
Abstract
Current androgen deprivation therapy often leads to androgen independence. However, mechanism of the therapeutic failure is still not well understood. Here, we demonstrate elevated expression of Zeb1 in androgen-independent prostate cancer cells and prostate tumors of castrated PTEN conditional knockout mice. While Zeb1 shRNA resulted in a sensitization of androgen-independent prostate cancer cells, forced Zeb1 expression caused androgen-dependent prostate cancer cells to be more resistant to androgen deprivation. Moreover, such effects appeared to be mediated by induction of pluripotent genes or stem cell-like properties. Collectively, these findings suggest that inhibition of Zeb1 might be a potential therapeutic strategy for treatment of androgen-independent prostate cancer.
Introduction
Although androgen deprivation therapy remains the mainstay therapeutic strategy for prostate cancer, there still exists a serious clinical problem, that is, initially androgen-dependent prostate cancer (ADPC) would eventually progress to androgen-independent prostate cancer (AIPC). Once it reaches the androgen independence stage, the disease will have a very poor prognosis, with a median survival of only approximately 12 months. 1 However, to date, the molecular mechanism underlying the androgen independence remains not well understood.
Our previous study has reported an interesting phenomenon that castration induces epithelial–mesenchymal transition (EMT) via a possible AR-Zeb1 bidirectional, negative feedback loop. 2 The EMT transcription factor Zinc-finger E-box-binding homeobox factor 1 (Zeb1), also known as TCF8 or deltaEF1, is proposed to play a crucial role in many types of human carcinomas such as breast cancer 3 and pancreatic cancer. 4 Very importantly, a recent study by Weinberg group indicates that Zeb1 is sufficient to switch the cells from a non-cancer stem cell to a cancer stem cell state and is required for the maintenance of the stemness of breast cancer stem cells. 3 However, whether and how down-regulation or up-regulation of Zeb1 influences androgen independence of prostate cancer have not been directly determined.
In the present study, we set forth to investigate the biological functions of Zeb1 in the development of androgen independence of prostate cancer. Our findings provide direct evidence indicating that Zeb1 promotes prostate cancer androgen independence and suggest that Zeb1 can be a potential therapeutic target for treatment of AIPC.
Materials and methods
Cell lines
Human prostate cancer cell lines, including LNCaP, C4-2B, PC3 AR(+), PC3, CW22-RV1, and DU145 (ATCC), were grown in RPMI-1640 or Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% FBS (Gibco), 100 U/ml penicillin, and 0.1 mg/mL streptomycin (Gbico) at 37℃ in humidified 5% CO2 atmosphere.
Quantitative real-time PCR
Total RNA was extracted with Trizol reagent (Invitrogen) and 2 µg total RNA was reverse transcribed for cDNA synthesis using the high-capacity reverse transcription kit (Applied Biosystems). Quantitative real-time PCR (qRT-PCR) amplification was performed on the Applied Biosystems Step One Plus® Real-Time PCR Systems (Singapore). A panel of primer pairs was used following the SYBR® Green method (Toyobo, Japan) (Supplementary Table 1). Specific gene expression was normalized to β-actin levels using the ΔΔCt method, with each sample being carried out in triplicates.
Immunoprecipitation and Western blotting
For immunoprecipitation (IP), cells were washed with ice-cold PBS and lysed in a lysis buffer supplemented with proteinase inhibitors (Cocktail Kit, Prod#78410, Rockford, USA) and phosphatase inhibitors (Roche). Cell lysates were incubated with a specific antibody or equal quality IgG for IP overnight at 4℃. Protein G Agarose anti-Rabbit IP Beads (Roche) were then added and incubated for another 4 h at 4℃. The immunoprecipitates were washed six times with the IP buffer and boiled for 10 min in 1 × SDS loading buffer. Immunoprecipitated proteins were detected by following Western blot.
For Western blot assays, total cell proteins were extracted from all the cell lines using a modified radioimmunoprecipitation buffer (RIPA, Prod#89901, Rockford, USA) supplemented with proteinase inhibitors. The protein concentration was examined by the BCA Protein Assay (Bio-Rad, Hercules, CA). All primary antibodies were used at a 1:1000 dilution and incubated at 4℃ overnight. Antibody against Zeb1 was purchased from Cell Signaling Technology Inc. (#3396). Antibody against E-cadherin was purchased from BD Transduction Laboratories™ (#610181). Antibody against Vimentin (#2707-1) and Sox2 (#2683-1) were purchased from Epitomics, an Abcam Company. After washing three times with TBST, the membrane was incubated with secondary antibodies (1:1000, Cell Signaling Technology) against horseradish peroxidase-conjugated rabbit or mouse immunoglobulin G and the signals were measured by enhanced chemiluminescence Western blotting system (Millipore). Antibody against β-actin (Cell Signaling Technology Inc. #4970) was used as an internal loading control.
Immunofluorescent staining
All the cell lines were grown on Chamber slides in an appropriate medium as described above for 24 h. The cells were then fixed with 4% paraformaldehyde for 15 min at room temperature. After washing three times with PBS for 5 min each, the cells were blocked with a blocking buffer for 1 h at room temperature. After blocking, the cells were incubated with primary antibodies at 4℃ overnight. Then, the cells were washed three times with PBS followed by incubation with a relevant fluorochrome-conjugated secondary antibody for 2 h at room temperature in dark. After three washes with PBS, the slides were mounted with VECTASHIELD Mounting Medium with DAPI (Vector Laboratories, Inc., #H-1200, Burlingame, CA).
Plasmids, transfection, and lentivirus production
Zeb1 shRNA were cloned into lentiviral vector pUCTP. The cDNA of Zeb1 was cloned into lentiviral expression vector pLV.Des2d.P/puro. The sequences of Zeb1 shRNA were listed in Supplementary Table 2.
Lentiviruses were produced by co-transfection of 293 T cells with a lentiviral vector, and packaging vectors pCMV-DR8 and pMD2.2 using the calcium phosphate transfection method. At 48 h post-transfection, lentiviral supernatant was collected, filtered and mixed with polybrene (8 µg/mL), then added directly into target cell lines for infection. Starting 48 h post-infection, infected cells were then selected with 5 µg/mL puromycin for about 14 days to generate the stable transfectants.
