
Abstract
Extensive bowel resection results in short bowel syndrome. Absence of the ileocecal valve and length of remaining bowel are important prognostic factors. Such patients require total parenteral nutrition for survival, which has significant side effects, thus understanding mechanisms driving total parenteral nutrition-associated complications in short bowel syndrome is a major research focus. We hypothesized that we could develop an ambulatory total parenteral nutrition-short bowel syndrome piglet model recapitulating human short bowel syndrome for advanced research. Fourteen neonatal pigs received duodenal, jugular catheters, and a jacket with a miniaturized pump. Animals were randomly allocated to enteral nutrition (n = 5), total parenteral nutrition only (n = 5) or total parenteral nutrition with 75% small bowel, ileocecal valve resection, and ileo-colonic anastomosis (n = 4). Blood, liver, and gut were analyzed. Animals underwent successful bowel resection and anastomosis. Increased bilirubin was noted in short bowel syndrome and total parenteral nutrition. Mean conjugated bilirubin (mg/dL)±SE was 0.036 ± 0.004 for enteral nutrition (P = 0.03), 1.29 ± 0.613 for total parenteral nutrition (P = 0.01), and 3.89 ± 0.51 for short bowel syndrome (P = 0.000064). Linear gut density was reduced in short bowel syndrome and total parenteral nutrition vs. enteral nutrition. The mean linear gut density (g/cm)±SE for distal gut was 0.30 ± 0.02 for enteral nutrition (P = 0.0005); 0.16 ± 0.01 for total parenteral nutrition (P = 0.01), and 0.11 ± 0.008 for short bowel syndrome (P = 0.0001). We noted gut adaptation in short bowel syndrome (P = 0.015) with significant reduction in gut FXR, gut FGF19, and enhanced hepatic CyP7A1 expression in short bowel syndrome and total parenteral nutrition (P < 0.05). We successfully created an ambulatory total parenteral nutrition-short bowel syndrome model with distal small bowel and ileocecal valve resection recapitulating human short bowel syndrome. Our model validated total parenteral nutrition-related hyperbilirubinemia and gut changes, as noted in human short bowel syndrome. This model holds great potential for future innovative research and clinical applications.
Impact statement
Short bowel syndrome is associated with significant comorbidities and mortality. This study is important as unlike current systems, it provides a validated piglet model which mirrors anatomical, histological, and serological characteristics observed in human SBS. This model can be used to advance knowledge into mechanistic pathways and therapeutic modalities to improve outcomes for SBS patients. This study is novel in that in addition to significant reduction in the remnant bowel and noted liver disease, we also developed a method to emulate ileocecal valve resection and described gut adaptive responses which has important clinical implications in humans.
Keywords
Introduction
Short bowel syndrome (SBS) is a debilitating condition, typically resulting from extensive bowel resection.1–3 Due to insufficient functional gut to sustain nutritional needs solely on enteral nutrition, parenteral nutrition (PN) is needed for survival in these patients.4–6 PN is the process of providing nutritional needs intravenously, and when all nutritional needs are met via the parenteral route, the process is called total parenteral nutrition (TPN). Although TPN is used world-wide and its benefits remain undisputed, enthusiasm for its use is tempered due to a significant association of adverse effects.7,8 Intestinal failure-associated liver disease (IFALD) is a well-defined progressive cholestatic liver injury occurring in SBS patients on TPN.9,10 Recent data also describe gut atrophic changes in SBS in the absence of enteral nutrition.11,12
Ongoing research has focused on the mechanisms that lead to these adverse effects. In fact, recent literature has explored the role of gut-systemic signaling in the context of TPN injury in SBS.4,13 It has been reported that a key hepatobiliary receptor, namely the gut Farnesoid X Receptor (FXR),14,15 and its modulation by bile acids appear to play an important role in the pathogenesis of liver injury.4,12,16 Data also suggest that there might be a decreased synthesis of the hepato-protective fibroblast growth factor 19 (FGF19) secondary to inadequate gut FXR activation.17,18 It has been postulated that targeting the FXR-FGF19 axis could yield incremental results in the development of ameliorative as well as preventative strategies. Indeed, understanding the signaling pathways contributing to TPN-associated injury in SBS is not only a major research focus in gastroenterology and hepatology, but it could also prove crucial in the care of such patients. 19 In order to bring to fruition these objectives, creating a model system that is replicative of human disease with bowel resection is crucial to interrogate pathways as well as therapeutic testing.
Considering this perplexity, we hypothesized that an ambulatory neonatal piglet model of short bowel syndrome with distal small bowel resection and ileo-colonic anastomosis replicative of the common human SBS could be developed. This would not only be a remarkable step forward in establishing a rigorous model to test SBS therapies but given the significant homology of the liver and gastrointestinal track of neonatal pigs 20 and the human infant, it would also be a platform for future mechanistic studies with rapid translatability to clinical medicine.
Methods and materials
Animal procurement
The United States Department of Agriculture (USDA) recognizes Saint Louis University (SLU) as a registered research facility. The study was conducted upon approval by the Institutional Animal Care and Use Committee of SLU (SLU No. 2657, US Department of Agriculture registration 43-R-011) and in accordance with the Guide for the Care and Use of Laboratory Animals.21,22 Neonatal piglets 7 to 10 days old were used for this study, procured from a University approved class A vendor. Ear tags were used to identify animals.
Acclimatization and housing
Animals were placed in heated cages immediately upon arrival. Three days for acclimatization were provided per university guidelines. The facility veterinarian mandated the temperatures and the animals were kept in a thermally controlled environment for the duration of the study.
Surgery and catheter placement
Ensuing three days of acclimatization, the piglets underwent placement of jugular and duodenal catheters as previously described. 12 Animals in the SBS group underwent distal bowel resection with ileo-colonic anastomosis. Individually, each piglet was taken to the veterinary operating room and placed in a chamber that contained 3% to 5% isoflurane for anesthetic induction. Thereafter, the animal was transferred to a heated surgery table and appropriate anesthesia was maintained using a cone mask (2%–4% isoflurane). Through the entirety of the surgical procedure, vital signs, oxygen saturation by pulse oximeter, temperature, heart rate, and respiratory rate were monitored continuously. Once the animal was deeply anesthetized, the neck and abdomen were surgically prepared and draped for aseptic surgery.
