
Abstract
Vascular endothelial growth factor (VEGF) is elevated in chronic rhinosinusitis with nasal polyps. Steroids have anti-inflammatory properties and are ideal candidates for treating chronic inflammatory airways. The aims of this study were to identify the inhibitory effects and mechanisms of steroids on lipopolysaccharide (LPS)-induced VEGF expression in nasal polyps. Nasal polyp-derived fibroblasts (NPDFs) were stimulated with LPS alone or with both LPS and steroids were used to determine the expression levels of toll-like receptor (TLR)-4, myeloid differentiation primary response gene 88 (MyD88), and VEGF by using reverse transcription-polymerase chain reaction (RT-PCR). VEGF protein level was analyzed by immunocytochemical staining and enzyme-linked immunosorbent assay (ELISA). Small interfering RNA (siRNA) for TLR4 was transfected to down-regulate TLR4 expression. Activation of Akt and nuclear factor κB (NF-κB) pathway on VEGF expression was determined by Western blot analysis, immunocytochemical staining, and ELISA. Nasal polyp organ cultures were stimulated with LPS alone or in conjunction with steroids or LPS-RS (TLR4 inhibitor) and accessed the expression of VEGF. Steroids decreased the expressions of TLR4, MyD88, and VEGF mRNA and VEGF protein in LPS-stimulated NPDFs. Steroids inhibited LPS-induced VEGF expression levels in dose-dependent manner. The suppression of TLR4 transcription by siRNA treatment reduced LPS-induced expression of both TLR4 and VEGF in NPDFs. Furthermore, steroids inhibited the production of VEGF by blocking Akt and NF-κB activation and preventing with NF-κB translocation. Also, steroid and TLR4 inhibitor decreased VEGF expression in nasal polyp organ cultures. These results indicate that steroids inhibit LPS-induced VEGF expression through the TLR4/Akt/NF-κB signaling pathway in chronic rhinosinusitis with nasal polyp.
Keywords
Introduction
Chronic rhinosinusitis, a prolonged inflammation of the sinonasal mucosa, leads to nasal obstruction, rhinorrhea, hyposmia, and facial pain. 1 Chronic rhinosinusitis without nasal polyposis and chronic rhinosinusitis with polyposis may present distinct clinical and pathological subtypes of chronic rhinosinusitis. 1 Nasal polyposis results in benign lesions in the sino-nasal mucosa. 1 Nasal polyps are characterized by the infiltration of inflammatory cells such as eosinophils or neutrophils, and by the proliferation and thickening of mucosal epithelium with focal squamous metaplasia, glandular hyperplasia, basement membrane thickening, and edematous stroma. 2 Although the common treatment for nasal polyps is endoscopic functional sinus surgery, steroids are often used to treat nasal polyps and previous studies show that the intranasal application of steroids can inhibit the growth of nasal polyps. 3 However, the therapeutic mechanisms of steroids on nasal polyposis are not fully understood.
Endothelial cells or epithelial cells are important for nasal polyp development. However, new concepts concerning the functions of fibroblasts have been defined recently. 4 The reasons are fibroblasts express many receptors for cytokines, growth factors, and hormones and play a key role in the structural modification of the sinonasal mucosa. Fibroblasts are major structural components of tissues, where they confer mechanical strength by providing a supporting framework for the extracellular matrix (ECM) such as collagen and fibronectin that is important to develop the nasal polyp formation. 5 In addition, recent studies have shown that fibroblasts are not just a structural modifier, but also produce a variety of cytokines such as vascular endothelial growth factor (VEGF). 6
VEGF is one of the main proteins involved in angiogenic sprouting or vasculogenesis. 7 VEGF production by fibroblasts may play an important role in the vascular changes, one of the features of airway remodeling. 8 VEGF plays a significant role in the regulation of capillary and basilar membrane permeability, which can influence both edema development and growth of nasal polyps. 9 Also, VEGF increase microvascular permeability and allows the extravasation of plasma proteins that participate in the accumulation of the ECM, thereby accelerating nasal polyp growth. 10 In our previous study, it was reported that the increased level of VEGF in cultured nasal fibroblasts involved with the pathogenesis of nasal polyps. 11 In the present study, we found that steroids inhibited the expression of VEGF via TLR4/Akt/NF-κB pathway in nasal polyp-derived fibroblasts (NPDFs).
Toll-like receptors (TLRs) are type-I transmembrane receptors that are composed of an extracellular domain with leucine-rich repeats and an intracellular signaling domain. TLRs play a crucial role in initiating inflammatory responses against invading microorganisms by the recognition of pathogen-associated molecular patterns (PAMPs) that are related with foreign pathogens from bacteria, viruses, and fungi. 12 After binding to PAMPs (such as lipopolysaccharide (LPS), lipopeptides, dsRNA, and bacterial DNA), TLRs and Toll/interleukin-1 receptor domain form a dimer and the receptor undergoes a conformational change that is required to recruit downstream signaling molecules such as myeloid differentiation primary response gene 88 (MyD88). TLRs directly recognize a series of PAMPs and induce the release of inflammatory cytokines, and link the innate and adaptive immune responses. Among the 10 TLRs, TLR4 has been extensively studied and is known to bind to LPS, which is found in the outer membrane of Gram-negative bacteria. 12 LPS is a PAMP that induces pathogenicity and binds the CD14/TLR4/MD2 receptor complex in many cell types. 13 Previous studies have reported that LPS induced VEGF up-regulation in murine macrophages, 14 rat lung pericytes, 15 and odontoblasts. 16
Steroids are involved in cholesterol metabolism. These hormones are effective immunosuppressant and anti-inflammatory agents, and are naturally produced in the adrenal glands upon stimulation by the hypothalamic–pituitary–adrenal axis. These steroid hormones are the main component of therapeutic agents used for the treatment of various diseases such as asthma, allergic rhinitis, and nasal polyposis. 17 Recent study has also suggested that VEGF expression in asthma is down-regulated by steroids. 18 Steroids may be used for treating chronic airway inflammatory diseases that progress through continued neovascularization.
The objectives of the present study were to identify the inhibitory effects of steroids on LPS-induced VEGF expression and to determine the underlying mechanism in nasal polyp.
Materials and methods
Reagents
LPS from Pseudomonas aeruginosa, dexamethasone (DEX, 2.5 µmol/L), fluticasone propionate (FP, 2.5 µmol/L), inhibitors of Akt (LY294002, 10 µmol/L), and nuclear factor κB (NF-κB) (BAY-11, 2.5 µmol/L) were all purchased from Sigma (St. Louis, MO). Antibodies against p50, phospho-Akt, and β-actin were obtained from Santa Cruz Biotechnology (Santa Cruz, CA).
