
Abstract
Interferons play important roles in defense mechanisms against viral infection, and thus interferon therapy has been a standard treatment in chronic hepatitis B patients. Interferons signaling pathways promote interferon-inducible genes including microRNAs. In this research, we aimed to determine microRNAs expression profiles in vitro and in vivo. For in vitro model, Huh7 cells were transfected with or without hepatitis B virus plasmid for 6 h, and then treated with 100 ng of pegylated-interferon alpha-2a for 24 h. In vivo, we defined microRNAs expression profiles in pair-liver tissues of chronic hepatitis B patients in comparison between before and after treatment of pegylated-interferon alpha-2a for 48 weeks. Cellular small RNAs were extracted followed by library preparation. To determine microRNAs expression profiles, the next-generation sequencing was carried out on MiSeq platform (Illumina®). In vitro analysis demonstrated that microRNAs can be classified into up-regulated and down-regulated microRNAs in response to hepatitis B virus, interferon, and combination of hepatitis B virus and interferon. Moreover, in vivo analysis revealed microRNAs profiles in non-responders, responders without hepatitis B surface antigen clearance, and responders with hepatitis B surface antigen clearance. The target genes of the candidate microRNAs were determined in terms of roles in cellular pathways and immune response, which might be related to treatment in chronic hepatitis B patients. Results revealed that two down-regulated microRNAs including miR-185-5p and miR-186-5p were correlated in both in vitro and in vivo studies. These two microRNAs might be represented as specific hepatic microRNAs responding to hepatitis B virus and pegylated-interferon alpha-2a treatment, which may remarkable and attractive for further study involving in the association of their target genes and prediction of pegylated-interferon alpha-2a response. Interestingly, microRNAs expression patterns might be useful for understanding the response mechanism and serve as biomarkers for prediction of pegylated-interferon alpha-2a treatment response in patients with chronic hepatitis B.
Introduction
Hepatitis B virus (HBV) is the main cause of acute or chronic liver disease which up to 70–90% of patient can lead to cirrhosis and hepatocellular carcinoma (HCC), according to 50% of HCC patients have presence of HBV infection.1,2 Almost 360 million people worldwide have chronic hepatitis infection with HBV, and 0.5 to 1.2 million people die each year from HBV-related liver disease. Prevalence of this disease depends on prevalence rates of HBV that can be varied by geographical regions and nationality population. 3 During HBV infection, host genome was integrated by HBV DNA and produced viral proteins such as HBx, hepatitis B surface antigen (HBsAg), and HBeAg. These viral proteins can trigger cancer-related genes, motivate several signaling pathways including genetic instability, inflammatory, and host immune responses, which will lead to hepatocyte injury and then transform to HCC. 4
Currently, anti-viral drugs have been used in the treatment of HBV infection. First of all, a nucleos(t)ide analog affects the inhibition of viral polymerase and also viral production. The second type of drugs is an interferon (IFN) alpha, which can be called as immunomodulatory drug triggering host immune responses and leads to inhibition of viral proliferation. 5 For the treatment of HBV patients, IFN alpha was the first permitted drug to use since 1980s.5,6 IFN alpha was attached with polyethylene glycol (PEG) to improve pharmacological advantages such as more resistance to protease activity and less renal clearance leading to extension of drug circulation time. 7 Treatment with IFN provides better treatment response without drug resistant occur. Furthermore, IFN treatment can show the definite duration of therapy.5,8 However, there are various side-effects of IFN treatment such as flu-like symptoms, headache, nausea, fatigue, weight and hair loss. 9 Furthermore, the Peg-IFN alpha treatment response rate is just 32–36% with high cost of treatment.10,11 Prediction of Peg-IFN alpha response in HBV-infected patients will be useful for personalized medicine or alternative treatments.
