
Abstract
Prenatal dexamethasone has been shown to increase blood pressure in male offspring but the mechanism for the increase in blood pressure is unclear. The present study examined if prenatal programming by maternal injection of dexamethasone on days 15 and 16 of gestation affected the blood pressure comparably in female and male offspring. Our hypothesis was that males would be affected by prenatal dexamethasone to a greater extent than females and that either an increase in renal tubular transporter abundance or an increase in renin or aldosterone system would be associated with hypertension with prenatal programming. Prenatal dexamethasone increased blood pressure at two months and six months of age and resulted in proteinuria and albuminuria at six months in male but not female rat offspring. There was no effect of prenatal dexamethasone on blood pressure and proteinuria at one month in male and in female offspring. While prenatal dexamethasone increased male renal thick ascending limb sodium potassium two chloride cotransporter protein abundance at two months, prenatal dexamethasone on days 15 and 16 of gestation did not affect transporter abundance in males at other ages, nor did it affect proximal tubule sodium/hydrogen exchanger or distal convoluted tubule sodium chloride cotransporter protein abundance at any age. There was no difference in systemic renin or aldosterone in the prenatal dexamethasone group compared to same sex controls. In conclusion, male but not female offspring have an increase in blood pressure and urinary protein excretion with prenatal dexamethasone. The increase in blood pressure with prenatal programming was not associated with a consistent increase in renal tubular transporter protein abundance, nor plasma renin activity and serum aldosterone.
Keywords
Impact statement
Prenatal dexamethasone is often administered to accelerate pulmonary maturation but the effect on other organs is unclear. We administer the same dose of dexamethasone per kg body weight for two days in rats and study the offspring. While males have been studied with more severe prenatal insults such as multiple and high-dose dexamethasone and low maternal protein diets, the effect of a dose of dexamethasone/body weight comparable to humans and the effect on females is unclear. Furthermore, the pathogenesis for the hypertension and the effect of prenatal dexamethasone on the female kidney have not been examined.
Introduction
We have previously shown that prenatal dexamethasone and a maternal low protein diet increased blood pressure in adult male rats. In prior studies, we administered 4 daily doses of dexamethasone on days 15–18 or fed pregnant rats a 6% protein diet compared to 20% in controls from day 12 gestation until birth. We found a parallel increase in blood pressure in male offspring and a significantly greater proximal tubule sodium/hydrogen exchanger (NHE3), thick ascending limb sodium potassium 2 chloride cotransporter (NKCC2), and distal convoluted tubule sodium chloride cotransporter (NCC) but not the epithelial sodium channel (ENaC) protein abundance in programmed male rats compared to controls.1–3 These results are consistent with an increase in renal sodium transporter abundance and thus sodium transport mediating the hypertension seen in prenatal programmed rats. However, while both observations may be true, it was not certain if the increase in transporter abundance was the sole cause for the increase in blood pressure in programmed rats. It is also unclear if the increase in blood pressure and factors such as the renin angiotensin system were comparably affected in male and female offspring.
Previous studies have examined if prenatal insults affected blood pressure in both male and female offspring in rats. A moderate reduction in maternal protein intake in pregnant rats from 19% to 8.5% resulted in male offspring that had hypertension as adults, 4 while female programmed adult offspring had comparable blood pressures as controls. 5 However, more severe maternal protein restriction to 5% or 6% resulted in hypertension in both male and female rat offspring.6,7 Male offspring born to mothers with surgically induced uteroplacental insufficiency had hypertension, whereas female offspring had comparable blood pressure to controls..8,9 Others have found that the uteroplacental insufficiency model of prenatal programming causes hypertension in both males and female offspring in 1 and 2 month old as well 12-month-old rats.10,11 However, at three and six months of age only programmed males, but not the females were hypertensive suggesting that there is an age dependence to the effect of programming on females with the uteroplacental insufficiency model.10,11
Studies have also been performed on offspring of sheep whose mothers were administered glucocorticoids during gestation to determine if prenatal programming affected males and females differently. Administration of prenatal glucocorticoids to sheep either on days 26–28 gestation 8,12 or on days 80 and 81 gestation 13 (term 145–150) increased in blood pressure in both male and female offspring. Adult male sheep that were the offspring of mothers that were administered glucocorticoids on days 80 and 81 of gestation had a reduction in glomerular filtration rate and an attenuated natriuresis in response to a sodium load compared to control males. However, there was no difference between female control and prenatal glucocorticoid-treated female sheep. 14 In addition, prenatal glucocorticoid-treated males had a natriuresis upon sodium loading with angiotensin-(1–7) infusion compared to control males, but females had no significant response to angiotensin-1–7. 14 Proximal tubule cells from betamethasone-treated male offspring, but not female, had an increase in sodium uptake in response to angiotensin II. 15 Thus, while both male and female sheep have an increase in blood pressure in response to prenatal glucocorticoids, there is a sexual disperity in the renal response.