Nasal tissues and fibroblast culture
Eight patients with nasal polyps were recruited from the Department of Otorhinolaryngology, Korea University Medical Center in Korea. They are all non-smokers and have no history of nasal allergy, asthma, or aspirin hypersensitivity, and they had not been treated with oral or topical anti-allergic agents during the previous eight weeks. Written informed consent was obtained from each patient, and the study was approved by the Korea University Medical Center Institutional Review Board (KUGH12041). The patients did not have allergies, asthma, or aspirin sensitivity. NPDFs were isolated from surgical tissues by enzymatic digestion with collagenase (500 U/mL, Sigma), hyaluronidase (30 U/mL, Sigma), and DNase (10 U/mL, Sigma). Cells were cultured in Dulbecco’s Modified Eagle Medium containing 10% (v/v) heat-inactivated fetal bovine serum (Invitrogen, Carlsbad, CA), 1% (v/v), 10,000 U/mL penicillin, and 10,000 µg/mL streptomycin (Invitrogen). NPDF purity was confirmed by microscopically assessing the cell morphology and characterizing the cell markers by flow cytometry. 19
Reverse transcription-polymerase chain reaction (RT-PCR)
For determining VEGF mRNA expression, NPDFs were stimulated with LPS (1 µg/mL) for 12 h. For determining TLR4 mRNA expression, NPDFs were stimulated with LPS for 24 h. Total RNA was isolated by using Trizol reagent (Invitrogen). MMLV transcriptase (Invitrogen) was used to reverse transcribe 2 µg RNA. PCR was performed using the primer pairs targeting specific genes as follows: VEGF (sense sequence, 5′-GCC TTG CCT TGC TGC TCT ACC-3′; anti-sense sequence, 5′-CAC ACT CCA GGC CCT CGT CAT TG-3′; 251 bp), TLR4 (sense sequence, 5′-TGA GCA GTC GTG CTG GTA TC-3′; anti-sense sequence, 5′-CAG GGC TTT TCT GAG TCG TC-3′; 167 bp), MyD88 (sense sequence, 5′-CCA GTT TGT GCA GGA GAT GA-3′; anti-sense sequence, 5′-AGG ATG CTG GGG AAC TCT TT-3′; 287 bp), GAPDH (sense sequence, 5′-GTG GAT ATT GTT GCC ATC AAT GAC C-3′; anti-sense sequence, 5′-GCC CCA GCC TTC TTC ATG GTG GT-3′; 271 bp). Amplification reactions were performed as follows: an initial denaturation step at 94℃ for 5 min, followed by 30 cycles at 94℃ for 45 s, 55–65℃ for 45 s, 72℃ for 45 s, and a final extension step at 74℃ for 5 min. All reactions were performed in a 20 µL volume. Products were electrophoresed on a 1.5% agarose gel and visualized by staining with ethidium bromide. The gels were visualized by using Molecular Imager ChemiDoc XRS+ (Bio-Rad, Hercules, CA).
Transfection with small interference RNA (siRNA) of TLR4
NPDFs were washed twice with phosphate-buffered saline and then the cells were dissociated. The cells were pelleted by centrifugation at 13,000 r/min for 3 min; thereafter, the cells were suspended in 1 mL phosphate-buffered saline and dispersed using a pipette. The cells were pelleted at 1000 r/min for 1 min. The supernatant was discarded, and the cells were suspended in Neon Resuspension buffer (Invitrogen) at a concentration of 6 × 105 cells/mL. Universal negative control siRNA (siControl; Santa Cruz) and small interference oligonucleotide RNA directed against TLR4 (siTLR4; Santa Cruz) were used as controls. Neon electrolytic buffer (Invitrogen) was added into the Neon transfection tubes, and the tubes were then placed in the Neon transfection system device (Invitrogen) that was set to 1400 V and 30 pulses. Gold tips were used to aspirate 100 µL RNA cell mixture and placed in the device station. After electroporation, an appropriate amount of complete medium was immediately added to each cell aliquot, and the cells were re-plated on to culture dishes.
VEGF quantitation by enzyme-linked immunosorbent assay (ELISA)
NPDFs were stimulated with LPS (1 µg/mL) for 48 h. VEGF production was determined by using ELISA (R&D Systems, Minneapolis, MN). Standards and samples were added and incubated at room temperature for 2 h. The cells were then washed three times, followed by the addition of VEGF conjugate and incubated for 2 h at room temperature. The reaction was stopped with stop solution, and the optical densities of the standards and samples were measured at 450 nm by using a microplate reader (Bio-Rad, Hercules, CA).
Immunofluorescence staining of VEGF and NF-κB
For immunofluorescence staining of VEGF, NPDFs were stimulated with LPS for 48 h. For immunofluorescence staining of NF-κB, NPDFs were stimulated with LPS for 1 h. NPDFs were fixed on coverslips using 4% paraformaldehyde, followed by treatment with 0.2% Triton X-100 in 1% bovine serum albumin for 10 min for membrane permeabilization. The cells were then blocked with 5% bovine serum albumin for 1 h at room temperature, followed by overnight incubation at 4℃ with monoclonal anti-VEGF and polyclonal anti-p50 antibodies. NPDFs were then incubated with anti-mouse Alexa 488 (Invitrogen) or anti-rabbit Alexa 555 (Invitrogen) secondary antibodies. Finally, NPDFs were counterstained using 4′-6-diamidino-2-phenylindole and visualized under a confocal laser scanning microscope (LSM700; Zeiss, Oberkochen, Germany).
Western blot analysis
NPDFs were incubated with LPS (1 µg/mL) for 1 h. For determining the steroid-mediated inhibition of TLR4 signaling pathways, NPDFs were stimulated with LPS after pretreatment with signaling pathway inhibitors: LY294002 (Akt inhibitor, 10 µmol/L) and BAY-11 (NF-κB inhibitor, 2.5 µmol/L). NPDFs were lysed in PRO-PREP™ protein extraction solution (iNtRON Biotechnology, Seongnam, Korea). Lysates were separated using 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride (PVDF) membranes (Millipore Inc., Billerica, MA). Membranes were blocked using 5% skim milk solution and incubated with the following antibodies: phospho-Akt, p50, and β-actin. Blots were visualized by using horseradish peroxide-conjugated secondary antibodies and an ECL system (Pierce, Rockford, IL).
