
Abstract
Asthma is a common chronic inflammatory disease characterized by the infiltration and accumulation of memory-like Th2 cells and eosinophils. Viral infection has emerged as the most common cause of severe episodes of asthma. For the treatment of bronchial asthma, the root of liquorice (Glycyrrhiza glabra) has been used as a traditional medicine in the East and West. Licochalcone A is the predominant, characteristic chalcone in liquorice root. To determine whether licochalcone A possesses an anti-inflammatory effect, we tested its effect on the expression and production of thymic stromal lymphopoietin (TSLP) in BEAS 2B cells and primary bronchial epithelial cells. We found that polyinosinic-polycytidylic acid (poly-IC)-induced TSLP expression was suppressed by treatment with licochalcone A in a dose- and time-dependent manner. We also found that poly-IC-induced mRNA expression of other proinflammatory mediators such as MCP-1, RANTES, and IL-8 was suppressed by licochalcone A. Furthermore, licochalcone A suppressed poly-IC-induced nuclear factor kappa B (NF-κB) nuclear translocation and DNA-binding activity by suppressing the Iκβ kinase (IKK) activity but not by direct phosphorylation of p65 at serine 276. Collectively, our findings suggest that licochalcone A suppresses poly-IC-induced TSLP expression and production by inhibiting the IKK/NF-κB signaling pathway, which might be involved in the pathogenesis of virus-exacerbated asthma. Further elucidation of the mechanisms underlying these observations can help develop therapeutic strategies for virally induced asthma.
Keywords
Introduction
Asthma is a common chronic inflammatory disease characterized by the infiltration and accumulation of memory-like Th2 cells and eosinophils. 1 Rhinovirus (RV) infection has emerged as the most common cause of severe episodes of asthma in children and adults,2,3 and other viruses such as respiratory syncytial virus (RSV) have also shown predisposing attributes. 4
The root of liquorice has been used in Eastern and Western systems of traditional medicine for the treatment of bronchial asthma, gastric ulcer, and inflammation. 5 Licochalcone A is a phenolic compound found in the roots of both Glycyrrhiza glabra and Glycyrrhiza inflata and is the main characteristic chalcone isolated from the root of Xinjiang liquorice. 6 Studies have shown that licochalcone A possesses radical-scavenging effects 7 ; antileishmanial activity; and antimicrobial activity, including growth inhibition of Staphylococcus aureus and suppression of Helicobacter pylori activity.8,9 However, little is known about the effect of licochalcone A on asthma.
Thymic stromal lymphopoietin (TSLP) is an interleukin-7-like epithelial cell-derived cytokine (pro-allergic cytokine), which might be a major factor in the pathogenesis of asthma. Bronchial biopsy studies have demonstrated the overexpression of TSLP mRNA in the bronchial epithelial lining in severe asthma. 10 The potential role of TSLP in virally exacerbated asthma is additionally supported by findings that double-stranded RNA—a viral infection surrogate stimulus—can evoke overexpression of TSLP and overproduction of its protein in primary bronchial epithelial cells (PBECs) obtained from asthmatic individuals compared to epithelial cells from non-asthmatic individuals. 11 Inducible expression of TSLP in airway epithelial cells has been shown to be controlled by nuclear factor kappa B (NF-κB).12,13
Recent studies show that TSLP expression is induced in airway epithelial cells exposed to pro-inflammatory mediators, including tumor necrosis factor alpha (TNFα), lipopolysaccharide (LPS), and polyinosinic-polycytidylic acid (poly-IC), by NF-κB activation.12,14 Furthermore, licochalcone A has been found to inhibit TNFα-induced NF-κB activation by the inhibition of IκB kinase complex activation. 15 Interestingly, another study has demonstrated that licochalcone A inhibits LPS-induced NF-κB activation by direct inhibition of p65 phosphorylation at serine 276 (Ser 276). 16
Therefore, in the present study, we aimed to investigate the inhibitory effect of licochalcone A on poly-IC-induced TSLP expression and related mechanisms.
Materials and methods
Cell culture
BEAS 2B cells and PBECs were obtained from the American Type Culture Collection (Manassas, VA). BEAS 2B cells were grown in RPMI-1640 with 10% fetal bovine serum (FBS) and maintained at 37℃ in a humidified atmosphere of 5% CO2 and 95% air. PBECs were cultured in bronchial epithelium growth medium (Lonza) in flasks coated with collagen and fibronectin for at least three weeks in total as described. 17 Cells were plated in 24 - or 6-well plates (EM), grown to confluence and placed overnight in BEBM containing transferrin, insulin, gentamicin, and amphotericin B (Sigma-Aldrich).
