
Abstract
Leptin promotes the growth of gastric cancer cells in vitro. It is, however, unknown if leptin induces gastric cancer in vivo. This study therefore investigated the effect of leptin on the histology and expression of tumorigenic genes in the stomach of rats following 40 weeks of leptin treatment. Male Sprague-Dawley rats, aged 6 weeks, were randomized into control and experimental groups (n = 8 per group). The experimental group was given intraperitoneal injections of leptin (60 µg/kg/day) once daily for 40 weeks, whereas the control group received intraperitoneal injection of an equal volume of normal saline daily. Rats were housed in polypropylene cages for the duration of the study. Body weight was measured weekly. Upon completion of treatment, rats were euthanized and their stomachs were collected for histopathological examination, microarray, and RT-qPCR. Data were analyzed using one-way ANOVA and Fisher’s exact test. On histology, one rat (12.5%) in the leptin-treated group had a large red-colored tumor nodule at the pyloric antrum of the stomach. Microscopically, stomachs of two leptin-treated rats (25%) showed hyperplasia or dysplasia. Microarray analysis revealed significant upregulation of a number of genes in the stomachs of leptin-treated rats that have been shown to be associated with tumorigenesis in other tissues, including Furin (protein maturation), Eef1a1 and Eif4g2 (translation factors), Tmed2 (vesicular trafficking), Rab7a (plasma membrane trafficking), Rfwd2 (protein degradation), Fth1 and Ftl1 (oxygen transport), Tspan8, Tspan1, Fxyd3, and Rack1 (cell migration), Pde4d (signal transduction), Nupr1 and Ybx1 (transcription factors), Ptma and Tmem134 (oncogenes), Srsf2 (mRNA maturation), and Reep5 (cell proliferation). None of the known oncogenes were, however, significantly up-regulated. In conclusion, although the overall effect of leptin on gastric carcinogenesis seems inconclusive, the findings of dysplasia and the up-regulation of some of the cancer-related genes nevertheless warrant further scrutiny on the role of leptin in gastric cancer.
Impact statement
Gastric cancer is the third most common cause of death due to cancer in the world. Obese individuals are at risk of developing gastric cancer, and the reason for this is unknown. Serum leptin levels are high in obese individuals and leptin is known to induce proliferation of gastric cancer cells in vitro. However, to date, no reports exist on the tumorigenic effects of leptin on the stomach in vivo. This study therefore determines if chronic leptin administration induces gastric carcinogenesis in non-obese rats, which might serve as a useful animal model for future studies. Although the findings are somewhat inconclusive, to our knowledge, however, this is the first study to show the up-regulation of numerous potential driver genes that highlight the potential role of leptin in the higher prevalence of gastric cancer among obese individuals. The findings certainly necessitate further scrutiny of leptin gastric cancer.
Keywords
Introduction
Leptin is encoded by the ob gene and is primarily synthesized by white adipose tissues. 1 It is also produced and secreted, albeit in small quantities, by the anterior pituitary gland, mammary epithelial cells, colonic mucosa, placenta, human ejaculated spermatozoa, and gastric mucosa.2,3 Leptin acts on the hypothalamus to regulate food intake and energy expenditure in order to maintain normal body weight. 4 Furthermore, leptin is essential in the regulation of glucose homeostasis, immune functions, and sexual maturation. 5
Leptin in excess, particularly in obese individuals, has been associated with a number of obesity-related diseases, including hypertension, infertility, and carcinogenesis. 5 Obesity has been identified as a risk factor in gastric cancer, colorectal cancer, prostate cancer, endometrial cancer, lung cancer, and breast cancer.6,7 An association between adipokines, in particular leptin, and the increased risk of developing breast cancer, colorectal cancer, prostate cancer, ovarian cancer, lung cancer, and gastric cancer has been reviewed recently. 7
Leptin has been shown to promote the proliferation of human breast 8 and gastric cancer cells 9 in vitro. However, there is no direct evidence linking leptin with gastric cancer in vivo. Gastric cancer is the third leading cause of cancer-related death in the world. 10 There is, therefore, a need to further investigate and identify the causes of gastric cancer, particularly in obese individuals where the prevalence of gastric cancer is high. This study therefore investigated the effect of chronic leptin administration on the stomach of rats. Histopathological examination was performed to identify the microscopic changes caused by chronic leptin treatment in rats. In addition, microarray analysis was done to screen for changes in gene expression in these stomachs, in particular those genes related to carcinogenesis.
Materials and methods
Animals and diets
Male Sprague-Dawley rats were obtained from Laboratory Animal Care Unit (LACU), Faculty of Medicine, UiTM Sungai Buloh, (Selangor, Malaysia). The rats were housed in polypropylene cages with commercial wood chip bedding in the animal room of LACU at a room temperature of 22–24°C, relative humidity of 50–55%, and a 12-h light/dark cycle. Their beddings were changed twice a week. Rats had access ad libitum to rat food (Specialty Feeds Pty Ltd Australia) and tap water. The handling of animals and all experimental treatment procedures were conducted in accordance with the Universal Guidelines for Care and Use of Laboratory Animals and approved by the Animal Care and Use Committee of the Faculty of Medicine (ACUC, Faculty of Medicine, UiTM, Malaysia) (Ref. No. ACUC-8/12).