Cell proliferation assay and drug sensitivity assay
Cell Counting Kit 8 (CCK-8) (Dojindo, Kumamoto, Japan) was used to analyze the rate of cell proliferation. Cells were seeded into 96-well plates with 5000 cells per well and cultured for 24 h, 48 h, 72 h, and 96 h. For drug sensitivity assay, cells were treated with different doses of bicalutamide. Then, 10 µL of CCK-8 solution was added to each well and incubated for 2 h. The optical density (OD) value was detected at a wave-length of 450 nm on the microplate reader (BioTek). Cell survival rate was calculated follow the formula below: (OD (treated) − OD (blank))/(OD (control) − OD (blank)) × 100%.
Colony assay and sphere assay
For colony assay, 1000 cells per well were plated in 6-well culture plate and cultured for about two weeks. Then, colony cells were fixed with 4% paraformaldehyde and using 0.1% crystal violet for stain. For sphere assay, 500 DU145 cells and 1000 LNCaP cells per well were plated in six-well ultra low cluster plate (Corning, NY, USA) and cultured in DMEM:F12 with EGF (20 ng/mL), bFGF (20 ng/mL)and B27 (2%) for about one week. Further analysis was performed by counting the colony and sphere numbers under a Nikon microscope.
Cell migration assays
Transwell assay
The top chambers were seeded with 3 × 104 cells in serum-free media and 20% FBS was used as the chemoattractant in the bottom chamber (Corning, #3422). Then, migrated cells were fixed with 4% paraformaldehyde and using 0.1% crystal violet for stain. For further quantification of the migrated cells, five random regions of the filter were counted under a Nikon microscope. Three filters were used and the experiments were carried out in triplicates.
Wound healing assay
Scratch wounds were made in confluent cell monolayers using a pipette tip. Monolayers were washed twice with PBS and the cells were cultured in serum-free media. Cell migration was recorded in five different microscopic fields.
In vivo growth assay and mouse strains
Five-week-old male BALB/C nude mice (SLAC, Shanghai) were injected s.c. in the flank with 5 × 106 LNCaP cells or 2 × 106 DU145 cells mixed with an equal volume of matrigel (BD Biosciences, Bedford, MA, USA). Tumor growth was evaluated by measuring the length and width with a caliper and the volume of each tumor was calculated by the following formula: volume = π/6 × tumor length × tumor width 2 .
PTEN conditional knockout mice were generated as previously described. 5 To investigate the association of Zeb1 with androgen independence, some PTEN deletion mice and male athymic nude mice which harbored prostate tumor cells were castrated and some received sham surgery as controls. All mice were housed and bred in the SPF animal facility of Ren Ji Hospital in a pathogen-free environment with controlled temperature and humidity.
Statistical data analysis