Real-time reverse transcriptase-PCR
Total RNA was isolated from BEAS 2B cells and PBECs using an Easy-BLUE Total RNA Extraction Kit (iNtRON Biotechnologies, Seoul, Korea) after exposure to poly-IC and/or licochalcone A. Total RNA (2 µg) was reverse transcribed using the oligo (dT) primer and murine leukemia virus (MMLV) reverse transcriptase (Promega, Madison, WI) at 42℃ for 90 min. Real-time polymerase chain reaction (PCR) was performed using an ABI Prism 7500 instrument according to the manufacturer’s instructions (Applied Biosystems, Foster City, CA). The following primer pairs were used: TSLP, forward 5′-TATGAGTGGGACCAAAAGTACCG-3′ and reverse, 5′-GGGATTGAAGGTTAGGCTCTGG-3′; MCP-1, forward 5′-TGAGGTGGT TGTGGAAAAGG-3′ and reverse, 5′-CCTGCTGTTCACAGTTGCC-3′; RANTES, for ward 5′-TCCCCATATTCCTCGGAC-3′ and reverse 5′-GATGTACTCCCGAACCCA-3′; IL-8, forward5′-GGCACAAACTTTCAGAGACAG-3′ and reverse 5′-ACACAG A GCTGCAGAAATCAGG-3′; and GAPDH, forward 5′-GGCCAAAAGGGTCATCATC-3′ and reverse 5′-GTGATGGCATGGACTGTGG-3′. After an initial hot start for 10 min, amplification was performed for 40 cycles consisting of denaturation for 10 s at 94℃, annealing for 30 s at 56℃, and extension for 40 s at 72℃. The amplification kinetics was recorded as sigmoid progress curves for which fluorescence was plotted against the number of amplification cycles. The threshold cycle number (CT) was used to define the initial amount of each template. The CT was the first cycle for which a detectable fluorescent signal was observed. The mRNA expression levels were determined and compared with the GAPDH standard.
Preparation of whole cell lysates and nuclear extracts
The whole cell lysates were prepared by radio immunoprecipitation assay (RIPA) lysis buffer (50 mmol/L Tris-HCl, 150 mmol/L NaCl, 1% NP-40, 0·25% sodium deoxycholate, 2 mmol/L ethylenediamine tetraacetic acid [EDTA]) containing a mixture of protease inhibitors (Sigma, St Louis, MO). Nuclear extracts were prepared using 200 mL of lysis buffer (10 mmol/L HEPES, pH 7·9, 10 mmol/L KCl, 0·1 mmol/L EDTA, 0·1 mmol/L ethyleneglycol tetraacetic acid [EGTA]) and incubated on ice for 15 min. At the end of this incubation, 10 mL of 10% NP-40 was added and the tube was vortexed for 10 s. After centrifugation at 12,000 g for 10 min at 4℃, supernatants were discarded and the pellets were processed further to obtain nuclear extracts. The pellets were resuspended in the extraction buffer (20 mmol/L HEPES, 1 mmol/L glycerol, 0·4 mmol/L NaCl, 1 mmol/L EDTA, 1 mmol/L EGTA) and incubated for 30 min at 4℃. Nuclear extracts were isolated by centrifugation at 12,000 g for 30 min at 4℃. The supernatants were stored at 80℃ until used for Western blot analysis.
Western blot analysis
The cell extracts were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto a nitrocellulose membrane. The membranes were blocked in blocking solution (5% non-fat dried milk in phosphate-buffered saline [PBS]) for 2 h at room temperature and then probed with anti-NFκB p50, anti-NFκB p65, anti-P-p65(s276), anti-P-p65(s536), anti-Iκβ kinase α (anti-IKKα), anti-IKKβ, anti-IKKγ, anti-β-actin, anti-IκBα, and anti-Lamin B (Santa Cruz Technology, Santa Cruz, CA) for 1 h at room temperature. After washing three times in PBS containing 0.1% Tween-20 (PBS-T), the membranes were incubated with secondary antibodies (Jackson Immunoresearch, West Grove, PA) for 1 h at room temperature. After washing an additional three times in PBS-T, the membranes were developed using an electrochemiluminescence (ECL) solution (Pierce, Rockford, IL, USA) and exposed to Kodak X-ray film.
Electrophoretic mobility shift assay (EMSA)
EMSA was performed as described previously. 15 In brief, a synthetic double-strand oligonucleotide corresponding to NFκB motif of TSLP promoter, 5′-CTGCTAGGGAA-ACTCCATTATTAC-3′ was radiolabeled with [g32P]ATP and T4 polynucleotide kinase (Promega, Madison, WI) and purified using Sephadex G-25 Quick Spin Columns (Roche, Mannheim, Germany). DNA–protein complexes were separated on 6% (w/v) polyacrylamide gels in Tris/glycine buffer, pH 8.3. The dried gels were exposed to X-ray film.
In vitro IKK assay
Beas 2B cells (1 × 107 cells) were preincubated with or without 30 µmol/L Licochalcone A (Sigma) for 1 h at 37℃ after stimulation with or without 5 µg/mL Poly-IC (Sigma) for 30 min. Cell lysates were immunoprecipitated with anti-IKKγ antibody with protein G-Sepharose (Zymed Laboratories, South San Francisco, CA) for 2 h at 4℃ and then washed three times with lysis buffer and twice with kinase buffer (25 mmol/L HEPES-NaOH, pH 7.5, 20 mmol/L MgCl2, 20 mmol/L glycerophosphate, 0.1 mmol/L Na3VO4, 2 mmol/L dithiothreitol, and 20 mmol/L p-nitrophenylphosphate). The kinase reaction in 20 µL of kinase buffer including [γ-32 P]ATP was carried out with 1 µg of GST-IκBα as a substrate for 20 min at 30℃. Samples were resolved by SDS polyacrylamide gel electrophoresis, and phosphorylated GST-IκBα was visualized by autoradiography.
Measurement of TSLP secretion
BEAS 2B cells and PBECs were incubated with indicated doses of Licochalcone A for 24 h after stimulation with Poly-IC (5 µg/mL) for 30 min, and then the TSLP concentration in each supernatant was quantified using an human TSLP quantikine ELISA kit from R&D Systems (Minneapolis, MN, USA) according to the manufacturer’s instructions.
