
Abstract
Cup-shaped secretory portals at the cell plasma membrane called porosomes mediate the precision release of intravesicular material from cells. Membrane-bound secretory vesicles transiently dock and fuse at the base of porosomes facing the cytosol to expel pressurized intravesicular contents from the cell during secretion. The structure, isolation, composition, and functional reconstitution of the neuronal porosome complex have greatly progressed, providing a molecular understanding of its function in health and disease. Neuronal porosomes are 15 nm cup-shaped lipoprotein structures composed of nearly 40 proteins, compared to the 120 nm nuclear pore complex composed of >500 protein molecules. Membrane proteins compose the porosome complex, making it practically impossible to solve its atomic structure. However, atomic force microscopy and small-angle X-ray solution scattering studies have provided three-dimensional structural details of the native neuronal porosome at sub-nanometer resolution, providing insights into the molecular mechanism of its function. The participation of several porosome proteins previously implicated in neurotransmission and neurological disorders, further attest to the crosstalk between porosome proteins and their coordinated involvement in release of neurotransmitter at the synapse.
Introduction
A variety of physiological processes in cells of the bakers yeast to cells in humans are governed by the release of intracellular products from within cells to the outside. Cell–cell communication such as neurotransmission, the release of histamines from mast cells following exposure to allergens, or the release of hormones from endocrine cells to maintain homeostasis, all result from the release of membrane-bound vesicle encased products to the outside either via complete collapse of the vesicle membrane at the cell plasma membrane termed total fusion, or the transient engagement and fusion of secretory vesicles at the cell plasma membrane termed kiss-and-run. This later process of kiss-and-run, enabling the release of a portion of intravesicular contents to the outside, while retaining the chemical and morphological integrity of the secretory vesicle following a secretory episode, supports the observation in electron micrographs of partially empty vesicles following cell secretion. However, since the high surface tension of secretory vesicles would favor its collapse at the cell plasma membrane, the presence of a permanent docking and transient fusion port at the cell plasma membrane was hypothesized and discovered by our laboratory over 20 years ago, and named the ‘porosome’. Studies now demonstrate that the ‘fusion pore’, an opening between the membrane of the vesicle and the plasma membrane is established at the porosome base oriented toward the interior of the cell.
Historically, the kiss-and-run or transient mode of docking and fusion of secretory vesicles at the cell plasma membrane were proposed in 1973, 1 that could explain the fractional release of intravesicular contents from cells during cell secretion. How this transient mechanism could be accomplished, remained a mystery. Subsequently in 1990, it was suggested that the fusion pore results from a ‘preassembled ion channel-like structure that could open and close.’ 2 In a review 3 from the early 1990s, the authors had proposed that the primary difficulty in observing such preassembled structures at the cell plasma membrane and fusion pore formation at such a structure was due to the absence of tools for ultrahigh resolution live cell imaging. The invention of force microscopy in the mid-1980s changed all that. In the mid-1990s, the hypothesis of the presence of secretory portals was confirmed by the discovery of the porosome, a new cellular structure at the cell plasma membrane 4 using the atomic force microscope (AFM), 5 a imaging tool that has revolutionized nano science especially nano cell biology. Using AFM, 100–180 nm in size secretory portals were observed at the plasma membrane of live pancreatic acinar cells 4 that secreted digestive enzymes through them. The presence and morphology of these secretory portals were further confirmed using electron microscopy (EM),6,7 and their presence was established in all secretory cells examined including neurons,7–20 initially by our group7–20 and subsequently by other laboratories.21–30 The function of the porosome as a universal secretory portal, its chemical composition,7–9,13,19,20 and its functional and structural reconstitution,6,8 has greatly advanced our understanding of this vital cellular organelle. Furthermore in neurons, the native porosome with docked synaptic vesicle at the presynaptic membrane has been determined at sub-nanometer resolution using small-angle X-Ray solution scattering (SAXS), 12 providing a molecular understanding of its structure–function. Consequently in the past 18 years, several articles report on the porosome and the importance of its discovery in contributing to our understanding of transient or the kiss-and-rum mechanism of cell secretion.31–47
Careful examination of the porosome proteome demonstrates the participation of several porosome-associated proteins previously implicated either in neurotransmitter release or in various neurological disorders,48–98 further confirming the involvement of porosome-associated proteins in crosstalk within the complex and in the regulation of neurotransmitter release. Therefore, certain porosome proteins could be potential drug targets to help ameliorating various neurosecretory disorders. In this brief review focused on the neuronal porosome complex, the structure, composition, reconstitution, and the involvement of porosome proteins on neurotransmitter release, and in various neurological disorders is presented.
Structure, function, and isolation of the neuronal porosome
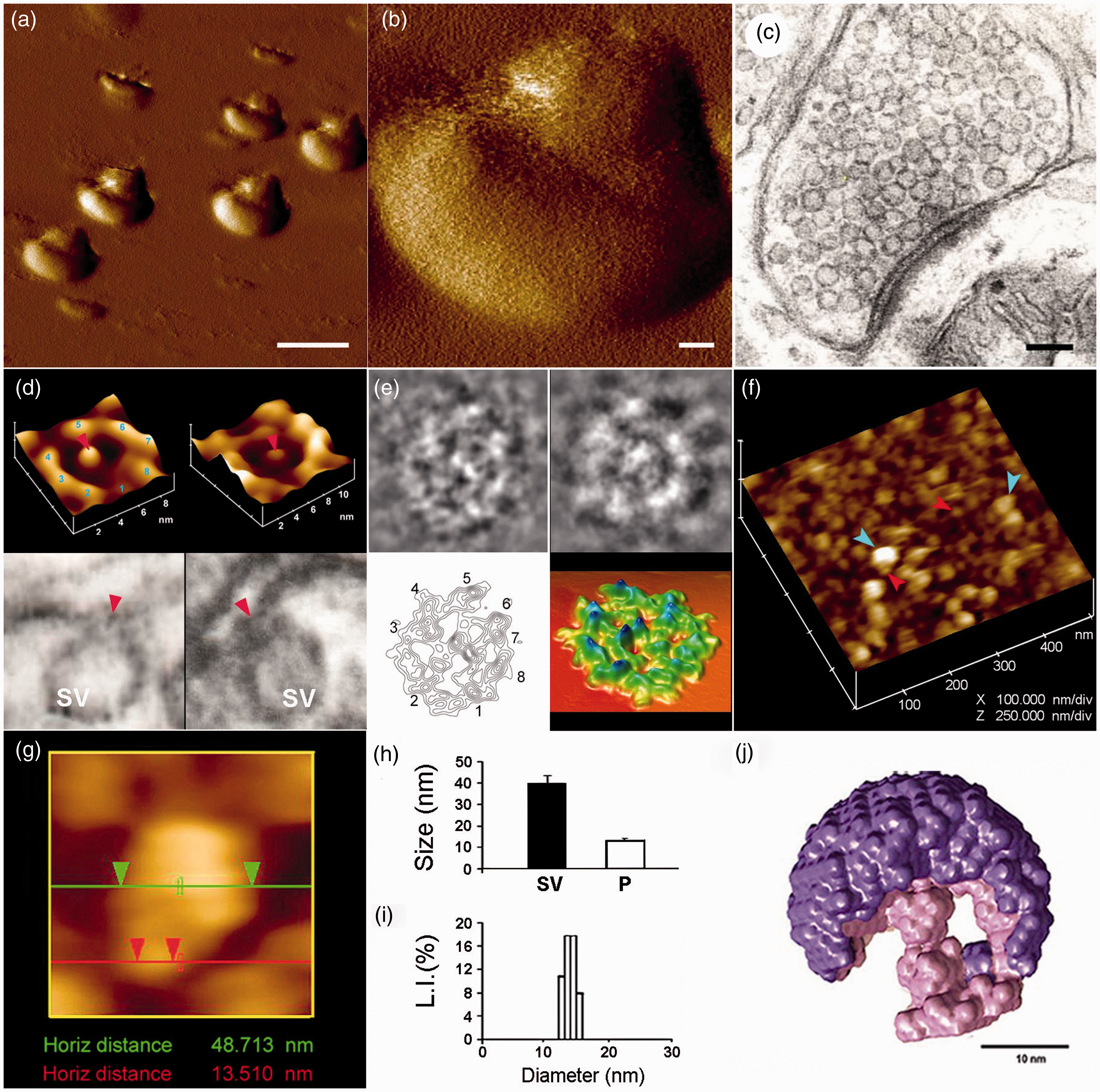
Using AFM,
8
EM,
8
and SAXS,
12