In the present study, we examined if female offspring were comparably affected by maternal dexamethasone. We administered a less severe maternal insult (two daily doses vs. four doses previously) to determine if the increase in transporter abundance and/or increase in renin angiotensin system could be dissociated from the increase in blood pressure. Our hypothesis was that males would be affected by prenatal dexamethasone to a greater extent than females and that either an increase in renal tubular transporter abundance or an increase in renin or aldosterone would be responsible for the hypertension with prenatal programming.
Materials and methods
Animals
Female Sprague Dawley rats were administered one dose of dexamethasone (0.2 mg/kg body weight) or saline (vehicle) on days 15 and 16 of pregnancy as we have performed previously.16,17 The pregnant rats and their offspring had free access to water and Teklad global 18% protein rodent diet #2018–0.2% sodium (Madison, WI). No more than three male and three female rats per litter were used in these studies. Litters greater than 10 pups were reduced to 10 pups. We studied male and female offspring of vehicle and dexamethasone-treated rats at one, two, and six months of age. These studies were approved by the IACUC of the University of Texas Southwestern Medical Center (Protocol 2017–101904).
Blood collection and kidney harvest
Rats were anesthetized with an intraperitoneal injection of Inactin (10 mg/100 g body weight). Blood was collected from the left ventricle of the heart using a winged infusion set coated with EDTA. Plasma was placed in a chilled tube containing 20 µmol/L enalaprilat, 5 mmol/L EDTA, 10 µmol/L pepstatin A, and 1 mmol/L 1,10-phenanthrolene (all chemicals were from Sigma Chemical Co., St. Louis, MO). Serum samples were collected in separate tubes. The blood samples were centrifuged at 1200g for 10 min at 4°C and the supernatants were stored at −80°C. The kidneys were immediately clamp frozen in liquid nitrogen and stored at −80°C.
Blood pressure
Blood pressure was measured in male and female rats at one, two, and six months of age. The investigator that measured the blood pressure was blinded as to whether the rats were from the group whose mother received vehicle or dexamethasone. The rats were first placed in Lucite tubes and a blood pressure cuff was inflated several times for four consecutive days to acclimate the rat to the procedure as we have described previously.18–20 On the fifth day, blood pressures were measured using the CODA Blood Pressure Non-Invasive Pressure Analyzer (Kent Scientific Corporation, Torrington, CT). This instrument uses volume pressure recording that has been shown to correlate with measurements made using telemetry. 21 The average of at least five readings was used as the blood pressure of the rat.
Urine collection
At one, two, or six months of age, the rats were placed in metabolic cages and acclimated for 24 h. The rats had free access to food and water. Urine was then collected in a container that had 50 µmol pepstatin, 10 mg sodium azide, 125 μmol EDTA to prevent protein degradation.18,22–24 Urine protein was measured using the Bio-Rad protein colorimetric assay based on the Bradford assay (Bio-Rad Laboratories; Hercules, California), and urine albumin using a rat urinary albumin enzyme immunoassay kit (Nephrat; Exocell; Philadelphia, Pennsylvania). Urine and serum creatinine were assayed using capillary electrophoresis. Urine sodium was measured by flame photometry.