Organ culture of nasal polyps
Nasal polyps were cut, using scissors, into 2 × 3 mm3 pieces under sterile conditions. Tissue fragments were washed three times with phosphate-buffered saline. The washed tissue fragments were placed on a prehydrated gelatin sponge (10 mm × 10 mm × 1 mm; Spongostan, Johnson & Johnson, San Angelo, TX) in six-well plates. And then, filled with 1.5 mL of culture medium containing Dulbecco’s Modified Eagle Medium (Invitrogen), 2% fetal bovine serum (Invitrogen) per well. 20 Nasal polyp tissues were stimulated with LPS (10 µg/mL) alone or co-treated with either steroids (DEX 5 µmol/L or FP 5 µmol/L) or LPS-RS (10 µg/mL) for 72 h. The plates were maintained at 37℃ in 5% CO2. The supernatant media of cultured nasal polyp tissues were used to determine the protein expression level of VEGF. Nasal polyp tissues were also used for the detection of the VEGF mRNA and protein levels.
Statistical analysis
Results were obtained from at least three independent experiments. The statistical significance of the differences between control and experimental data was analyzed by using unpaired t test or one-way analysis of variance (ANOVA) followed by Tukey’s test (GraphPad Prism, version 5, Graph Pad Software; San Diego, CA). Significance was determined at 95% confidence level. P values less than 0.05 were considered statistically significant.
Results
Steroids inhibit LPS-induced VEGF expression
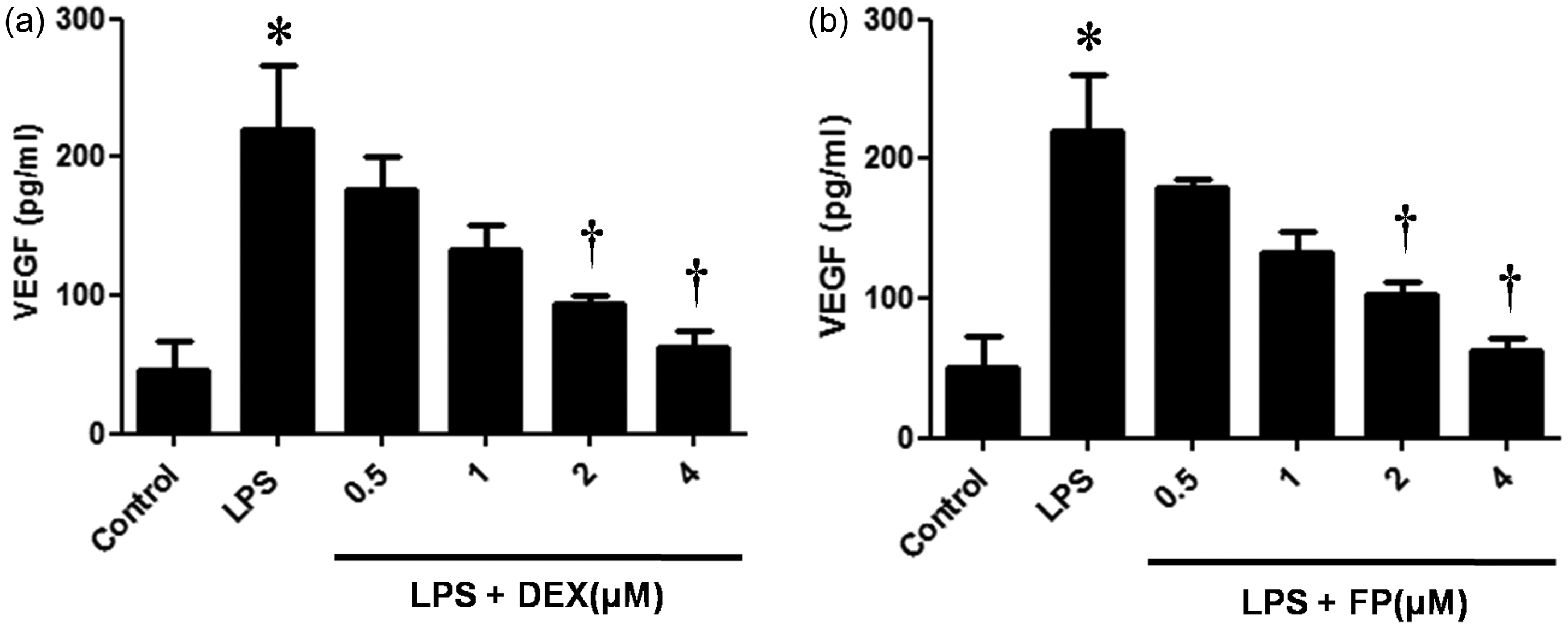
In this study, we investigated the effects of steroid treatment on VEGF expression in LPS-stimulated NPDFs. The steroids DEX and FP significantly inhibited LPS-induced VEGF mRNA expression (Figure 1a). The inhibitory effects of steroids on LPS-induced VEGF protein production were confirmed by ELISA and immunofluorescence staining (Figure 1b and c). Both steroids decreased LPS-induced VEGF protein levels in a dose-dependent manner (Figure 2).