the ultrahighresolution imaging of the presynaptic membrane of the neuronal synapse, reveal the presence of 15 nm cup-shaped lipoprotein porosomes, each having a rod-like structure at its center (Figure 1). The outer peremiter of the porosome structure facing the postsynaptic membrane is marked by eight protein densities equally spaced around the perimeter of the complex (Figure 1(d) and (e)). The eight-fold symmetry of the protein densities has been previously identified in native neuronal porosomes (Figure (d) top left) and in immunoisolated porosome preparations reconstituted into artificial lipid membrane (Figure (d) top right). As observed in AFM micrographs of the porosome complex, eight interconnected protein densities are identified in purified neuronal porosomes as shown via EM (Figure 1(e)). Electron density and contour mapping with ensuing three-dimensional topology profile of the porosome reveal the arrangement of proteins within the complex. The rod-like structure or plug at the center of the porosome is connected via distinct spoke-like elements to the eight peripheral densities (Figure 1(e) lower left). The three-dimensional topology of the neuronal porosome determined by electron density maps, further attest to a semi-circular porosome morphology, containing a central rod connected via spokes to the densities present at the outer margin of the porosome opening (Figure 1(e) lower right]. AFM measurements on the cytosolic side of isolated synaptosomal membrane demonstrate the presence of 12–17 nm inverted porosomes, some with 50 nm synaptic vesicles attached to the base of the complex (Figure 1(f) and (g)). AFM, EM, and photon correlation spectroscopy (PCS) (Figure 1(h) and (i)), collectively show neuronal porosomes average 15 nm in size ranging from 12 to 17 nm. SAXS studies
12
of the native porosome complex within the synaptic terminal found the structure associated with a synaptic vesicle (Figure 1(j)), providing further details of the neuronal porosome architecture in its native state in the cell. AFM micrographs of synaptosomal membrane demonstrate the porosome central plug to be positioned in distinct conformational states, extended outward, pushed inward, or completely withdrawn into the structure (Figure 2). These AFM studies suggest the central plug of the porosome complex to be capable of linear movement, perhaps facilitating the necessary rapid formation and closure of the fusion pore (Figure 3). Furthermore, the structural details of the neuronal porosome complex associated with synaptic vesicle from SAXS studies
12
provides information on the possible involvement of the central plug in the establishment of the fusion pore at the porosome base. SAXS studies lend support to the fact that the synaptic vesicle is required to be closely appose to the porosome base and to establish the fusion pore (Figure 3). The SAXS structure of the porosome-synaptic vesicle complex further supports the hypothesis that following stimulation of neurotransmission, the central plug is pushed inward, resulting in a small 3–4 nm protrusion at the porosome base and the tightly apposed synaptic vesicle membrane. Consequently, the protrusion or bump established in both the opposed membranes are under enormous tension, facilitating SNARE-mediated and calcium-induced membrane fusion. The resultant arrangement of t-/v-SNARE complex in a rosette conformation99–109 (Figure 4) between the porosome base and the docked synaptic vesicle membrane, and the consequent continuity or fusion pore establishment between the opposing bilayers results in neurotransmitter release. The central plug is then pushed outward to relieve tension of the opposing membranes, allowing swift closure of the transiently established fusion pore. The presence of actin, myosin, and other structural and motor proteins in the neuronal porosome complex
13
(Tables 1 and 2) may aid in the motility of the central plug and possible utilization of ATP and calcium for its movement.
Neuronal porosome structure and organization.8–13 (a) Low- (Scale = 1 µm) (b) and high-resolution (Scale = 100 nm) atomic force microscope (AFM) micrographs of isolated rat-brain nerve terminals or synaptosomes in buffer. (c) Electron microscope (EM) picture of a bouton (Scale = 100 nm). (d) Native neuronal porosome complex at the presynaptic membrane (Figure d top left), and of an isolated porosome complex reconstituted into lipid membrane (Figure d top right). Note the native and reconstituted porosomes are morphologically identical. Lower panels are two EM micrographs demonstrating synaptic vesicles (SV) docked at the base of cup-shaped porosome, with a central plug (red arrowhead). (e) EM, electron density and 3D contour maps provide at nanometer resolution of protein arrangement in the porosome. (f) The cytosolic compartment of an isolated bouton or synaptosome demonstrating SV (blue arrow-head) docked at the base of porosomes (red arrow-head). (g) an SV docked at a porosome. (h) Measurements (n = 15) using AFM of SV (SV, 40.15 ± 3.14) and porosomes (P, 13.05 ± 0.91) at the presynaptic membrane. (i) Photon correlation spectroscopy performed on isolated neuronal porosomes measure 12–17 nm. (j) X-ray solution scattering (SAXS) of averaged 3-D structure of a SV (purple) docked at the base of a native neuronal porosome (pink).
19
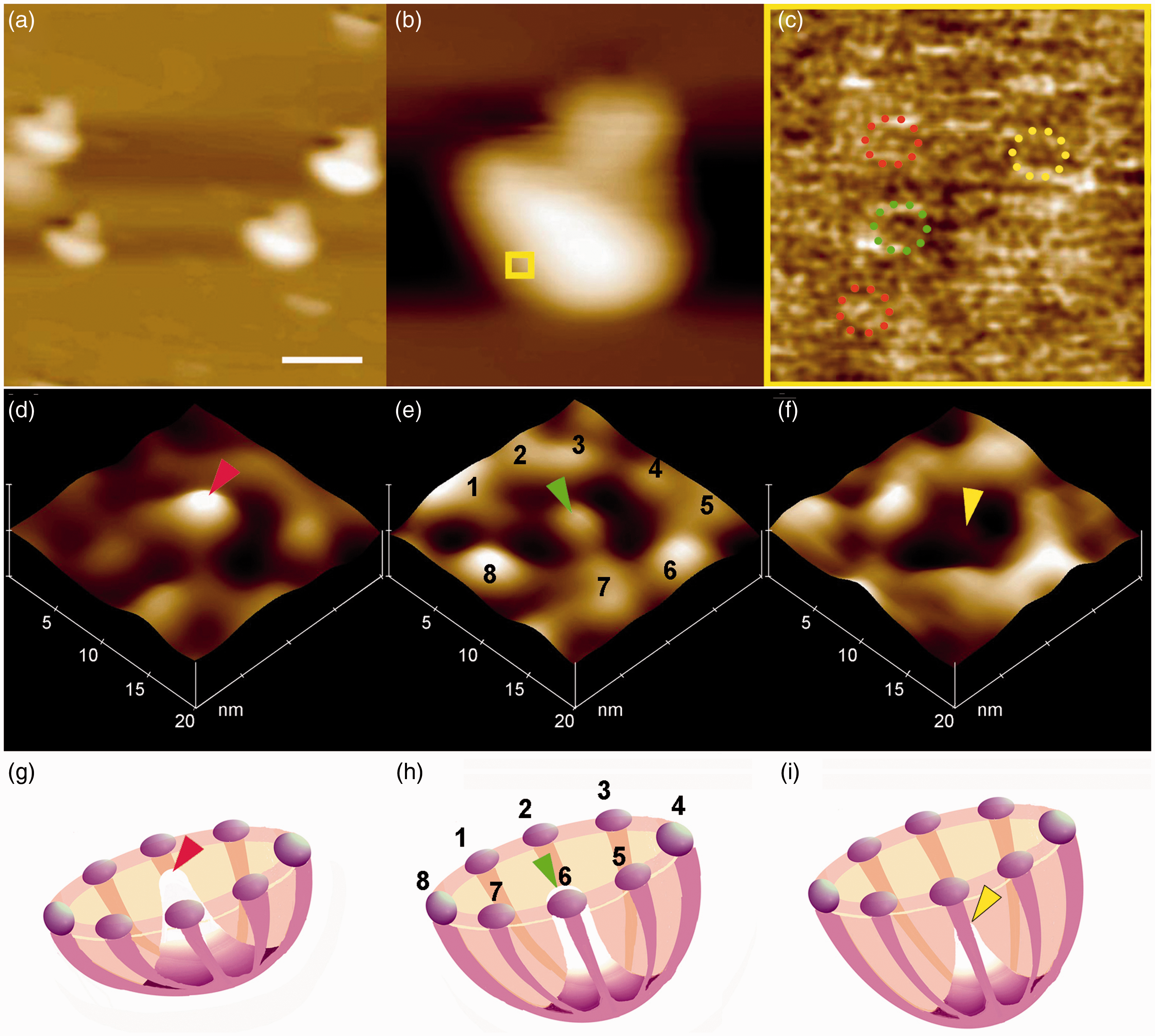
SV–porosome complex using EM, AFM and SAXS demonstrate similarity morphology.8–13 (A color version of this figure is available in the online journal.) The neuronal porosome exhibiting the central plug at various conformational states.
11
(a) Low magnification (Bar = 1 µm) (b) and high magnification AFM micrograph Box = 100 nm2 of isolated synaptic terminals in physiological buffer. (c) The boxed area in (b) is presented at higher magnification with the rod-like density at the center showing fully extended out (Figure c red rings; d, g) suggesting that the porosome opening is in its close conformation. The centrally located rod when mid-way extended (Figure c green ring; e, h) represents a semi-open state and when fully retracted (Figure c yellow ring; f, 2) represents an open state.