Plasma renin activity and serum aldosterone
Plasma renin activity was measured using a Plasma Renin Activity ELISA kit (ALPCO, Salem, NH). Angiotensin-1 was generated by incubating plasma samples for 90 min at 37°C with the buffer and protease inhibitors supplied in the kit. A duplicate reaction was incubated on ice. The samples and calibration solutions were applied to the rabbit Anti-Ang-I-coated 96-well plate and incubated on a shaker for 60 min with the angiotensin-I-biotin conjugate. After five microplate well washes, the bound Ang-I-Biotin samples were incubated with a streptavidin-horseradish peroxidase conjugate for 30 min. The wells were washed again before incubating with the TMB (3,3',5,5'-tetramethylbenzidine) substrate for 15 min. After stopping the reaction, the absorbance was read at 450 nm.
Serum aldosterone was measured using the manufacturer’s instructions using the Aldosterone EIA Kit (Enzo, Plymouth Meeting, PA). Briefly, the samples were incubated with an aldosterone antibody and conjugate on a donkey anti-sheep IgG microtiter plate overnight at 4°C. The wells were then washed and subsequently incubated for 1 h at room temperature with a p-Nitrophenyl Phosphate, Disodium Salt (pNpp) substrate. After stopping the reaction, absorbance was read at 405 nm.
SDS-PAGE and immunoblotting
At the time of study, renal samples were sliced from the frozen kidney and homogenized in a buffer containing 250 mmol/L sucrose, 10 mmol/L triethanolamine, 100 μg/mL phenylmethylsulfonyl fluoride (Research Products International, Mount Prospect, IL), and 10 µL/mL protease inhibitor cocktail (Sigma-Aldrich Chemical Company, St. Louis, MO). The samples were subsequently centrifuged for 15 min at 1000g. Brush-border membrane vesicles were isolated from the kidney homogenate by magnesium precipitation and differential centrifugation as previously described. 1 The protein content was assessed by Bradford assay (BioRad, Hercules, CA). Protein samples were denatured for 10 min at 37°C; 20 ug of protein was loaded and then separated on a 7% polyacrylamide gel using SDS-PAGE.25–27 Proteins were transferred to polyvinylidene difluoride membranes at 100 MA overnight at 4°C. The blots were blocked with Blotto (1% nonfat milk and 0.1% Tween-20 in PBS, pH 7.4) before incubation overnight at 4°C with primary antibody. Rabbit antibodies to NKCC2 and NCC both at a 1:1000 dilution (Stressmarq Biosciences, Victoria, Canada) were added to the Blotto. A mouse antibody to NHE3 #1568 (a gift from Dr. Orson Moe) was added at a 1:10 dilution. 28 The blots were extensively washed in blotto before the addition of horseradish peroxidase-conjugated anti-mouse IgG secondary antibody at a 1:10,000 dilution (Cell Signaling Technology, Danvers, MA). Bound antibody was detected using enhanced chemiluminescence (Perkin-Elmer, Boston, MA). An antibody to β-actin was added at 1:10,000 dilution to verify equal loading (Sigma-Aldrich, St. Louis, MO). Densitometry was used to assess the relative abundance of NHE3, NKCC2, NCC, and β-actin. While we had previously examined the effect of prenatal dexamethasone on blood pressure, protein, and albumin excretion in male and female rats at 14 months of age, we did not examine transporter abundance which we examined in this study. 29
Chemicals
All chemicals were obtained from Sigma Chemical Company unless otherwise specified (St. Louis, MO).
Statistics
All data are expressed as the mean ± the standard error of the mean. Statistical analysis was assessed using Sigma Plot 13.0. The four groups were compared using two-way analysis of variance with a Student–Newman–Keuls post hoc analysis.