Experimental animals and grouping
Sixteen , six-weeks-old male Sprague Dawley rats weighing 185–205 gm were randomized into a control and an experimental group. The experimental group was given 60 µg/kg/day of leptin once daily via the intraperitoneal route for 40 weeks (LEPT). The control group was given daily intraperitoneal injections of an equal volume of normal saline (CTRL). Body weights were measured weekly. The choice of the dose of leptin was based on our previous studies investigating the effect of leptin on blood pressure, blood pressure and proteinuria during pregnancy, and on sperm parameters in the rat, where it was found to show significant adverse effects but little effect on body weight.11–13 The duration of treatment was based on the N-Methyl-N'-Nitro-N-Nitrosoguanidine (MNNG) model of gastric adenocarcinoma. 14 Rats were fasted for 24 h before they were euthanized at 40 weeks. They were first lightly anesthetized with diethyl ether and then euthanized using a small animal guillotine. Their stomachs were collected and sectioned longitudinally, dividing the lesser and greater curvatures. Half of the stomachs were kept in 10% neutral-buffered formalin for histopathological examination using hematoxylin and eosin (H&E) staining. The other half of the stomachs was kept at −80°C for microarray and RT-qPCR analysis.
Histological examination
The stomachs were fixed in 10% formalin, processed, and then embedded in paraffin. Sections were made and stained with H&E. The H&E stained slides were examined under a digital compound microscope at 1000× magnification.
RNA extraction and cDNA preparation for microarray analysis
RNA was extracted using innuPREP_RNA Mini Kit (845-KS-2040050) (Analytik Jena, Germany); 3 µL of 10 ng RNA (3.33 ng/µL) was used to synthesize cDNA using Whole Transcript (WT) Pico Reagent Kit (P/N 703262) (Affymetrix, USA). The whole transcriptome expression of the cDNA was analyzed using GeneChip WT Expression Arrays in accordance with the manufacturer’s instructions (Affymetrix, USA). The data collected were analyzed using Expression Console Software 1.4 and Transcriptome Analysis Console (TAC) Software 3.1, which showed 30,429 genes, their fold changes, and P-values determined automatically using one-way ANOVA. Microarray analysis was performed on three stomach tissues from each group that showed close clustering of data intensity. The minimum cut-off fold change for LEPT-treated group compared to control was calculated and set at two standard deviations from the mean fold change (fold change ≥ 4.87 and ≤ −6.69) and with P < 0.05.
RT-qPCR
RNA from the same tissues used for microarray analysis was used for RT-qPCR. cDNA was reversely transcribed from RNA (100 ng/µL) using Maxima First Strand cDNA synthesis kit for RT-qPCR (Thermo Scientific Inc., USA). The cDNA was used as a template for real-time PCR using Luminaris Colour HiGreen qPCR Master Mix (2X) (Thermo Scientific, USA) incubated in CFX 96 PCR Thermal Cycler (Bio-Rad, USA). Primers were designed using Beacon Designer 8 and synthesized by Integrated DNA Technologies (IDT), USA. Table 1 shows the genes selected to confirm the result of the microarray analysis, including their sequences, annealing temperatures, and PCR efficiency values. Reference genes used were beta-actin (ActB) and ribosomal protein L29 (Rpl29). The selected target genes from microarray have validated full sequences (Nupr1, Tmem134, Rack1, Srsf2).
Primer sequences for RT-qPCR.