Jugular catheter placement
A vascular cut down technique was used to place catheters into the left and the right jugular veins. Purse string sutures were used to secure the catheter to the vessels and 3 mL heparinized saline was injected to confirm the patency. The catheters were then tunneled subcutaneously to exit the skin just caudal to the scapula, following which, a catheter flange was used to suture the catheter to the skin. Additionally, medical grade silicone glue was applied in order to secure the catheter and minimize its slide.
Duodenal catheterization
A midline, 5-inch abdominal incision was made cranial to the umbilicus. Placement of a catheter into the duodenal lumen was performed as previously published. 12 Once the catheter was sutured and tunneled, we proceeded with distal bowel resection.
Creation of short bowel
The ileocecal valve was identified (Figure 1(a)), and to perform a 75% resection, we measured the bowel using sterile silk ribbon placed along the anti-mesenteric border of the gently mobilized small intestine. We measured in retrograde fashion starting at the ileocecal valve.
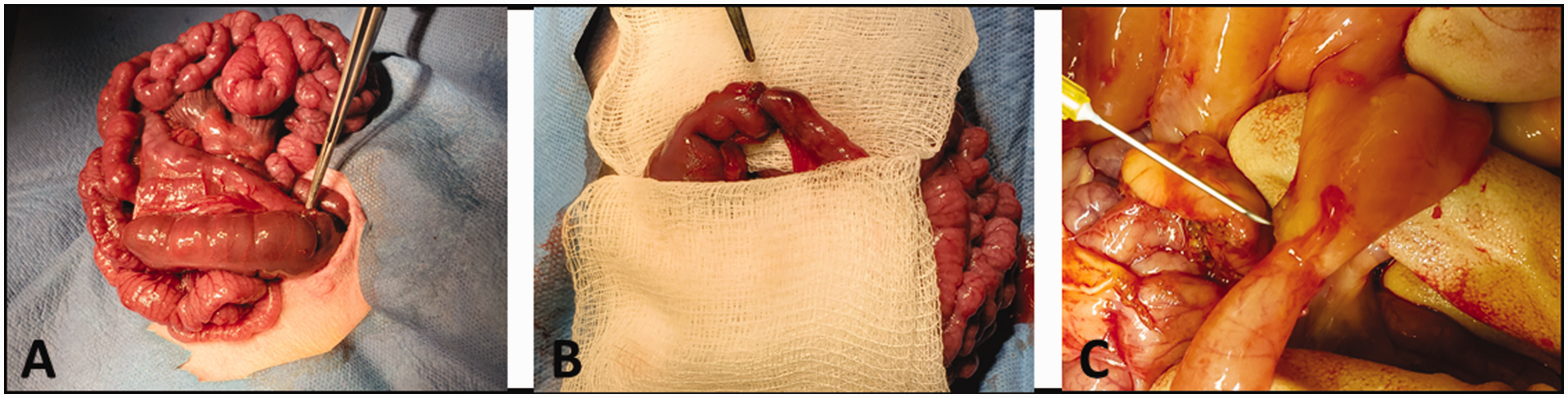
SBS with ileo-colonic anastomosis: (a) Unresected ileo-colonic junction. (b) Electrocautery is used to obliterate the mesentery to accomplish the resection. Noted here is post resection ileo-colonic anastomosis. (c) Intact anastomosis noted at animal sacrifice. (A color version of this figure is available in the online journal.)
We chose 75% bowel resection as this amount of resection is an important criteria to warrant referral of patients in clinical medicine to intestinal rehabilitation programs. 23
Using doyen-clamps, the proximal and distal ends were clamped to prevent gross contamination. The small bowel in between these two segments was resected using electrocautery. We then ligated the terminal ileum proximal to the valve using running 5–0 Vicryl. Once resected, the segment was freed from the mesentery, a small colotomy was made on the cecum, and the proximal segment of small bowel was brought together. Stay sutures at the mesenteric and anti-mesenteric borders were used to ensure correct apposition. Luminal continuity of the two remnant sections was restored by an ileocolonic end to end hand-sewn anastomosis using 5–0 Vicryl sutures (Figure 1(b)).
The anastomosis was copiously washed in a bowl of warm saline before being returned to the abdomen. The abdominal incision was closed in two layers with 2.0 Vicryl (absorbable; all layers of abdominal wall except skin) and staples. Piglets were administered buprenorphine (0.1 mg/kg, subcutaneously) for postoperative pain relief and monitored until full recovery. Animals in the control group underwent “sham” surgery, which involved making the abdominal incision and mobilizing the small bowel; however, no bowel resection was performed. The abdominal incision was closed as described above.
Jacket placement
Animals were fitted into jackets with a pocket on each side. The nutrition bag was placed in the left pocket of the jacket. In the right pocket, a miniaturized battery-operated infusion pump (Orchestra 500) was placed. Both the Jugular and Duodenal catheters ran through the right pocket fabric. One of the jugular catheters was connected to a battery-operated miniaturized pump via a PEGA tube mechanism, while the other was heparin locked. Subsequently, enteral nutrition or TPN (with lipids) in EVA bags (EVA, code 66050; Medtec Medical, IL, USA) was provided. This was connected via clamps to the inflow tubing supplying the pump. The veterinarian staff closely monitored the piglets until they fully recovered.
Animal monitoring
A veterinarian examined the animals and they were weighed each morning. Furthermore, the animals were monitored with scheduled visits by research personnel in accordance with the Institutional Animal Care and Use Committee and the Guide for the Care and Use of Laboratory Animals.