11
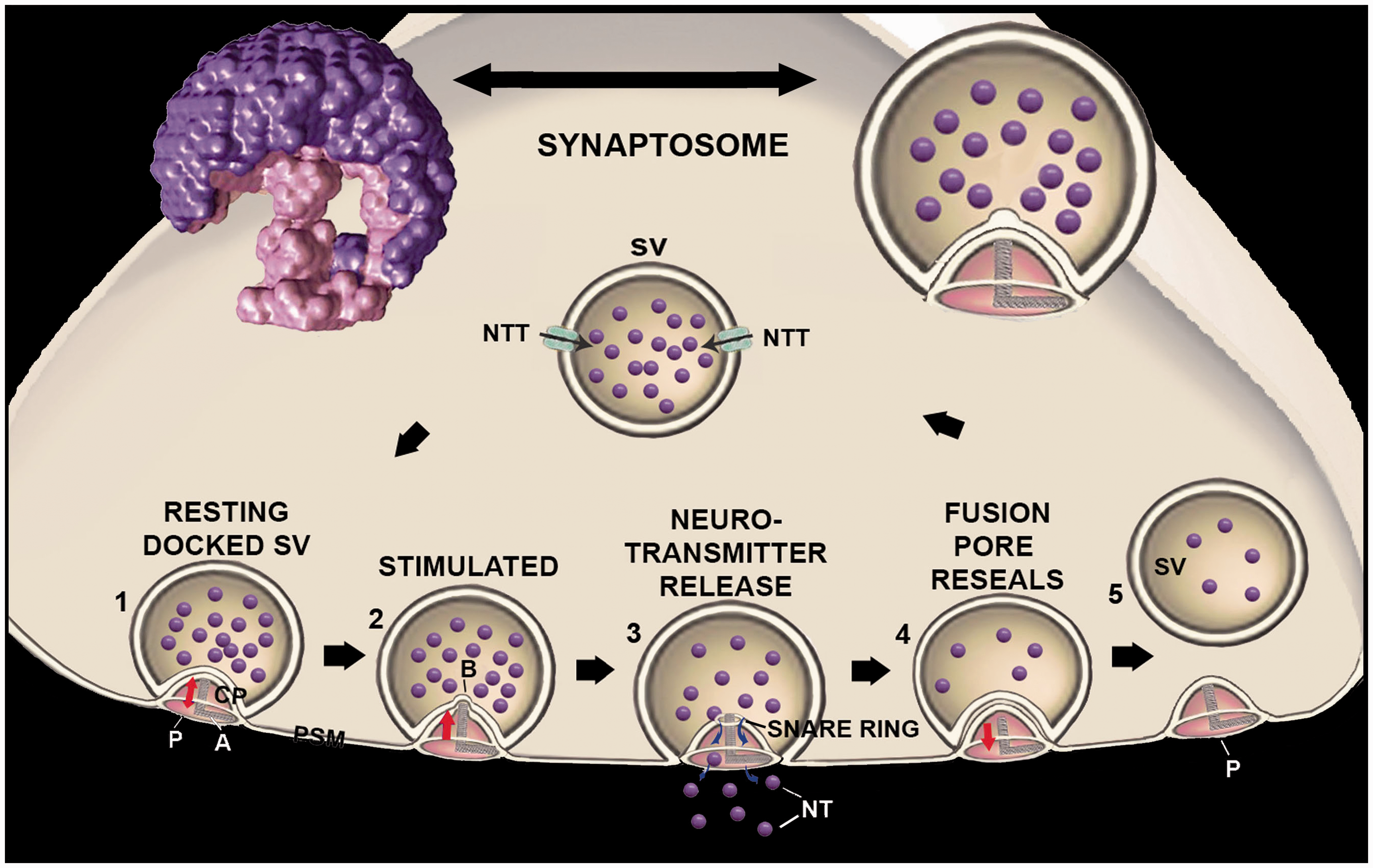
(A color version of this figure is available in the online journal.) Central plug’s involvement in neurotransmission.
12
To the top left and right are images of the native three dimensional X-ray solution scattering structure of a synaptic vesicle (SV, purple)-porosome (P, pink) complex, and a schematic drawing of such a complex. The sequence of events that may occur at the nerve terminal involving SV-P interactions and at the presynaptic membrane (PSM) and neurotransmitter release is presented.1–5 A cantilever arm (A) attached to the central plug (CP), provides vertical motion of the structure.
1
Following stimulation of neurotransmission, the foot of the CP within the porosome complex is pushed inward resulting in the establishment of a small bump (b) whose radius corresponds to the base of the CP around 1.5–2 nm. As a result, the membrane at the base of the porosome is under great stress resulting in the establishment of SNARE ring complex,
3
and membrane fusion and fusion pore (FP) formation leading to neurotransmission. Retraction of the porosome plug outward toward the opening of the structure, results in resealing the FP due in part to release of membrane tension.
4
Partially empty SV can dissociate from the porosome and get refilled through the neurotransmitter transporters (NTT) available at the membrane of the SV.
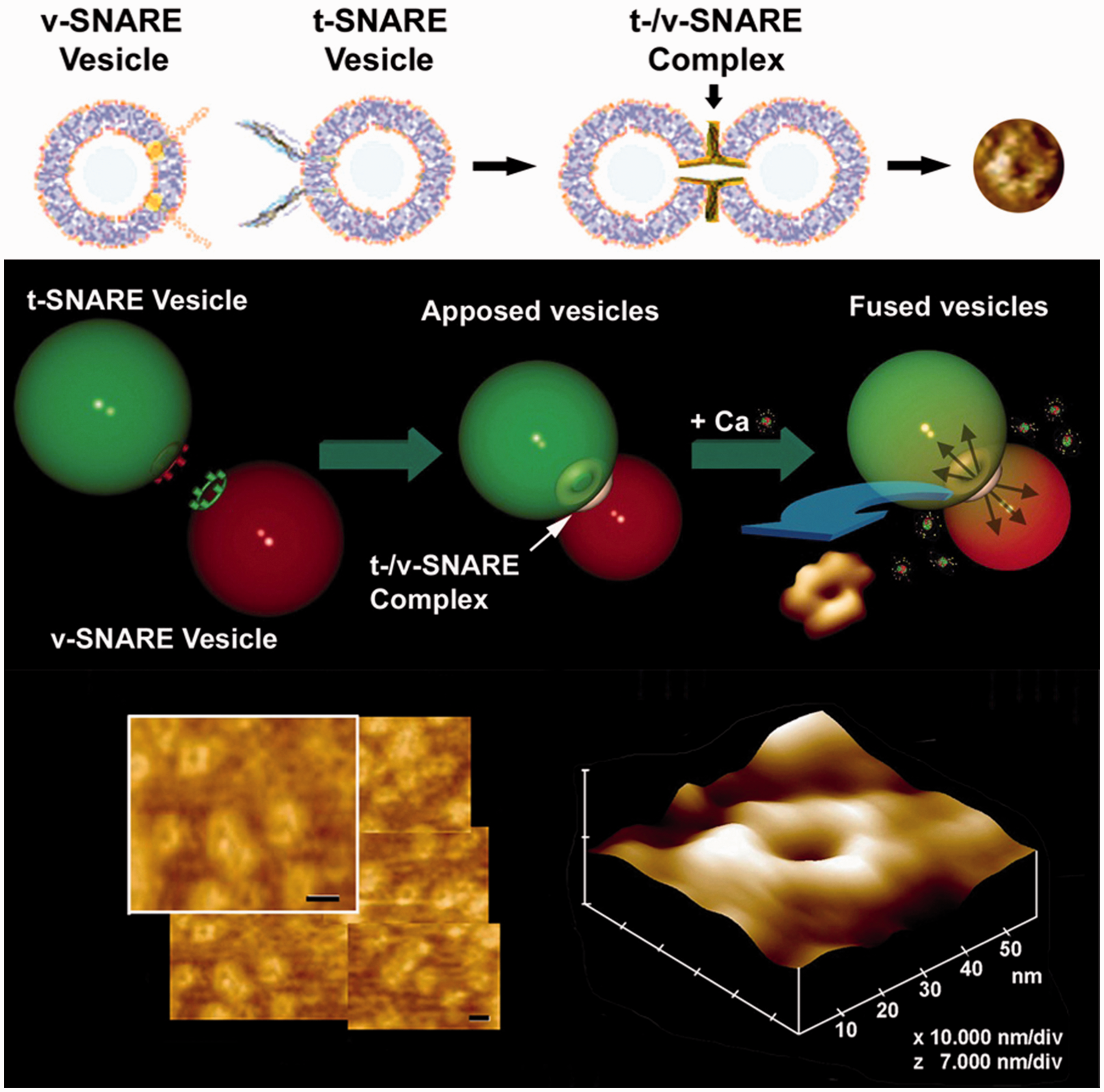
5
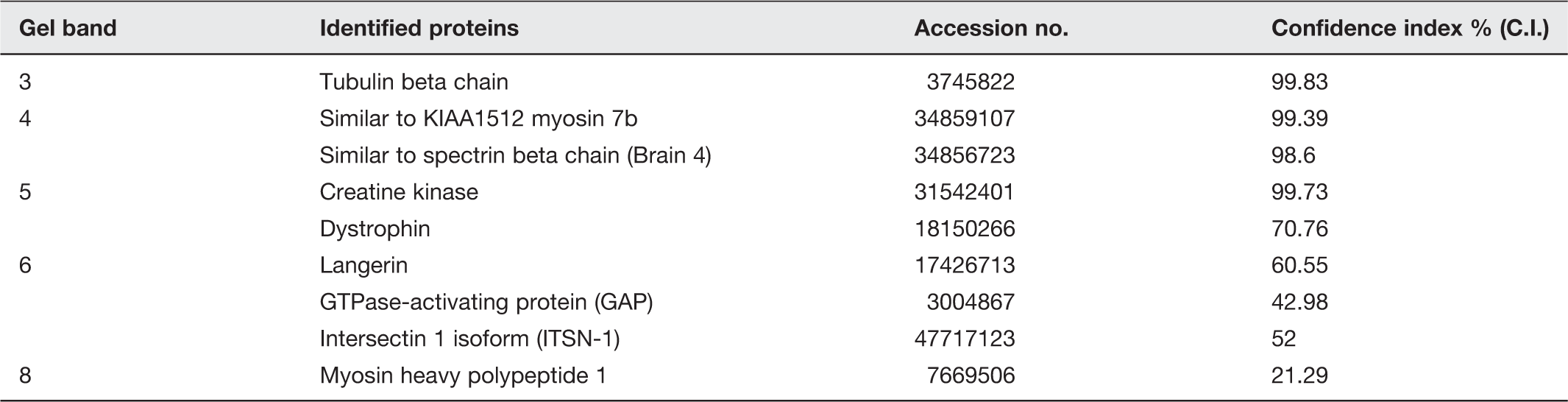
The refilled SV is then ready for reuse in the next round of neurotransmission. (A color version of this figure is available in the online journal.) Interaction of t-SNARE-associated lipid vesicle or lipid membrane with v-SNARE-associated liposomes establishes SNARE ring complexes. T-/v-SNARE ring complexes or rosettes established following interactions between t-SNAREs and v-SNAREs reconstituted lipid membranes.99–106 Top and center panels are interaction between t-SNARE liposomes with v-SNARE liposome in opposing vesicles. Atomic force micrographs of actual SNARE rosettes formed whose size is proportional to the vesicle size hence vesicle curvature. Besides vesicle size, vesicle composition is also known to influences the size of SNARE rosette. (A color version of this figure is available in the online journal.) MALDI-TOF/TOF results on specific bands in SDS-PAGE-resolved SNAP-25 immunoisolated neuronal porosome complex obtained using 1% Triton-Lubrol-solubilized rat brain synaptosome preparation.
13
Purified rat brain porosomes from two separate experiments were analyzed by LC-MS/MS on both LTQ and QSTAR XL. Only proteins identified in both samples are reported here all of which had protein confidence ≥95% with at least two unique peptides each having 95% confidence or above. Proteins also found in earlier immunoisolation studies are marked in a separate column. indicates proteins identified using MALDI-TOF/TOF. proteins identified using immunoblot analysis.