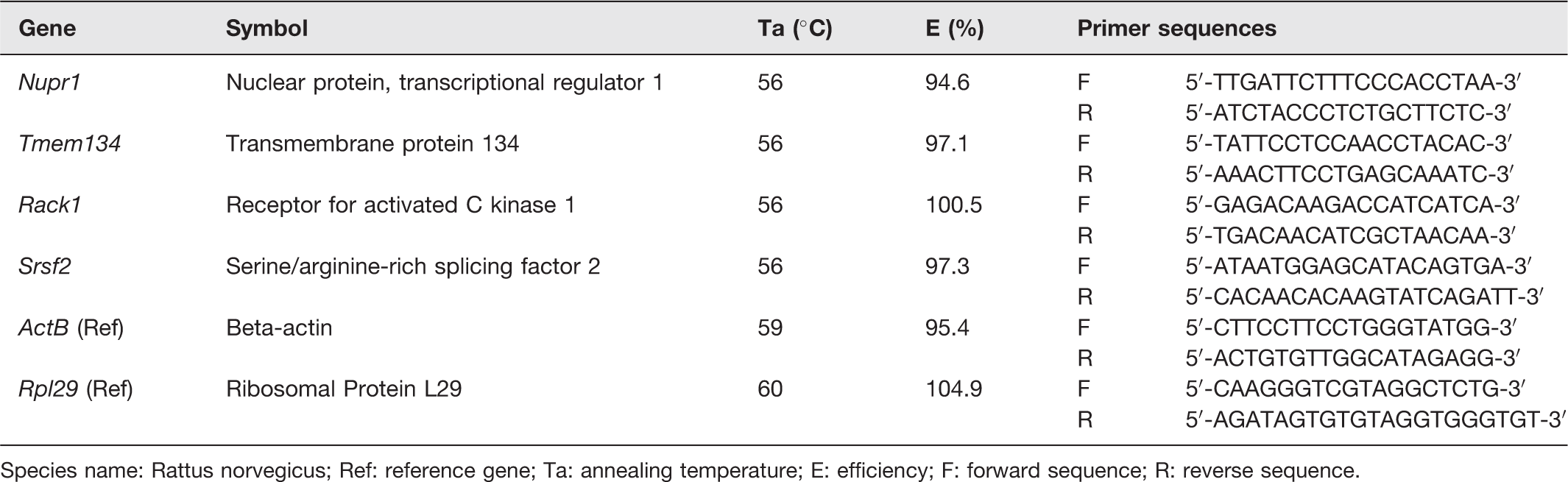
Species name: Rattus norvegicus; Ref: reference gene; Ta: annealing temperature; E: efficiency; F: forward sequence; R: reverse sequence.
Statistical analyses
Body weight data were analyzed using two-way ANOVA repeated measure, histopathological data were analyzed using Fisher’s exact test, and PCR data were analyzed using one-way ANOVA with Tukey as post hoc test in SPSS. P-value of <0.05 was considered statistically significant.
Results
Macroscopic examination of stomachs of rats
Figure 1 shows the macroscopic view of the stomachs with a large red-colored nodule at the center of the pyloric antrum in one of the rats in the LEPT group (Figure 1(b)). No macroscopic tumors were found in the stomach of control rats (Figure 1(a)).
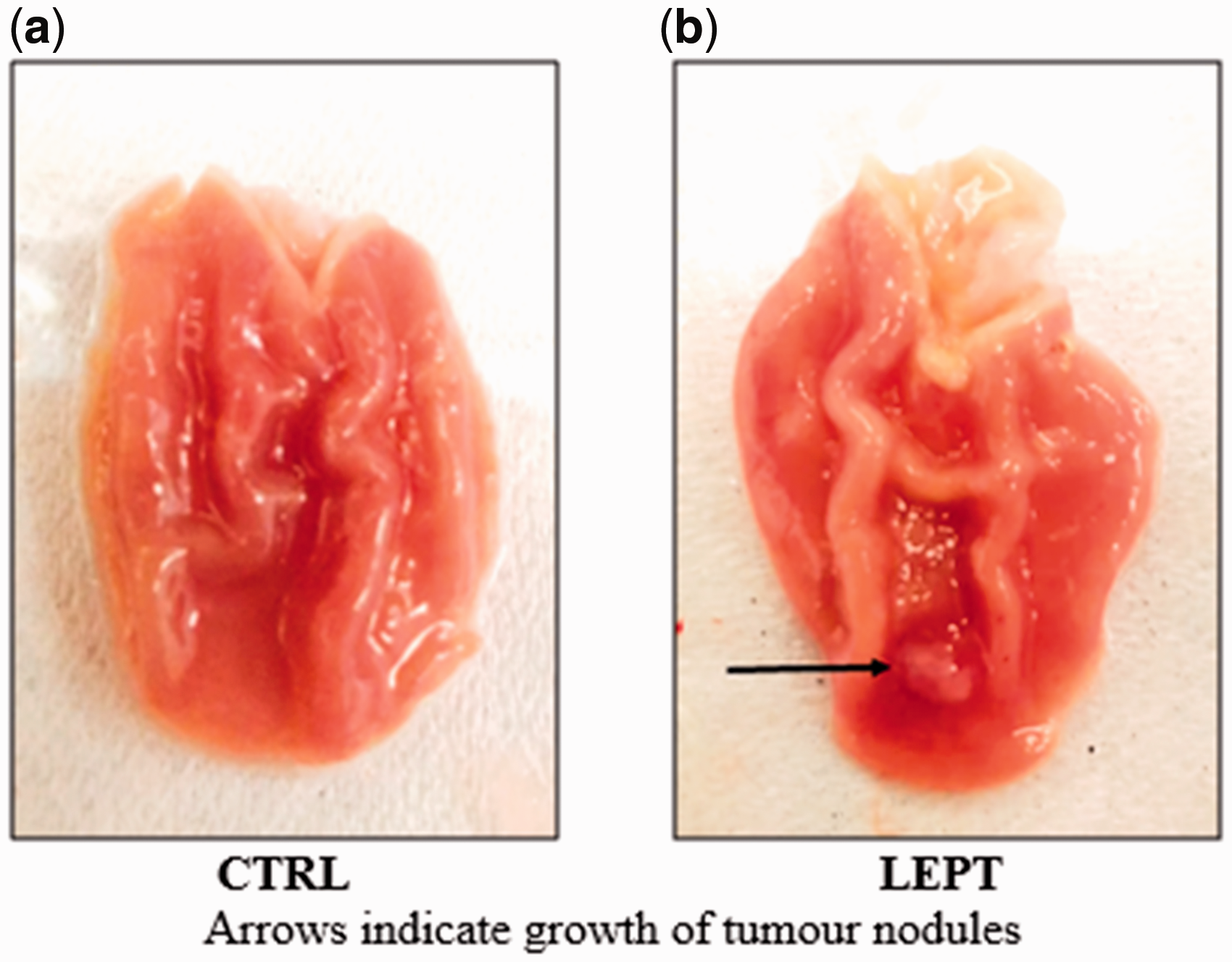
Macroscopic view of stomach of rats. (a) No tumor nodules were found in the stomach of control rats, (b) LEPT-treated stomach had one large red-colored tumor nodule located at the center of the pyloric antrum. (A color version of this figure is available in the online journal.)