Group allocation and statistical analysis
Randomly, 14 animals were allocated into three different groups (Table 1), after 48 h post-surgery recovery. The enteral nutrition (EN) animal group (n = 5) received enteral nutrition and no TPN. The TPN animal group (n = 5) received no EN but was provided TPN via the jugular catheter. The SBS animal group (n = 4) underwent bowel resection, ileo-colonic anastomosis, and received TPN via the jugular catheter but no EN. Mean and standard deviation were calculated for each group for weight gain, serum bilirubin, and the gut density. The Students’ t-test was used for statistical analysis. All tests were two sided using a statistical significance level of P = 0.05.
Study groups.

EN: enteral nutrition; TPN: total parenteral nutrition; SBS: short bowel syndrome.
Nutrition intake
Isocaloric and isonitrogenous nutrition was given to all animals for approximately two weeks. The EN group received swine replacement formula LitterLife (Merrick’s Inc.) via the duodenal catheter, while the TPN and SBS piglets received parenteral nutrition via the jugular catheter as previously published.8,12,16 TPN or EN was placed in nutrition bags with nutritional constituents as noted in Supplementary Table S1. All nutrition bags were replaced every 12 h via a clean technique to avoid contamination.
Animal euthanasia and sample collection
On day 14, the jugular catheter was assessed, and a blood sample was collected. Then, pentobarbital sodium (100 mg/kg) euthanasia solution (Beuthanasia-D; Schering-Plough Animal Health, Kenilworth, NJ, USA) was given intravenously for euthanasia. Immediately following euthanasia, the abdomen was opened, and the liver was obtained in its entirety. The proximal and distal segments were identified based on the ligament of Treitz, ileocolonic junction, or suture location and removed completely. Resected bowel was preserved. A 20 cm segment from the proximal and distal gut from all groups was immediately flushed with cold saline and weighed.
RNA extraction and real time quantitative PCR analysis
RNA was extracted from gut and liver tissue using Sigma Aldrich GenElute Mammalian Total RNA Miniprep Kit (RTN70-1KT) and Trizol (ThermoFisher, 15596018) respectively, as per the manufacturer protocol. Isolated RNA (1 ng) was reverse transcribed into complementary DNA using the Verso cDNA Synthesis Kit (ThermoFisher, AB1453B). Primers were designed utilizing the Integrated DNA Technologies (IDT) and primer sequences (Table 2) for each of the transcript were validated. Real-time quantitative polymerase chain reaction (RT-qPCR) was subsequently performed in triplicate using the Bio-Rad CFX Connect Real-Time System. Relative mRNA expression was calculated using the comparative threshold cycle method with Beta-Actin and cyclophilin B as an internal control.
Porcine primer sequences.

Results
All catheters remained secured throughout the study. At the time of animal euthanasia, we inspected the bowel anastomosis in all SBS animals, which remained intact (Figure 1(c)).
Weight gain
Adequate of caloric intake is reflected by weight gain. We weighed each animal daily. Daily weight gain did not show any statistically significant differences among the groups, confirming adequate caloric support. The mean daily weight gain ± SE was 130.12 ± 15.23 for EN (P = 0.27 EN vs. TPN), 121.8 ± 5.97 for TPN (P = 0.08 TPN vs. SBS), and 1113.25 ± 16.27 for SBS (P = 0.11 SBS vs. EN) as shown in Figure 2.
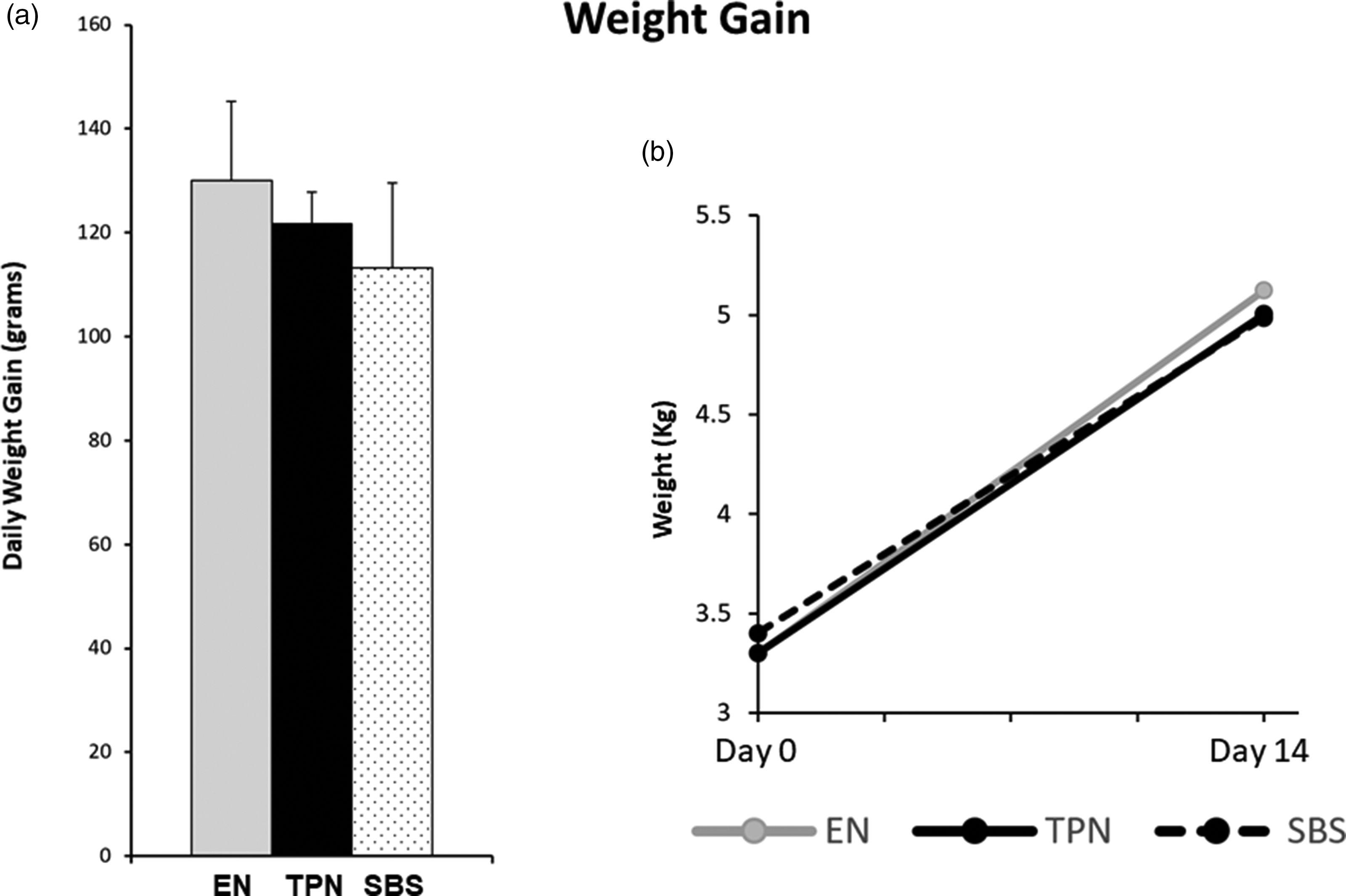
Weight gain: (a) Daily weight gain for animals in each group. (b) Absolute change in weight from Day 0 to Day 14 of the study. Note no differences in weight gain between EN, TPN, or SBS.
