
Abstract
Kidney dysfunction, including chronic kidney disease and acute kidney injury, is a globally prevalent health problem. However, treatment regimens are still lacking, especially for conditions involving kidney fibrosis. Stem cells hold great promise in the treatment of chronic kidney disease and acute kidney injury, but success has been hampered by insufficient incorporation of the stem cells in the injured kidney. Thus, new approaches for the restoration of kidney function after acute or chronic injury have been explored. Recently, kidney organoids have emerged as a useful tool in the treatment of kidney diseases. In this review, we discuss the mechanisms and approaches of cell therapy in acute kidney injury and chronic kidney disease, including diabetic kidney disease and lupus nephritis. We also summarize the potential applications of kidney organoids in the treatment of kidney diseases.
Impact statement
Stem cells hold great promise in regenerative medicine. Pluripotent stem cells have been differentiated into kidney organoids to understand human kidney development and to dissect renal disease mechanisms. Meanwhile, recent studies have explored the treatment of kidney diseases using a variety of cells, including mesenchymal stem cells and renal derivatives. This mini-review discusses the diverse mechanisms underlying current renal disease treatment via stem cell therapy. We postulate that clinical applications of stem cell therapy for kidney diseases can be readily achieved in the near future.
Introduction
Kidney diseases are prevalent all over the world, with chronic kidney disease (CKD) 1 comprising more than 10% of kidney disease diagnoses, and acute kidney disease (AKI) a wide range between 1% and 66% in different regions globally. 2 In the case of AKI, approximately 41% of patients not recovering renal function before hospital discharge will progress into CKD. 3 Treatment regimens for CKD and AKI are currently limited. Current efforts have focused on improving outcomes and reducing comorbidities for both CKD and AKI.