Small (18–24 nt in lengths) non-coding RNAs called microRNAs (miRNAs) play an important role in gene silencing via the mechanism of RNA interference pathway, which leads to the degradation or translational repression of mRNA target after matching the complement sequences between miRNAs itself and target mRNA. 12 Approximately 30% of human genes can be regulated by miRNAs. 13 From the previous studies, many evidences suggested that various cellular pathways and events such as cell development, differentiation, and cell immunity can be regulated by miRNAs. 14 Besides, miRNAs can be used as outstanding biomarkers of cancer prognosis and diagnosis, because they are easily detected and measured in human serum and plasma. The risen possibility at present is to find the unique miRNA patterns in serum and plasma that can be useful for performing non-invasive disease markers. 15
The first aim of this research is to determine differential expression of miRNA profiling in Huh7 cells in response to HBV and Peg-IFN alpha-2a treatment. Second, to investigate hepatic miRNAs expression profiles in chronic hepatitis B (CHB) patients between before and after treatment with Peg-IFN alpha-2a. Finally, to compare miRNAs expression profiles in CHB patients treated with Peg-IFN alpha-2a between responders and non-responders.
Materials and methods
Patients and sample collection
Baseline characteristics of patients with chronic hepatitis B (CHB)
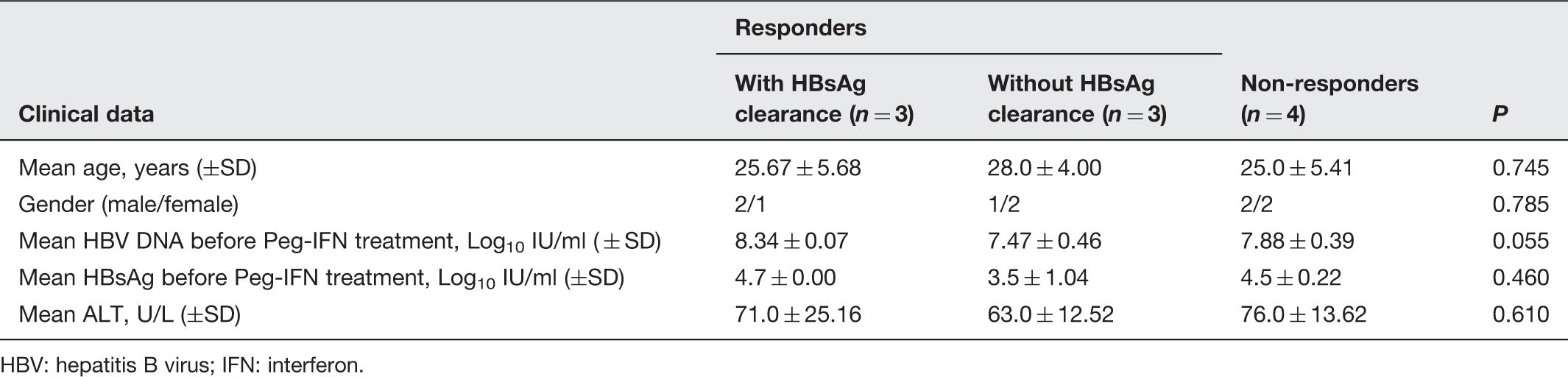
HBV: hepatitis B virus; IFN: interferon.
Plasmids and cell lines
The HCC cell lines (Huh7) and the plasmid expressing the HBV genome were generously provided by Prof. Yasuhito Tanaka, Nagoya City University Graduate School of Medical Sciences, Japan). The HBV plasmid was constructed by adding a full-length genome of HBV DNA genotype C (accession no. AB246345) into the pUC19 vector as described previously. 17 These plasmids were propagated in E. coli (DH5α) competent cells by heat shock transformation method. 18
Transfection of HBV plasmid
Huh7 cell line was cultured and maintained at 37℃, 5% CO2 humidified incubator in Dulbecco's Modified Eagle's Medium (Thermo Scientific) with 10% heat inactivated (56℃, 30 min) fetal bovine serum (Thermo Scientific) and 1% antibiotic-antimycotic (Gibco®). Cell cultures were divided into four different conditions in triplicates including mock infection, HBV expression plasmids transfection, Peg-IFNs alpha-2a treatment, and combined transfection of HBV expression plasmids and Peg-IFNs alpha-2a treatment. For transfection, the cells were seeded into 6-mm Petri dish (TPP) at a density of 7 × 105 cells/dish in medium without antibiotic-antimycotic and were incubated for 24 h. After the cells reached approximately 80% confluence, 3 µg of the HBV plasmid was diluted with Opti-MEM I reduced serum medium (Gibco®) and then transfected into cells by using 7.5 µL of Lipofectamine™ 2000 (Invitrogen) following the manufacturer’s guidelines. In IFN treatment condition, the cell cultures were treated with 100 ng of Peg-IFNs alpha-2a after 6 h of HBV transfection and then followed by 24-h incubation.