Statistical analysis
All results are expressed as the mean ± standard error of the mean (SEM). The statistical evaluation of the results was performed by an independent t-test and an analysis of variance (ANOVA) with a Tukey post hoc test. The results were significant with a value of P < 0.05.
Results
Licochalcone A significantly inhibited poly-IC-induced expression and secretion of TSLP
To evaluate the direct effect of licochalcone A on poly-IC-induced expression of TSLP in airway epithelial cells, we performed real-time PCR in BEAS 2B cells. When BEAS 2B cells were treated with various doses of licochalcone A and at different times after poly-IC stimulation for 30 min, TSLP expression was down-regulated in a dose- and time-dependent manner (Figure 1a, b), suggesting that licochalcone A directly down-regulates poly-IC-induced expression of TSLP in airway epithelial cells. In addition, licochalcone A potently suppressed poly-IC-induced secretion of TSLP in BEAS 2B cells (Figure 1c). No cytotoxic effect of licochalcone A treatment and/or poly-IC stimulation was observed in our experimental conditions (data not shown).
Effect of licochalcone A on poly-IC-induced TSLP expression and secretion. Beas 2B cells were incubated with indicated doses of licochalcone A for 3 h after stimulation with poly-IC (5 µg/mL) for 30 min, and then real-time PCR was performed. The values are normalized relative to the GAPDH standard (a). Beas 2B cells were incubated at indicated times of licochalcone A (30 µmol/L) after stimulation with poly-IC (5 µg/mL) for 30 min, and then real-time PCR was performed (b). Beas 2B cells were incubated with indicated doses of licochalcone A for 24 h after stimulation with poly-IC (5 µg/mL) for 30 min, and then the TSLP concentration in each supernatant was quantified using a ELISA kit (c). All data are representative of three independent experiments. Values represent the means ± SEM. *P < 0.05, **P < 0.005 vs. poly-IC. ND: not detected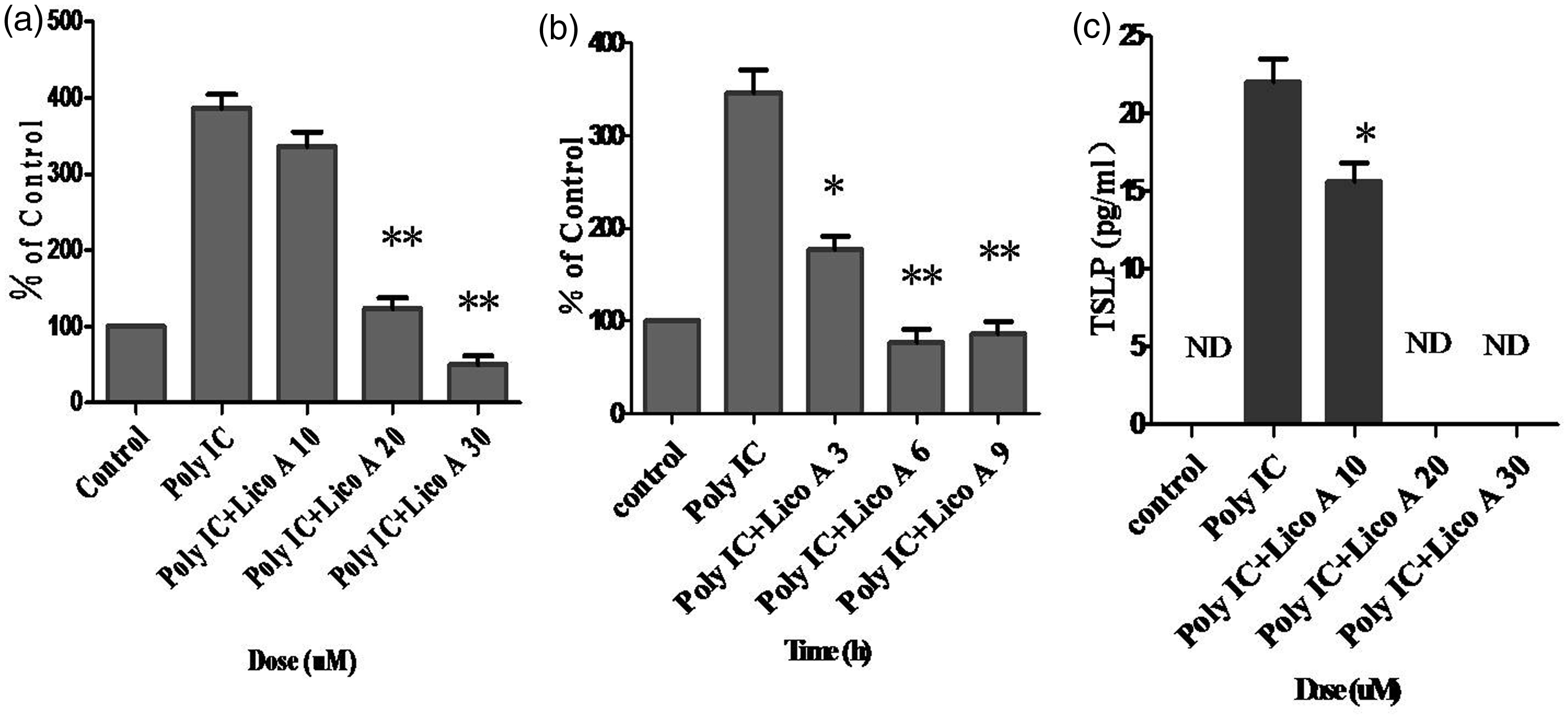
We also investigated whether licochalcone A could inhibit the poly-IC-induced expression and secretion of TSLP in PBECs. When PBECs were treated with various doses of licochalcone A after poly-IC stimulation for 30 min, TSLP mRNA was down-regulated dose dependently (Figure 2a). In addition, licochalcone A potently suppressed poly-IC-induced secretion of TSLP in PBECs (Figure 2b).