Hyperbilirubinemia
An increase in conjugated bilirubin is indicative of cholestasis and is commonly noted in patients with SBS on TPN. We measured the conjugated bilirubin fraction at the end of 14 days, prior to animal euthanasia. Animals on TPN had significantly higher conjugated bilirubin in comparison to EN animals. While the SBS animals also had significant elevation of serum conjugated bilirubin, the levels for SBS were higher than those animals on TPN (without gut resection) as noted in Figure 3(a). The mean conjugated bilirubin ± SE was 0.036 ± 0.004 mg/dL for EN (P = 0.03 EN vs. TPN), 1.29 ± 0.613 mg/dL for TPN (P = 0.01 TPN vs. SBS), and 3.89 ± 0.51 mg/dL for SBS (P = 0.000064 SBS vs. EN).
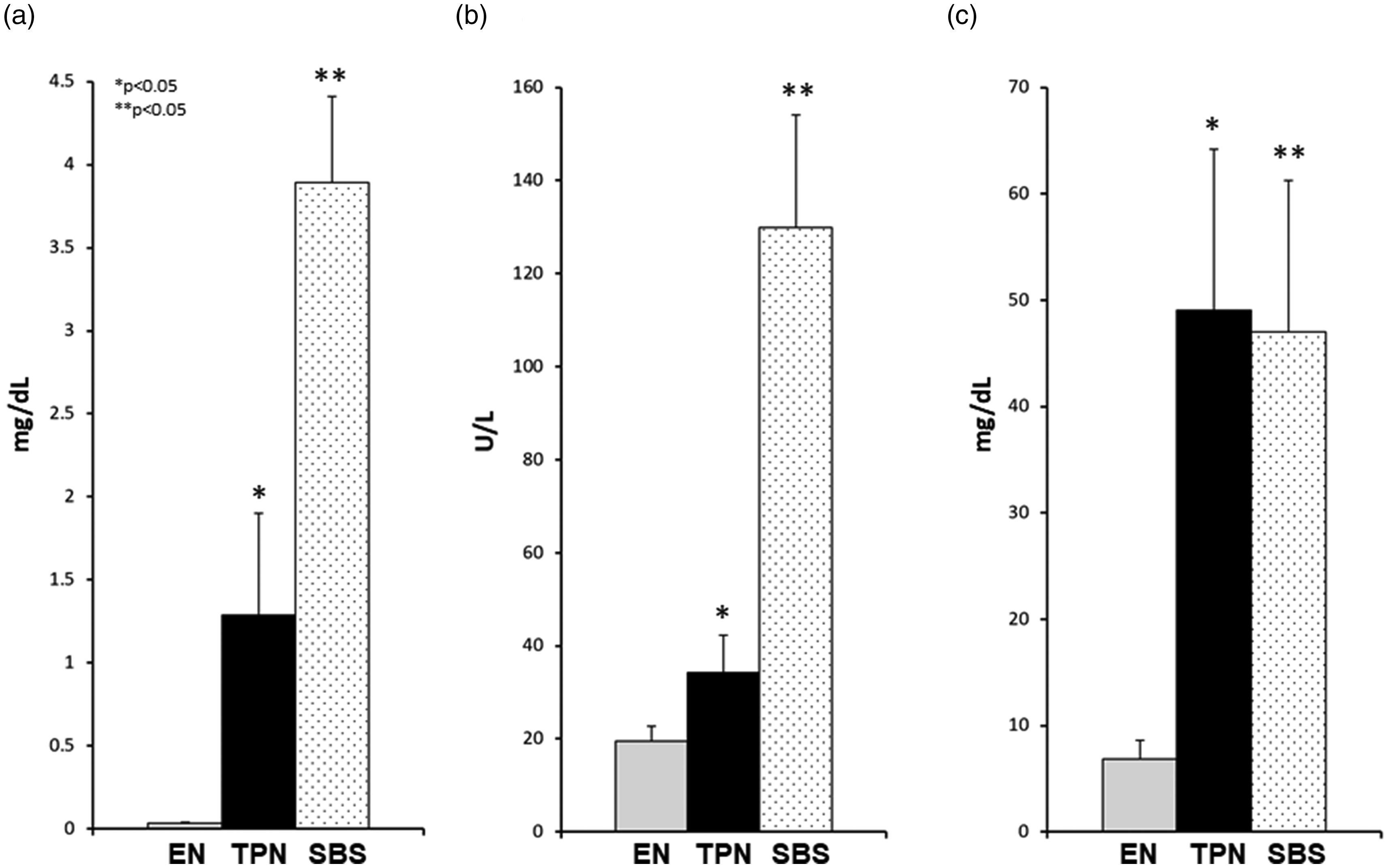
Serum markers: (a) Conjugated bilirubin, (b) GGT, (c) bile acids. Note: Significant increase in serum bilirubin, GGT, and bile acids in TPN and SBS groups vs. EN. Both bilirubin and GGT was also higher in SBS vs. TPN. Each column reflects the mean for that group. Error bars represent standard error. Differences between groups are based on the T test. All tests were two-sided using a significance level of 0.05.