Results
The present study examined whether there was a difference in blood pressure in male and female rats in response to prenatal programming. Blood pressure of pregnant rats treated with dexamethasone administered on days 15 and 16 of gestation was compared to the vehicle-treated groups at one, two, and six months of age. As shown in Table 1, there was no difference in blood pressure in males and females at one month of age and prenatal dexamethasone did not affect blood pressure in either sex. Prenatal dexamethasone resulted in significant increase in blood pressure in male but not female rats at two and six months of age (P < 0.001). At six months of age, vehicle and dexamethasone-treated male rats had a higher blood pressure than vehicle and dexamethasone female rats.
Effect of dexamethasone on systolic blood pressure in male and female rats at various ages (blood pressure in mm Hg).

^P< 0.001 male vs. female.
*P < 0.001 vs. all other groups.
Table 2 shows the effect of prenatal dexamethasone on the weights of rats at various ages. Male rats weighed more than female rats at all ages. Prenatal dexamethasone had no effect on the weight of rats at one and two months of age. Vehicle control male rats weighed more than the prenatal dexamethasone male rats at six months of age.
Effect of prenatal dexamethasone body weight in male and female rats.
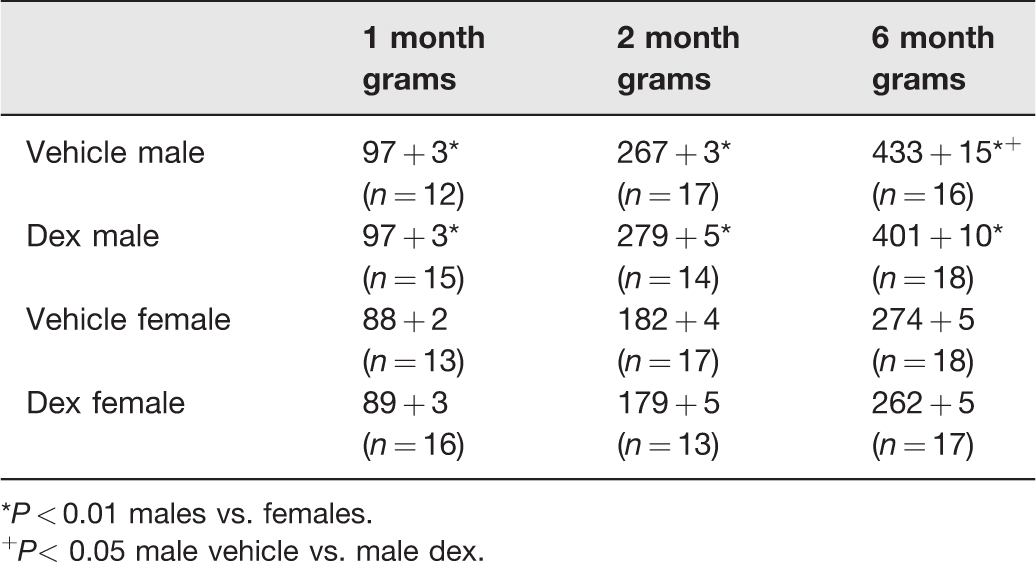
*P < 0.01 males vs. females.
+P< 0.05 male vehicle vs. male dex.
The effect of sex and prenatal dexamethasone on 24-h urinary protein and albumin excretion is shown in Tables 3 and 4, respectively. At one month of age, there was no difference in protein or albumin excretion between males and females and prenatal vehicle-treated compared to prenatal dexamethasone-treated rats. In the two-month-old and six-month-old rats, males had a greater protein and albumin excretion than females. Prenatal dexamethasone did not affect protein or albumin excretion at two months of age in either sex. However, at six months of age, male prenatal dexamethasone rats had a greater urine 24-h protein and albumin excretion than vehicle male rats (P < 0.05). There was no effect of prenatal dexamethasone in six-month-old female rats.
Effect of prenatal dexamethasone on 24-h urinary protein (mg) in male and female rats.

+P < 0.05 male vehicle > female vehicle.
*P< 0.05 Dex male > other groups.
Effect of prenatal dexamethasone on 24-h urinary albumin excretion (mg) in male and female rats.