Effect of licochalcone A on poly-IC-induced TSLP expression and secretion in PBECs. PBECs were incubated with indicated doses of licochalcone A for 3 h after stimulation with poly-IC (5 µg/mL) for 30 min, and then real-time PCR performed. The values are normalized relative to the GAPDH standard (a). PBECs were incubated with indicated doses of licochalcone A for 24 h after stimulation with poly-IC (5 µg/mL) for 30 min, and then the TSLP concentration in each supernatant was quantified using a ELISA kit (b). All data are representative of three independent experiments. *P < 0.05, ***P < 0.001 vs. poly-IC. ND: not detected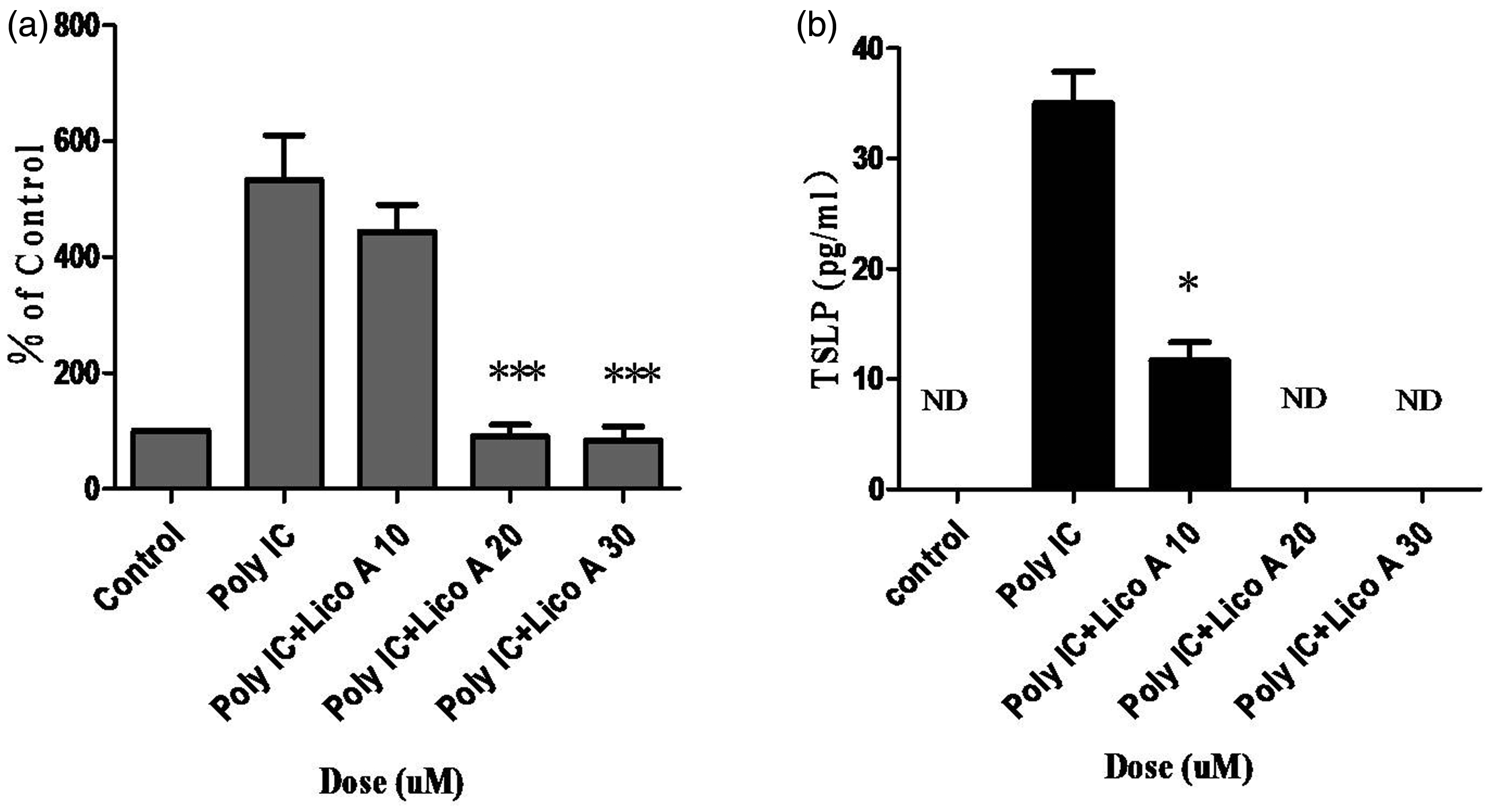
Licochalcone A significantly inhibited poly-IC-induced DNA-binding activity of NF-κB
The expression of human TSLP mRNA is controlled by NF-kB in various cells such as epithelial cells and fibroblasts.12,18 To investigate the inhibitory effect of licochalcone A on NF-κB activation, we first evaluated the effect of licochalcone A on poly-IC-induced NF-kB expression by Western blot assay using nuclear extracts. When BEAS 2B cells were treated with various doses of licochalcone A for 24 h after poly-IC stimulation for 30 min, the expression of both NF-κB p50 and p65 was down-regulated dose dependently (Figure 3a, b). Next, to decide which type of NF-κB subunit was involved in TSLP transcription, we obtained nuclear extracts from BEAS 2B cells treated with or without licochalcone A. We also applied anti-p50 and anti-p65 antibodies after poly-IC stimulation for 30 min and tested them using an oligonucleotide containing the NF-κB-binding motif from the human TSLP gene promoter. As shown in Figure 3c, licochalcone A markedly inhibited poly-IC-induced DNA-binding activity of NF-κB, and addition of antibodies against p65 and p50 each induced a supershift of the DNA-binding complex compared to the control antibody, indicating that licochalcone A down-regulated poly-IC-induced expression and secretion of TSLP by inactivating NF-κB, and that both NF-κB subunits p50 and p65 bound to the human TSLP gene promoter.