Stem cells are a class of cells that maintain the ability to self-renew and differentiate into multiple cell lineages. They can be divided into three categories: (a) pluripotent stem cells (PSCs) such as embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs); (b) multipotent stem cells with partial differentiation ability; and c) unipotent stem cells, which can only differentiate into one kind of cell lineage. PSCs have been differentiated into kidney organoids, 4 which are useful for kidney functional replacements in vivo and in vitro. Multipotent stromal cells such as mesenchymal stem cells (MSCs) that are derived from organs or tissues, such as bone marrow (BM-MSCs), amniotic fluid (AFSCs), urine (USCs) and umbilical cord (UC-MSCs), are used to treat kidney disease through mechanisms including paracrine signaling, initiation of autophagy, anti-apoptotic, or anti-inflammatory effects. Thus, stem cell treatment is a very promising therapy for kidney diseases. In this article, we review the mechanisms underlying the effects of stem cells on different kidney diseases (Figure 1, Table 1 and Table 2). We also summarize the progress in the development of kidney organoids for research and clinical application.
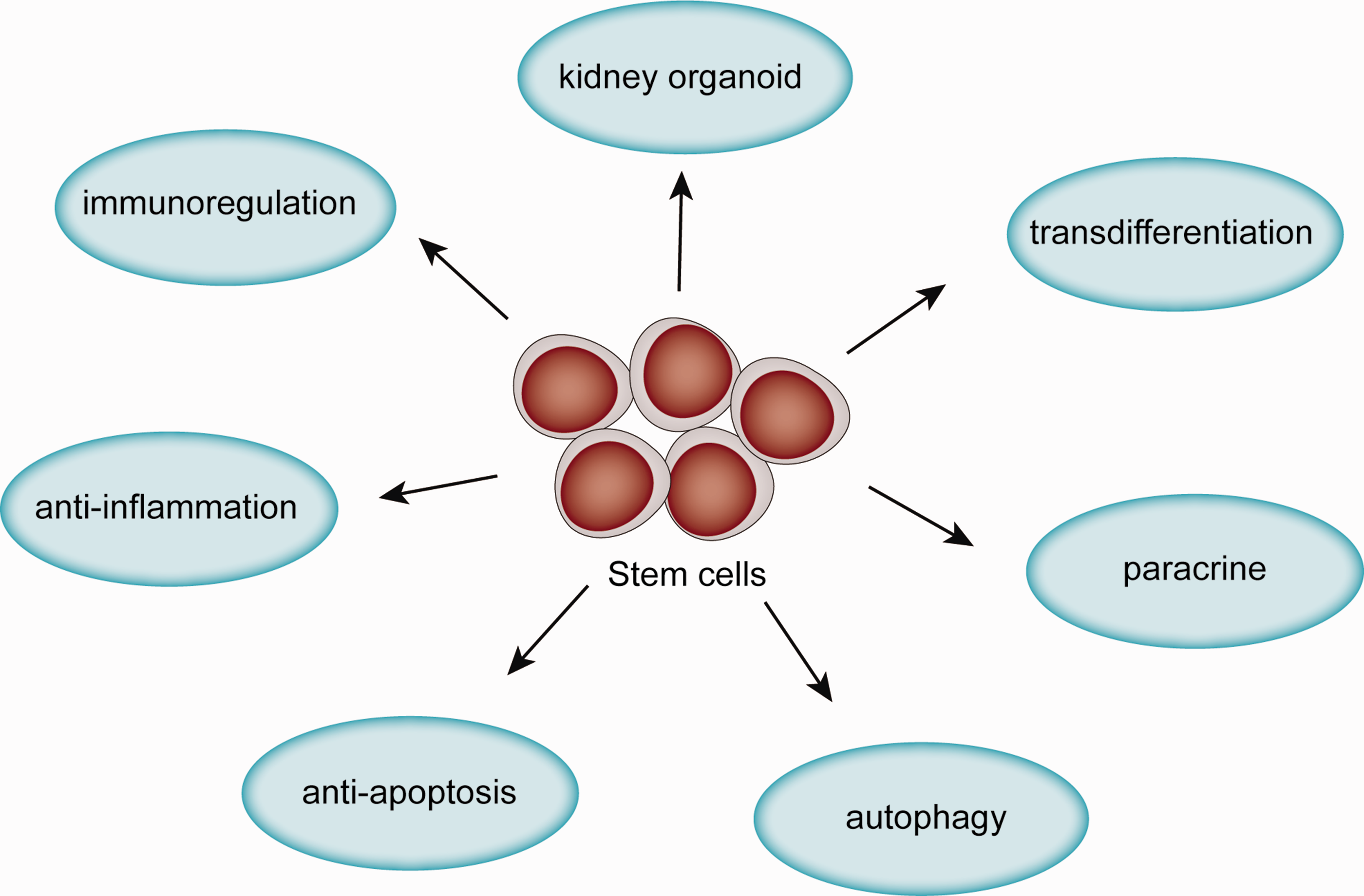
Mechanisms of stem cells in treatment of kidney disease.(A color version of this figure is available in the online journal.)
Sources of stem cells and their roles in the treatment of kidney diseases.
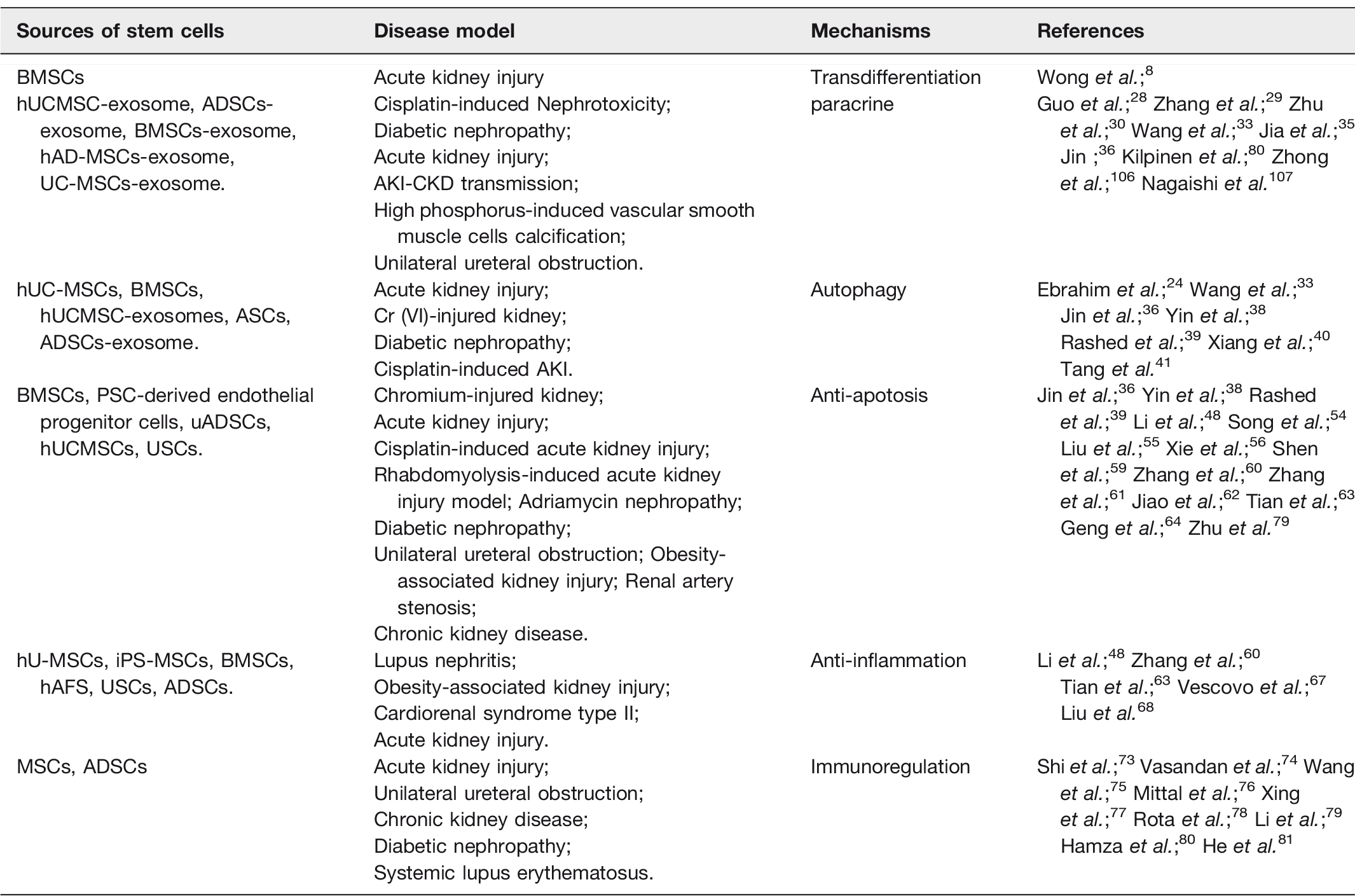
Application of mesenchymal stem cells and exosomes in the treatment of kidney diseases.

Note: Clinical use of stem cells has focused on applications for lupus nephritis 100 and monoclonal immunoglobulin-related renal disease, 101 including AL amyloidosis, light chain deposition disease, heavy chain deposition disease, and myeloma-related renal disease. Because of immunogenicity, autologous peripheral blood stem cells are used in treatment. Study has shown using stem cells could improve remission rate and patient survival in monoclonal immunoglobulin-related renal disease. 