13
Chemistry of the porosome complex in neurons
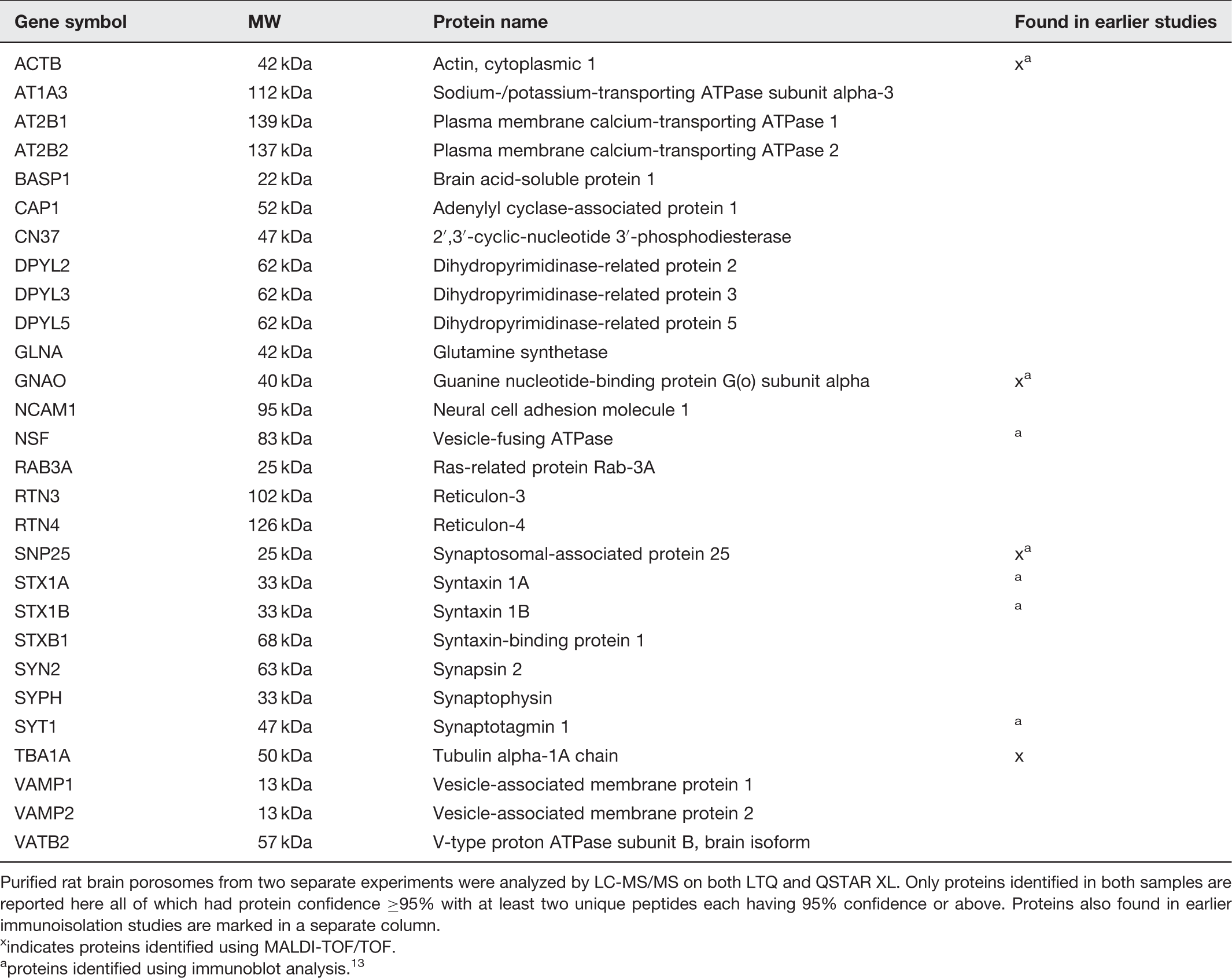
Chemistry of purified neuronal porosomes has been determined by mass spectrometry revealing the presence of approximately 40 proteins in the complex. 13 Utilizing information from earlier studies, the intermolecular interactions with other proteins and within the neuronal porosome proteome have been predicted using the STRING 9.0 database of protein–protein interactions. 110 Additionally, the dynamic nature of the porosome is reflected from association and dissociation of proteins from the complex during neurotransmission. 13 Immunoisolated porosomes from rat brain synaptosome preparations demonstrate the presence of a number proteins, among them P/Q-type calcium channel, actin, vimentin, the N-ethylmaleimide-sensitive factor (NSF), SNAP-25, syntaxin-1, synaptotagmin-1, alpha subunit of the heterotrimeric GTP-binding Gαo and the chloride channel CLC-3.8,9,13 A number of these porosome proteins have previously been implicated in cell secretion.6,7,111–114 Neuronal porosome-associated proteins previously implicated either in neurotransmitter release or in various neurological disorders48–98 have been further discussed in this article. Immunoisolated porosomes from neuronal tissue resolved using sodium dodecyl sulfate polyacrylamide gel electrophoresis, followed by SYPRO staining of the resolved protein bands in the gel, demonstrate the presence of 15 bands ranging from >205 kDa to >6.5 kDa in molecular weight, and show the presence of nine additional proteins besides those determined using mass spectrometry (Table 1). 13 GTPase activating protein (GAP), tubulin, myosin 7b, spectrin beta chain, creatine kinase, dystrophin, langerin, intersectin 1, and myosin heavy chain 1, have also been identified (Table 1). Furthermore, dynamin was hypothesized to be involved with the porosome complex based on its known association with intersectin 115 and its presence was confirmed by Western blot analysis. 13 The dynamics of dynamin association–dissociation at the neuronal porosome complex has also been demonstrated, since dynamin is increased in porosomes isolated from brain slices following stimulation. 13 In contrast to the increase in dynamin association with the porosome, a dissociation of Gαo is observed following stimulation. 13
In addition to determining the proteome of the immunoisolated rat brain porosomes, the porosome has also been purified using gel filtration chromatography and analyzed by LC-MS/MS on both LTQ and QSTAR XL. Analysis of the isolated porosome using gel filtration, revealed the presence of a number of new proteins, in addition to those obtained using immuno pull down (Table 2).