Statistical evaluation was conducted using the Student’s t-test and presented as the mean ± SEM of at least three independent experiments. Two-sided probability value less than 0.05 was considered to be statistically significant. Prism GraphPad5 was used to perform all the plots and data analysis.
Results
Zeb1 is up-regulated in AIPC cells compared to ADPC cells
To examine whether Zeb1 expression is up-regulated during development of androgen
independence, we utilized three pairs of AIPC and ADPC cell lines including
C4-2B vs. LNCaP, PC3 vs. PC3-AR+, and DU145 vs. CW22-RV16,7 and compared
their expression levels of Zeb1. As shown in Figures1(a) to (d), androgen-independent cells showed a
higher level of Zeb1 and Vimentin, but a lower level of E-cadherin at both mRNA
and protein levels, as revealed by qRT-PCR and immunoblotting analyses.
Immunofluorescence staining also showed the elevated expression of Zeb1 in the
AIPC cells as compared to those ADPC cells (Figure 1(e)). The level of Zeb1 is higher in AIPC cell lines
than ADPC cell lines. (a) to (d) qRT-PCR and Western blot analysis
of Zeb1, Vimentin, and E-cadherin expression in ADPC cell lines
(LNCaP, PC3 AR(+), and 22RV1) and AIPC cell lines (C4-2B, PC3, and
DU145). Actin was used as a negative control. (e) Further validation
of Zeb1, Vimentin, and E-cadherin expression in all the cells
mentioned above using immunofluorescent staining. Bars, 50 µm; **,
P < 0.01.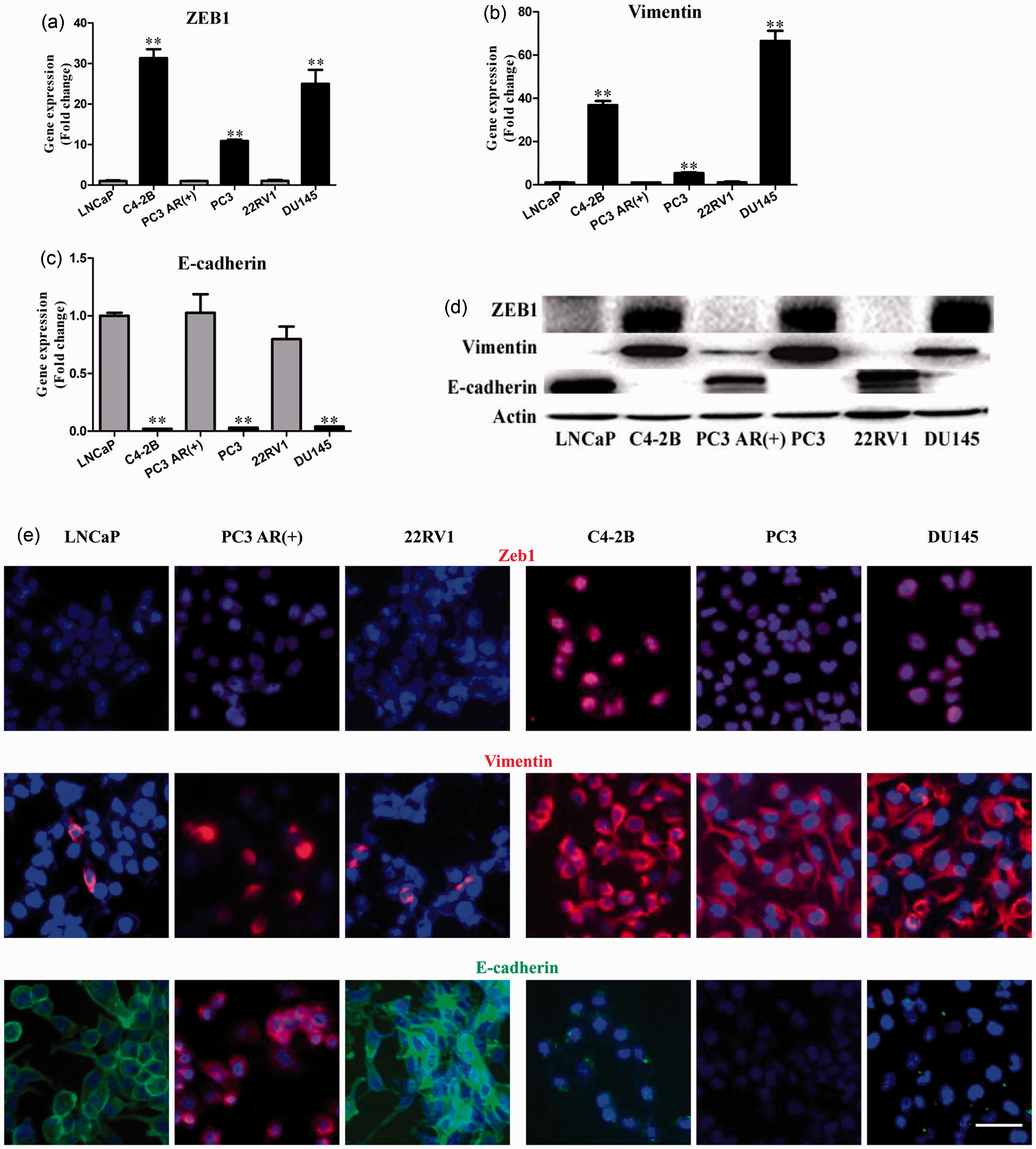
To extend the observations from these cell lines to clinical scenarios, we
analyzed microarray data from the Gene Expression Ominibus (GEO) database and
found that Zeb1 was indeed significantly increased in the patients suffering
from AIPC compared with those with ADPC (Figure 2(a),
P < 0.05) (GDS1390). To provide additional supporting data
for the positive correlation between Zeb1 expression and AIPC, we used the PTEN
conditional knockout mouse prostate cancer model.
8
As shown in Figure 2(b), prostate
tumors from the Pten−/− mice after castration displayed enhanced Zeb1
expression compared to those mice with a sham surgery. Similar enhanced level of
Vimentin was also observed in the Pten−/− prostate tumors after
castration (Figure
2(c)). Interestingly, double immunostaining for Vimentin and E-cadherin
showed that some of the tumor cells co-expressed both Vimentin and E-cadherin
(Figure 2(c)),
supporting the notion that these cells are in the process of undergoing EMT.
Collectively, these data strongly suggest that there is a positive correlation
between Zeb1 expression and AIPC. Zeb1 is increased in
AIPC clinical specimens or prostate tumors in castrated PTEN
conditional knockout mice. (a) Zeb1 expression levels in
androgen-dependent and androgen-independent patients. The data are
based on the analysis of GDS1390 from the NCBI/GEO database. (b) and