102 Studies on CKD have shown that infusing BMSCs or autologous CD34+ cells was safe and tolerable for CKD patients, with no significant change of kidney function.103,104 However, in AKI after cardiac surgery, using allogeneic MSCs did not improve the time to kidney function recovery, indicating the mechanisms of action behind the use of stem cell treatments in AKI patients have yet to be fully elucidated.
Differentiation of stem cells into functional kidney cells
Kidney cells contribute to many different physiological functions. For example, podocyte processes prevent proteinuria and tubule cells reabsorb filtrated materials. Damage to these cells induces proteinuria or kidney dysfunction.5–7 Studies have shown that culturing MSCs with injured mesangial cells induced by hydrogen peroxide in vitro allowed MSCs to differentiate into mesangial cells, 8 and that MSCs injected into the suprarenal aorta could trans-differentiate into podocyte-like cells in a puromycin aminonucleoside induced acute proteinuria model in rat. 9 However, stem cells have not been shown to trans-differentiate into tubule cells. 10 Using iPSCs in adriamycin nephropathy revealed labeled-iPSCs were trapped in the lungs, 11 and very few donor cells were observed in the kidney.10,12–14 This was attributed to the fact that only few stem cells infused into the soma could home into the injured areas during kidney injury and multiple reasons may contribute to these challenges.15–17 First, detrimental environment of stem cells may affect their differentiation abilities. Second, targeting stem cells to the location of injury is a major challenge in clinical applications. Third, the internal environment in AKI and CKD, including oxidative stress, inflammation, uremic toxin, hypoxia and other factors, can influence stem cell vitality.