13
Plasma membrane calcium-transporting ATPase 1 and 2 are both identified in the neuronal porosome and are known to be responsible for ATP-dependant removal of calcium from the cell, a process necessary for maintaining the delicate calcium balance required for exocytotic processes. Similarly, sodium/potassium-transporting ATPase is identified in the neuronal porosome, allowing for the required electrochemical gradient for maintaining the membrane resting potential, in addition to regulating cell volume during neurotransmission. Mass spectrometry on isolated neuronal porosome also demonstrates the presence of 2′,3′-cyclic-nucleotide 3′-phosphodiesterase, which has been shown to associate with microtubules, function similar to microtubule-associated protein, link tubulin to the cell membrane, and is involved in regulating microtubule distribution within the cell.
116
It is hypothesized that this enzyme may be utilized to maintain the structural integrity of the porosome. Similarly, dihydropyrimidinase-related protein is present in the porosome complex, and is known to participate in the reconstruction of the cytoskeleton and in maintaining polarity of the cell.
117
NSF and dynamin in the porosome complex are essential for SNARE complex disassembly, allowing closure of the fusion pore and dissociation of the synaptic vesicle from the base of the porosome cup. Such a mechanism is required for the disengagement of the synaptic vesicle from the porosome base following completion of neurotransmitter release. Since SNAREs and calcium channels are present at the porosome base,6,102 and known to physically interact,
118
their coordinated involvement in the establishment of the fusion pore at the porosome base during neurotransmission is suggested. The central plug similarly would be composed of myosin, actin, and associated proteins required for its vertical movement to establish the fusion pore at the base of the porosome complex. In order to further advance our understanding of the porosome complex, and the possible association of other minor proteins within the complex, STRING 9.0 database search of known protein–protein interactions (Figure 5)
110
has been implemented. Over five million proteins from 1133 organisms are covered by STRING, which integrates protein interaction data to develop a likely model of protein–protein interactions. STRING maps generated on the neuronal porosome proteome identifies two clusters of protein–protein interactions (Figure 5),
13
one cluster representing cytoskeletal and signaling proteins, while the other cluster contains proteins involved in membrane fusion such as SNAREs, their associated regulatory proteins, and calcium channels. Although the predicted functional interactions using STRING represents >99% confidence, further chemical cross-linking studies followed by mass spectrometry, and structural studies using electron crystallography, SAXS, neutron scattering, and molecular dynamic simulations will provide a molecular arrangement model of the neuronal porosome complex.
Protein–protein interactions within the neuronal porosome complex using STRING database search.
13
Two clusters of protein–protein interactions are identifiable within the porosome complex. The cluster to the left likely represents the open end of the porosome facing outside, and is composed of cytoskeletal and signaling proteins. The other cluster includes membrane fusion proteins like SNAREs and calcium channels. The right cluster would therefore represent the porosome basal. (A color version of this figure is available in the online journal.)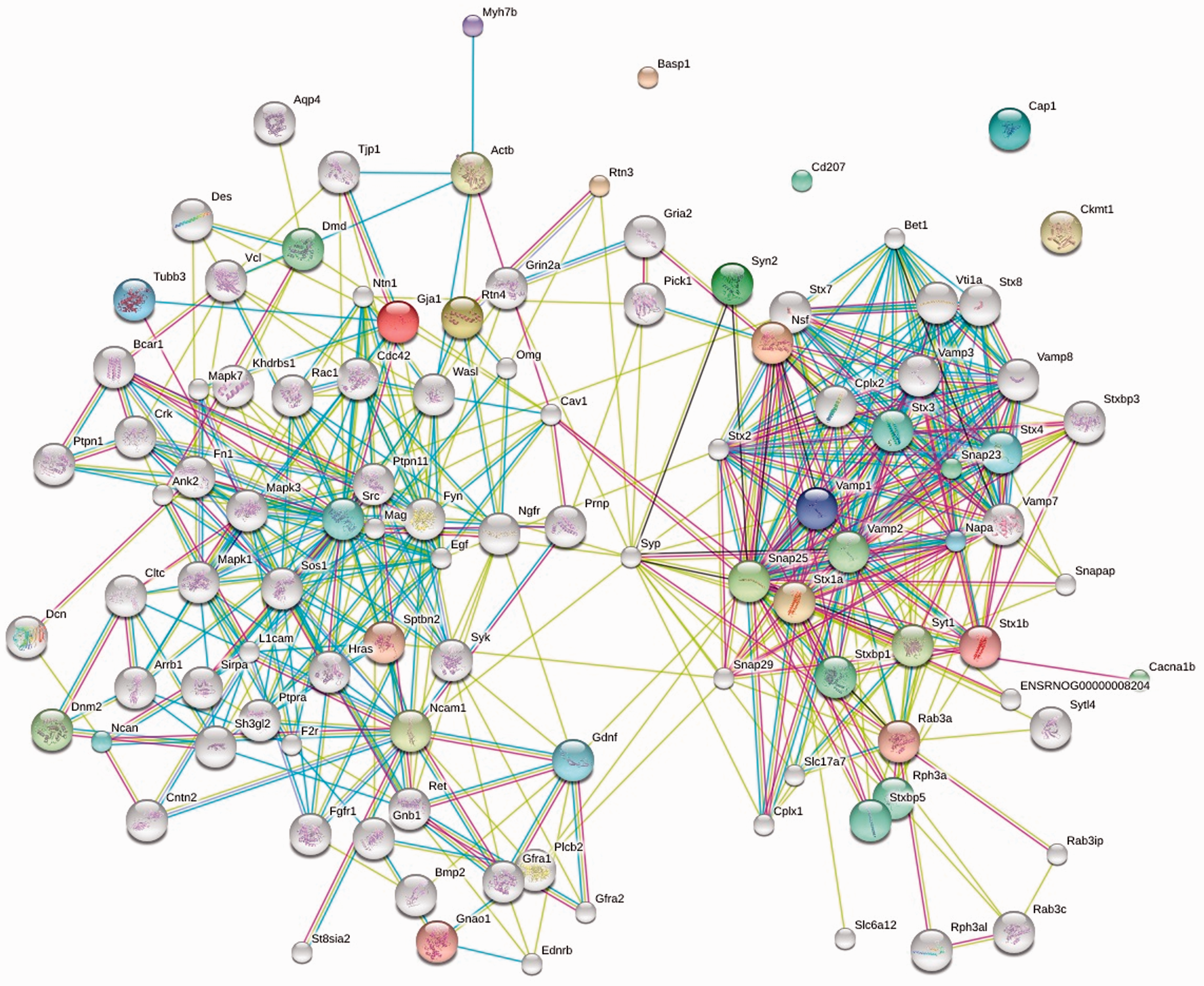
Major lipid species associated with the neuronal porosome complex identified using mass spectrometry
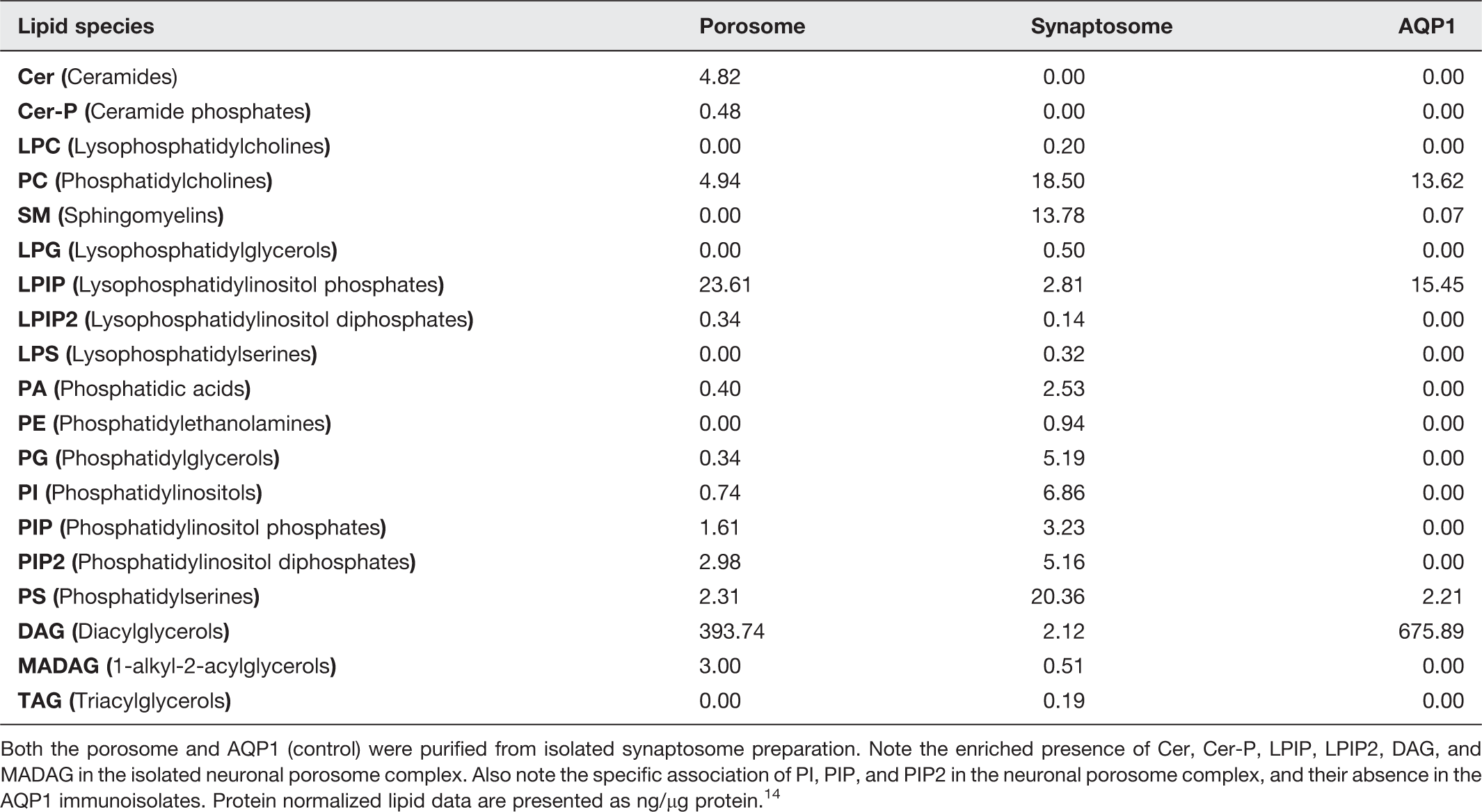
Both the porosome and AQP1 (control) were purified from isolated synaptosome preparation. Note the enriched presence of Cer, Cer-P, LPIP, LPIP2, DAG, and MADAG in the isolated neuronal porosome complex. Also note the specific association of PI, PIP, and PIP2 in the neuronal porosome complex, and their absence in the AQP1 immunoisolates. Protein normalized lipid data are presented as ng/µg protein. 14
List of neuronal porosome proteins containing various lipid-binding domains. 14

To be able to understand protein–lipid interactions within the porosome complex known protein–lipid-binding domains within the neuronal porosome proteome have been analyzed (Table 4). 14 This study, besides shedding light on the interactions between porosome proteins and the identified lipids in the complex, further suggests the interactions of other lipids that may interact with lipid-binding domains of porosome proteins. The various known lipid-binding domains that have been considered in understanding protein–lipid interactions within the neuronal porosome complex include the Cystein-rich C1 domain, the calcium-dependent lipid binding C2 domain, the PH or pleckstrin homology domain, the FYVE zinc-binding domain, the 120 amino acid phox or PX homology domain, the ENTH or Epsin N-terminal homology domain, the AP180 N-terminal or ANTH homology domain, the BAR or Bin-Amphiphysin-Rvs domain, the FERM or the Babd 4.1-Ezrin-Radixin–Moesin domain, and the Tubby domains. The porosome protein synaptotagmin possess the lipid-binding C2 domain capable of binding a wide range of lipids including phosphatidylcholine (PC) and phosphatidylserine (PS). The neuronal porosome protein spectrin beta possesses a