GGT, bile acids
Another marker noted in SBS patients is an elevation in serum gamma glutamyl transferase (GGT) and serum bile acids. We noted a significantly elevated GGT (Figure 3(b)) and bile acids (Figure 3(c)) in animals on TPN as well as those with SBS in comparison to EN controls. Statistically, GGT was also higher in SBS vs. TPN animals (Figure 3(b)). The mean GGT and bile acids ± SE was 19.56 ± 3.12 U/L and 6.84 ± 1.80 mg/dL for EN (P = 0.03; P = 0.02 EN vs. TPN), 34.28 ± 8.01 U/L and 49.04 ± 15.11 mg/dL for TPN (P = 0.0004; P = 0.02 TPN vs. SBS), 129.97 ± 24.11 U/L and 47.02 ± 14.22 mg/dL for SBS (P = 0.001; P = 0.015 SBS vs. EN), respectively.
We also evaluated liver transaminases. No statistical differences in serum alanine aminotransferase (ALT) (P = 0.22) or serum aspartate aminotransferase (AST) (P = 0.44) were noted among the groups. We believe that longer duration of TPN would enhance these differences to reach statistical significance. While this remains an unusual finding, such has been reported by us before.8,12,16
Linear gut density and villous to crypt ratio
We have postulated gut mucosal atrophy with TPN. To objectively quantify the gut atrophic changes, we determined the linear gut density of the gut as measured by the weight in grams per centimeter of bowel length. We noted a significant reduction in linear gut density in SBS and TPN animals in comparison to EN controls in both proximal (Figure 4(a)) and distal (Figure 4(b)) gut segments. Additionally, while there was no statistical difference in the proximal linear gut density between TPN and SBS, we noted that the linear gut density was statistically lower in SBS for the distal segment. The mean linear gut density ± SE for proximal and distal gut was respectively 0.20 ± 0.01 gm/cm and 0.30 ± 0.02 gm/cm for EN (P = 0.0007; P = 0.0005 EN vs. TPN), 0.11 ± 0.02 gm/cm and 0.16 ± 0.01 gm/cm for TPN (P = 0.07; P = 0.01 TPN vs. SBS), 0.14 ± 0.01 gm/cm and 0.11 ± 0.008 gm/cm for SBS (P = 0.02; P = 0.0001 SBS vs. EN).
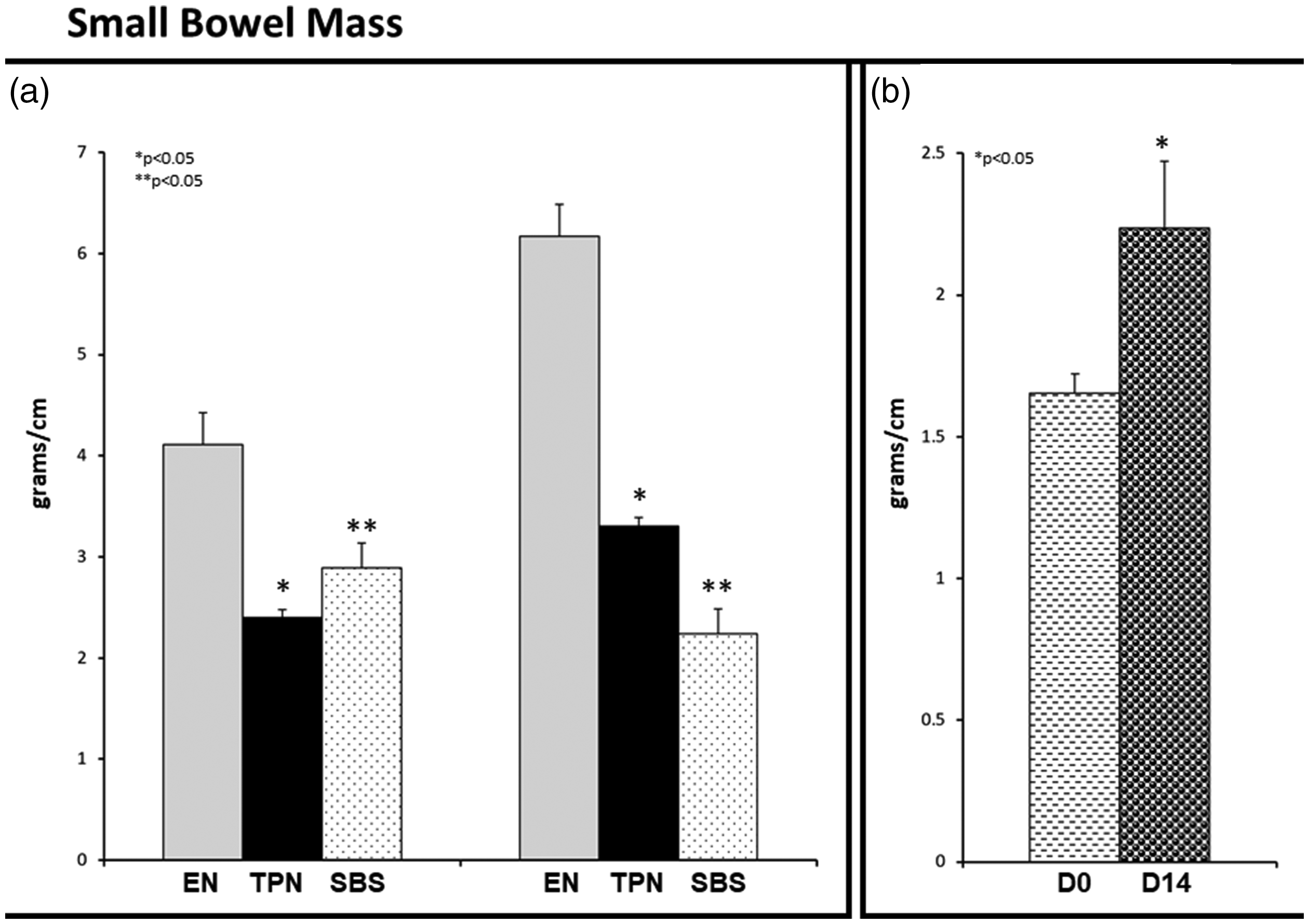
Linear gut density: (a) Proximal and distal gut segments. Note: Significant reduction in proximal and distal linear gut density (g/cm) in TPN and SBS group vs. EN. SBS linear gut density was higher than TPN for the proximal bowel. (b) Gut adaptation in SBS. Note that the D0 segment (proximal end of the resection) adapted over 14 days – D14 in SBS animals. Each column reflects the mean for that group. Error bars represent standard error. Differences between groups are based on the T test. All tests were two-sided using a significance level of 0.05.