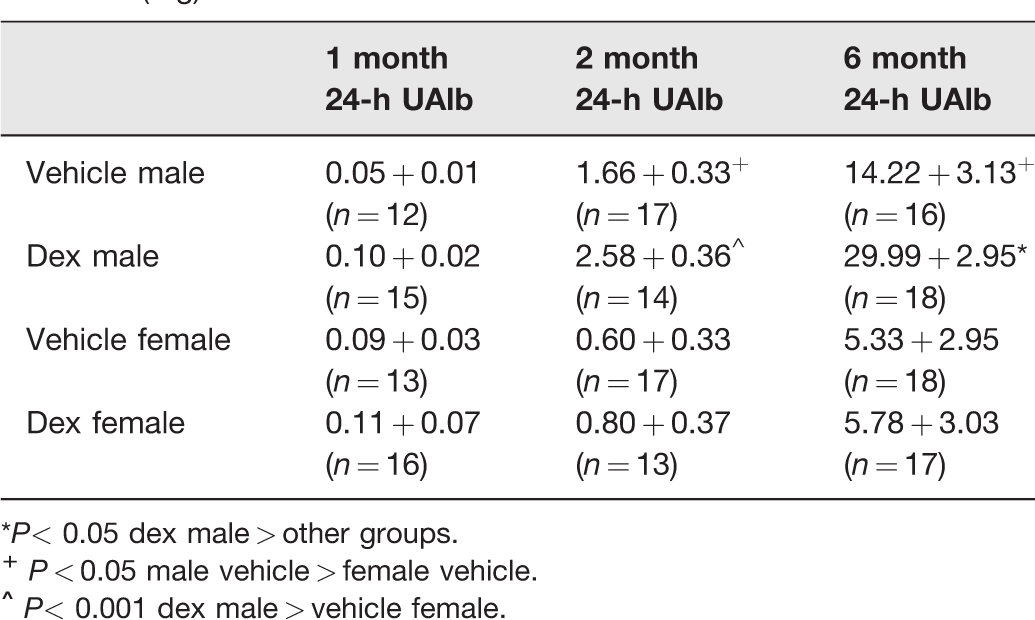
*P< 0.05 dex male > other groups.
Table 5 shows the 24-h urine sodium excretion and sodium excretion normalized for urine creatinine in the four groups at one, two, and six months of age. There was no difference between the dexamethasone and the control group at any age either in males or females. The dexamethasone and vehicle males had a greater sodium excretion at two months of age compared to the females, but this did not persist at six months of age. The female vehicle and dexamethasone groups had a greater urine sodium/creatinine than the males.
Effect of prenatal dexamethasone 24-h urinary sodium excretion (UNa, mmole) and Na/Cr ratio (mmol/mg) in male and female rats.
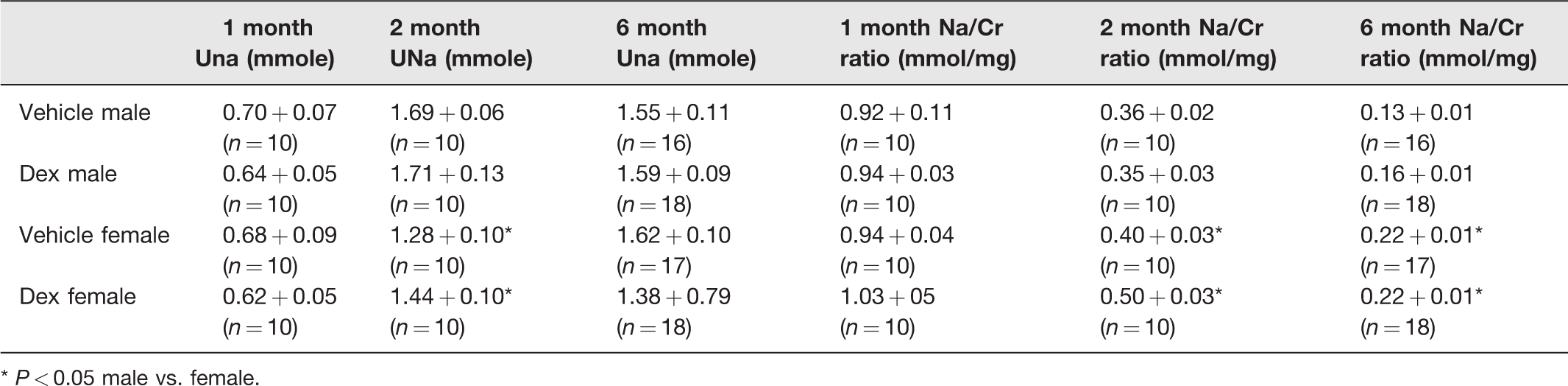
* P < 0.05 male vs. female.