Microscopic examination of stomachs of rats
Hyperplasia was evident in the stomach of one rat (1/8) and dysplasia (1/8) in another rat in the LEPT group (Figure 2(b) and (c)). There were no microscopic changes seen in the stomachs of the age-matched control rats (Figure 2(a)).
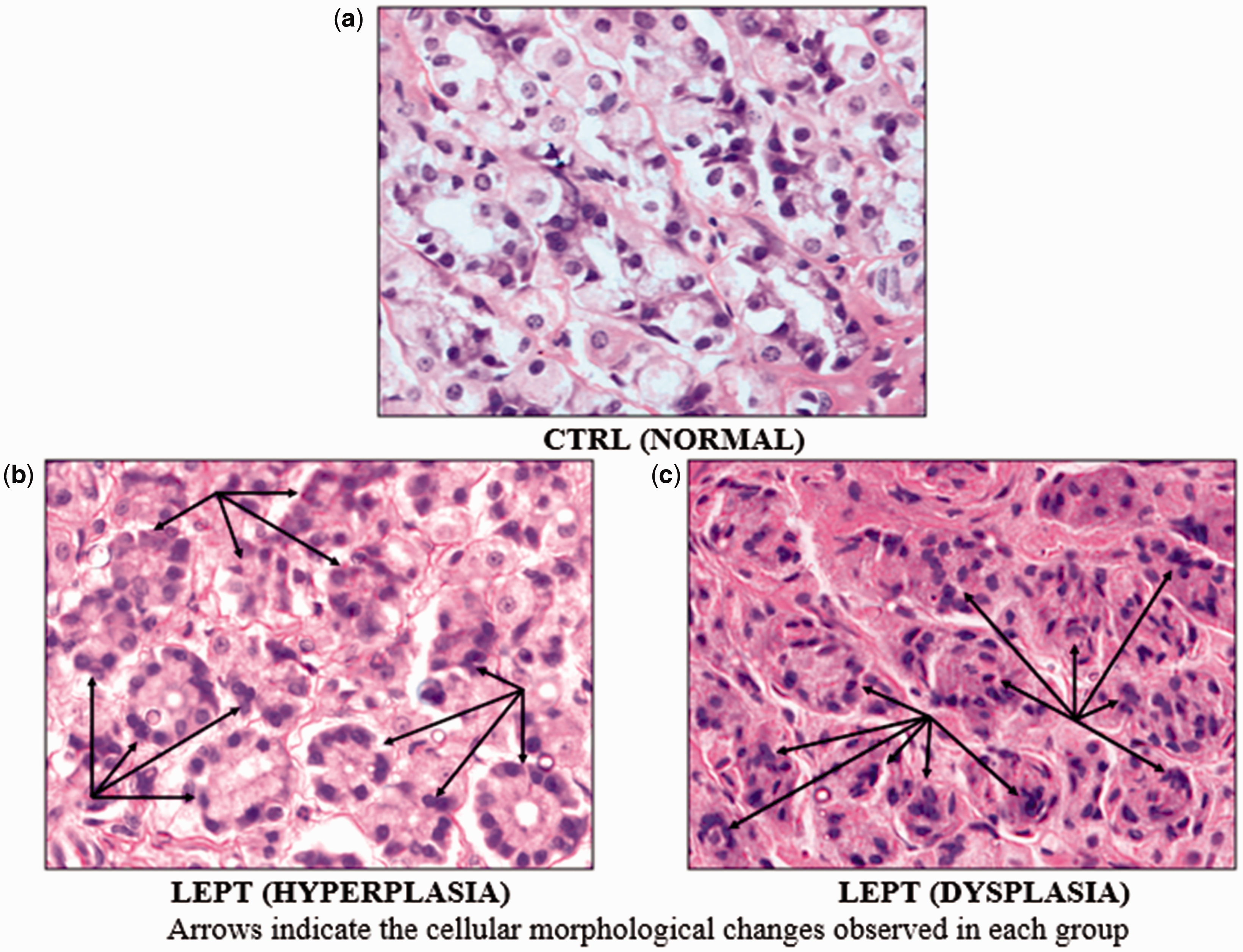
Microscopic view of the stomach of rats (×1000). (a) No microscopic changes in the control stomachs, (b) LEPT-treated stomach had hyperplasia, or (c) dysplasia. (A color version of this figure is available in the online journal.)