It is important to understand that the distal segment in SBS at animal sacrifice (D14) represented the proximal end of the resection (D0) during the SBS surgery. In order to quantify adaptation 24 in this segment, we compared D0 against the distal segment D14 obtained after the 14 days of the experiment at animal sacrifice. We noted that the linear gut density for the D14 segment was statistically higher than that for D0 (P = 0.015), indicating adaptive responses (Figure 4(c)).
Severe villous atrophy was noted in animals on TPN and SBS in comparison to EN animals as shown in Figure 5 (P = 0.028). To quantify morphometric differences, we evaluated in each group the villous/crypt ratio (V/C). The mean V/C ratio ± SE for distal and proximal segment was EN 4.27 ± 0.14; 3.81 ± 0.23, for TPN 1.86 ± 0.11; 1.73 ± 0.09, and SBS 2.12 ± 0.07; 1.92 ± 0.17.
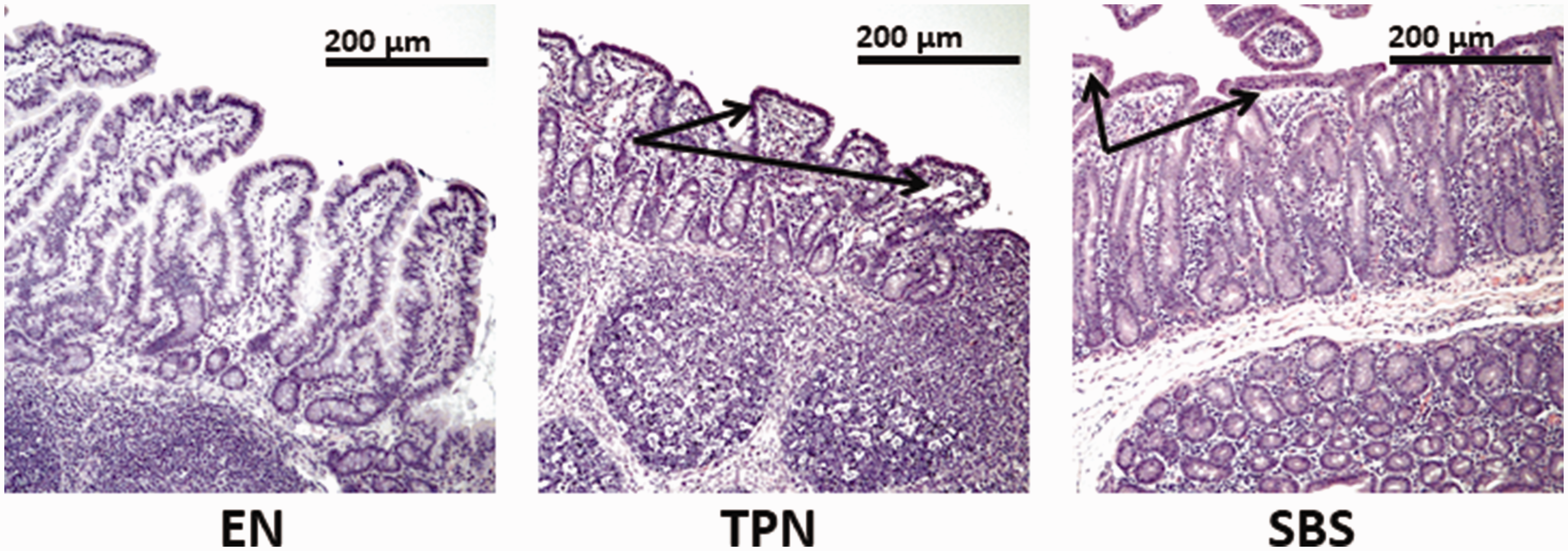
Small bowel histology (200× magnification): EN, TPN, and SBS groups. Note villous atrophy (arrows) in TPN and SBS animals in comparison to EN animals. (A color version of this figure is available in the online journal.)
Messenger RNA expression
It has been reported that there is decreased activation of gut FXR during TPN, with a resulting decrease in gut FGF19. 8 Mechanistically, this pathway regulates hepatic bile acid synthesis via regulation of cholesterol 7 alpha-hydroxylase (CyP7A1). While mRNA levels do not represent protein levels, mRNA fold change can highlight changes in gene expression of key hepato-biliary receptors.
Thus, in order to quantify mRNA expression of these major hepato-biliary receptors, which have a predominance in the distal segments, we performed quantitative reverse transcription (qPCR) on the bowel segment in each of the study groups (Figure 6). When comparing fold change against the EN group, there was almost a 50% reduction in gut FXR expression in both TPN and SBS groups (P = 0.0006, P = 0.007, respectively). Gut FGF19 was over 2-fold lower for both TPN and SBS (P = 0.0001, P = 0.038, respectively). No statistical differences were noted between SBS and TPN with either gut FXR (P = 0.91) or gut FGF19 (P = 0.95). CyP7A1 activity was almost double with TPN and SBS against EN controls (P = 0.002, P = 0.047, respectively). Since CyP7A1 is the rate limiting step for bile acid synthesis, this un-inhibited CyP7A1 activity was in line with the significantly higher bile acid levels noted in both TPN and SBS animals.