Isolation of miRNAs
Total miRNAs in cell cultures and clinical liver tissues were extracted by miRNA purification kit (Norgen) and the concentration of small RNA was quantitated by Qubit® BR RNA Assay kit (Invitrogen) following the manufacturer’s protocol.
Library preparation and next-generation sequencing
Small RNA libraries preparation begins with at least 100 ng/sample of total small RNAs using NEBNext® Multiplex Small RNA Library Prep Set for Illumina® (New England Biolabs). Briefly, small RNA was ligated with 3′ SR adaptor, and 5′ SR adaptor was then reverse transcribed into a cDNA library. After that, the cDNA library obtained from each sample was amplified by PCR with different indexes (distinctive 6 base-labeled). The size-selection of DNA library was performed through 3% agarose gel electrophoresis. The specific product size of DNA library (the length of miRNA with adaptors approximately 147 bp) was extracted by QIAquick gel extraction kit (QIAGEN). The DNA libraries were quantified by KAPA SYBR FAST qPCR Master Mix (2 × ) (Kapa Biosystems) according to the manufacturer’s recommendation. DNA libraries with different indexes were pooled together with equal concentration to make a 2 nM master DNA library. After that the DNA library was denatured by NaOH and then diluted using HT1 buffer in order to yield 10 pM DNA library which is suitable for next-generation sequencing (NGS). The NGS was performed using MiSeq® v2 reagent kit (Illumina®) carried out on MiSeq platform (Illumina®) according to Illumina’s instructions manual.
MicroRNA expression profiling
The CLC genomic workbench version 8 (http://www.clcbio.com/) was used for data processing and analysis of miRNA expression profiles. Low-quality reads (Q-score < 30) and adapter sequences were trimmed. The pass filtered reads (Q-score ≥ 30) were aligned with human genomic DNA (hg19), mature and precursor human miRNA (from miRbase), and contaminant RNA (human tRNA, rRNA, and mRNA). The sequencing reads which matched to the miRNA database but not contaminant RNA were considered as miRNA. The miRNAs were identified and counted based on the number of reads matched to the miRbase (www.mirbase.org/). Fold change between control and cases was calculated by log2 of normalized expression in case divided by normalized expression in control. Fold changes higher than 2 were considered as the significant difference. For miRNA profiling in vivo, candidate miRNAs were selected based on similar expression profiles (up-regulated or down-regulated) observed in the same group (responders with HBsAg clearance, responders without HBsAg clearance or non-responders). RNAhybrid (https://bibiserv2.cebitec.uni bielefeld.de/rnahybrid) was used for prediction of hybridization pattern between miRNA and their target genes.
Results
Analysis of miRNAs profiling by NGS
In cell culture model, miRNAs collected from the triplicate wells of each group were pooled together and used for preparation of four DNA libraries, including mock, Peg-IFN alpha-2a treatment, HBV transfection and HBV and Peg-IFN alpha-2a combination. In CHB patients, miRNAs extracted from 10 pair-liver biopsy of pre-treatment (week 0) and post-treatment (week 48) with Peg-IFN alpha-2a were used to construct 20 DNA libraries for individual sample. All 24 DNA libraries were pooled with equal concentration and subjected to run at the same time on MiSeq platform. The DNA library obtained from each sample was identified by different indexes. The results obtained from NGS yielded raw data 18,598,911 total reads. After trimming of low-quality reads (Q-score < 30) and adapter sequences, the passed filter reads (Q-score ≥ 30) were 17,761,014 reads (97.37% of total reads). Average reads per index were 740,042 reads. High-quality filtered reads were achieved and used for further analysis. The miRNAs were identified and counted based on the number of reads matched to the miRbase (www.mirbase.org/).