Effect of licochalcone A on poly-IC-induced NF-κB translocation and binding activity. Beas 2B cells were incubated with indicated doses of licochalcone A for 24 h after stimulation with poly-IC (5 µg/mL) for 30 min, and nuclear extracts were prepared and Western blotting analysis for NF-κB p50 was performed. Lamin B was used as a loading control (a). Beas 2B cells were incubated with indicated doses of licochalcone A for 24 h after stimulation with poly-IC (5 µg/mL) for 30 min, and nuclear extracts were prepared and Western blotting analysis for NF-κB p65 was performed (b). Beas 2B cells were incubated with 30 µmol/L licochalcone A for 3 h after stimulation with poly-IC (5 µg/mL) for 30 min, and electrophoretic mobility shift assay (EMSA) was performed as described in “Methods” section. The arrow indicates specific binding activity. For supershift assay nuclear extract was preincubated with 1 g of normal IgG, anti-NF-κB p50, or anti-NF-κB p50 for 1 h before the addition of labeled oligonucleotide probe (c). Values represent the means ± SEM. **P < 0.005, ***P < 0.001 vs. poly-IC. ND: not detected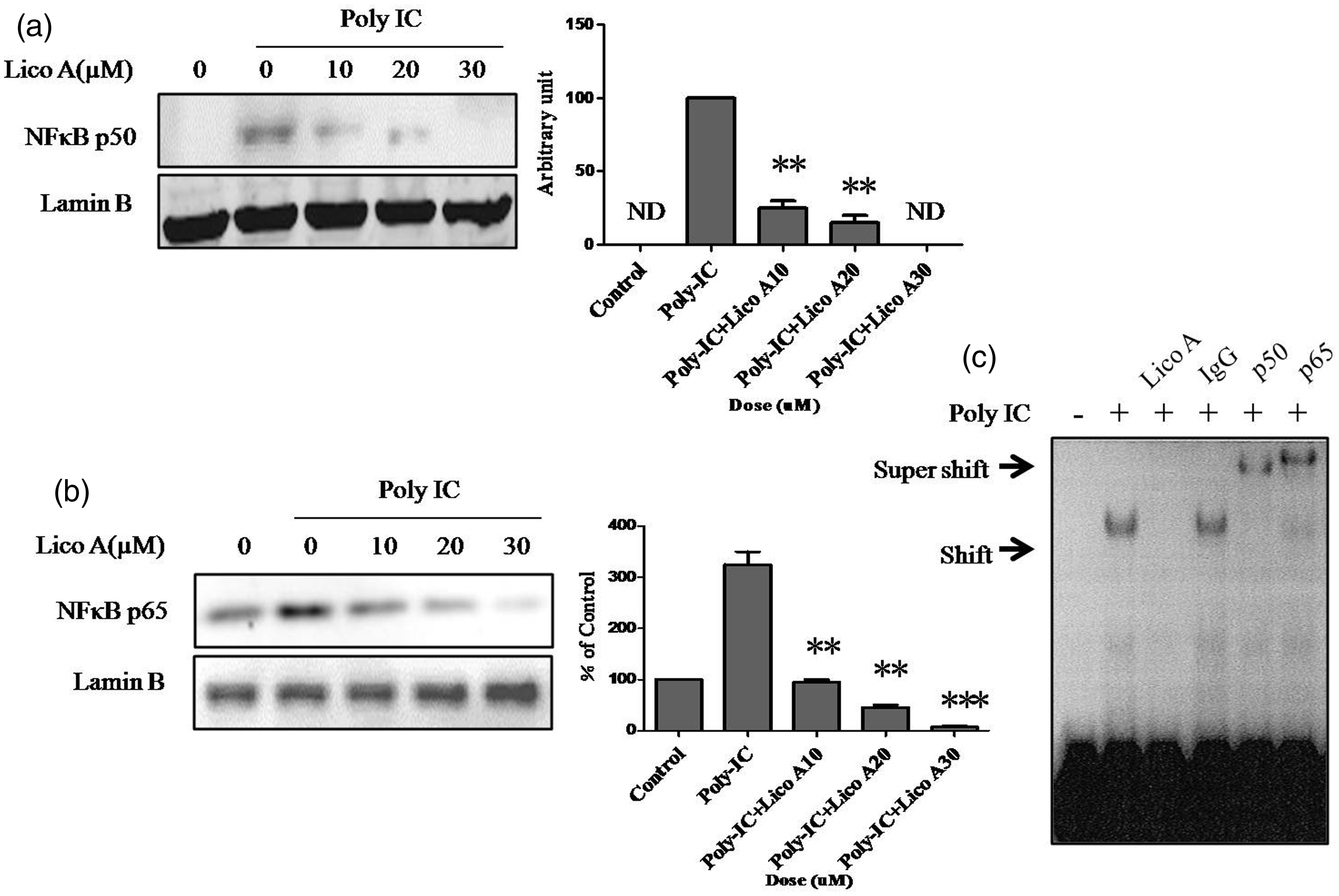
Licochalcone A significantly inhibited poly-IC-induced IκB kinase activation and IκBα degradation
In the canonical NF-κB pathway, stimulants activate IκB kinases (IKKs), which comprise two catalytic subunits, IKKα and IKKβ, and a regulatory subunit, IKKγ. In unstimulated cells, NF-κB remains inactive in the cytoplasm owing to its association with inhibitor proteins of the IκB family. Activated IKKs phosphorylate IκBs, leading to their ubiquitination and proteasomal degradation. These events release NF-κB dimers in the cytosol, allowing them to translocate to the nucleus, where they enhance the transcription of target genes.15,19
To determine how licochalcone A inhibited NF-κB activation induced by poly-IC, we investigated its effect on the expression of IKKα, IKKβ, and IKKγ. The expression levels of IKKα, IKKβ, and IKKγ were unchanged in cells treated with 30 µmol/L licochalcone A in the absence and presence of poly-IC simulation (Figure 4a), indicating that the inhibition of NF-κB activation by licochalcone A was not due to the altered expression of IKK subunits.