Global expression profile of LEPT-treated stomachs
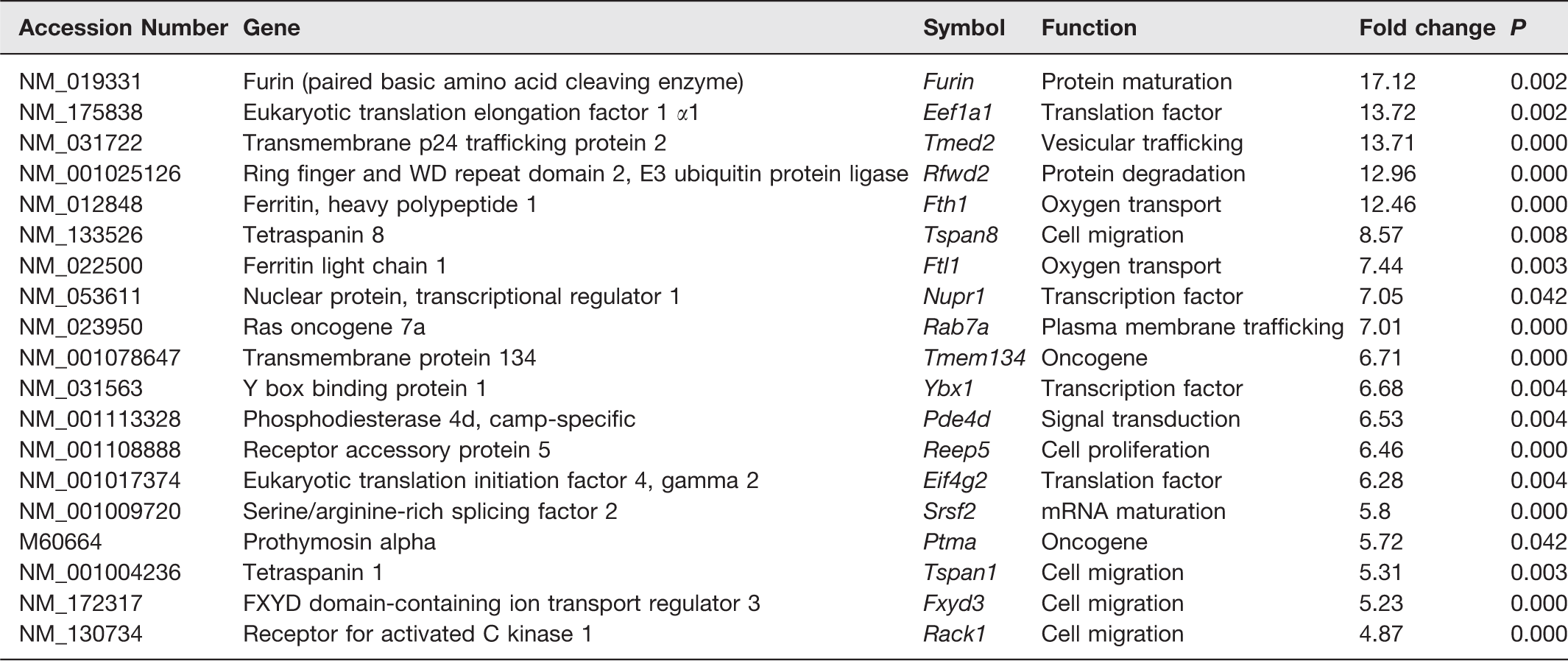
Global expression profile of LEPT-treated stomachs by microarray analysis set at a fold change of two standard deviations showed significant upregulation of 133 genes and downregulation of 18 genes. After further analysis, there were 16 genes that are thought to be most probably involved in gastric carcinogenesis (Table 2). These include Furin (protein maturation), Eef1a1 and Eif4g2 (translation factors), Tmed2 (vesicular trafficking), Rfwd2 (protein degradation), Fth1 and Ftl1 (oxygen transport), Tspan8, Tspan1, Fxyd3, and Rack1 (cell migration), Nupr1 and Ybx1 (transcription factors), Rab7a (plasma membrane trafficking), Ptma and Tmem134 (oncogenes), Pde4d (signal transduction), Reep5 (cell proliferation), and Srsf2 (mRNA maturation).
Genes differentially expressed in LEPT-treated stomachs.
Validation of microarray analysis by RT-qPCR
Figure 3(a) to (d) shows the expression of four target genes normalized to two reference genes (ActB and Rpl29) and analyzed using RT-qPCR. The expression of Nupr1 (transcription factor) (Figure 3(a)), Tmem134 (oncogene) (Figure 3(b)), Rack1 (cell migration) (Figure 3(c)), and Srsf2 (mRNA maturation) (Figure 3(d)) genes was significantly higher in LEPT-treated stomachs compared to control.