MiRNAs in Huh7 cells responding to Peg-IFN-alpha 2a and HBV
Up-regulated and down-regulated miRNAs in Huh7 cells responding to Peg-IFN-alpha-2a treatment and HBV transfection
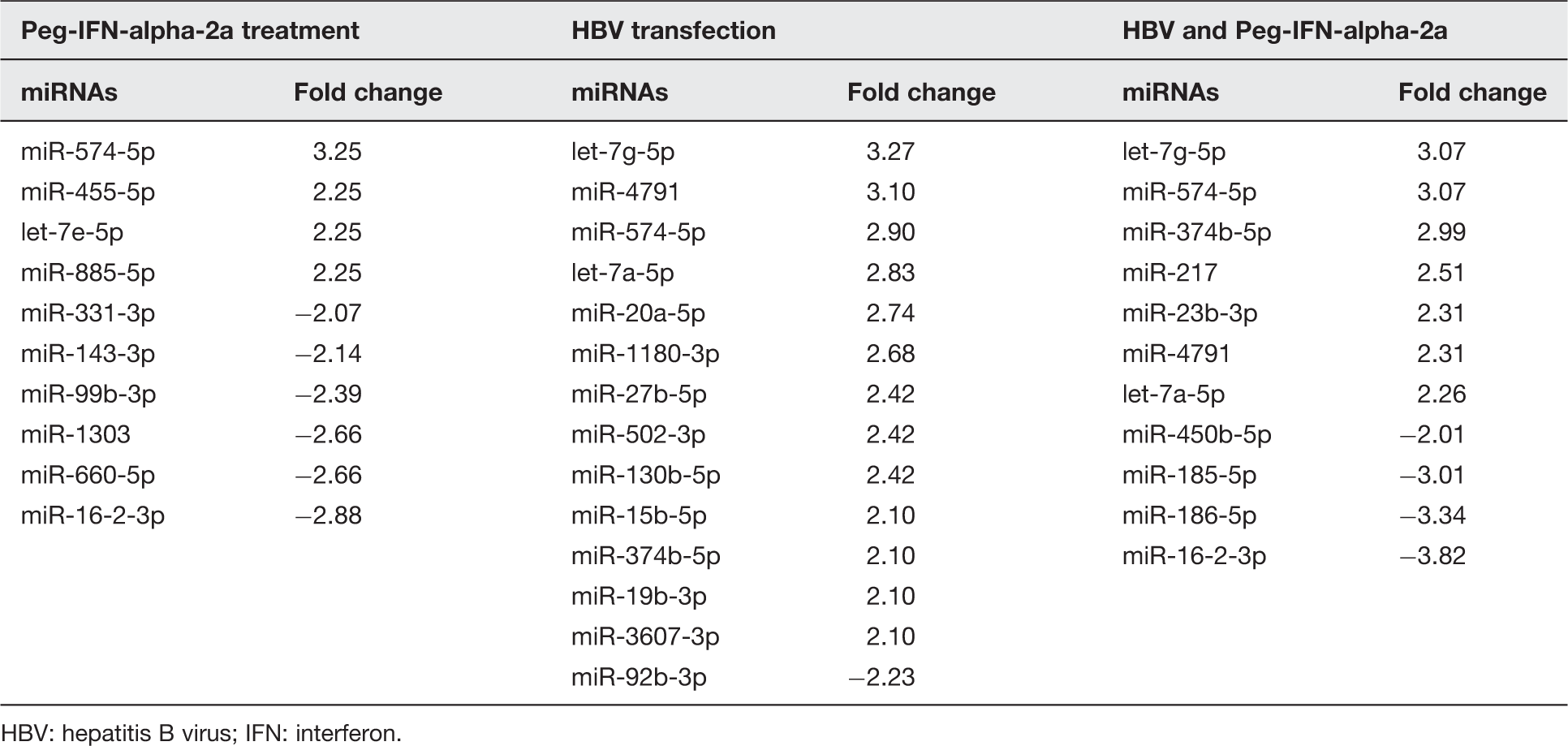
HBV: hepatitis B virus; IFN: interferon.