pleckstrin-binding domain and plays a significant role in membrane anchoring and signaling by binding actin, dystrophin, and PI-(4,5)-P2-; PI-(3,4,5)-P3-; PI-(3,5)-P2-; and Ins(1,4,5)P3. Similarly dystrophin, a porosome protein that contributes to resistance in contraction–relaxation cycles contains 24 spectrin-like repeats and binding domains specific for PI-(4,5)-P2-; PI-(3,4,5)-P3-; PI-(3,5)-P2-; and Ins(1,4,5)P3. The influence of membrane lipids on ion channels such as calcium channel is well known. Studies also demonstrate that slight changes in lipid membrane composition may alter lipid domain formation, and thus modify the activity of calcium channels. Furthermore, lipid-G-protein-coupled receptor x-ray crystallography demonstrates that the signaling lysosphingolipid sphingosine 1-phosphate is able to modulate lymphocyte trafficking, endothelial development and integrity, heart rate, and vascular tone by activating GTP-binding G-protein-coupled receptors. Understanding of the direct or indirect lipid–protein interactions within the neuronal porosome complex has greatly progressed our understanding of the structure–function of several neuronal porosome proteins, and of the porosome complex.
It is well documented that membrane proteins are surrounded by an annulus of lipid molecules that associate with the membrane-penetrating surface of the protein; similarly, non-annular lipid molecules in the membrane bilayer are known to bind between the transmembrane α-helices of membrane proteins. It has been demonstrated that non-annular lipids and cholesterol can associate with membrane proteins outside of transmembrane regions. In contrast, protein association of annular lipids requires fatty acyl chain of specific lengths suggesting hydrophobic matching between membrane proteins and the lipid bilayer. Furthermore, it has been reported that although annular lipids are highly mobile and undergo diffusion with the remainder of the lipid bilayer, non-annular lipids tend to remain associated with the protein, and therefore the protein–lipid interactions within the neuronal porosome complex will profoundly influence its structure–function. Interestingly, studies report that Na-K-ATPase porosome protein is functionally dependent on the acidic glycerophospholipid PS, which provides stability by binding the protein between the 8, 9, and 10 transmembrane segments of its α-subunit. Studies further demonstrate that the neutral glycerophosphophospholipids PC and phosphatidylethanolamine (PE) can bind to and activate the Na-K-ATPase, and Ca-ATPase in neuronal porosome bind to PE. These findings attest to the critical role of protein–lipid and protein–protein interactions within the neuronal porosome complex crucial to its structure and function.
Structural and functional reconstitution of the neuronal porosome
The structural and functional reconstitution of the neuronal porosome complex has been accomplished over a decade ago
8
using AFM and electrophysiology. AFM micrographs of isolated porosome complex reconstituted into lipid membrane demonstrated nearly identical morphological features with the native complex found at the presynaptic membrane (Figure 1). To determine the function of the isolated neuronal porosome complex, a EPC9 bilayer electrophysiological setup (EPC9) has been used (Figure 6).
8
In the EPC9 setup, membrane capacitance and conductance are continually monitored following reconstitution of immunoisolated neuronal porosomes into the artificial membrane and following exposure to isolated synaptic vesicles. Addition of synaptic vesicles to the cis compartment of the bilayer chamber of the EPC9 apparatus results in the transient docking and fusion of synaptic vesicles with the porosome-reconstituted membrane bilayer (Figure 6).
8
Several synaptic vesicles are observed to fuse at the bilayer, reflected in the large stepwise increases in membrane capacitance (Figure 6(d)); however, exposure to 50 µM ATP results in NSF-mediated t-/v-SNARE disassembly and the release of the transiently docked synaptic vesicles reflected in the return of the bilayers membrane capacitance to resting levels. Addition of full length recombinant NSF has no further effect on capacitance of the membrane bilayer suggesting that native NSF associated with the neuronal porosome complex is sufficient for t-/v-SNARE complex disassembly, resulting in the dissociation of synaptic vesicles from the bilayer. To further determine the dissociation of docked synaptic vesicles after treatment with ATP, isolated synaptosomal membranes have been subjected to ATP exposure and the resulting supernatant fraction evaluated for the presence of synaptic vesicles by detecting the presence of synaptic vesicle proteins SV2 and VAMP-2 using Western blot analysis (Figure 6(e)). These results exhibit that both SV2 and VAMP-2 proteins are enriched in supernatant fractions after treatment of synaptosomal membrane with ATP, in further confirmation of the electrophysiological measurements.
8
The briefly outlined aforementioned studies on the neuronal porosome complex (Figures 1 and 6) is evidence that the neuronal porosome is a unit fusion machinery or secretory portal, that can be isolated in its intact, stable, and functional state. This isolated neuronal porosome complex was used in the proteomic and lipidomic analysis presented in the previous section of the review.
Functional reconstitution of the neuronal porosome complex into artificial lipid membrane.