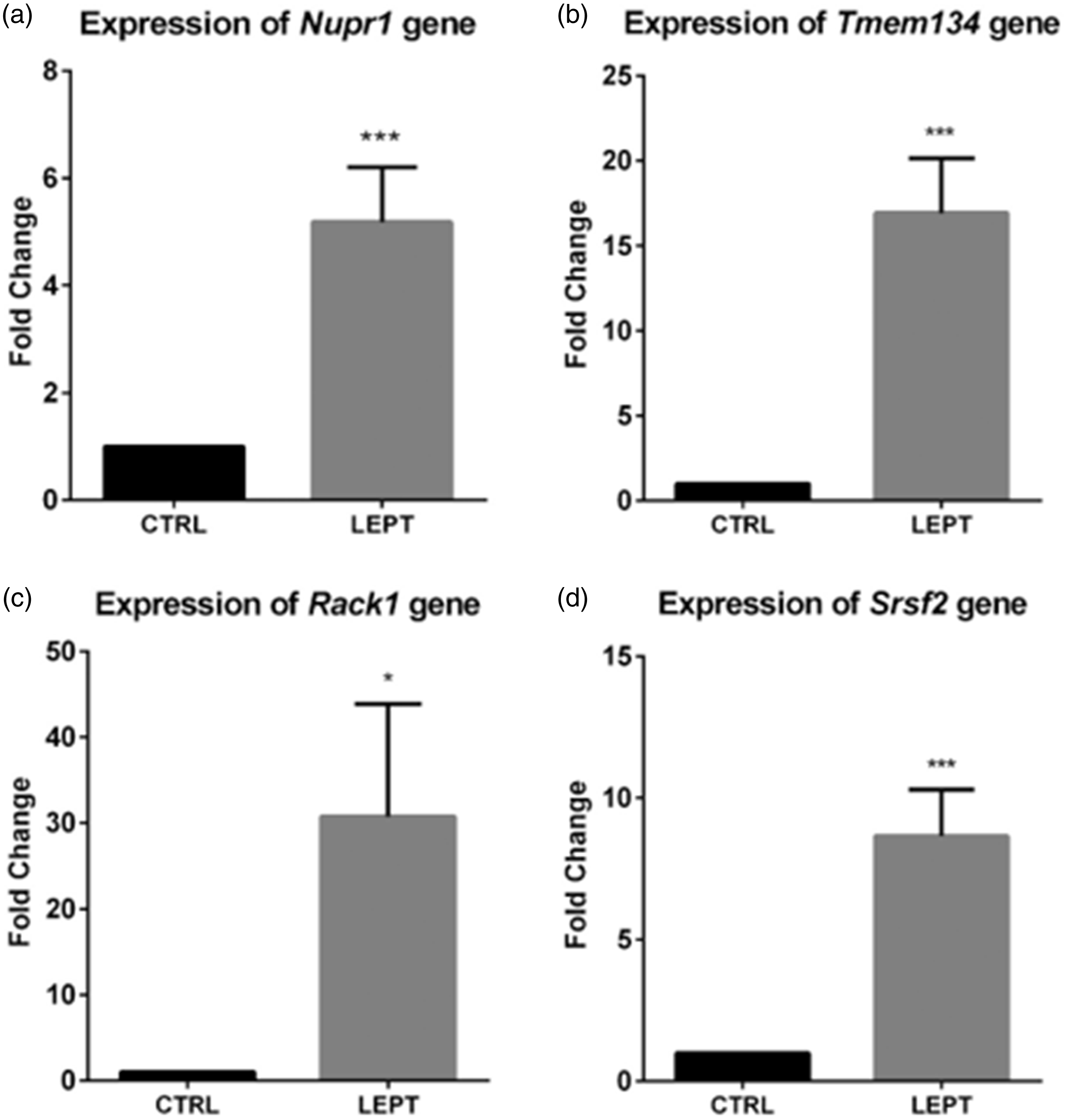
Expression of (a) Nupr1, (b) Tmem134, (c) Rack1, and (d) Srsf2 Gene in CTRL and LEPT-treated Rats. These expressions were significantly higher in LEPT-treated stomachs compared to control. *P < 0.05 compared to control, ***P < 0.001 compared to control.
Body weight
Body weight of rats increased significantly over the 40 weeks, but there was no significant difference in body weight between the groups across the study period (Figure 4).