Effect of licochalcone A on poly-IC-induced IKK activity. Beas 2B cells were incubated with or without 30 µmol/L licochalcone A for 3 h after stimulation with or without poly-IC (5 µg/mL) for 30 min. A whole cell lysates were immunoblotted with antibodies recognizing IKKα, IKKβ, IKKγ, or β-actin (a). A whole cell lysates were immunoprecipitated with anti-IKKγ antibody. IKK immunoprecipitates were assayed for kinase activity using purified GST-IB as a substrate and immunoprecipitates were immunoblotted with anti-IKKγ antibody (b). A whole cell lysates were immunoblotted with antibodies recognizing IκBα or β-actin as a control (c). Values represent the means ± SEM. *P < 0.05, **P < 0.005 vs. poly-IC
Next, using an in vitro kinase assay, we determined whether licochalcone A inhibited poly-IC-induced IKK activation. Cells were pre-treated with licochalcone A after poly-IC stimulation, and the IKK complex was immunoprecipitated with anti-IKKγ antibody. IKK activity was measured using GST–IκBα as a substrate. As shown in Figure 4b, licochalcone A significantly inhibited poly-IC-induced IKK activation. Furthermore, when the degradation of IκBα after poly-IC stimulation was examined by immunoblotting, licochalcone A strongly inhibited poly-IC-induced IκBα degradation (Figure 4c). These data indicate that licochalcone A inhibited NF-κB activation by suppressing IKK activity.
Licochalcone A and poly-IC had no effect on the phosphorylation of NF-κB p65 at Ser 276
NF-κB p65 has been shown to be phosphorylated at Ser 276 by protein kinase A (PKA), while the phosphorylation at serine 536 (Ser 536) depends on the IKK- or PI3K-dependent pathway.20–23 In addition, a recent study showed that licochalcone A inhibits the LPS-induced signaling pathway by inhibiting NF-κB p65 phosphorylation at Ser 276 without affecting IKK activation or the DNA-binding activity of NF-κB.
16
To determine whether both poly-IC and licochalcone A affected NF-κB p65 phosphorylation at Ser 276, we investigated their effect on the expression level of p65 at Ser 276 and Ser 536. We observed that licochalcone A significantly reduced poly-IC-induced phosphorylation of p65 at Ser 536 (Figure 5 middle). In contrast, poly-IC had no effect on the phosphorylation of p65 at Ser 276 (Figure 5 upper).