Effect of steroids on lipopolysaccharide (LPS)-induced vascular endothelial growth factor (VEGF) expression. Nasal polyp-derived fibroblasts (NPDFs) were treated with LPS (1 µg/mL) alone or co-treated with either dexamethasone (DEX, 2.5 µmol/L) or fluticasone propionate (FP, 2.5 µmol/L). Expression level of VEGF mRNA was determined by reverse transcriptase-polymerase chain reaction (RT-PCR) analysis (a). Protein production of VEGF was measured by using enzyme-linked immunosorbent assay (ELISA) (b) and visualized by immunofluorescence staining (c). Data are represented as means ± SEM taken from three independent experiments. Images were acquired by using confocal laser scanning microscopy. *P < 0.05 as compared to the mean VEGF values of control cells; †P < 0.05 as compared to the mean VEGF values of the LPS stimulated cells. Scale bar = 50 µm. (A color version of this figure is available in the online journal.) Steroids inhibit lipopolysaccharide (LPS)-induced vascular endothelial growth factor (VEGF) protein expression in a dose-dependent manner. Nasal polyp-derived fibroblasts (NPDFs) were stimulated with LPS (1 µg/mL) alone or co-treated with dexamethasone (DEX) (a) or fluticasone propionate (FP) (b). VEGF protein levels were determined by using enzyme-linked immunosorbent assay (ELISA). Data are represented as means ± SEM. *P < 0.05 as compared to the mean VEGF values of control cells; †P < 0.05 as compared to the mean VEGF values of the LPS-stimulated cells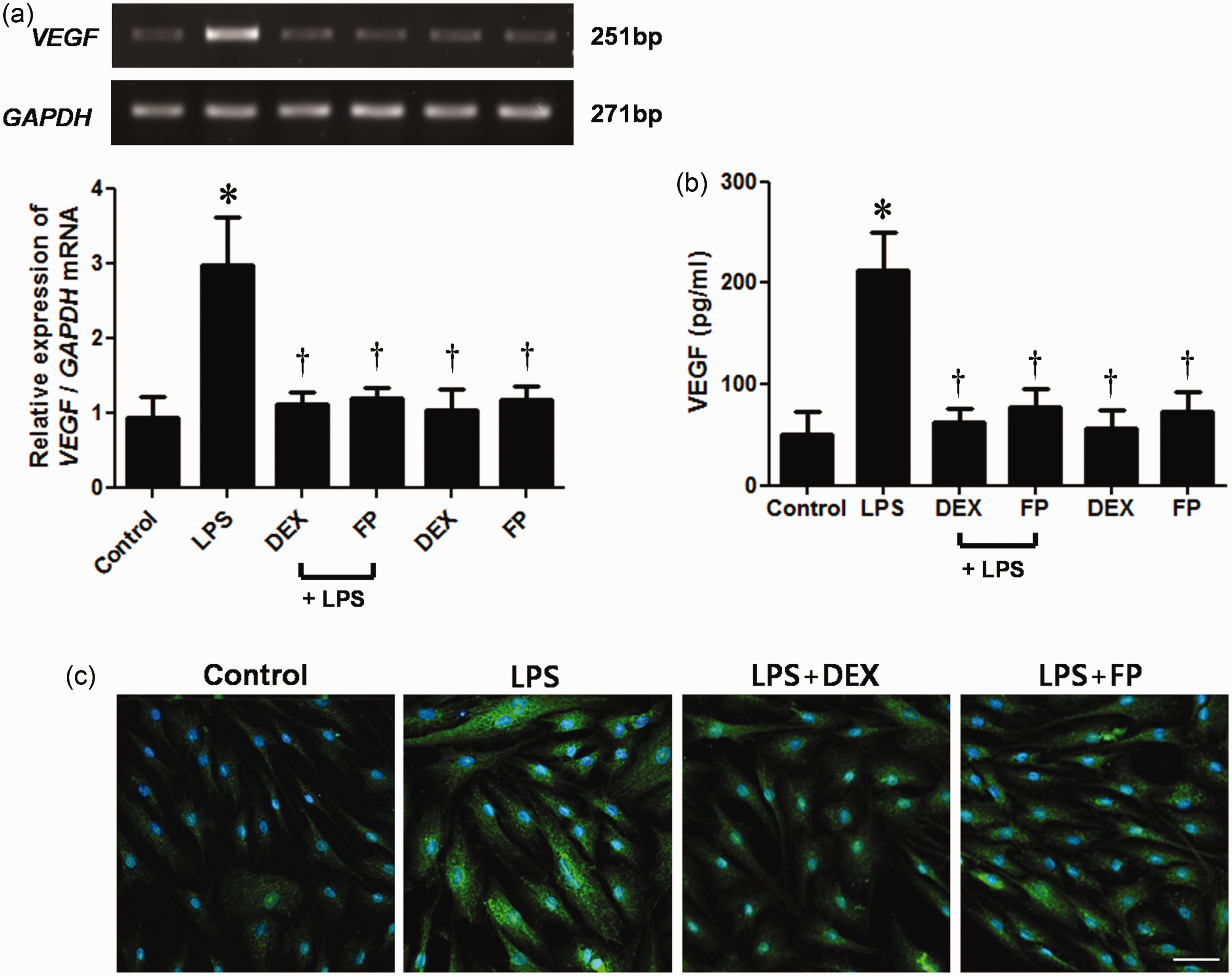
Steroids inhibit LPS-induced TLR4 and MyD88 expression
After LPS binds to TLR4, the receptor induces downstream signaling molecules such as MyD88.12,21 To elucidate the effects of steroids on TLR4 and MyD88 expression in LPS-treated NPDFs, we determined the mRNA expression levels of TLR4 and MyD88 by RT-PCR analysis. LPS stimulation of NPDFs resulted in increased TLR4 and MyD88 mRNA expressions; however, steroids (DEX and FP) treatment suppressed their expressions in LPS-treated NPDFs (Figure 3).