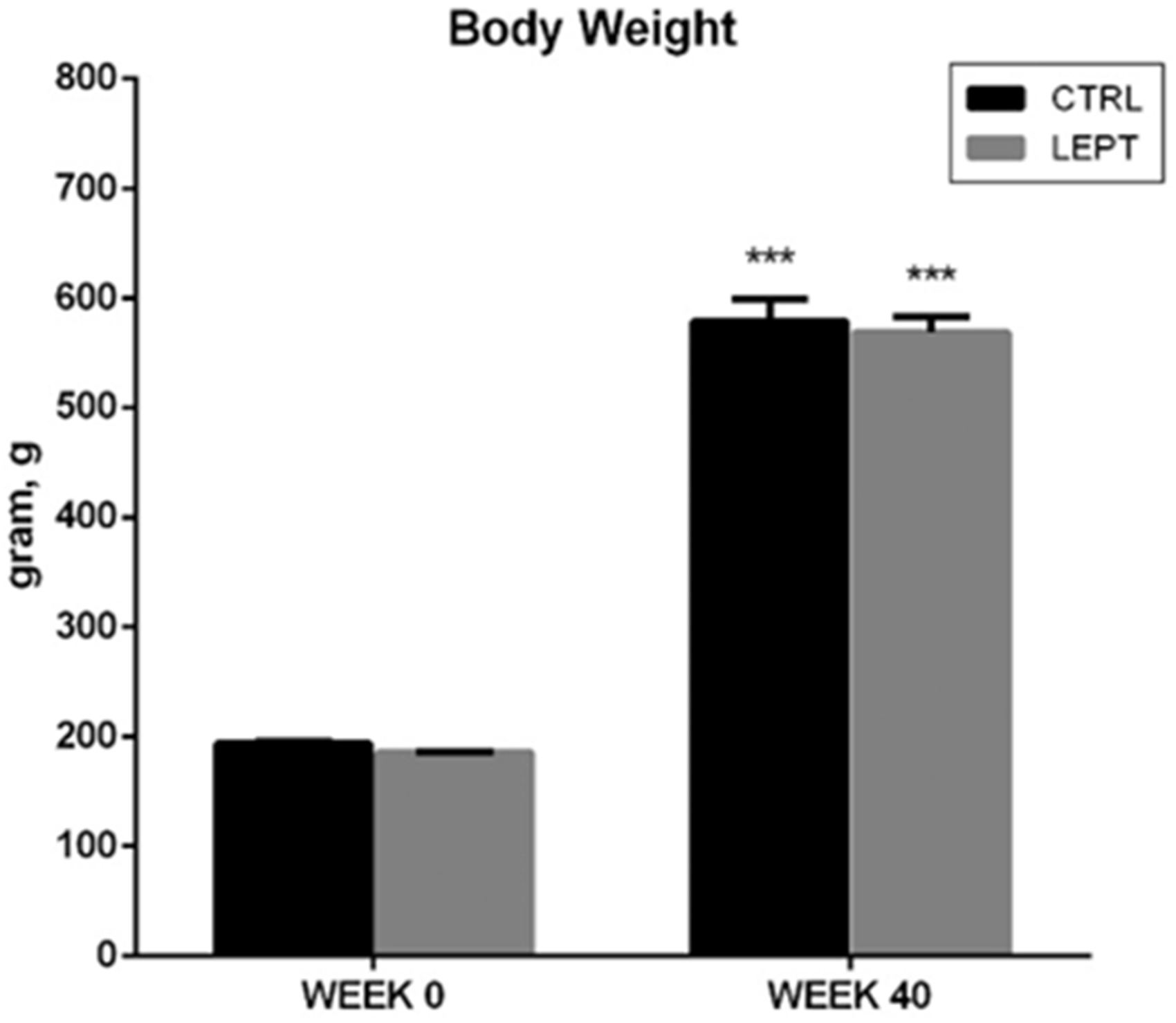
Mean body weight of rats at week 0 and week 40. The body weight of rats increased throughout the 40-week study period. No significant difference was seen in body weight between the two groups. ***P < 0.001 compared to their respective weights at week 0.
Discussion
The major findings of this study include, (i) the finding of a large tumor nodule (Figure 1(b)) in the stomach of one rat, and (ii) hyperplasia or dysplasia in stomachs of two of the eight rats treated with leptin (Figure 2(b) and (c)). No changes were seen in the saline-treated age-matched controls. Statistical analysis, however, did not reveal any significant differences between control and leptin-treated rats. To the best of our knowledge, there are no reports in the literature on the effect of chronic leptin treatment on the development of cancer in the stomach of rats. These findings, albeit inconclusive at this stage, are the first and seem to suggest the need for further study on the role of leptin in gastric cancer.
Although only one LEPT-treated rat developed a large tumor in the stomach, and dysplasia was evident in only one rat, but when these are read together with the micro-array analysis findings and information on the cell proliferative properties of leptin in the literature,8,9 the possibility that leptin might have a role in carcinogenesis certainly emerges. The reason why more leptin-treated rats did not show macroscopic and microscopic changes is unclear, but it is possible that a longer duration of leptin treatment or a higher dose of leptin may be required for these changes to take place in the stomach. It is also possible that leptin’s ability to induce carcinogenesis is enhanced in the presence of known carcinogens or vice versa. Examining the effects of leptin in the presence of known gastric carcinogens, like MNNG or Helicobacter pylori might be worthwhile. It might also be worthwhile examining the effect of leptin on the stomachs of other strains of rats or rodents as strain differences to MNNG-induced carcinogenesis has been reported before15,16.