Effect of licochalcone A on NF-κB p65 phosphorylation. Beas 2B cells were incubated with or without 30 µmol/L licochalcone A for 3 h after stimulation with or without poly-IC (5 µg/mL) for 30 min, and then a whole cell lysates were immunoblotted with anti-phospho p65 antibody (Ser276), anti-phospho p65 antibody (Ser536) or p65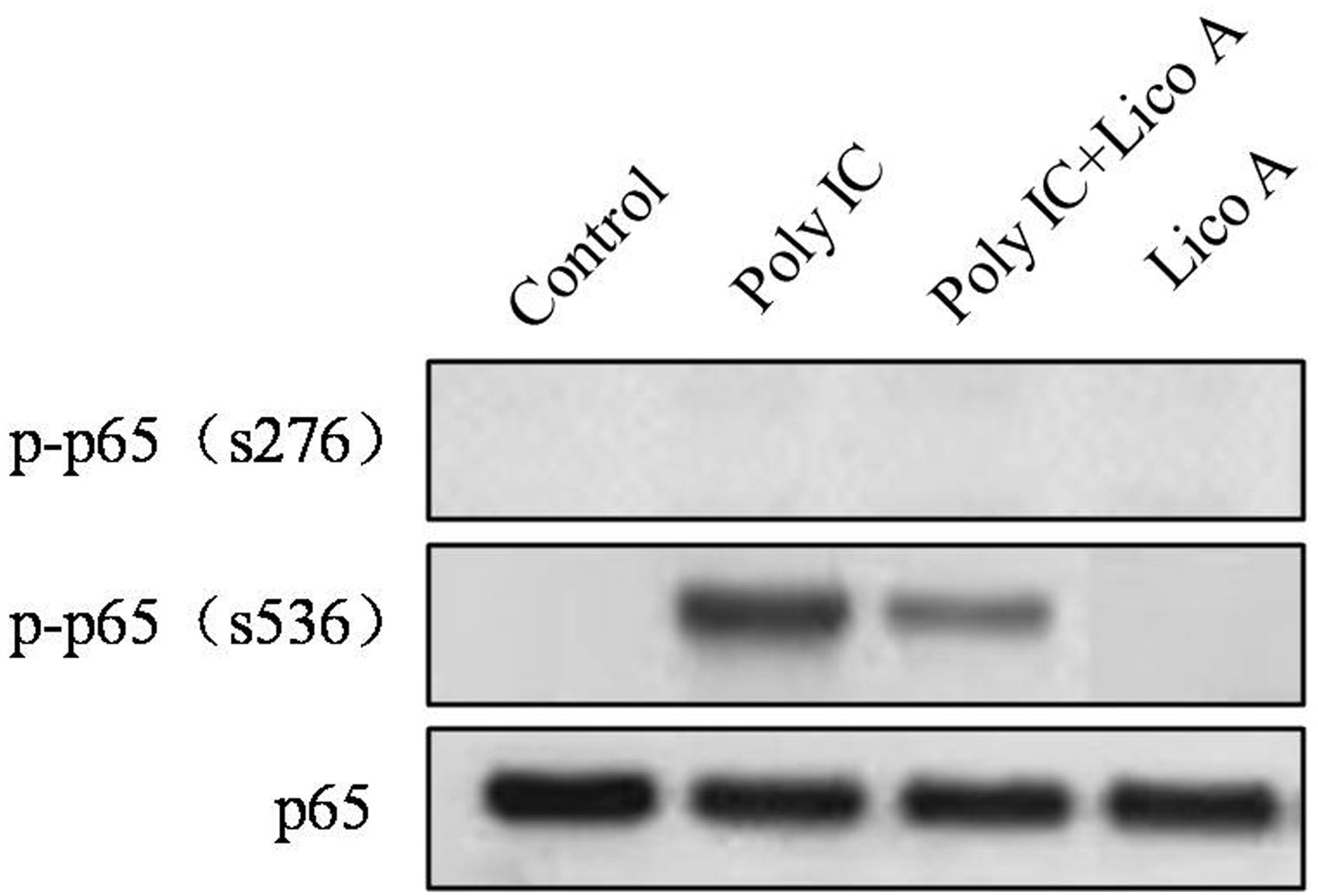
Licochalcone A significantly inhibited poly-IC-induced expression of MCP-1, RANTES, and IL-8
To examine whether licochalcone A also affects the expression of cytokines involved in asthma, its effects on poly-IC-induced expression of MCP-1, RANTES, and IL-8 were investigated by real-time PCR. In BEAS 2B cells, poly-IC significantly induced the expression of MCP-1, RANTES, and IL-8. Interestingly, licochalcone A potently inhibited mRNA expression of MCP-1, RANTES, and IL-8 (Figure 6a, b, and c).
Effect of licochalcone A on poly-IC-induced gene expression of MCP-1, RANTES, and IL-8. Beas 2B cells were incubated with or without 30 µmol/L licochalcone A for 3 h after stimulation with or without poly-IC (5 µg/mL) for 30 min, and then real-time PCR performed for MCP-1 (a), RANTES (b), and IL-8 (c). The values are normalized relative to the GAPDH standard. *P < 0.05, **P < 0.005, ***P < 0.001 vs. poly-IC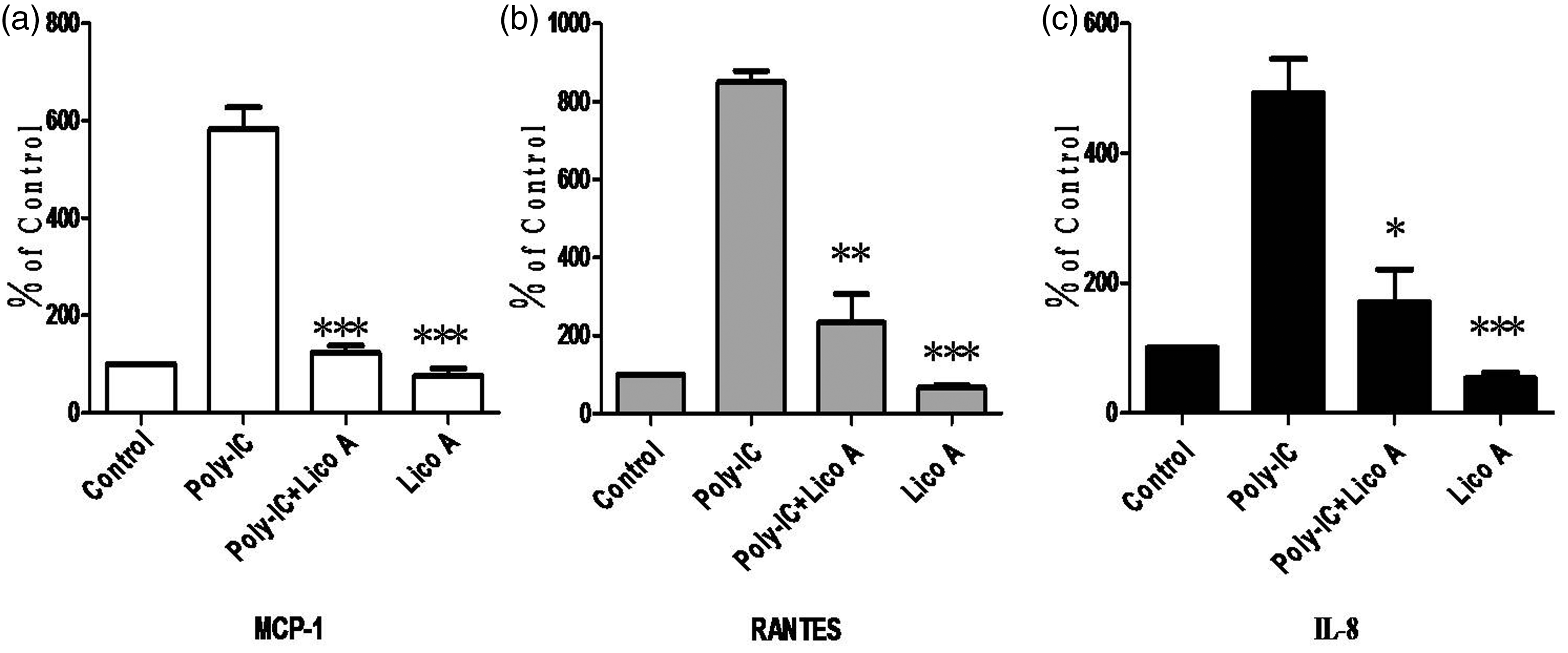
Discussion
Natural product-based medicinal compounds are considered superior to synthetic compounds in certain aspects, such as the absence of severe side effects. 24 The benefits of natural product-based medicinal compounds are evidenced from the use of plant products in traditional systems of medicine. For many years, the root of liquorice, G. glabra and G. inflata, has been used to treat various disorders, including bronchial asthma, gastric ulcer, and inflammation.6,25 Licochalcone A, the major flavonoid isolated from the root of G. inflata, harbors potent anti-inflammatory effects, but the detailed molecular mechanism of such activity has not been explored. Therefore, we investigated the effect of the plant-based compound licochalcone A on the expression and production of TSLP.
As is well known, asthma is a common chronic inflammatory disease characterized by the infiltration and accumulation of memory-like Th2 cells and eosinophils. In the lung, TSLP is expressed primarily by airway epithelial cells, and it has been shown to be necessary and sufficient for the development of airway inflammation in mice. 26 Elevated levels of TSLP mRNA have been found in the airway epithelium of asthma patients. 