Effects of steroids on Toll-like receptor 4 (TLR4) and myeloid differentiation primary response gene 88 (MyD88) mRNA expression levels in lipopolysaccharide (LPS)-induced nasal polyp-derived fibroblasts (NPDFs). NPDFs were stimulated with LPS (1 µg/mL) alone or co-treated with dexamethasone (DEX) (2.5 µmol/L) and fluticasone propionate (FP) (2.5 µmol/L). TLR4 and MyD88 mRNA expression levels were determined by using reverse transcriptase-polymerase chain reaction (RT-PCR). Data are represented as means ± SEM. *P < 0.05 as compared to the mean relative mRNA levels of control cells; †P < 0.05 as compared to the mean relative mRNA levels of the LPS-stimulated cells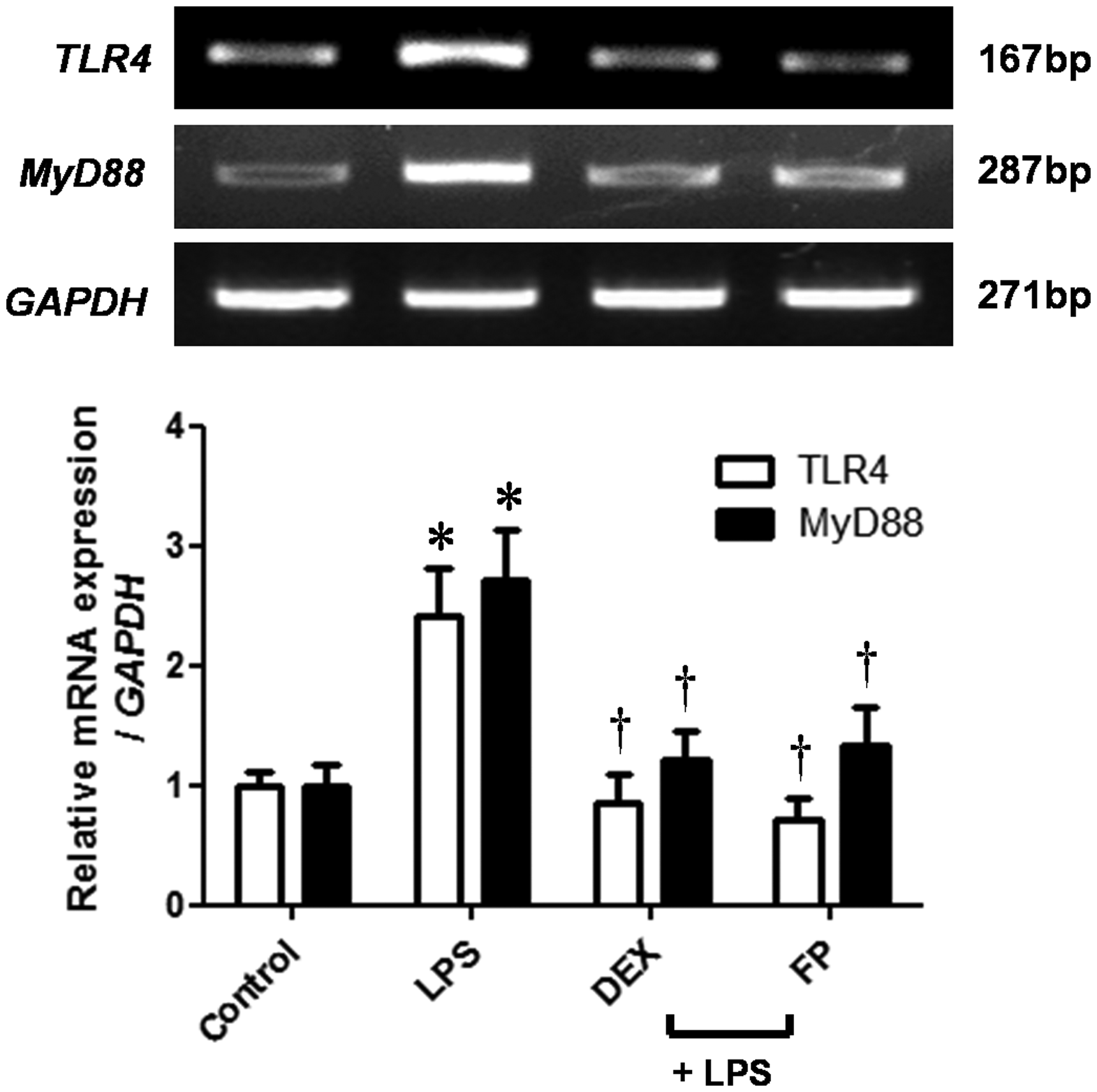
TLR4 mediates LPS-induced VEGF expression
NPDFs were transfected with siTLR4 before stimulating the cells with LPS for evaluating the role of TLR4 in mediating VEGF expression. RT-PCR analysis and ELISA were used for measuring VEGF expression. TLR4 mRNA expression was inhibited in siTLR4-transfected NPDFs (Figure 4a). Moreover, VEGF mRNA and protein expression were significantly inhibited by TLR4 silencing in LPS-induced NPDFs (Figure 4a and b).
Role of Toll-like receptor 4 (TLR4) on vascular endothelial growth factor (VEGF) expression in lipopolysaccharide (LPS)-induced nasal polyp-derived fibroblasts (NPDFs). NPDFs were transfected with siControl or siTLR4 and stimulated with LPS (1 µg/mL). (a) TLR4 and VEGF mRNA expression levels were accessed by using reverse transcriptase-polymerase chain reaction (RT-PCR). (b) VEGF production was determined by using enzyme-linked immunosorbent assay (ELISA). Data are represented as means ± SEM. *P < 0.05 as compared to the mean values of control cells; †P < 0.05 as compared to the mean values of the LPS-stimulated cells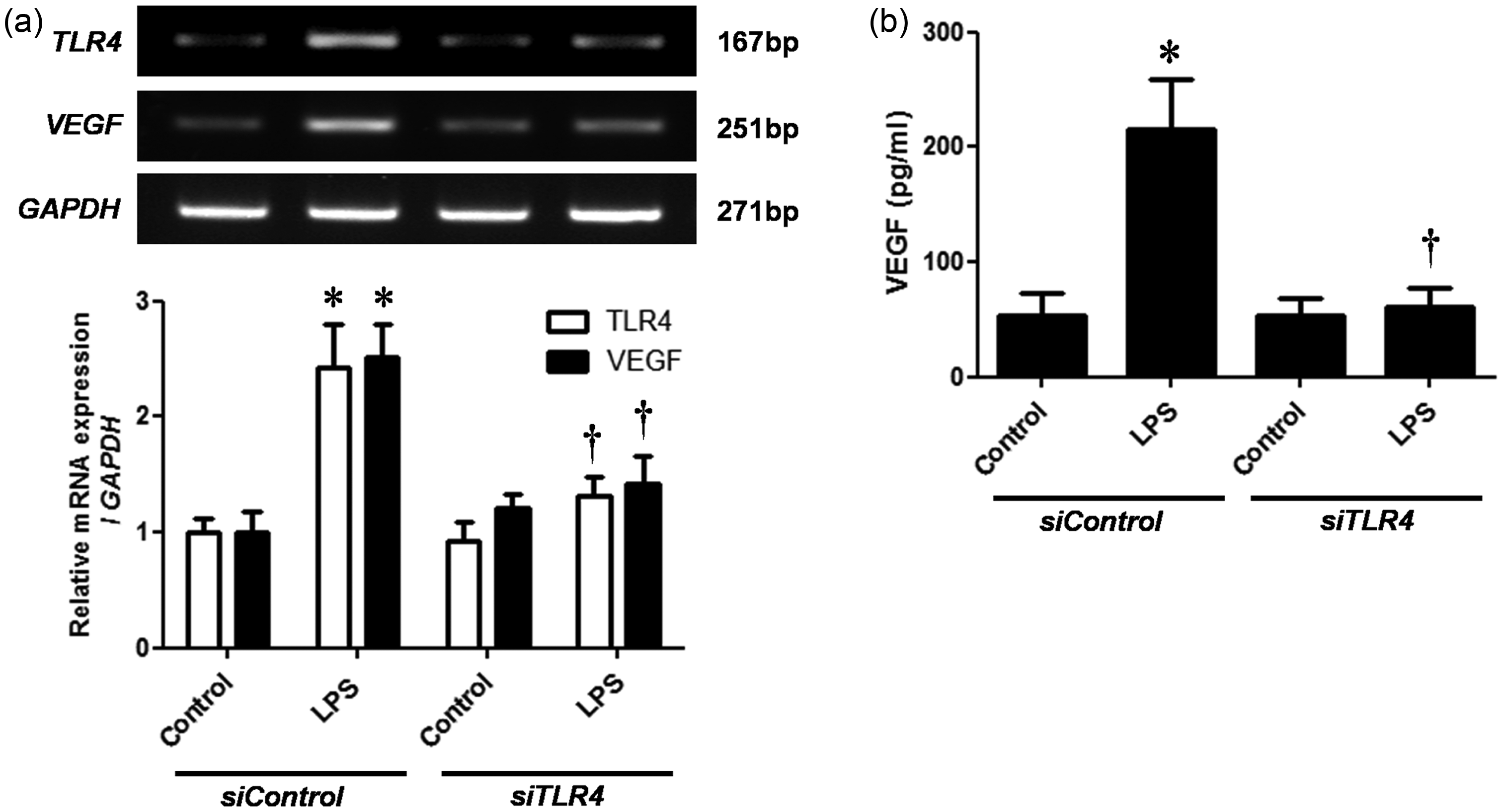
Akt regulates LPS-induced VEGF expression
VEGF secretion is Akt dependent, and Akt signaling pathway is critical for inducing LPS-induced inflammatory responses. To verify whether the Akt signaling pathway is involved in VEGF expression, we conducted NPDF stimulation studies by using LPS, steroids, and Akt inhibitor (LY294002). Western blot analysis showed that LPS activated Akt. However, steroids and LY294002 inhibited Akt activation (Figure 5a). In addition, LPS-induced VEGF production was significantly blocked by Akt inhibitor (Figure 5b).