8
(a) a neuronal porosome complex at the presynaptic membrane with a synaptic vesicle (SV) docked at the base. (b) an EPC9 electrophysiological bilayers apparatus, for continual monitor of changes in the capacitance of porosome-reconstituted membrane following introduction of isolated SV into the cis compartment, in the presence and absence of ATP and the purified N-Ethylmaleimide sensitive factor, NSF. (c) SV docking, fusion, and ATP-NSF induced dissociation, at an EPC9 membrane reconstituted porosome complex. (d) Capacitance measurements of porosome-reconstituted bilayers demonstrating increase in membrane capacitance following SV fusion and a return to resting state on ATP exposure. No further change is observed on the addition of exogenous NSF, suggesting that the endogenous NSF is adequate for the observed NSF-ATP mediated t-/v-SNARE disassembly and disengagement of the SV from the membrane (n = 6). (e) Exposure of lysed synaptosomes to ATP dislodges SVs from the membrane into the buffered suspension, which is reflected as an increase in SV-specific proteins SV2 and VAMP-2, in the suspension buffer. (A color version of this figure is available in the online journal.)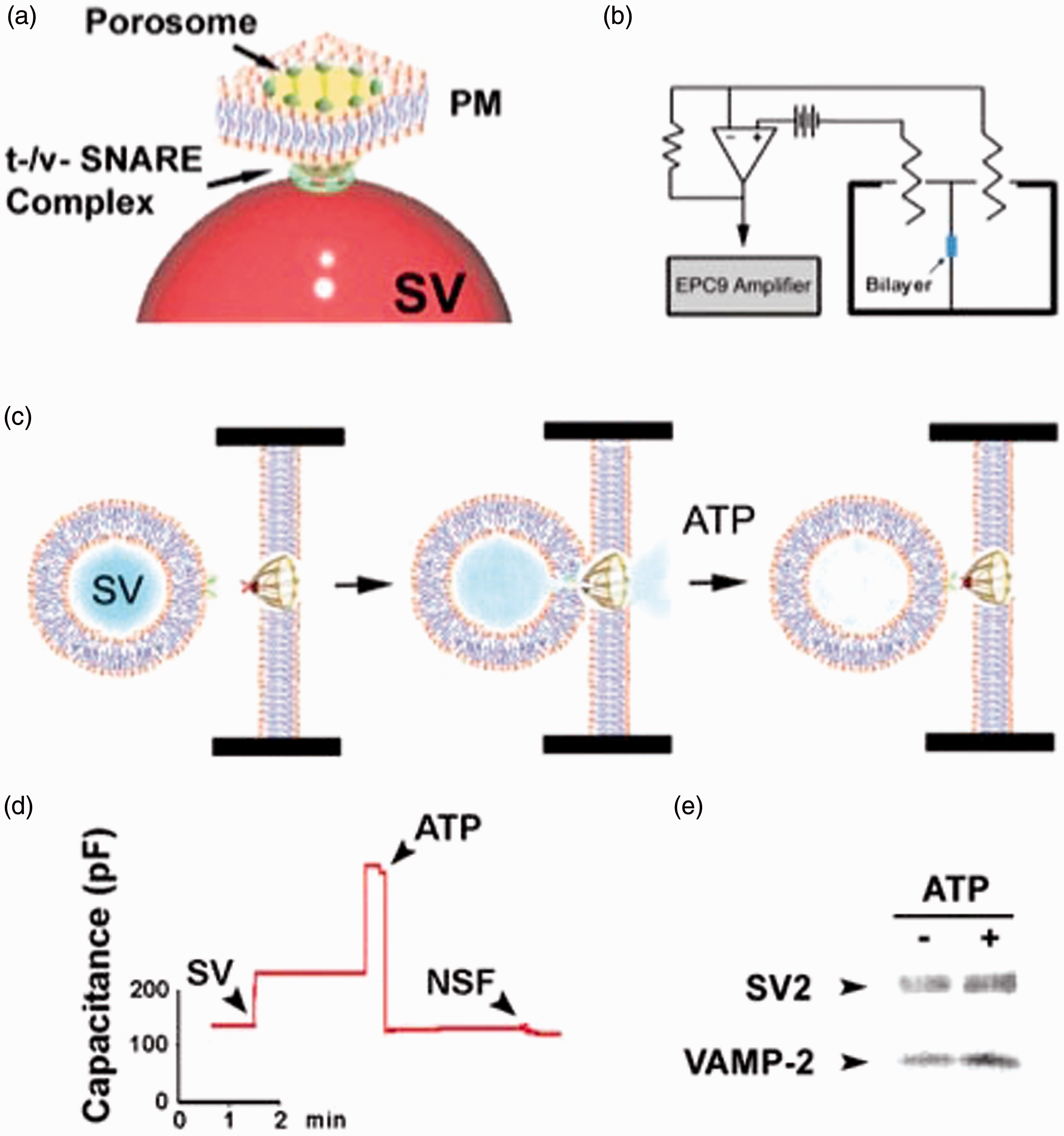
Porosome proteins implicated in neurotransmitter release and disease
Cytoskeletal proteins such as actin and the alpha chain of tubulin are two of the several neuronal porosome proteins detected using mass spectrometry. It has long been established that actin is involved in neurotransmission. 48 Studies performed in presence of latrinculin A, an actin depolymerizing agent, partially blocked neurotransmitter release at the presynaptic terminal of motor neurons. 49 Actin β protein, a post-translational product of actb mRNA, is important in formation of excitatory synapses, which is promoted by interaction of actb mRNA with Src associated in mitosis (Sam68) protein. Loss in Sam68 diminishes its interaction with actb mRNA leading to lower levels of synaptic actin β, which in turn leads to neurological disorders involved with abnormal synaptic transmissions. 50 Although tublin involvement in neurotransmission is not fully understood, its association with several synaptosomal proteins at the presynaptic membrane51,52 suggests an important role in neurotransmission. Besides actin and tubulin, other important classes of proteins identified in the neuronal porosome are the membrane integrated ion-channel proteins that are important in maintaining cellular homeostasis. The alpha subunit 3 of the universal Na+/K+ ATPase, identified in the porosome complex, seems to be involved in neuronal secretion. Na+/K+ ATPase activity is known to be blocked by dihydrooubain (DHO). 53 Activation of Na+/K+ ATPase inhibits synaptic transmission, which results from the secretion of a presynaptic protein ‘follistatin-like 1’ which activates the alpha 1 subunit of the ATPase. 54 Transient blocking of Na+/K+ ATPase pump using DHO is reported to result in an increase in both the amplitude and number of action potentials at the nerve terminal. 55 Interestingly, Na+/K+ ATPase inhibition is calcium dependent and it has been demonstrated that increased intracellular calcium levels inhibit Na+/K+ ATPase, which increases excitability of neurons. 56 Similarly, the porosome protein plasma membrane calcium ATPases (PMCA) are an important class of proteins known to be involved in maintaining calcium homeostasis within cells including neurons. PMCA2 has been shown to co-localize in synaptosomes with synaptohysin. 57 At the presynaptic membrane, syntaxin 1, also a porosome protein, has been demonstrated to co-localize with PMCA2 and the glycine transporter 2 (GlyT2) that is found coupled to the Na+/K+ pump suggesting the presence of a protein complex involved in neurotransmission.58–60 Similarly, deletion of PMCA2 generates an ataxic phenotype in mice where the neurons possess prolonged hyperpolarized membrane resulting from an increase in the basal levels of calcium within these neurons. 61 Mutation in the PMCA2 encoding gene is known to result in homozygous deafwaddler mice (dfw/dfw) and they show high levels of calcium accumulation within their synaptic terminals. 62 NAP-22, also known as BASP-1, is a protein found in the neuronal porosome complex, and due to its localization on the pre and postsynaptic membranes and also in synaptic vesicles, it has been long speculated to be involved in synaptic transmission.63–65 NAP-22 is known to bind to the inner leaflet of lipid rafts suggesting interaction with cholesterol. Adenylyl cyclase-associated protein-1 (CAP-1) is known to regulate actin polymerization 66 and both actin and CAP-1 are present in the porosomal complex. Cells depleted of CAP-1 using RNAi result in lamellopodia growth and F-actin accumulation along with other cytoskeletal abnormalities. 67 In Alzheimer’s, the levels of CNPase (2,3-cyclic nucleotide phosphodiesterase) and the heat shock protein 70 (HSP70) are found to increase, while the levels of dihydropyrimidinase-related protein-2 (DRP-2) decrease. 68 Decreased levels of CNPase have been detected in the frontal and temporal cortex of patients with Alzheimer’s disease and or Down’s syndrome.69,70 Low CNPase levels have also been detected in postmortem anterior frontal cortex in schizophrenic patients. 71 Additionally, an allele that is associated with low levels of CNPase is also linked to schizophrenia. 72 CNPase is also detected as a marker for oligodendroglia and myelin and several diseases associated with low levels of CNPase indicating low myelin on neurons. 73 CNPase positive cells have also been shown to increase in corpus callosum of rats exposed to an enriched environment, meaning given to perform a task. 74 Similarly, alterations in the levels of several of the SNARE proteins are associated with various neurological disorders. For instance, SNAP-25 and synaptophysin are significantly reduced in neurons of patients with Alzheimer’s disease.75–77 Mice that are SNAP-25 (+/−) show disabled learning and memory, and exhibit epileptic like seizures. 78 Similarly, over-expression of SNAP-25 also results in defects in cognitive function, 79 and loss of SNAP-25 is also associated with Huntington’s disease and a reduction in rabphilin3a, another protein involved in vesicle docking and fusion at the presynaptic membrane. 80 SNARE knockout mice are neonatally lethal, and mice with a dominant mutation in SNAREs are known to develop ataxia and show impairment in vesicle recycling capability. 81 Increase in synaptophysin levels along with SNAP-25 are also observed in Broddmann’s area in the postmortem brain of patients with bipolar disorder I. 82 Protein levels of synaptotagmin and syntaxin 1 are highly up regulated in areas of cerebral ischemia, which are known to have very active neurotransmission. 83 Mutations in certain regions of the protein syntaxin 1A, such as the Ca+2 channel-binding region, is known to increase neurotransmitter release suggesting syntaxin 1A to be involved in regulating Ca+2 channel function. 84 A point mutation in syntaxin 1A also results in augmented release of neurotransmitters in models of flies. 85 Synaptotagmin similar to syntaxin has Ca+2-binding domains, and is known to form dimers and interact with syntaxin to form complexes in the presence of calcium. 86 Binding of synaptotagmin to SNAP 25 is also calcium dependent. 87 Reticulons are proteins that contribute to lipid membrane curvature and are found in the neuronal porosome. The presence of reticulons with the porosome and diseases associated with their deregulation lend credence to the role of membrane curvature in modulating synaptic vesicle fusion at the porosome complex. These early molecular studies reflect on the critical role of various neuronal porosome proteins in protein–protein and protein–lipid interactions within the porosome complex and in their participation in normal porosome-mediated neurotransmission at the nerve terminal and in disease. Although beyond the scope of this review, it need mention that similar to the neuron, porosome proteins in other cell types such as the insulin-secreting beta cells of the endocrine pancreas and mucin-secreting human airways epithelia have similarly been implicated in secretion and in secretory diseases such as diabetes and cystic fibrosis.