Extracellular vesicles (EVs) from stem cells: Paracrine effects
EVs or exosomes in the size of approximately 40–200 nm can be secreted from endosomal compartments by most cell types and have been reported to be involved in the cell-to-cell communication and delivery of biomolecules including proteins, DNA, messenger RNAs (mRNAs), microRNAs (miRNAs) and lipids to recipient cells. 18 Exosomes from MSCs have been reported to be a new regimen for AKI and CKD. 19 Treatment with EVs via caudal vein after reperfusion could improve cortical microvascular and peritubular capillary density as well as the expression of angiogenic factors, such as vascular endothelial growth factor (VEGF) 20 and hepatocyte growth factor. 21 VEGF subsequently activated Notch1 22 in human arterial endothelial cells to trigger arteriogenesis and angiogenesis, 23 which improves microcirculation through more efficient delivery of oxygen. As a result, oxidative stress and apoptosis were attenuated, resulting in better renal function and histological conditions.24–27
Furthermore, EVs exhibited anti-calcification and anti-fibrosis characteristics. In the end stage renal disease, vascular calcification is commonly attributed to high phosphorus-induced vascular smooth muscle cell calcification which lacks of effective treatment, while the disease could induce sudden death, increase fragility of blood vessels and remote ischemia. It has been reported that EVs could secret miRNAs to activate Wnt, mammalian target of the rapamycin (mTOR), and mitogen-activated protein kinase pathways to inhibit vascular calcification. 28 Therefore, using EVs to ameliorate the calcification would be a promising method in end stage renal disease patients. In addition, kidney fibrosis is a final common pathology of CKD attributed to hypoxia and ischemia. In the fibrosis model of unilateral ureteral obstruction, miRNA-26a inhibited the levels of connective tissue growth factor and transforming growth factor-β (TGF-β1). 29 Also, exosomes from MSCs activate tubular Sox9 to prevent the transition of tubular epithelium cells into a pro-fibrotic phenotype induced by TGF-β1 30 ; however, this transition model has been challenged 31 and whether EVs can inhibit kidney fibrosis needs to be validated.
The effect of stem cells on autophagy
Autophagy is a self-clearance process in which excess or damaged organelles are selectively degraded mediated by autophagosome. There are usually three types of autophagy: macroautophagy, microautophagy, and chaperone-mediated autophagy. Mutation of autophagy-related protein, ATG5 or ATG7, resulted in mild dysfunction of podocytes and tubules within two months and severe glomerular and tubular changes by four months, and then organ failure by six months. 32 Similar changes were observed in human idiopathic focal segmental glomerular sclerosis kidney biopsy specimens, 32 suggesting that dysregulated autophagy could influence kidney structure. Transmission electron microscopy has revealed large amounts of autophagosomes in MSC-exosome-treated rat kidney epithelial cells, 33 indicating autophagy was activated marked by elevated expressions of LC3B. 34 The mechanisms of activating autophagy include: a. Increasing autophagy proteins ATG5 and ATG7 to activate the mTOR pathway 34 ; b. 14–3-3ζ protein contained in MSC-exosomes interacts with ATG16L to promote the localization of ATG16L at autophagosome precursors, resulting in activation of autophagy 35 ; c. Secreting miRNA-486 inhibits mTOR activation, resulting in the increase of autophagy and the reduction of podocyte apoptosis via decreasing smad1. 36 But whether autophagy improves or perturbs kidney function in AKI and CKD remains unclear. It is known that rapamycin can inhibit mTOR pathway to activate mitophagy. In ischemia and cisplatin-induced kidney injury, rapamycin did not enhance autophagy or ameliorate kidney injury although it blocks the mTOR pathway. 37 In parallel, BMSCs inhibited mitophagy, manifesting as decreased Beclin1, PINK1, Parkin, p-Parkin, and LC3B, to restore chromium-injured kidney via upregulating phosphorylation of extracellular signal-regulated kinases and downregulating phosphorylation of p38 and c-Jun-N-terminal kinase. 38 On top of that, decreased beclin1 was also observed in diabetic nephropathy rats treated with MSCs. 39 In contrast, human UC-MSCs could secrete miRNA-145 to induce HK-2 cellular autophagy via inhibition of the phosphoinositide-3-kinase (PI3K)/AKT/mTOR signaling pathway. 40 Moreover, LC3B and ATG5 as well as ATG7 were increased in cisplatin-induced AKI rats after receiving the treatment of MSC-exosomes, 33 suggesting autophagy was activated to preserve kidney function after injury. This kind of protection effect is not only on cisplatin-induced AKI but also in diabetic nephropathy model. It has been shown that glucose metabolism and kidney fibrosis could be improved 41 by using ASCs via decreasing mTOR pathway.