Effects of steroids on lipopolysaccharide (LPS)-induced Akt signaling pathway in nasal polyp-derived fibroblasts (NPDFs). NPDFs were stimulated with LPS (1 µg/mL) alone or co-treated with either steroids (dexamethasone [DEX] or fluticasone propionate [FP]) or pretreatment with Akt inhibitor (LY294002, 10 µmol/L). (a) Activation of Akt pathways was determined by using Western blot analysis. (b) Vascular endothelial growth factor (VEGF) protein expression was measured by enzyme-linked immunosorbent assay (ELISA). Data are represented as means ± SEM. *P < 0.05 as compared to the mean values of control cells; †P < 0.05 as compared to the mean values of LPS-stimulated cells
NF-κB mediates LPS-induced VEGF expression
To identify NF-κB signaling pathways that are involved in VEGF expression, we treated LPS-stimulated NPDF cells with steroids or NF-κB inhibitor (BAY-11). Protein expression of p50 was inhibited by steroids, the Akt and NF-κB inhibitors (Figure 6a). Immunofluorescence staining studies showed that LPS stimulation induced p50 translocation to the nucleus. However, the p50 translocation was abrogated by the treatment with steroids, the Akt inhibitor, and the NF-κB inhibitor (Figure 6b). In addition, LPS-induced VEGF production was significantly blocked by the NF-κB inhibitor (Figure 6c).
Nuclear factor κB (NF-κB) activation in lipopolysaccharide (LPS)-stimulated nasal polyp-derived fibroblasts (NPDFs). NPDFs were stimulated with LPS (1 µg/mL) alone or co-treated with either steroids (dexamethasone [DEX] or fluticasone propionate [FP]) or pretreatment with inhibitors (Akt inhibitor LY294002, 10 µmol/L or NF-κB inhibitor BAY-11, 2.5 µmol/L). (a) NF-κB p50 protein expression levels were determined by using Western blotting. (b) NF-κB p50 protein translocation was determined by using immunofluorescence staining. (c) Vascular endothelial growth factor (VEGF) production was measured by using enzyme-linked immunosorbent assay (ELISA). Data are shown as the means ± SEM. *P < 0.05 as compared to the mean values of the control samples; †P < 0.05 as compared to the mean values of the LPS-stimulated cells. Images were acquired by confocal laser scanning microscopy. Scale bar = 50 µm. (A color version of this figure is available in the online journal.)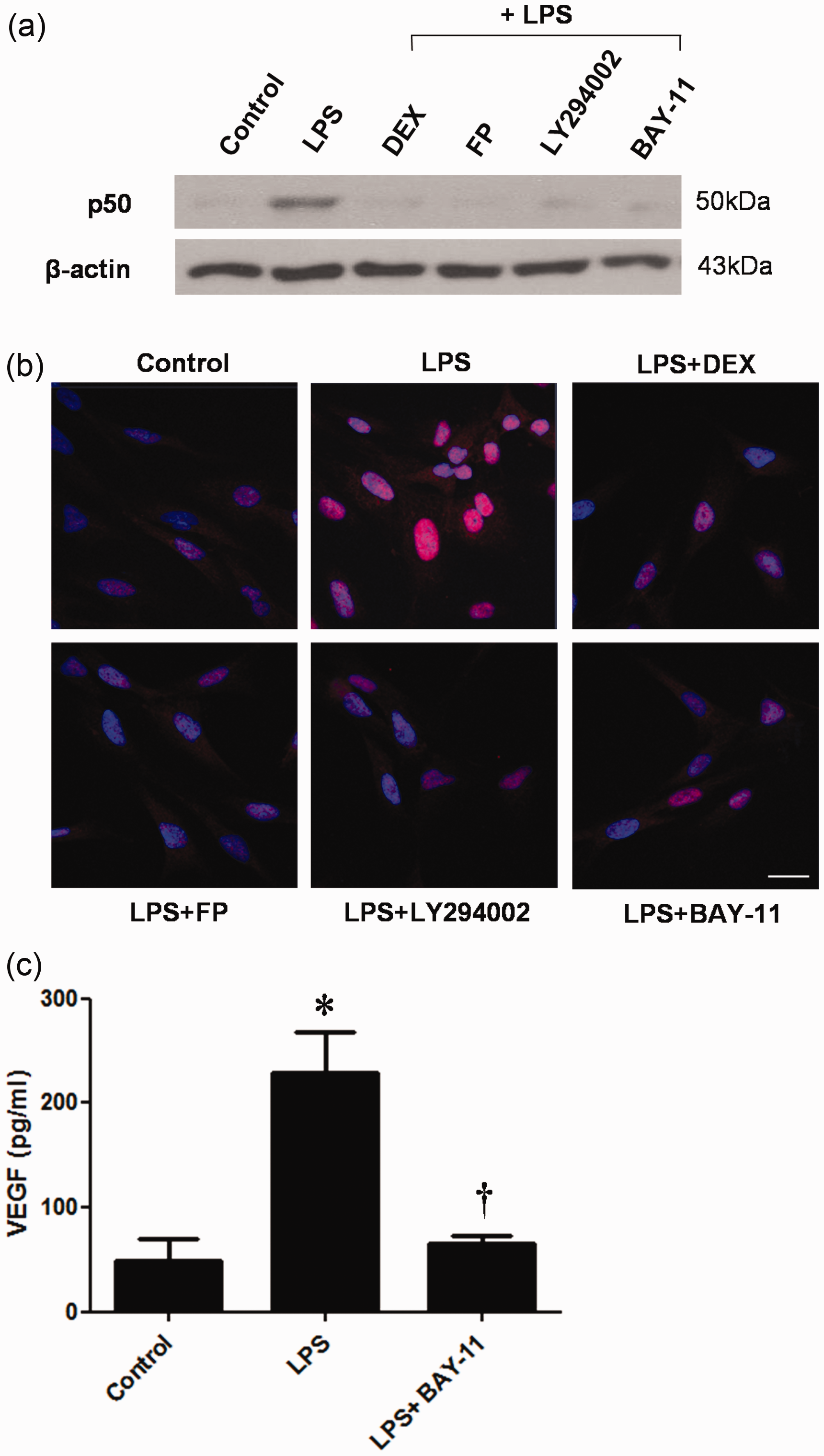
Steroids inhibit LPS-induced VEGF expression in nasal polyp tissues
To find VEGF expression in nasal polyp organ tissues, we treated the NPDFs with LPS with or without TLR4 antagonist (LPS-RS, LPS from Rhodobacter sphaeroides) and the steroids. VEGF mRNA level was inhibited by LPS-RS and steroids (Figure 7a). Furthermore, the production of VEGF was significantly reduced (Figure 7b).