The effect of prenatal dexamethasone on creatinine clearance is shown in Table 6. At one month of age, the vehicle group had a greater uncorrected creatinine clearance than the dexamethasone group, but the creatinine clearance was no different when corrected for body weight. Male vehicle and dexamethasone male rats had a higher creatinine clearance than the respective female vehicle and dexamethasone rats at six months of age. However, when corrected for body weight, there was no effect of sex or prenatal dexamethasone at any age.
Effect of prenatal dexamethasone on creatinine clearance (CrCl mL/min) in male and female rats.
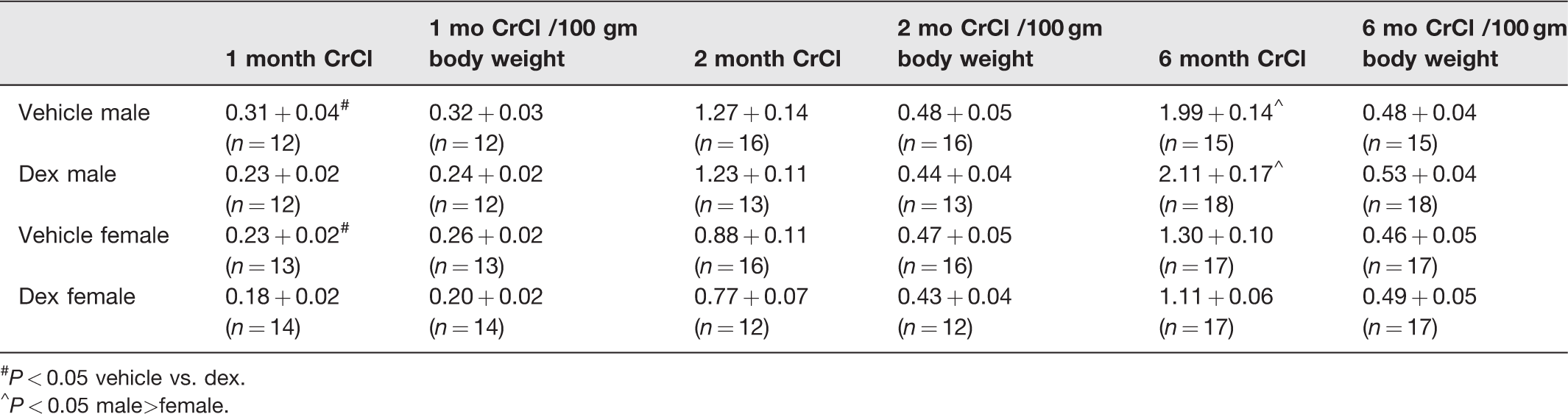
#P < 0.05 vehicle vs. dex.
^P < 0.05 male>female.
We next examined if prenatal programming by maternal administration of dexamethasone affected plasma renin activity and serum aldosterone as shown in Table 7. There was no significant difference in plasma renin activity at one and two months of age in the four groups. Surprisingly, the plasma renin activity was higher in the vehicle male and females compared to the dexamethasone-treated rats at six months of age. Serum aldosterone levels were elevated in female dexamethasone group at two months compared to control but were not different in the male dexamethasone group at this age or in the one and six-month-old rats. Thus, while there are some differences in renin and aldosterone at some ages, none are consistent with renin or aldosterone mediating the prenatal programming of hypertension seen in male rats at two and six months.
Effect sex on plasma renin activity and aldosterone.