Common miRNAs in Huh7 cells responding to Peg-IFN-alpha 2a and HBV
For the characterization of miRNAs which is commonly expressed among groups and those exclusively expressed in each group of interest, Venn diagram was used to distinguish all miRNAs into three groups as shown in Figure 1. The results revealed miR-574-5p in the center of Venn diagram which was found to be up-regulated in all the three groups. In addition, up-regulated miRNAs observed between HBV transfected group and combination of HBV and Peg-IFN alpha-2a group were let-7g-5p, let-7a-5p, 374b-5p and miR-4791. These miRNAs possibly expressed when the cell infected with HBV and involved with endogenous IFNs response in the cell which enough to induced miRNAs in this group. The miRNA expression in cells treated with Peg-IFN alpha-2a between with and without HBV transfection revealed that only miR-16-2-3p was down-regulated which probably resulted from treatment with IFNs.
Venn diagram representing unique and common miRNAs which significantly expressed (fold changed > 2) in Huh7 cells culture model. Cells were divided into four groups including mock control, HBV transfected, Peg-IFN alpha-2a treated, and combination of HBV and Peg-IFN alpha-2a. (a) The up-regulated miRNAs in each group compared with mock control. (b) The down-regulated miRNAs in each group compared with mock control
Hepatic miRNAs profiling in CHB treated with Peg-IFN alpha-2a
MiRNA profiles were determined in pair-liver tissue samples obtained from 10 patients with CHB during pre- and post-treatment (48 weeks) with Peg-IFNs alpha-2a. The miRNA expression profiles were compared between pre-treatment and post-treatment in each individual patient. Then the patients were classified based on the outcomes of the treatment including responders without HBsAg clearance (n = 3), responders with HBsAg clearance (n = 3) and non-responders (n = 4). Therefore, miRNA profiles were compared among patients within the same group in order to determine the consensus miRNA expression patterns which might be useful as predictive biomarkers for the outcomes of treatment. The results revealed that all of the consensus miRNAs observed from patients within the same group were down-regulated. For responders without HBsAg clearance revealed the decrease of 5 miRNAs (let-7d-5p, let-7f-5p, miR-155-5p, miR-181a-5p and miR-423-5p). The reduction of six miRNAs expression in responders with HBsAg clearance such as miR-183-5p, miR-185-5p, miR-186-5p, miR-199b-5p, miR-422a and miR-424-5p is influenced by the treatment of Peg-IFN alpha-2a, whereas non-responders showed a decline of miR-145-5p and miR-199a-5p. There is no overlapping miRNA expression pattern observed among patients in different groups as shown in Figure 2.