Based on these studies, autophagy should be an important mechanism correlating with different kidney injury models. Previous studies show that MSC delivery via peripheral veins results in decreased autophagy,38,39 while MSCs via kidney injection 33 or subrenal capsule increase autophagy. 41 Although peripheral delivery of MSCs also show some labeled stem cells homing into kidney tissue, it is still unclear why different injecting methods influence the activity of autophagy in injured kidney.
The anti-apoptosis effects on AKI models
The initiation of apoptosis is induced by BAK and BAX 42 via mitochondrial outer membrane permeabilization, 43 which subsequently releases pro-apoptotic protein, cytochrome c, which activates caspase-9 protease in the cytosol. 44 Thereafter, caspase-9 activates caspase-3,-6,-7 to induce the cascade that culminates in apoptotic cell death. Apoptosis is a common pathophysiological process in AKI. 45 It has been shown that intraluminal debris appeared after AKI, 45 which are apoptotic tubule cells induced by oxidative stress and endoplasmic reticulum stress (ER). Oxidative stress can induce mitochondria to transmit apoptotic signals via release of cytochrome c into cytoplasm. 46 ER could induce unfolded protein response (UPR). Mildly enhanced UPR protects against glomerular injury, 47 while severe ER stress results in overwhelming UPR and UPR-related apoptosis emerges.48,49 Studies have shown using MSCs could inhibit oxidative stress50–56 and ER stress57,58 to attenuate apoptosis. Also, previous studies have revealed the effect of MSCs on anti-apoptosis in different kidney injury models to improve kidney outcomes.36,38,59–64 In the chromium-injured kidney, BMSCs treatment revealed decreased apoptosis-related proteins Bax, cytochrome c, and caspase-3, 38 which were also observed in ischemia AKI after dealing with PSC-derived endothelial progenitor cells 59 via decreasing indoxyl sulfate and interleukin-1β (IL-1β). 63 Furthermore, this kind of anti-apoptosis could be enhanced by resveratrol to promote MSCs to secrete platelet-derived growth factor-DD, activating extracellular signal-regulated kinases, which inhibit renal tubular cells apoptosis. 61 In addition, cisplatin-induced nephropathy also exhibits reduced tubular apoptosis after treating with BMSCs through activating Wnt/β-catenin pathway. 62 Wnt/β-catenin pathway has also been defined to induce kidney fibrosis. 65 Therefore, the effect of MSCs on kidney fibrosis needs to be validated. Rhabdomyolysis-induced AKI model is another kidney injury model, manifesting as muscle pain and hematuria as well as kidney dysfunction requiring dialysis. Collagen graft packed with MSCs has been reported to ameliorate outcome of rhabdomyolysis-induced AKI via activating PI3K/Akt pathway to inhibit apoptosis. 64 Delivery of MSCs via biological membrane provides a useful tool in the treatment of AKI and CKD. Potentially, it would be more useful if biological membrane can be made into small capsule to deliver MSCs as subrenal capsule.