Effect of steroids on lipopolysaccharide (LPS)-stimulated vascular endothelial growth factor (VEGF) expression in nasal polyp organ culture. Organ cultured nasal polyp tissues were stimulated with LPS (10 µg/mL) alone or co-treated with either steroids (dexamethasone [DEX] 5 µmol/L or fluticasone propionate [FP] 5 µmol/L) or LPS-RS (10 µg/mL). (a) VEGF mRNA expression levels were accessed by using reverse transcriptase-polymerase chain reaction (RT-PCR). (b) VEGF protein production was determined by using enzyme-linked immunosorbent assay (ELISA). Data are represented as means ± SEM. *P < 0.05 as compared to the mean values of the control samples; †P < 0.05 as compared to the mean values of the LPS-stimulated samples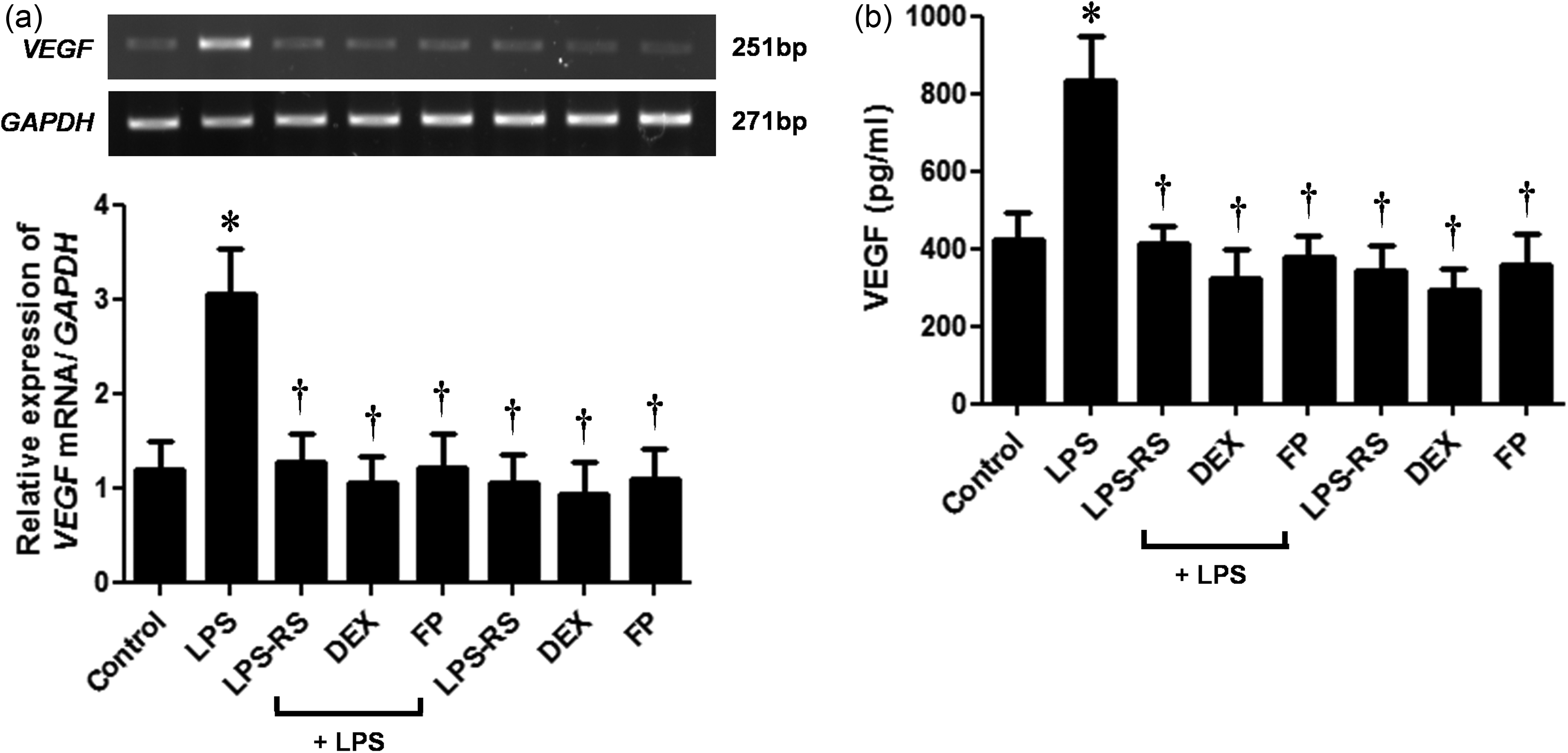
Discussion
Our study data show that LPS treatment significantly elevates VEGF production in NPDFs. TLR4 and MyD88 expressions are significantly increased in LPS-stimulated NPDFs. LPS stimulation results in Akt phosphorylation and NF-κB activation. In addition, steroids inhibit VEGF production induced by LPS via the TLR4/MyD88/Akt/NF-κB signaling pathway in a dose-dependent manner.
Chronic rhinosinusitis with nasal polyp is prolonged inflammation of the sinonasal mucosa. Nasal polyp is associated with tissue remodeling and increased immature microvessels. 10 Angiogenesis and vascular remodeling possibly play an important role in the growth process of nasal polyposis. 22 In this study, we demonstrated the inhibitory effects of steroids on VEGF production in NPDFs.
TLRs are pattern recognition receptors that bind to PAMPs. Characterization of TLR expression and immune responses in nasal polyps is essential for understanding the mechanisms involved in the formation and remodeling of nasal polyposis. 23 Previous mRNA studies indicate that all 10 mammalian TLRs are expressed in the sinonasal mucosa. 24 Airborne fungi stimulate TLR2, TLR3, and TLR4 expression and trigger immune responses in nasal polyp epithelial cells. 23 The sinonasal mucosa is continuously exposed to environmental agents and is thus a common site of microbial infection and microbial byproducts such as LPS. Nasal microorganisms are found in individuals with or without sinonasal diseases. LPS, a cell wall component of gram-negative bacteria, is a PAMP that is recognized by TLR4 expressed in the host tissue. 25 Previous studies have reported that LPS up-regulates VEGF expression in murine macrophages 14 and odontoblasts. 16 TLR4 signaling may be either MyD88-dependent or MyD88-independent. MyD88-dependent signaling leads to mitogen-activated protein kinase, Akt, and NF-κB activation, whereas MyD88-independent signaling involves TRIF-IRF3 activation. 12 Our data shows that LPS induces TLR4 signaling pathway that culminates in increased TLR4 and MyD88 expression.
