This is especially challenging for the brain as it consist of so many different cell types (different kind of neurons, astrocytes, oligodendrocytes, microglia, pericytes, and endothelial cells) that need to be present in precise ratio and have substantial cell–cell interactions to reach organ functionality. In addition, the various cell types of the CNS have their specific differentiation and maturation patterns that often require long time culturing. Complex 3D brain models are also often more difficult to evaluate than traditional monolayer cultures. Two-dimensional models are normally more easily accessible, and several assays and endpoints have been developed for 2D cultures and need to be optimized for 3D conditions. An example is the challenge to study 3D brain models at the single-cell level using, e.g., morphological studies, immunohistochemistry, and reporter gene techniques. This is especially important in neuronal cultures, as morphological observations such as neurite outgrowth120,132 and synaptogensis 122 have shown to be some of the most promising assays for neurotoxicity assessment. This also challenge the usual neuronal functionality assays, e.g., calcium imaging and patch clamp. MEA is still a tool that can be used for 3D tissue like cultures, though mainly the activity of the surface will be recorded. 38 Furthermore, new/adapted techniques for 3D cultures are being explored.118,133,134 Even though we are far from building the perfect brain, MPS complex enough to mimic some brain functionality will be far superior to traditional monolayer cultures.
Once the MPS of the different organs have been successfully developed the next challenge will be to combine them for the human-on-a-chip approach. A compromise cell culture medium to maintain all the different organotypic cultures needs to be established and the organs and perfusion liquid compartments require balanced size to allow close physiological kinetics. Too large MPS will have limited access to essential nutrients and oxygen and require additional perfusion systems to avoid necrotic and/or apoptotic tissue. It is not clear, how well the brain organoids will tolerate co-cultures with other organoids, especially under re-circulating perfusion conditions. Re-circulation will be necessary to allow full organoid interactions and also accumulation of released products (also to be used as biomarkers), but organoids might compete for nutrients and intoxicate each other with waste products. This urges even more the combination with a functional BBB.
This is not just a biological and bioengineering challenge but also a standardization and reproducibility challenge. In general, the more complex models tend to have lower reproducibility. The development of 3D culturing techniques, such as use of ECM, 55 micro-contact printing, 135 and micro well arrays, 58 has enabled systems capable of control and modulate shape and size to increase the homogeneity of the MPS. However, the use of ECM should be carefully evaluated as the wrong material can decrease cell viability and mobility of the cells. 136 This is particularly important for neuronal and glial cells as they secrete their own specific components (see Modeling the complexity of the human brain). Still, neural cells are often cultured in ECM likely not optimal for neural cultures, e.g., Matrigel or Geltrex while neural 3D models cultured without ECM could be an advantage. Furthermore, over-engineering could induce further artificial effects.
These are important challenges when considering applications of the MPS, especially for scientific or regulatory implementation. In the case of, e.g., toxicity testing, a key lesson from the validation of alternative methods 137 is the pivotal role of this challenge, and usually this requires simplicity, as too many artificially chosen and difficult to control conditions influence our experiments. 6 To find a balance between the complexity of the system, the relevance and the reproducibility are crucial in order to generate useful MPS and this will represent additional challenges as to standardization of design and generation of cultures and devices. This holds especially true for the brain with its inherent complexity. Further, in order to apply this approach in regulatory procedures, some kind of formal validation as developed by European Centre for the Validation of Alternative Methods (ECVAM), adapted and expanded by Interagency Coordinating Committee of the Validation of Alternative Methods (ICCVAM) and other validation bodies, and, finally, internationally harmonized by Organisation for Economic Co-operation and Development (OECD) is needed. 138 Validation is the independent assessment of the scientific basis, the reproducibility, and the predictive capacity of a test. It was re-defined in 2004 in the Modular Approach 139 but needs to be seen as a continuous adaptation of the process to practical needs and a case-by-case assessment of what is feasible.140,141 A key problem for the novel technologies is the absence of a point of reference, i.e., a “traditional test” or “gold standard.” To some extent, data on organ toxicities in repeated-dose systemic toxicity studies, reproductive and developmental toxicity studies as well as dedicated neurotoxicity and DNT studies are available for the brain. In the absence of reference data, the scientific validation needs to be stressed. 142 The framework of evidence-based medicine is increasingly being translated to toxicology, 143 and it recently led to the creation of the evidence-based toxicology collaboration (EBTC).144,145 Most recently this has led to a proposal for mechanistic validation, 146 which might be a major support to the validation of MPS. The basic idea is to analyze to which extent a given test reflects current scientific understanding in a formalized way (a systematic review); in case of toxicity, this would be the established modes of action, pathways of toxicity and adverse outcome pathways. This contrasts to traditional validation, which largely considers the test system a black box and correlates results with those of (black box) animal models.
The investment into superior in vitro models will promote toxicology, pharmacology, and medicine for the 21st century, even if it does not result in a routine predictive tool. The added benefit in pursing this challenge is that the in vitro, human-based, mechanistically oriented organ-on-a-chip or combined to human-on-a-chip, and other test systems developed for that purpose will also serve to meet the needs of the other disciplines that currently rely on animal tests to predict human responses.
Footnotes
Author contributions
All authors contributed equally.
ACKNOWLEDGEMENTS
This project is supported by the NCATS grant “A 3D Model of Human Brain Development for Studying Gene/Environment Interactions” (1U18TR000547). Additional work by the group referred to in the article is supported by the US Food and Drug Administration grant “DNTox-21 c Identification of Pathways of Developmental Neurotoxicity for High Throughput Testing by Metabolomics” (U01FD004230).