*P < 0.05 vehicle vs. dex.
+ P < 0.05 female dex vs. male and female vehicle.
Finally, we examined if the selective increase in blood pressure in males by prenatal dexamethasone was due to an increase in renal transporter abundance. We previously demonstrated that a total of four daily doses of 0.2 mg/kg body weight on days 15–18 of gestation increased renal NHE3, NCC, and NKCC2 protein abundance in male offspring at two months of age. 3 The results of the present study examining the effect of dexamethasone on days 15 and 16 of gestation are shown in Tables 8 (NHE3), 9 (NKCC2), 10 (NCC) and a representative blot is shown in Figure 1. We include studies of male and female vehicle- and dexamethasone-treated rats at 14 months of age to examine aged rats. We found that two daily prenatal doses of dexamethasone in the current study increased NKCC2 protein abundance in prenatal dexamethasone-treated male rats compared to male vehicle controls at two months of age as we have found previously with four doses, but not at 1, 6, and 14 months of age. NHE3 (neither whole kidney nor brush border membrane) and NCC protein abundance was not increased in prenatal dexamethasone-treated male rats at two months in this study using two daily doses of dexamethasone compared to vehicle controls. We find that there were differences between male and female rats transporter protein abundance in some age groups, but despite the greater NKCC2 and NCC protein abundance in female rats there was no effect of dexamethasone as females had a lower blood pressure than males. Importantly, the increase in blood pressure in male rats could not be consistently accounted for by an increase in transporter abundance. Thus, the increase in blood pressure in programmed male rats was unlikely due to an increase in transporter abundance.
Effect of sex and age on NHE3 protein abundance.

aBrush border membrane vesicles.
*P < 0.05 dex male vs. dex female.
†P < 0.05 males vs. females.

Representative immunoblots for NHE3, NKCC2, and NCC. Immunoblots assaying NHE3, NKCC2, and NCC protein abundance at six months of age for in vehicle- and prenatal dexamethasone-treated male and female rats. MV: male vehicle; MDX: male dexamethasone; FV-female vehicle; FDX: female dexamethasone.
Effect of sex and age on NKCC2 protein abundance.

$Male dex vs. male vehicle P < 0.05.
*Dex male vs. dex female P < 0.001.
^Females vs. males P < 0.05.
#Vehicle female vs. vehicle male P < 0.05.
Effect of sex and age on NCC protein abundance.

+ P < 0.05 dex male vs. dex female.
^P < 0.05 female vs. male.
*P < 0.05 vehicle female vs. vehicle male.
Discussion
The present study examined whether there was a difference in blood pressure in male and female rats in response to prenatal programming with dexamethasone administered on days 15 and 16 of gestation compared to vehicle-treated offspring. In the present study, we found that only male rats of mothers treated with prenatal dexamethasone had hypertension at two and six months of age. While we had previously hypothesized that an increase in transporter abundance was responsible for hypertension with prenatal programming, we found that neither an increase in NHE3, NKCC2, nor NCC protein abundance were the primary factors leading to hypertension in programmed male offspring.
We have previously found that administration of dexamethasone on days 15 and 16 of life resulted in a comparable reduction in nephron number in male and female offspring.16,17,29 Here we showed that only males developed hypertension with two prenatal doses of dexamethasone. We and others have examined if prenatal insults affected blood pressure and glomerular abundance in male and female offspring. A moderate reduction in maternal protein intake in pregnant rats from 19% to 8.5% resulted in male offspring that had a reduction in nephron number and hypertension as adults, 4 while female adult offspring had comparable blood pressures and nephron endowment as vehicles. 5 However, more severe maternal protein restriction to 5% or 6% resulted in hypertension and a reduction in nephron number in both male and female rat offspring.6,7 Thus, the severity of the insult may determine if both a reduction in glomerular number and increased blood pressure may occur with prenatal insults. Pertinent to the purpose of this investigation, a reduction in glomerular number in female rats seen previously with two days of prenatal dexamethasone was associated with hypertension in both male and female offspring at 60–90 days of age but did not result in hypertension in females at 6 months of age nor at 14 months of age.17,29 Thus, a reduction in nephron number is unlikely to play a significant role to increase in blood pressure with prenatal programming. Furthermore, in almost all studies examining the effect of prenatal programming, the reduction in nephron number is usually between 20 and 30% of that of controls making a reduction in nephron number an unlikely major factor for hypertension in and of itself.