VEGF is the most extensively characterized endothelial cell-specific angiogenic factor. VEGF, which is produced by many types of cells including neutrophils, fibroblasts, and endothelial cells, induces endothelial cell proliferation, increases vascular permeability, and is a key regulator of physiological and pathological angiogenesis. 7 Enhanced VEGF expression is localized in the endothelium, basement membrane, perivascular space, and epithelium of nasal polyps.9,22 Immunohistochemical analysis of nasal polyps showed increased VEGF staining within the vascular endothelium and increased mean blood vessel count, both of which correlated with the size of the nasal polyps. 26 Several studies have reported high levels of VEGF in the epithelium and endothelium of nasal polyps.10,22 VEGF plays an important role in tissue edema formation associated with nasal polyp development. 10 LPS triggers proinflammatory cytokine production and disrupts the microcirculation by increasing vascular permeability in experimental models. 16 Our study shows that LPS-induced activation of the TLR4 signaling pathway can lead to increased VEGF expression in NPDFs via TLR4/MyD88 pathway.
Steroids have been successfully used for nasal polyp treatment and have been shown to alleviate clinical symptoms, nasal peak flow, and decrease nasal polyp size. 27 Steroid-treated nasal polyp studies have shown down-regulation of proinflammatory cytokines and adhesion molecules that attract and activate eosinophils. 28 Steroids such as DEX and FP are among the most widely prescribed anti-inflammatory drugs. Several studies have demonstrated that steroid treatment down-regulates proinflammatory and proangiogenic cytokines in nasal polyps. 29 Numerous studies have also shown that DEX can inhibit VEGF expression in various cell types such as chondrocytes, 30 gastric mucosa cells, 31 and renal cell carcinoma cells. 32 However, the inhibitory mechanisms of steroids have not been elucidated. Therefore, we investigated the effects of DEX and FP on VEGF expression in NPDFs. In our study, steroids significantly decreased LPS-induced TLR4, MyD88, and VEGF expression. These results suggest that steroids inhibited LPS-stimulated VEGF production through TLR4 signaling pathway.
VEGF secretion is phosphatidyl-inositol 3-kinase (PI3K)/Akt/mammalian target of rapamycin (mTOR)-dependent. 33 Previous studies have indicated that Akt signaling pathway plays a critical role in LPS-induced in vitro and in vivo inflammatory responses. 33 Interestingly, Akt inhibition dramatically reduced LPS-induced reactive oxygen species production, lowered expression of inflammatory mediators such as VEGF, and inhibited NF-κB activation. 34 Taken together, these findings indicate that Akt signaling may regulate vascular leakage associated with the aggravation of airway inflammation through suppression of reactive oxygen species production and the expression of VEGF and various other inflammatory mediators that are induced by NF-κB activation. Our study shows that LPS-induced Akt phosphorylation and Akt inhibitor treatment decreased LPS-induced VEGF production in NPDFs. In addition, we showed that DEX and FP inhibited LPS-induced Akt activation. Thus, our data show that steroids inhibit LPS-stimulated VEGF production via the Akt signaling pathway.
PI3K/Akt pathway participates in various signaling pathways that lead to NF-κB activation that culminates in NF-κB-dependent transcription. 35 NF-κB is a transcription factor that is involved in proinflammatory cytokine production and can initiate inflammatory responses in various cells including inflammatory cells, cancer cells, and fibroblasts. 36 Akt-dependent phosphorylation of IκBα leads to degradation of IκBα thereby allowing translocation of NF-κB to the nucleus. 35 Recent evidence suggests that NF-κB activation increased angiogenesis as a result of elevated VEGF levels. LPS can induce VEGF gene expression via NF-κB activation in macrophages, 37 and dendritic cells. 38 Our studies show markedly increased NF-κB activation and nuclear translocation for transcriptional activation in LPS-stimulated NPDFs whereas DEX and FP treatment of LPS-stimulated NPDFs resulted in inhibition of NF-κB activation. Thus, these data show that steroids inhibit LPS-induced VEGF production via NF-κB signaling pathway. PI3K/Akt pathway has also been implicated in the regulation of hypoxia-inducible factor-1α (HIF-1α). 39 HIF-1α is regulated transcriptionally in an NF-κB-dependent manner and acts as a master transcription factor. 40 HIF-1α binds on VEGF gene promoter and transactivates VEGF gene expression. 41 To examine the effects of LPS or steroids on HIF-1α expression, NPDFs were treated with LPS with or without steroids. LPS induced HIF-1α expression, whereas steroids inhibited the increased HIF-1α expression (data not shown).
Nasal polyp organ cultures have been well established. 20 Nasal polyp organ cultures provide accessible means to mimic the in-vivo situation because native cell–cell contacts, cell–matrix integrity, and three-dimensional structure are maintained. Because an in-vivo model system for nasal polyposis has not been established, ex-vivo organ cultures of a nasal polyp would be a useful tool for research investigating nasal polyp treatment. In our study, we show LPS stimulates VEGF expression through TLR4 in nasal polyp organ cultures and steroids significantly inhibit the LPS-induced VEGF expression.
The underlying mechanisms for inhibitory effects of steroids on LPS-induced VEGF expression in nasal polyps are poorly understood. In summary, we have shown that the TLR4/MyD88/Akt/NF-κB pathway is mainly involved in LPS-induced VEGF production in NPDFs. In addition, our data indicate that steroids inhibit the VEGF production induced by LPS via TLR4/MyD88/Akt/NF-κB pathway NPDFs. Therefore, our findings have elucidated some of the mechanisms of nasal polyp pathogenesis and indicate that steroids could be used for nasal polyposis treatment.
Footnotes
Author contributions
All authors contributed to the design, interpretation of the studies, analysis of the data and review of the manuscript. J-SC and J-HK conducted the experiments and wrote the manuscript. H-ML supervised the project.
ACKNOWLEDGMENTS
This work was supported by the National Research Foundation of Korea (NRF) grant (no. 2011-0015346) and the Korean Health Technology R&D Project (Ministry of Health & Welfare, no. HI13C1501) funded by the Korea Government and a Korea University Grant.