(c) Zeb1,Vimentin, and E-cadherin expression in the intact and
castrated PTEN conditional knockout mice by immunofluorescent
staining. Note that higher level of Zeb1 is seen in castrated mouse
prostate tumors. E-cadherin/Vimentin co-localization is observed
after castration. Bars, 50 µm; **,
P < 0.01.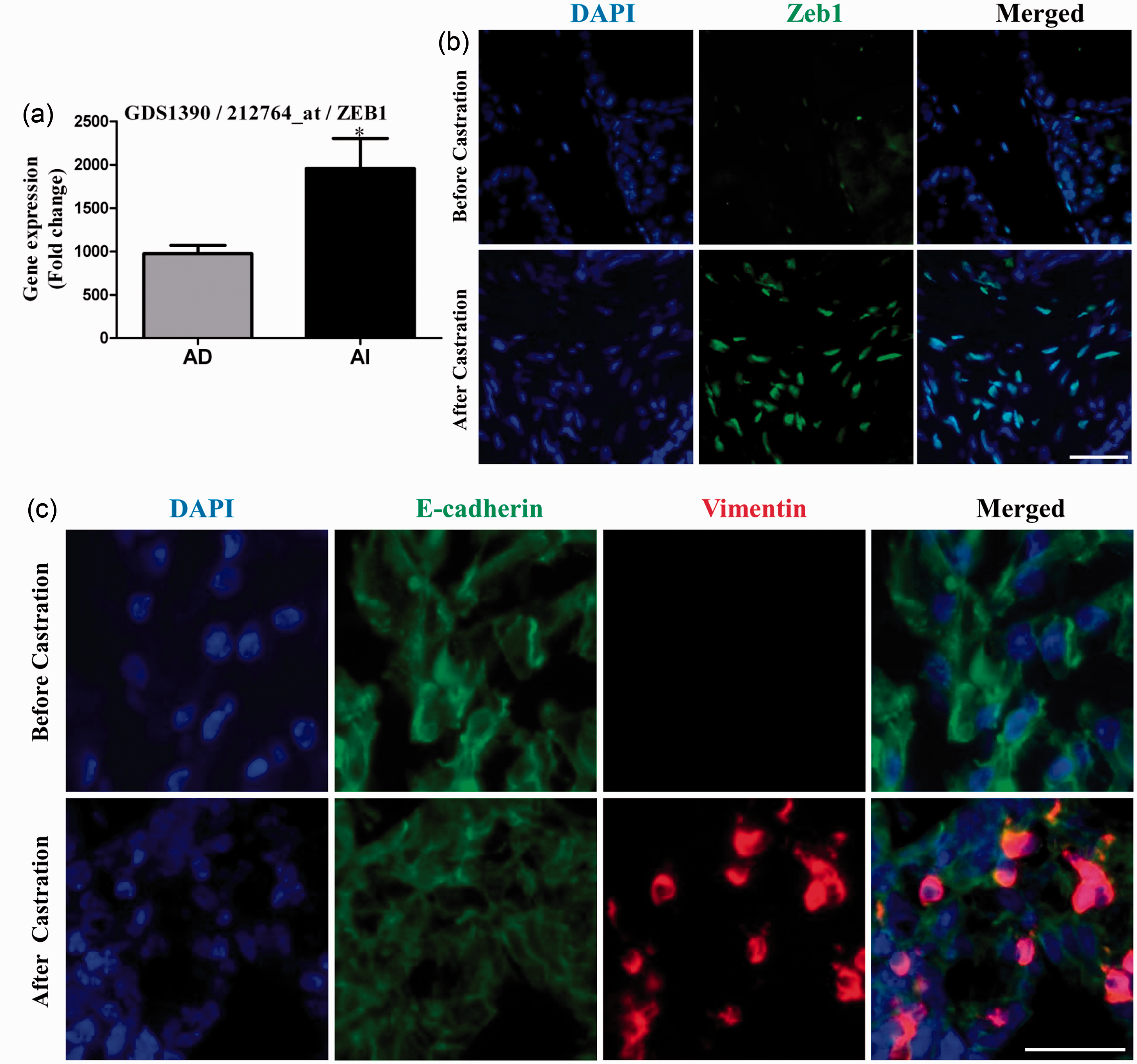
Zeb1 expression is elevated in prostate cancer cell lines that show higher migration ability and in clinical prostate tumor specimens with angio-lymphatic invasion
An important clinical symptom of AIPC is the distal metastasis. As expected, AIPC
PC3 cell had a higher migration ability than ADPC PC3 AR (+) cell (Figure 3(a) to (b),
P < 0.01). In addition, data analysis from the Oncomine
database revealed significantly increased Zeb1 expression in the patients with
angio-lymphatic invasion compared with those without angio-lymphatic invasion
(Figure 3(c),
P < 0.05).9,10 To further investigate
whether Zeb1 plays a role in the metastasis of AIPC, we analyzed the GEO
database that contains benign, malignant, and metastatic prostate cancer
specimens. Notably, compared to the benign prostates, Zeb1 level was
significantly higher not only in prostate cancer tissues but also in metastatic
prostate tumor specimens (Figure 3(d)) (P < 0.01, GDS 3289). Taken
together, these results demonstrate that Zeb1 expression is elevated in AIPC
cells that have a higher migration capability and its expression level is
positively correlated with the tumors with angio-lymphatic invasion. Elevated Zeb1 expression in AIPC cell lines
shows higher migration ability and is associated with
angio-lymphatic invasion. (a) and (b) Representative transwell
migration assays display a higher migration rate in AIPC cells (PC3)
compared with the ADPC cells (PC3 AR+). (c) Zeb1 expressions between
specimens with or without angio-Lymphatic invasion. The data are
based on the Wallace study. (d) Differential Zeb1 expressions in the
benign, prostate intraepithelial neoplasia (PIN), prostate cancer
and metastatic prostate cancer tissues. The data are based on
analysis of GDS3289 from the NCBI/GEO database. Bars, 50 µm; *,
P < 0.05; **, P < 0.01.
(A color version of this figure is available in the online
journal.)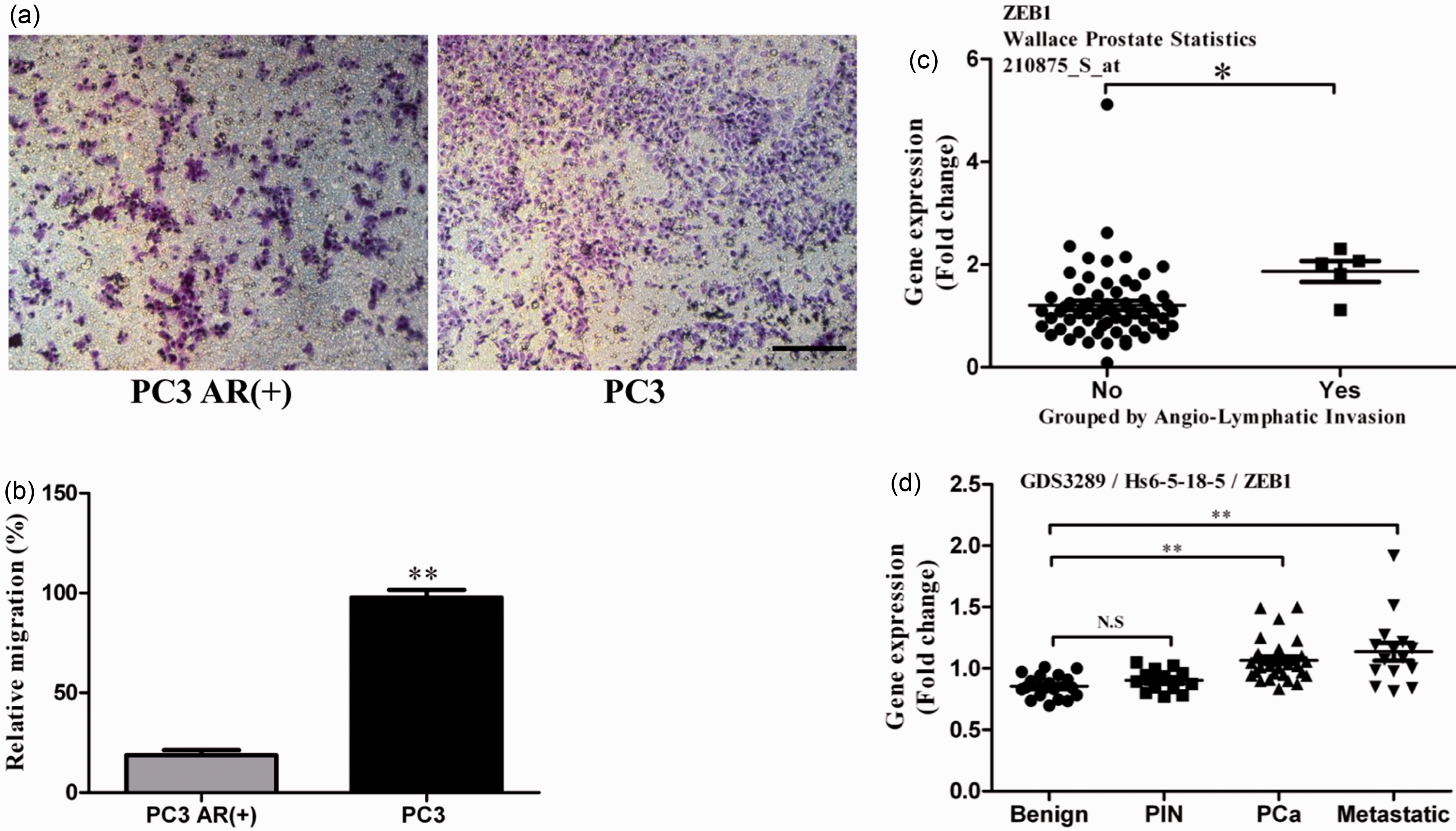
Forced expression of Zeb1 enhances androgen independence and migration ability of prostate cancer cells
To directly validate the significance of Zeb1 expression in the development of
AIPC, we transfected a lentivival vector containing ectopic Zeb1 expression into
LNCaP cells which showed relatively low Zeb1 expression at both mRNA and protein
levels (Figure 1(a) and
(d)). Both RT-PCR
and immunostaining confirmed overexpression of Zeb1 in LNCaP cells at both mRNA
and protein levels (Supplementary Figures 1(a) and (b)). As shown in Figure 4(a), after