Anti-inflammation and immunoregulation
Aseptic inflammation, a common pathophysiology process in AKI and CKD, is induced by multiple signaling pathways that are activated by the binding of ligands, termed “damage-associated molecular patterns,” to toll-like receptors. These pathways include the nuclear factor kappa B, mitogen-activated protein kinase, and type I interferon pathways. 66 The activated inflammatory pathways release IL-1β, tumor necrosis factor-α (TNF-α), interferon-γ to recruit chemokines resulting the migration of mononuclear leucocytes into the injured areas inducing local inflammation. Inflammation can result from different pathogenesis. Ischemia kidney injury could induce hypoxia and increase release of inflammatory factors by necrotic proximal tubule cells, which could be reversed by treatment with USCs, 63 adipose-derived mesenchymal stem cells (ADSCs), 60 and human AFSCs 67 via decreasing the mRNA expression levels of interferon-γ and IL-1β and increasing IL-10 and TGF-β1, resulting in decreased lumen expansion and loss of tubular epithelial cells. 63 Another common etiology of CKD, lupus nephritis, induces kidney injury by immune system activation resulting in elevation of inflammatory markers which could also be inhibited by human UC-MSCs via suppressing nuclear factor kappa B pathway to downregulate the expression of TNF-α, intercellular cell adhesion molecule-1, and plasminogen activator inhibitor-1. 68 Besides the classical kidney injury model, there are still chronic infiltration inflammation in kidney, such as obesity-associated kidney injury manifesting as elevated inflammation markers of IL-6, chemokine (C-X-C motif) ligand 1 and 2 which were also mitigated after treatment with MSCs. 48
During ischemia–reperfusion injury, T cells accumulation in kidney tissues is observed to reside in the infarction boundary zone for about 14 days. The CD4+ and CD8+ T cells have a detrimental role in the kidneys after ischemia–reperfusion injury. 69 In contrast, T-regulatory cells exhibit a protective role in ischemia and reperfusion by secreting IL-10 to reduce the ischemia–reperfusion injury. 70 On top of that, plenty of innate immune cells, including mast cells, neutrophils, macrophages, myeloid-derived suppressor cells, dendritic cells and natural killer cells are engaged in ischemia–reperfusion injury regulated by MSCs. 71 MSCs can secret prostaglandin E2, 72 kynurenic acid, 73 TNF-stimulated gene 6 74 to promote macrophage polarization from an M1 phenotype towards an M2 phenotype to ease inflammatory status. MSCs also inhibited the infiltration of macrophages in kidney tissue.75,76 In addition, MSCs inhibit lipopolysaccharide-stimulated rat peritoneal macrophages by downregulating inflammation-related cytokines such as IL-6, TNF-α, IL-1β, 77 and IL-8 78 to prevent diabetic nephropathy. Moreover, decreasing IL-17 and increasing CD4+CD25+Foxp3+Tregs by ADSCs 79 or EVs 80 could relieve immune inflammation.
Kidney organoids open a new avenue for stem cell therapy
The kidney organoids are developed from ESCs or iPSCs,81,82 and transcriptional congruence of nephron cell type-specific markers, as well as stromal and endothelial markers were similar to human fetal kidney. 83 The induced organoids begin from the primitive streak, induced by Wnt pathway agonist, CHIR99021, to intermediate mesoderm. 84 Thereafter, ureteric epithelium, metanephric mesenchyme, progenitors of renal interstitium, and endothelium are induced from intermediate mesoderm by using a transwell filter. The above progenitors then aggregate at an air-media interface.4,85 After about 20 days, kidney organoids can be obtained. During this procession, exosomes from stem cells are involved in the impaction of kidney organoids by entering the cytoplasm and nucleus via fluorescently labeled exosomal RNA. 86 Kidney organoids contain all kinds of kidney cell lineages. However, lacking of capillary loops suggests that kidney organoids may be insufficient to filtrate urine yet. 87 It has been reported that modulating Wnt pathway activity could generate capillary loops that is adding CHIR99021 at different time period. 88 Although a vascular network is developed, no significant filtration ability was still observed by the organoids.88,89 There should be other factors or microenvironment influencing the function of kidney organoids. Therefore, the kidney organoids were transplanted into vivo resulting functionally vascular filtration,88,89 suggesting a soft environment promotes the maturation of kidney organoids.88,90 Likewise, high fluidic shear stress also revealed generation of capillary loops using a millifluidic chips in vitro. 87 This study indicates a future direction to combine kidney organoids and dialyzer together to use fluidic shear stress to promote the maturation of kidney organoids. The matured kidney organoids then could replace dialysis membrane to serve as kidney replacement therapy.