Venn diagram representing consensus of down-regulated miRNAs observed among groups of CHB patients after treated with Peg-IFN alpha-2a for 48 weeks including responders without HBsAg clearance, responders with HBsAg clearance, and non-responders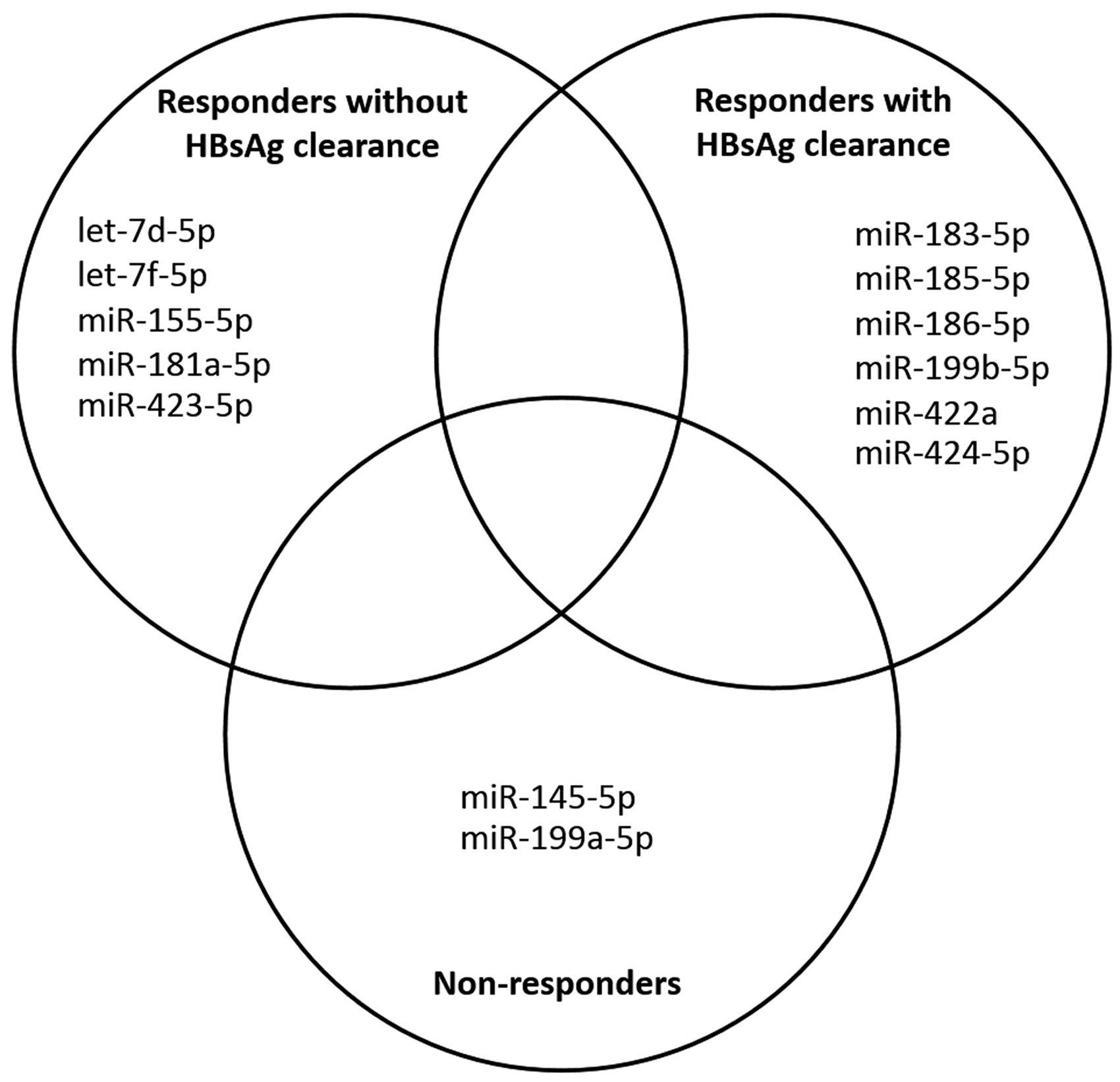
Target genes of candidate miRNAs
Hybridization patterns between candidate miRNAs and target genes.

AKAP12/AKAP250/Gravin: A kinase anchor protein 12; BTK: Burton tyrosine kinase; CD3G: CD3g molecule; gamma; DACH1: Dachshund family transcription factor 1; KRAS: Kirsten rat sarcoma viral oncogene homolog; Mzb1: Marginal zone B and B1 cell-specific protein; MFE: minimum free energy; PAX6: Paired box 6; PTEN: phosphatase and tensin homolog; PDCD4: programmed cell death 4; SMAD3: SMAD family member 3; v-kit Hardy-Zuckerman 4 feline sarcoma viral oncogene homolog (KIT); Wee1: WEE1 G2 checkpoint kinase.
Discussion
Currently, cell culture models have been used for studying the mechanism of cancer cells. HepG2, HepG2.2.15 and Huh7 cell lines have been favored for representing HCC cells. However, previous studies demonstrated that the expression level of IFN-α mRNA in Huh7 cells was more than those found in HepG2 cells, 32 meaning that Huh7 cells could respond to IFN better than HepG2 cells. Moreover, a previous study found that the HepG2 cells completely lacked primary response genes of IFN pathway. 33 Therefore, Huh7 cell culture would be more suitable than HepG2 cells for being a model to investigate the effect of Peg-IFN alpha-2a in this study. The result found that Huh7 could respond to HBV transfection and Peg-IFN alpha-2a treatment as differential miRNAs expression. MiRNAs expression in each group would affect the related target genes by the regulation of cellular gene expression. The HBV plasmid was used instead of HBV infection due to the fact that the HBV still lacks effective cell culture system in a laboratory. Therefore, 6-h HBV plasmid transfection was used to mimic HBV acute hepatitis B infection, but it could not represent CHB infection.