treatment with different doses of bicalutamide, an androgen agonist, cell
survival rate was much higher in LNCaP-Zeb1 cells than in the vector control
cells, indicating that overexpression of Zeb1 significantly enhanced androgen
independence in the LNCaP cells that were originally androgen-dependent. To
further investigate the effect of Zeb1 on the development of AIPC
in vivo, we performed xenograft experiments in adult male
athymic nude mice with LNCaP-Ctl and LNCaP-Zeb1 cells in Matrigel. When the
tumors reached approximately 600 mm3 in size, the mice were
castrated. As shown in Figure
4(b), the LNCaP-Zeb1 cells had a significantly higher growth
advantage following castration of the host than those LNCaP-Ctl cells. These
data indicate that overexpression of Zeb1 causes these cells to become more
resistant to androgen deprivation treatment. Ectopic expression of
Zeb1 promotes androgen-independent and migration in prostate cancer
cell lines. (a) A dose–response curve of LNCaP cells transfected
with control or Zeb1 vectors displays relative bicalutamide
sensitivity by CCK8 assay. (b) Response of LNCaP-Zeb1 cell growth in
athymic male nude mice to castration. LNCaP-Ctl and LNCaP-Zeb1 cells
mixed with Matrigel are injected into the flank of the mice. When
the volume of the tumors reached approximately 600 mm3,
the mice were castrated and the size of the tumors was monitored by
weekly measurements with a caliper. ▪, average size of LNCaP-Zeb1
tumors (n = 5); Δ, average size of LNCaP-Ctl tumors
(n = 6). The growth curves are significantly
different (P < 0.05). Data are expressed as
Mean ± SE. (c) and (d) Representative transwell migration assays
display a higher migration rate in prostate cancer cells transfected
with the Zeb1 vector compared with the control vector. Bars, 50 µm;
*, P < 0.05; **, P < 0.01.
(A color version of this figure is available in the online
journal.)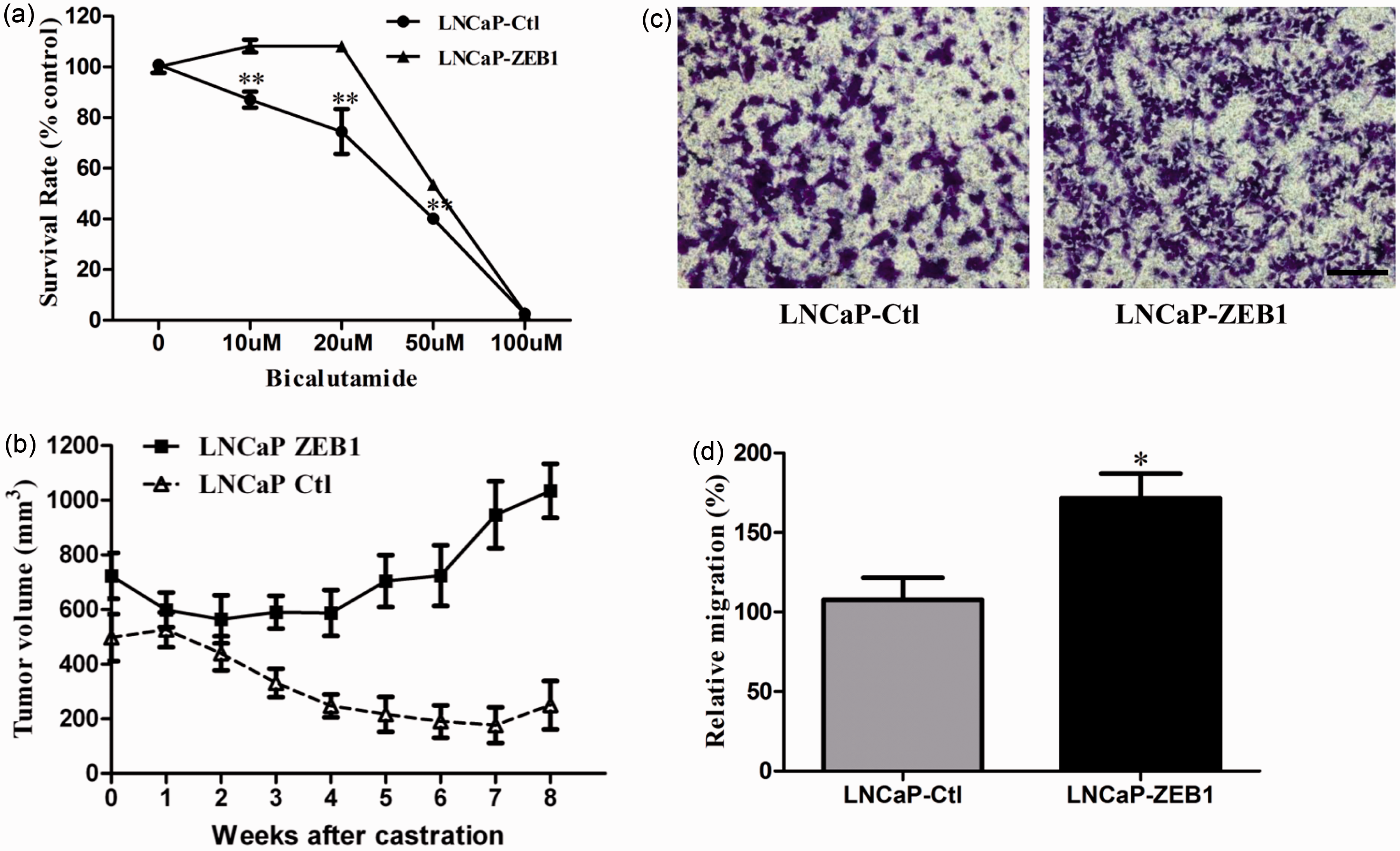
In addition, overexpression of Zeb1 in LNCaP cells also led to an elongated and scattered mesenchymal morphology in vitro (Supplementary Figure 2(a)), one of the properties of cells undergoing EMT. We also found that overexpression of Zeb1 in LNCaP cells caused a significant decrease of E-cadherin expression compared with the vector control (Supplementary Figure 2(b)). Because expression of the epithelial marker E-cadherin is negatively associated with the ability of cell migration, we studied the migration capacity between Zeb1 transfected cells and vector control cells using a transwell migration assay. As shown in Figure 4(c) to (d), overexpression of Zeb1 resulted in a higher migration rate in LNCaP cells (P < 0.05).
Knockdown of Zeb1 inhibits androgen independence and migration ability of prostate cancer cells
To determine whether knockdown of Zeb1 could influence androgen independence and