Microarray analysis in all three samples of LEPT-treated stomachs revealed significant upregulation of a number of genes, which have been shown to be involved in tumorigenesis in other tissues. Although none of the known oncogenes particularly those associated with gastric cancer (EGFR, HER2, VEGF) 17 were significantly upregulated, there was nevertheless significant upregulation of genes that are known to induce dysplasia or carcinogenesis in other tissues (Table 2). It has to be added that microarray analysis was done on stomach samples from just three rats from each group and it is possible that examining the transcriptomes of only three rats could return many significant results, given the large number genes. Nevertheless, the corroborative evidence provided by the higher prevalence of gastric cancer in hyperleptinaemic obese individuals, 7 cell proliferative properties of leptin in vitro in the literature, 9 the necessary editing has been done and the finding of a tumor in one of the leptin-treated rats (Figure 1b) do support the microarray findings in this study, particularly when it shows the up-regulation of a number cancer genes, albeit, those associated with other organs or tissues. The genes that were found up-regulated include Furin (protein maturation), Eef1a1 and Eif4g2 (translation factors), Tmed2 (vesicular trafficking), Rab7a (plasma membrane trafficking), Rfwd2 (protein degradation), Fth1 and Ftl1 (oxygen transport), Tspan8, Tspan1, Fxyd3, and Rack1 (cell migration), Pde4d (signal transduction), Nupr1 and Ybx1 (transcription factors), Ptma and Tmem134 (oncogenes), Srsf2 (mRNA maturation), and Reep5 (cell proliferation). Past studies on gastric cancer have documented escalated expression of Rab, 18 Tspan8, 19 Tspan1, 20 Fxyd3, 21 and Ptma gene. 22 Furin modifies inactivated cancer-associated substrates to form matured functional proteins 23 and was found overexpressed in ovarian cancer. 24 Translation initiation factor Eif and translation elongation factor Eef are commonly upregulated in lung cancer, ovarian cancer, and breast cancer. 25 Tmed2 upregulation was associated with increased proliferation of ovarian cancer cells. 26 Overexpression of Rfwd2 was observed in hepatocellular carcinoma, ovarian adenocarcinoma, and breast cancer. 27 Cancer cells promote the production of Fth1 and Ftl1 (subunits that make up ferritin) to increase the transport of oxygen, as cancer cells thrive in an oxygenated micro-environment. 28 Pde4d overexpression was associated with an increase in cell proliferation of prostate cancer 29 and angiogenesis of lung cancers. 30 Transcription factor Nupr1 31 and mRNA maturation Srsf2 32 were overexpressed in hepatocellular carcinoma. Ybx1 was found to be upregulated in breast cancer cells, 33 and was identified as a prognostic biomarker for lung cancer. 34 Tmem134 showed oncogenic property in boosting the growth of prostate cancer. 35 Overexpression of Reep5 has been associated with increase in growth and invasion of lung cancer cells. 36
Despite the large range of differentially expressed genes induced by leptin treatment, it is possible that the genetic effects exerted by leptin might require a longer duration for their phenotypic expression. As suggested earlier, perhaps a longer duration of leptin treatment at this dose could have manifested more severe effects of leptin across the protein level, which would have provided microscopic evidence of the tumorigenic effects of leptin in inducing gastric carcinogenesis.
The body weight of male Sprague-Dawley rats increased significantly during the 40-week study period. More importantly, no significant changes in body weight were evident between the two groups at week 0 and at week 40 (Figure 4). Despite the changes found in the stomach of LEPT-treated rats, it appears that leptin treatment at the dose used in this study does not affect the body weight of the rats.
In conclusion, although the overall effect of leptin on gastric carcinogenesis seems inconclusive, the findings of hyperplasia, dysplasia, and the up-regulation of some of the cancer-related genes following 40 weeks of leptin treatment suggest a potential role for leptin in gastric carcinogenesis, which warrants further scrutiny.
Footnotes
Authors’ contributions
FI conducted the experiments, analyzed the data, interpreted the results, and wrote the manuscript; MKK participated in the study design, analyzed histology slides and microarray data, helped in the preparation of the manuscript and reviewed the manuscript; DD interpreted the results and reviewed the manuscript; NS analyzed microarray and RT-qPCR data; HJS participated in the study design, provided the overall supervision, interpreted the results, helped in the preparation of the manuscript and reviewed the manuscript.
ACKNOWLEDGEMENTS
Special thanks to all the members of the Adipokine Interest Group, Faculty of Medicine, UiTM for their comments and their continuous support. The authors wish to acknowledge the staff of Laboratory Animal Care Unit (LACU) and Institute of Medical Molecular Biotechnology (IMMB) for their technical help and support.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
This study was supported by 600-IRMI/MyRA 5/3/GIP (019/2017) research grant.