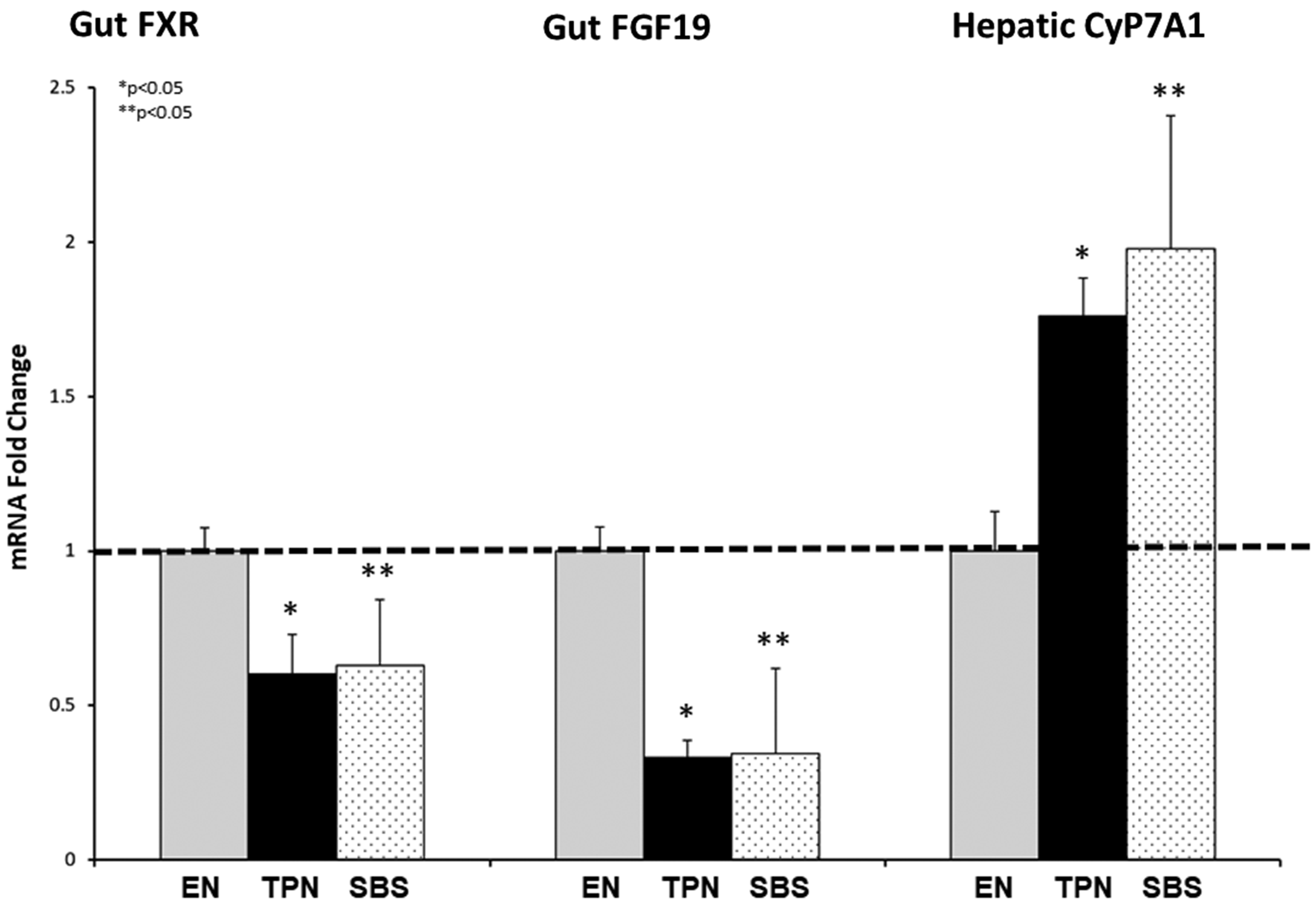
(a) mRNA expression for gut FXR, gut FGF19, and Hepatic CyP7A1. Note a significant fold reduction in both gut FXR and gut FGF19 in the TPN and SBS group vs. the EN group. CyP7A1 was much higher in TPN and SBS against the EN controls. Each column reflects the mean for that group. Error bars represent standard error. Differences between groups are based on the T test. All tests were two-sided using a significance level of 0.05.
We also set out to measure plasma 7α-hydroxy-4-cholesten-3-one (C4) levels which is known to correlate with CYP7A1 enzymatic activity and could serve as a biomarker for bile acid synthesis. C4 is an oxidative enzymatic product of cholesterol metabolism via cholesterol 7α-hydroxylase, an enzyme also known as cholesterol 7-alpha-monooxygenase. C4 is a stable intermediate in the rate-limiting pathway of bile acid biosynthesis.25,26 In line with the CyP7A1 mRNA expression results, we noted a significant increase in C4 activity for both TPN and SBS animals (P = 0.026).
Discussion
Short bowel syndrome is a complex condition in which a loss of functional bowel results in impaired digestive and absorptive processes. Although there are rare cases of congenital short bowel, SBS is typically a result of extensive small bowel resection resulting in decreased mucosal surface area. 27 There are several factors contributing to the overall prognosis in individuals with SBS, with the length of the remaining functional small bowel playing a major role.28,29 The absence of the of the ileocecal valve (IVC) is also a major driver of outcomes.2,9
Due to the inability to absorb needed nutrition and fluids for survival, TPN 30 has become an integral part in the management of patients with intestinal failure and SBS. Although TPN can be life-sustaining and lifesaving, it does not come without risks.11,31,32 Some complications include intestinal failure-associated liver disease (IFALD) and gut mucosal atrophy for which there has been significant research focused on developing preventative and ameliorative therapies. 10
In order to better study the mechanisms driving TPN-associated hepatic and intestinal injury, as well as to develop therapies for SBS, establishment of an SBS animal model is imperative; however, a major limitation in moving the field forward is the absence of a robust SBS animal model replicating human ambulatory TPN infusion, a critical step for translational studies.