migration ability of prostate cancer cells, we transfected a lentiviral shRNA
targeting Zeb1 into DU145 and PC3 cell lines which displayed relatively high
Zeb1 expression at both mRNA and protein levels (Figure 1 (a) and (d)). RT-PCR analysis confirmed an
approximate 70% reduction in mRNA level (Supplementary Figure 1(c) and (d)).
Down-regulation of Zeb1 caused DU145 cells to be more sensitive to bicalutamide
(Figure 5(a),
P < 0.05). In addition, we also found that
down-regulation of Zeb1 in PC3 cells caused a reversed EMT process, called as
mesenchymal-to-epithelial transition (MET), with morphological changes from an
elongated to a more round, tightly packed shape (Supplementary Figure 2(c)).
Furthermore, as expected, increased E-cadherin expression was observed in these
cells after transfection with sh-Zeb1 (Supplementary Figure 2(d),
P < 0.01). We then investigated whether down-regulation
of Zeb1 led to an inhibition of prostate cancer cell migration capacity. Using a
transwell assay and a wound healing assay, we found that the migration rate of
DU145 sh-Zeb1 cells was significantly lower than DU145 vector control cells
(Figure 5(b) to
(d),
P < 0.01), indicating that Zeb1 knockdown suppresses the
ability of cell migration. Similar results were also observed in PC3 cells
(Supplementary Figure 3(a) to (c)). Zeb1 knockdown
attenuates androgen-independence and inhibits migration and
tumorigenesis ability in prostate cancer cell lines. (a) Knockdown
of Zeb1 induces prostate cancer cells to become more sensitive to
bicalutamide treatment by CCK8 assays. (b) to (d) Representative
transwell migration assays and wound closure assays display a lower
migration rate in prostate cancer cells transfected with sh-Zeb1
compared with the control vector. (e) An in vivo
xenograft model shows an inhibition of growth in DU145 cells
transfected with sh-Zeb1 as compared to the scramble shRNA. Bars,
50 µm; *, P < 0.05; **,
P < 0.01. (A color version of this figure is
available in the online journal.)
To determine whether Zeb1 knockdown could inhibit the tumorigenicity of the prostate cancer cells in vivo, we performed xenograft experiments using androgen-independent DU145 prostate cancer cells. Compared with DU145 vector control, DU145 sh-Zeb1 significantly suppressed tumorigenesis capacity in the xenograft mouse model in vivo (Figure 5(e), P < 0.01). The average size of the tumors decreased approximately 1.5-fold when Zeb1 was knockdown. Consistent with these results, cell proliferation assays in DU145 in vitro with sh-Zeb1 or LNCaP with Zeb1 also showed an inhibitory or stimulatory effect, respectively (Supplementary Figure 4(a) and (b)). Collectively, these results indicate that knockdown of Zeb1 attenuates androgen independence and inhibits migration and tumorigenesis ability of prostate cancer cells.