10 TSLP can potently activate myeloid dendritic cells (DCs) by up-regulating their surface expression of MHC class II, CD54, CD80, CD83, CD86, and DC-lysosomal-associated membrane protein. 27 TSLP-activated DCs can induce naïve T cells to differentiate into inflammatory Th2 cells producing the classical Th2 cytokines IL-4, IL-5, and IL-13, as well as a large amount of TNFα. 28 Moreover, in vitro, only TSLP-activated DCs can induce allergen-specific Th2 memory cells to undergo homeostatic expansion and further Th2 polarization, as well as to mediate recall responses. 29 In this study, we demonstrated that the microbial mimic poly-IC induces TSLP expression in human PBECs and BEAS 2B cells via an IKK/NF-κB-mediated pathway. Both PBECs and BEAS 2B cells have a low basal expression of TSLP, which can be elevated by poly-IC exposure. This increase in expression was seen at both the mRNA and protein levels, suggesting the role of a transcriptional mechanism.
The NF-κB transcription factor is a key player in the transactivation of a network of pro-inflammatory genes, including TSLP, MCP-1, RANTES, and IL-8,12,30,31 making it a potential therapeutic target for limiting the pro-inflammatory response. Airway viral infections are a dominant trigger for worsening asthma. 32 Ample preliminary studies have shown the importance of RV- and RSV-triggered IKK activation, p65 nuclear accumulation, and DNA binding, all of which are characteristic of the classical pathway for NF-κB activation.33–37 NF-κB regulation appears to be stimulus specific, and several recent studies have shown additional regulation by direct p65 phosphorylation at Ser 276 and Ser 536.38,39 Ser 276 is phosphorylated by PKA, while phosphorylation at Ser 536 depends on the IKK- or PI3K-dependent pathway.20–23,40 In this study, we demonstrated, for the first time, the inhibitory effect of licochalcone A on poly-IC-induced TSLP expression and production by the inhibition of IKK activation and subsequent NF-κB activation. Further research is needed to confirm the inhibitory effect of licochalcone A on RV- or RSV-infected bronchial epithelial cells.
Our overall findings underscore the potential of licochalcone A in the treatment of inflammatory diseases such as asthma through the suppression of the IKK/NF-κB/TSLP pathway.
Footnotes
Authors’ contributions
Acquisition of data: YQ; analysis and interpretation of data: SHK, JGX and XY; drafting of manuscript: SHK.
ACKNOWLEDGMENTS
This work was supported in part by the Tianjin Natural Science Foundation (Grant No 12JCYBJC33300) and in part by Science and Technology Fund of Tianjin Public Health Bureau (Grant No 2012KY08).