We hypothesized that an ambulatory SBS piglet model could be developed using miniaturized pumps carried by an animal. Thus, as part of this study, we developed surgical techniques to resect distal bowel in newborn piglets and achieve end-to-end anastomosis, while bypassing the ileocecal valve to mimic SBS patients with distal bowel and ICV resection. Our model also entailed surgical placement of intra-duodenal as well as jugular catheters tunneled subcutaneously to exit dorsally with minimal complications, allowing for improved mobility of animals. Furthermore, we were successfully able to deliver nutrition to these animals, using ambulatory pumps. With our model, equal weight gain in short bowel animals compared to animals without bowel resection on TPN and those on enteral nutrition was demonstrated. This indicates delivery of adequate nutrition to all groups.
As seen in human patients with SBS, we noted significantly higher conjugated bilirubin, GGT, and bile acid levels in animals on TPN, reiterating the cholestatic nature of TPN-associated liver injury. Interestingly, both conjugated bilirubin and GGT levels were also significantly higher in SBS animals when compared to animals on TPN with intact bowel. This reflects the complex nature of IFALD and may indicate that distal bowel and ICV resection may themselves lead to exacerbated cholestatic injury or that the functional length of the remaining bowel is a co-relate of serum bilirubin? These questions are part of ongoing research, made possible by such a model system.
Several recent studies also point to atrophic changes in the gut while on TPN. This is believed to occur due to the lack of trophic signals secondary to a lack of luminal nutrients during TPN. While animals in our study on TPN with intact gut had atrophic changes as seen by a reduction in the linear gut density in g/cm, we found that the proximal linear gut density in animals with SBS was higher. We postulate that this response is secondary to gut adaptation in response to the distal bowel resection. Elucidating mechanisms driving such gut changes remains an important research area. In SBS animals, another interesting observation was a significant gut adaptation after 14 days as noted in the difference in linear gut density in the D0 vs. D14 segments. A possible explanation other than purely adaptive changes could be a natural increase in linear gut density as the animals’ age and increase in body weight. Future studies assessing the respective segments of bowel from non-SBS animals receiving EN or TPN can provide further comparative rigor. Nevertheless, further research into such adaptive responses continues to challenge the established paradigms and will drive the field forward.
An important use of our model would be therapeutic drug testing. We postulate positive impact of gut TGR5 agonists like oleanolic acid16,33 in modulating linear gut density as well as the villous/crypt ratio in SBS based on our prior studies using these drugs in animals with intact gut. We have also evaluated the role of subcutaneously delivered epidermal growth factor34,35 (EGF), which is a known gut promoter, at sub-therapeutic levels, as proof of concept in SBS animals. At such doses while no difference in gut growth was noted (n = 2), we also did not note adverse effects, enabling the design of future experiments with physiological dose testing. An important focus for future studies is also to evaluate changes in gut integrity and gut translocation of microbiota with adaptation inducers as well as the role of the gut microbes in modulating disease in SBS.
We also noted a decrease in FXR and FGF19 36 expression in the gut in both TPN and SBS in comparison to EN. This is important as it validates prior publication documenting a decrease in luminal activation of key hepatobiliary receptors in patients on TPN. 8 In line with these results, we noted a significant elevation in CyP7A1 activity. Thus, targeting downstream molecules 37 released secondary to FXR activation or inhibitors of CyP7A1 may hold great promise in mitigating liver disease in SBS which could be tested using this model system.
Our SBS model also serves as an important tool in further defining mechanistic links mediating gut to systemic crosstalk. Given its ambulatory nature, our SBS model poses minimal animal stress in comparison to tethered animal models 38 and thus long-term effects of TPN infusion in SBS as occurring in humans could be ascertained and rigorously tested.
This study demonstrated that the establishment of an ambulatory SBS model with ileocolonic anastomosis on TPN is feasible. This system, by recapitulating the human disease could prove to be a key tool in interrogating intricate mechanistic pathways, develop therapeutics as well as diagnostics, and ultimately help mitigate unfortunate side effects from a live-saving therapy for SBS patients.
Conclusion
We successfully created an animal model of short bowel syndrome with ileo-colonic anastomosis using neonatal piglets with TPN delivered in an ambulatory manner using miniaturized pumps. We strongly believe that this model mimics ambulatory TPN delivery in human SBS patients. Our model with significant distal bowel resection along with TPN dependency had hyperbilirubinemia, compensatory gut changes, as well as alteration in hepatobiliary receptors. This is well in line with key elements noted in human short bowel syndrome and thus holds high potential for translational research supporting drug development, diagnostic testing, and in better understanding of the mechanisms in SBS leading to adverse side effects.
Supplemental Material
EBM915881 Supplemental Material - Supplemental material for Development and validation of an ambulatory piglet model for short bowel syndrome with ileo-colonic anastomosis
Supplemental material, EBM915881 Supplemental Material for Development and validation of an ambulatory piglet model for short bowel syndrome with ileo-colonic anastomosis by Chandrashekhara Manithody, Christine Denton, Amber Price, Keith Blomenkamp, Yogi Patel, Adam Welu, Ester Glbert, Himani Madnawat, Sonali Jain, Gustavo A Villalona and Ajay K Jain in Experimental Biology and Medicine
Footnotes
Authors’ contributions
AKJ contributed to the conception/design of the research. All authors contributed to acquisition, analysis, or interpretation of data, as well as drafting/editing the manuscript. AKJ, CM, CD, and AP conducted the experiment. KB, YP, AW, EG, SJ, GAV and HM assisted with the experiments. SJ and GAV performed animal surgeries. All authors agree to be fully accountable for ensuring the integrity and accuracy of the work and approved the final manuscript.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: AKJ serves as a consultant and speaker for Alexion Pharmaceuticals; however, this association is not relevant to the current manuscript. The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
FUNDING
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work was supported by the National Institutes of Health [grants numbers K08DK098623, R03EB015955-01, and R13DK109671-01], internal funding through Saint Louis University [Fleur-de-lis grant], and the DeNardo Foundation.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.