For miRNAs responding to HBV, a previous report found that miR-15b promotes HBV replication and expression of HBx protein which leads to repression of the miR-15b expression in long-term events. 34 However, this study found the up-regulation of miR-15b, which might be due to the acute phase (6 h) of HBV plasmid transfection. There were some studies about miRNAs expression in response to Peg-IFN alpha treatment. Cheng et al. studied the effect of IFN alpha-2b to miRNAs expression level in Huh7 cells by incubating with 1 ng/ml of IFN alpha-2b for 12 h and quantified miRNAs expression by TaqMan miRNA profiling. The study of Cheng and colleagues found up-regulation of let-7e, miR-455-5p and down-regulation of miR-1303 which similar to those found in our study. In contrast, miR-143, miR-99b, miR-331, miR-660 and miR-885-5p showed different expression levels between our study and the previous report. 35 The variations might be due to different doses of IFN and measuring time of the miRNA expression.
In combined treatment group (HBV and Peg-IFN alpha-2a), our study found that miR-23b-3p and miR-217 were up-regulated while miR-185-5p, miR-186-5p and miR-450b-5p were down-regulated. Putative target genes of these candidate miRNAs were predicted by RNAhybrid and confirmed by previous researches. In up-regulated miRNAs, miR-23b-3p has been found in targeting the 3′ UTR of SMAD family member 3 (Smad3), leading to down-regulation of transforming growth factor-beta1 (TGF-β)/bone morphogenetic protein signaling-induced apoptosis19,20 and association with cell proliferation, migration and differentiation.36,37 MiR-217 was reported as a biomarker for pancreatic injury. 38 Previous study has shown that miR-217 can target oncogenes or tumor suppressor genes. As in breast cancer, miR-217 can promote cell proliferation through targeting dachshund family transcription factor 1. 27 However, miR-217 acts as a tumor suppressor gene by targeting Kirsten rat sarcoma viral oncogene homolog in pancreatic ductal adenocarcinoma. 28
In down-regulated miRNAs, marginal zones B and B1 cell-specific protein (Mzb1) and Burton tyrosine kinase (BTK) are target genes of miR-185, which are related with T cells and B-cell developments. In DN3 cells and Jurkat T cells, there was the up-regulation of miR-185 leading to Mzb1 down-regulation and calcium responses effect in T-cell receptor (TCR), B-cell antigen receptor (BCR) and development of plasma cell. 24 Moreover, previous studies have found that BTK has a function in BCR cell signaling connected to Erk phosphorylation which is essential in follicular B-cell development regulated by miR-185.39,40 One target gene of miR-186 is a kinase anchor protein 12 (AKAP12/AKAP250/Gravin), which acts as a tumor suppressor gene by the decrease of cyclin D1 expression and also inhibits cell signaling of protein kinase A and protein kinase C through ERK2 and actin cytoskeleton. 25 Prior study revealed that down-regulation of Gravin expression was found in HCC and other cancers25,41,42 and also related to virulence of HCC progression. 25 MiR-450b-5p was found to target the pairbox 6 (Pax6) gene, which is important in fate decision of epidermal cell development 31 including pancreatic β-cell function.
In CHB patients, there were several significantly down-regulated miRNAs after treatment with Peg-IFN alpha-2a. In non-responders, down-regulation of miR-145-5p and miR-199a-5p were observed. MiR-145-5p was known as a tumor suppressor miRNA by regulating histone deacetylase 2 in liver cells.43,44 Down-regulation of MiR-199a-5p was induced by cisplatin leading to drug resistance through activating autophagy in liver cancer cell. 45 Moreover, a study found that miR-199a-5p could target discoidin domain receptor tyrosine kinase 1 and affected the increase of cell invasion in HCC. 46
The results obtained from CHB patients treated with Peg-IFN alpha-2a showed that down-regulation of let-7d-5p, let-7f-5p, miR-155-5p, miR-181a-5p and miR-423-5p were found in responders without HBsAg clearance group whereas down-regulation of miR-183-5p, miR-185-5p, miR-186-5p, miR-199b-5p, miR-422a and miR-424-5p were observed in responders with HBsAg clearance group. Generally, miR-183 acts as a potent oncogene by targeting tumor suppressor genes such as EGR1 and PTEN. 21 MiR-183 significantly up-regulates in HCC, targeting the programmed cell death 4 inducing cell growth and cancer development. 22 In this study, miR-183-5p was down-regulated in responders with HBsAg clearance which implied that the decrease of miR-183-5p might be a potential marker for treatment responsiveness in CHB patients. MiR-199b-5p was identified as a positive regulator of human erythropoiesis by targeting c-Kit expression. 26 CD3G is one of cluster of differentiation 3 (CD3) subunits assembled in TCR complexes targeted by miR-422a. 29 CD3 subunits interact with TCR to form a CD3-TCR complex to play a key part in cell recognition events. 47 MiR-424-5p targeted Wee1 kinase, similar to miR-128, leading to cell cycle arrest. 30 Target genes of miR-185-5p and miR-186-5p were explained above.