Zeb1 enhances stem cell properties including up-regulation of Sox2, colonogenesis and sphere formation of prostate cancer cells
As Zeb1 is an activator enhancing EMT, and EMT has been implicated in the
acquisition of cancer stem cell properties in pancreatic cancer
4
and breast
cancer
11
, we hypothesized that Zeb1 expression may affect the traits
of CSCs in prostate cancer cells. To test this hypothesis, we analyzed the CSCs
properties in prostate cancer cells in which Zeb1 was up-regulated or
down-regulated, respectively. The characteristics of CSCs include the ability of
self-renewal, differentiation, and homeostatic control as well as the expression
of pluripotent stem cell genes such as Sox2 and resistance to
apoptosis.4,12 As shown in Figure 6(a) and (b), overexpression of Zeb1 in LNCaP cells
resulted in elevated expression of stem cell markers such as Sox2 at mRNA and
protein levels. In contrast, knockdown of Zeb1 led to a reduced expression of
Sox2 in DU145 cells (Figure
6(c)). Interestingly, after analyzing the microarray data in the GEO
database of mouse prostate tissues, we found that both Zeb1 and Sox2 levels were
increased two weeks after castration and were reversed to a relatively normal
level following androgen replacement (P < 0.01, GDS 2562)
(Figure 6(d) to
(e)), suggesting a
positive correlation between Zeb1 and Sox2. Moreover, IP and Western blot
analyses revealed that Zeb1 protein might bind to Sox2 (Figure 6(f)). Support for this notion
comes from the work of Erik Engelen
13
using mass spectrometry
analysis. Zeb1 induces stem cell-like properties of prostate
cancer cells. (a) to (c) Stem cell marker expression in cells
transfected with ectopic Zeb1and sh-Zeb1. Note that the alteration
of Sox2 mRNA and protein levels is positively associated with Zeb1
expression. (d) and (e) Zeb1 and Sox2 expression changes after two
weeks post castration and three days of treatment with testosterone.
The data are based on the analysis of GDS2562 from the NCBI/GEO
database. (f) IP assay and Western blot analyses revealed that Zeb1
protein might bind to Sox2. (g) and (h) Colony formation assays
display a lower clonogenic rate in prostate cancer cells transfected
with sh-Zeb1 compared with the control vector. (i) and (j)
Colony-formation assays show that cells transfected with the Zeb1
vector grow faster than those transfected with the control vector.
(k) and (l) Sphere assays show reduced sphere-forming ability in the
sh-Zeb1 transfectants compared with vector control cells. (m) and
(n) Sphere assays show enhanced sphere-forming ability in the Zeb1
transfectants than those cells transfected with a control vector.
Bars, 50 µm; *, P < 0.05; **,
P < 0.01. (A color version of this figure is
available in the online journal.)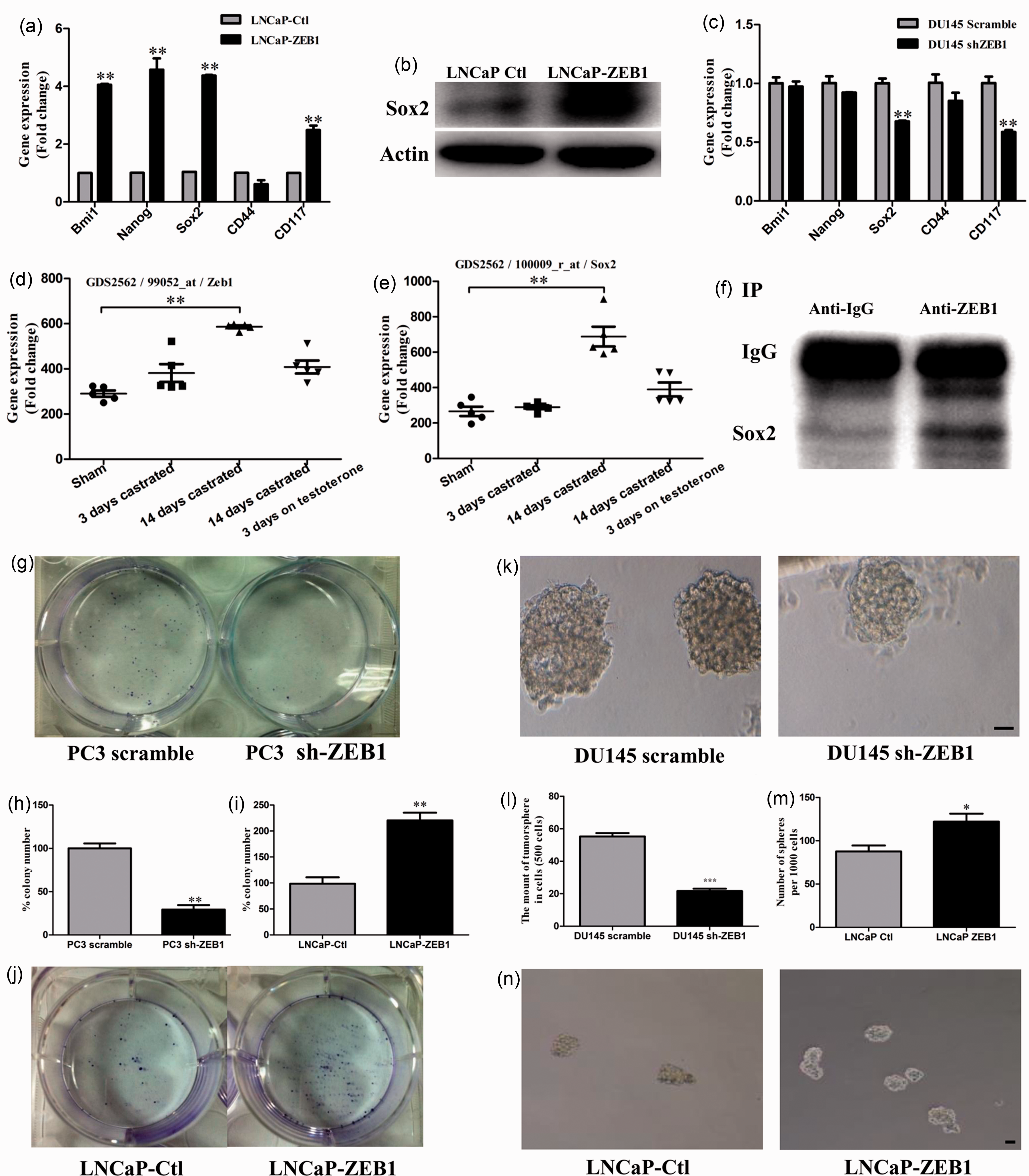
To provide further direct evidence for the correlation of Zeb1 and stem cell-like properties, we examined the ability of self-renewal using the colony formation and sphere formation assays. Compared to the control group, reduced colonogenesis (Figure 6(g) and (h)) and decreased number of prostasphere (Figure 6(k) and (l)) were observed in sh-Zeb1 transfected group under the same culture conditions. In contrast, enhanced colonogenesis (Figure 6(i) and (j)) and increased number of prostasphere (Figure 6(m) and (n)) were observed in Zeb1 transfected group than the control group. These results support the proposal that Zeb1 presumably enhances stemness of the prostate cancer cells.