Based on the similarity to kidney structure and function, kidney organoids have been used to screen drugs. Taking cisplatin kidney injury model as an example, after introduction of 5 µM cisplatin into the culture medium with kidney organoids, acute apoptosis of mature proximal tubular cells emerged at only one day. 91 In addition, kidney organoids can also be used as kidney disease models. For example, polycystic kidney disease is an autosomal dominant hereditary kidney disease that lacks an ideal disease model. Cruz et al. 92 have used kidney organoids to generate a polycystic kidney disease model with decreased cystogenes of polycystin-1 after removing stoma, while increased cysts when adding cyclic adenosine monophosphate. Furthermore, kidney organoids from iPSCs of a congenital nephrotic patient with a missense mutation of Nephrin revealed impaired silt diaphragm which could be restored by genetic correction of mutation in the single amino acid, resulting in normal silt diaphragm formation.93,94 Apart from establishing disease models, exploring gene expression in kidney intrinsic cell lines is available. Single cell transcriptional signature analysis of podocytes disease in kidney organoids revealed LYPD1, PRSS23, and CDH6 correlated with human glomerular disease, which had not been reported before as it is difficult to study the gene expression signatures of podocytes in vivo. 95
When it comes to transplantation application of kidney organoid, many questions remain to be resolved. First, the source of PSCs needs to be secured. While ESCs are viewed with potential ethic concerns, iPSC banks with MHC matching are not. Recently, urine-derived renal progenitor cells has been published that showed renal stem cell markers -SIX2, CITED1, WT1, CD24, and CD106 and pluripotency-associated proteins- TRA-1–60, TRA-1–81, SSEA4, C-KIT, and CD133 indicating a potential source for kidney organoids. 96 Second, current kidney organoids are too small to provide sufficient filtration ability. Therefore, kidney organoids shall contain a sufficient amount of nephrons for future transplantation; Third, as blood perfusion is essential to promote the maturation of kidney organoids,88–90 the transplantation site should be adjacent to great vessels and ureter. It has been reported using omentum 97 and retroperitoneal fat tissue 98 as the transplantation site. Retroperitoneal fat tissue resulted in better glomerular filtration rate. However, both studies transplanted metanephroi not kidney organoids into rats. A recent study 99 showed transplanted kidney organoids beneath kidney capsule could survive for several months and also be vascularized from endothelial cells of host mice. Finally, although kidney organoids exhibit better nephron-like structures after transplantation, structure of tubules brush border, filtration barrier, and capillary lumens were not as organized as in the mature kidney tissues, requiring further improvement of kidney organoids differentiation.
Conclusions
In our review, we focused on the mechanisms by which stem cells could help treat kidney diseases, as well as the use of kidney organoids as an emerging treatment approach and research resource. Although MSCs are versatile with many mechanisms during the treatment, exosomes may be an alternative to fulfill the therapeutic effects of MSCs. As transdifferentiation of MSCs into tubule cells is not successful in vivo, 10 MSCs may restore kidney injury via paracrine pathways including secreting exosomes. In fact, exosomes are being used in a clinical trial (NCT04173650).
Clinically, stem cells have been used to treat lupus nephritis patients, with good complete recovery rates. However, clinical trials using MSCs in diabetic nephropathy did not show sufficient effect and the treatment effects of stem cells on AKI are still controversial. As lupus nephritis or primary amyloidosis are systemic diseases, MSCs exhibit effective treatment results clinically. Based on current clinical methods, delivering exosomes into injured kidney through renal artery could be a more feasible and useful procedure. In addition, injection of exosomes subrenal capsule may also be a clinically relevant delivery method.
Although many studies have shown the safety of MSCs treatment, the side effects of stem cell treatment of kidney diseases remain to be investigated. Nevertheless, we foresee a great future for stem cell therapy of nephropathy. Through better understanding the pathophysiology of CKD and AKI, we will further clarify the protective mechanisms of stem cells in the treatment of kidney diseases.
Supplemental Material
EBM915901 Supplemental Material - Supplemental material for Stem cell-based treatment of kidney diseases
Supplemental material, EBM915901 Supplemental Material for Stem cell-based treatment of kidney diseases by Binbin Pan and Guoping Fan in Experimental Biology and Medicine
Footnotes
Authors’ contributions
All authors contributed to writing the article. All authors reviewed and approved the final submission.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors acknowlege the financial support from NIH RO1 DE 025474 for the research, authorship, and publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.