Comparisons of miRNAs expression profiling between Huh7 cell (in vitro) and liver tissues of CHB patients (in vivo) were performed in order to determine the hepatic miRNAs in response to HBV and Peg-IFN alpha-2a treatment. Results revealed that two down-regulated miRNAs including miR-185-5p and miR-186-5p were correlated in both in vitro and in vivo. This implied that these two miRNAs might be represented as specific hepatic miRNAs responding to HBV and Peg-IFN alpha-2a treatment. Thus, miR-185-5p and miR-186-5p may remarkable for further study involving in the association of their target genes and trends to predict Peg-IFN alpha-2a response. Other uncorrelated miRNAs observed between in vitro and in vivo might be due to several factors. Firstly, miRNAs observed in liver tissues of CHB patients were affected from various cell types (hepatic stellate cells, kupffer cells, liver sinusoids and endothelial cells), whereas miRNAs observed in Huh7 cells were expressed from only hepatic cells. Secondly, miRNAs in liver tissues might be affected by circulating miRNAs and various stimulants (hormones, cytokines and xenobiotics) from other tissues, while miRNAs in cell culture were homogenously obtained from Huh7 cells. In addition, HBV plasmid transfection in cell culture represented as an acute HBV infection, whereas HBV infection in CHB patients was in chronic stage. Moreover, the IFN treatment in Huh7 cells was a single dose short-term treatment (24 h), but CHB patients were treated with multiple doses in long-term treatment (48 weeks).
In conclusion, miRNAs expression patterns might be useful as biomarkers for prediction of Peg-IFN alpha-2a treatment response in CHB patients. However, this study has some limitations in terms of small clinical sample sizes and invasive sample collections. The small sample sizes are limited by the difficulty to obtain pair-liver tissue samples (before and after treatments). Moreover, liver tissue collections from CHB patients are done by invasive methods which implied that hepatic miRNAs are not suitable for biomarkers. Therefore, large sample sizes and non-invasive sample collection (serum or plasma miRNAs) should be further studied in order to identify suitable serum or plasma miRNAs as a biomarker for prediction of Peg-IFN alpha-2a treatment response in CHB patients.
Footnotes
Authors’ contributions
TJ analyzed the miRNA profiling and drafted the manuscript. NC assisted with liver tissues collection and cell culture. WP carried out the NGS process and data processing. KP performed NGS libraries preparation. JM assisted with data analysis and revised the manuscript. RK and KJ assisted with sample processing and libraries preparation. PT provided liver tissue samples and clinical data. SP designed the study, analyzed data, revised the manuscript, and coordinated the project.
Acknowledgements
We would like to gratefully acknowledge the Department of Biochemistry and Research affairs, Faculty of Medicine, Chulalongkorn University for the research facilities. This study was kindly supported by the Research Unit of Hepatitis and Liver Cancer, Chulalongkorn University. Funding was supported by a Joint Research Program between Thailand and Japan (NRCT-JSPS); the National Research Council of Thailand Grant under the Research University Network (RUN) initiative; the Thailand Research Fund (TRF: RSA5680031); Ratchadapiseksompotch Fund (Faculty of Medicine, Chulalongkorn University); the Research Chair Grant, the National Science and Technology Development Agency (NSTDA); National Research University Project; Office of Higher Education Commission; Chulalongkorn Academic Advancement into Its Second Century Project; The Scholarship from the Graduate School, Chulalongkorn University to commemorate 72nd Anniversary of his Majesty King Bhumibol Adulyadej and the 90th Anniversary Chulalongkorn University Fund (Ratchadaphiseksomphot Endowment Fund) are gratefully acknowledged.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.