Discussion
Although our previous study 2 reports that castration induces EMT in the normal prostate and prostate cancer, whether an EMT regulator can influence the androgen independence and metastasis via gene silence and overexpression approaches is undetermined. The present study demonstrates clearly that an important EMT modulator Zeb1 promotes androgen independence, migration and invasion of prostate cancer. Overexpression of Zeb1 causes the ADPC cells to become more androgen-independent and enhances migration. Conversely, interference of Zeb1 induces an opposite effect, that is, more sensitive to androgen deprivation and reduced migration and tumor formation abilities.
The role of Zeb1 in the development of therapeutic resistance has been implicated in pancreatic cancer 14 and glioblastoma cells 15 , however, to the best of our knowledge, our finding is the first to provide direct evidence that Zeb1 could influence androgen independence in prostate cancer. Firstly, the present experiments extend the observations reported in the previous study indicating the differential expression of EMT molecules including Zeb1 between androgen-dependent LNCaP and androgen-independent PC3 prostate cancer cells. 2 In the present experiments we examine three better paired androgen-dependent and independent prostate cancer cells: LNCaP vs. C2-4B; PC3 (AR+) vs. PC3; CW22-RV1 vs. DU145. We find that all three androgen-independent prostate cancer cells express higher levels of Zeb1 than those androgen-dependent prostate cancer cells. Secondly, clinical AIPC samples display a higher level of Zeb1 than those ADPC samples. Thirdly, in the PTEN conditional knockout mice model, castration causes a significant elevation in Zeb1 expression with co-expression of E-cadherin and Vimentin. Although it is possible that castration or androgen deprivation might induce production of EMT-inducing growth factors such as TGF-beta in the prostate tissue, which in turn causes the EMT process, future experiments are required to verify this possibility. Fourthly, overexpression of Zeb1 in ADPC cells leads to increased resistance to bicalutamide (a common anti-androgen drug) in vitro and continued tumor growth even after castration in vivo. Finally, knockdown of Zeb1 elicits opposite effects. These data together support the idea that Zeb1 can act as an important modulator for the development of AIPC.
The present study reinforces the notion that Zeb1 promotes invasion and migration of prostate cancer cells. Firstly, knockdown of Zeb1 in PC3 cells reverses EMT or causes MET with increased E-cadherin expression and epithelial morphology. As a result, Zeb1 knockdown leads to a reduced capacity of migration in both PC3 and DU145 cell lines. Secondly, overexpression of Zeb1 elicits opposite effects. Finally, our data analyses from the GEO and Oncomine databases provide further evidence that Zeb1 plays an important role in prostate tumorigenesis and invasion, that is, high level of Zeb1 has been observed in both prostate tumors and the tumors with angio-lymphatic invasion. Combining with our previous study, 2 these data together suggest that Zeb1 is a promoting factor for prostate tumorigenesis and invasion.
Increasing evidence suggest that EMT is often activated during cancer invasion and metastasis with the acquisition of stem cell-like properties. 11 In the present study, we investigate whether Zeb1 expression influences acquisition of stem cell characteristics by prostate cancer cells. We find that over-expression of Zeb1 increases the expression of the pluripotent stem cell markers such as Sox2, accompanied by enhanced colonogenesis and sphere-forming capacities, common properties of CSCs. Furthermore, IP assay suggests that Zeb1 protein might bind to Sox2, although whether such binding is specific and whether Zeb1 can directly regulate Sox2 remain to be verified in future studies. Together, these results suggest that the EMT-activator Zeb1 is strongly correlated with stem-cell like properties possibly through binding to Sox2 protein. Our findings are not only consistent with the previous studies indicating that normal prostate stem cells display higher expression levels of Zeb1 than those non-stem cells 2 but also are in agreement with the work done in pancreatic cancer, 4 glioblastoma, 15 and breast cancer. 3 For example, a recent study linking EMT and CSCs by Weinberg and colleagues demonstrates that Zeb1 is capable to convert non-CSCs to a CSC status and functions as an important modulator in the stemness maintenance of breast cancer cells. 3 Thus, these data together support the notion that high expression levels of Zeb1 facilitate the induction of stemness of CSCs in prostate cancer cells.
Taken together, the present study provides direct evidence that Zeb1, an important EMT modulator, promotes prostate cancer progression including androgen independence, invasion, and metastasis. Such progression-promoting effects by Zeb1 appear to be mediated via induction of stem cell-like properties. Therefore, reversing the EMT by targeting Zeb1 in combination with the traditional androgen deprivation therapy might be a conceivable approach in the war against AIPC.
Footnotes
Author contributions
PL, RY, and WQG participated in the design, execution, analysis and interpretation of the experiments and wrote the paper; JW, MC and KZ contributed to the experimental design and assisted with animal experiments. All authors have approved the final version of the manuscript.
ACKNOWLEDGEMENTS
This research project is supported by funds from the Chinese Ministry of Science and Technology (2012CB966800 to WQG and RY and 2013CB945600 to WQG and RY), the National Natural Science Foundation of China (81130038 and 81372189 to WQG), Science and Technology Commission of Shanghai Municipality (Pujiang program to WQG), Shanghai Education Committee Key Discipline and Specialty Foundation (J50208 to WQG), Shanghai Health Bureau Key Discipline and Specialty Foundation (to WQG), and KC Wong foundation (to WQG). Shanghai Jiao Tong University School of Medicine Doctor Innovation Foundation (BXJ201217 to PL).