In addition to the earlier molecular studies on the involvement of porosome proteins in neurotransmission, recent morphological studies on the neuronal porosome complex have also shed light on both the health and disease states of the neuronal porosome complex as examined using high resolution EM.88–92 Ultrastructure of the neuronal porosome complex in rats subjected to continuous white noise that is relevant to the increasing, random noise encountered by humans in present day environment, and also known to provoke diverse effects on different regions of the brain,93–98 demonstrates alteration in the porosome length. 92 Constant exposure to such noise is further known to sabotage the development and normal function of the auditory and some other brain regions, and ultimately impair hearing, language acquisition,93,94 memory performance,95,96 and other cognitive functions. The chemical basis of such alterations besides elongation of the porosome complex and the rapid synaptic vesicle fusion resulting from such morphological change to the porosome may reflect the corresponding abnormalities in neurotransmitter release that is currently being investigated.
Summary
All life processes are governed at the chemical level; hence, an understanding of cellular processes at the molecular stage is required. Bakers yeast to cells in humans secrete, which is required for a plethora of physiological functions such as neurotransmission and the release of hormones and digestive enzymes. Consequently, defects in cell secretion are responsible for a wide range of diseases, from neurological, immunological, to endocrine disorders such as diabetes. Secretory products are stored in membranous sacs or vesicles that dock and fuse with the cell plasma membrane to release intravesicular contents. It is known that during secretion secretory vesicles may collapse and fully merge with the cell plasma membrane to dump the entire contents of the secretory vesicle, or they may transiently dock, fuse, and release measured amounts of secretory products at membrane-associated supramolecular structures called porosomes. Porosomes are supramolecular cup-shaped lipoprotein structure at the cell plasma membrane in all secretory cells including neurons, where secretory vesicles transiently dock and fuse to expel a fraction of their contents to the cells exterior. Although the general role of porosomes as secretory portals is well established, and its overall structures at nm resolution known, a molecular understanding of its structure–function remains to be determined.
In neurons, porosomes are 15 nm cup-shaped lipoprotein structures composed of nearly 40 proteins, and enriched in several classes of lipids. Although STRING maps generated on the neuronal porosome proteome identifies associated proteins and their predicted interactions, further chemical cross-linking studies followed by mass spectrometry will be required to confirm these interactions, and to determine the stoichiometry of the proteins present within the complex, to be able to build a working model of the neuronal porosome structure. A great number of studies that implicate various porosome proteins in neurotransmission and in various neurological disorders further lend support that this secretory portal at the nerve terminal is critical to neurotransmission, and that its altered state leads to various neurological disorders and diseases requiring further molecular understanding of its structure–function. Being a membrane-associated supramolecular complex has precluded determination of the atomic structure of the porosome complex. Ultrahigh resolution AFM studies and SAXS have provided the three dimensional structure of the native neuronal porosome complex. SAXS and AFM studies have also helped to determine a portion of the molecular mechanism involved in porosome-mediated neurotransmitter exocytosis at the synapse. Despite the fact that the porosome in nerves are nearly 10 times smaller than those present in endocrine and exocrine cells, they all are similar if not identical in both their structure and composition. Nature has fine-tuned the porosome complex in cells of different tissues to optimally assist in their specialized functions. Therefore, both the porosome and secretory vesicle size may be a reflection of one aspect of the fine-tuning to accomplish specialized secretory function of a cell. Smaller secretory vesicles with greater curvature fuse more efficiently and rapidly in comparison to larger ones,119,120 suggesting that membrane curvature of both secretory vesicles and the porosome base would reflect on the potency and efficacy of membrane fusion during cell secretion. Neurons therefore utilize 30–50 nm in diameter synaptic vesicles and porosomes with small bases measuring 2–4 nm in diameter for rapid and efficient membrane fusion and neurotransmitter release. A slow secretory cell like the pancreatic acinar cell or the neuroendocrine growth hormone secreting cell or insulin-secreting beta cells possess larger secretory vesicles measuring 200–1200 nm that transiently dock and fuse at porosomes whose bases are much larger measuring 20–30 nm.
To determine the molecular structure of the neuronal porosome complex and to understand its function at that level, ongoing studies in the laboratory involve cryo EM, AFM, SAXS and neutron scattering analyses. Chemical cross-linking followed by mass spectrometry is being carried out to obtain molecular details of the neuronal porosome structure. Changes in the subunit composition of the porosome and the interactions between subunits during neurotransmission are also being carried out to further our knowledge of the porosome structure, dynamics, and composition. Experiments are also in progress involving multiple quantitative mass spectrometry and a number of biochemical and molecular approaches including CXL-MS to obtain further understanding of the porosome at the nerve terminal. To limit the number of false positives in chemical cross-linking studies, isolated porosomes are being utilized. Quantitative mass spectrometry (iTRAQ) and absolute quantification of proteins (AQUA) 121 are being performed to determine alterations in porosome subunit structure in different states of neurotransmitter release. Quantitative CXL-MS analysis with heavy isotope cross-linkers are planned, which will help in the identification of changes in molecular interactions within the porosome complex in both resting and stimulated states. Single particle cryo-EM tomography, 122 and SAXS studies combined with neutron scattering on isolated neuronal porosomes should serve to reveal the position of various proteins within the structure. Finally, computational approaches employing coarse-grain molecular docking studies,123–138 homology-modeled interactions,139–141 and fitting of known atomic structures of proteins and their interactions and the resultant complexes formed142–148 will be employed to further determine the molecular structure of the neuronal porosome. A molecular understanding of the neuronal porosome structure will help in revealing the crosstalk between proteins within the complex and in the regulation of neurotransmitter release, and hence the potential for the design and development of drugs to target neurological disorders and diseases.
Footnotes
Author’s contributions
ARN participated in writing portions of the manuscript. Limited survey of the literature on the role of porosome proteins in neurotransmitter release and their implication in various neurological disorders was performed by ARN and KTL. The overall layout and content, and part of the writing and revision of the manuscript was carried out by BPJ.
ACKNOWLEDGEMENTS
Work presented in this article was supported in part by the National Institutes of Health grants DK56212, NS39918 and the National Science Foundation grants EB00303, CBET1066661 (BPJ); the WSU Interdisciplinary Biomedical Systems Fellowship (ARN); the WSU Interdisciplinary Biomedical Systems Fellowship and the Thomas C. Rumble University Graduate Fellowship (KTL).