Measurement of blood pressure by telemetry in rats that were administered dexamethasone during the last week of pregnancy found that programmed rats had lower blood pressures at baseline than controls. 30 However, even minor disturbances in prenatal dexamethasone-treated rats resulted in blood pressures that were significantly higher than controls. 30 This is consistent with an increase in sympathetic nerve activity playing a role in mediating the hypertension in programmed rats. 19 The present study assessed blood pressure using tail cuff in rats that were trained for the procedure and appeared calm during the measurement of blood pressure. None-the-less tail cuff blood pressure even in trained rats is not entirely stress free.
We have previously found that six-month-old male offspring of rats whose mothers were fed a low protein diet (6%) during the last half of pregnancy had albuminuria but not total proteinuria in comparison rats whose mothers were fed a 20% control diet throughout pregnancy. 18 In the present study, we found that there was no significant difference in protein and albumin excretion in male and female offspring in controls vs. the prenatal dexamethasone groups at one and two months of age. The male dexamethasone group, but not the female group, had a greater 24-h protein and albumin excretion than vehicle controls at six months of age. The proteinuria and albuminuria are likely a harbinger of glomerular injury. Indeed, we previously found that prenatal dexamethasone male rats had increased proteinuria and albuminuria and a reduction in glomerular filtration rate at 14 months of age. 29 We also measured 24-h urine sodium excretion in one, two, and six-month-old rats. Only the two-month-old dexamethasone and vehicle males had a greater sodium excretion than the females but there was no difference between the dexamethasone and vehicle groups at any age. Corrected for urine creatinine, urine sodium was higher in females than males at two and six months of age, but also not different between controls and dexamethasone groups. Extracellular volume expansion due to sodium retention and the lack of a pressure natriuresis by the male dexamethasone group may be a factor to explain the hypertension in these rats. Unfortunately, diuretics were not administered to assess transporter activity.
We have shown that prenatal administration of dexamethasone on days 15 and 16 days of age resulted in ∼30% reduction in glomerular number compared to vehicle-treated controls. 16 Despite the reduction in nephron number, we showed that glomerular filtration rate measured using inulin clearance in male rats at 60–90 days of age and 6 to 9 months of age was comparable to control rats at both ages.16,17 In the present study, we did not find a decrease in creatinine clearance in one-, two-, and six-month prenatal dexamethasone-treated rats compared to control rats which is consistent with our previous results. However, we have previously shown that there is a decrease in glomerular filtration rate using inulin clearance in male offspring of prenatal dexamethasone-treated rats at 14 months of age compared to prenatal vehicle-treated male offspring. 29 Female glomerular filtration rate was not affected by prenatal dexamethasone at 14 months of age. Thus, the effect of prenatal programming to decrease glomerular filtration rate was only found in aged males and may be the result of hypertension and the reduced nephron number. In humans, there is evidence that the decrease in renal function occurs early in life. Using the National Health and Nutrition Examination Survey (NHANES), Khalsa et al. showed a higher prevalence of reduced eGFR in 12–15-year-old children who were of low birthweight (<2500 g) compared to children of normal birthweight (2500–4000 g). The prevalence of a low estimated glomerular filtration rate was even higher in very low birthweight (<1500 g) children. 31
A previous study had examined the effect of prenatal programming on plasma renin and aldosterone levels at one and two months of age. 6 In this study, prenatal programming was due to maternal ingestion of a low-protein diet during the last half of pregnancy. This study found that both male and female programmed rats at four and eight weeks had a lower plasma renin activity than the controls whose mothers were fed a normal protein diet. While we found comparable values of plasma renin activity with prenatal dexamethasone at one and two months of age, prenatal dexamethasone resulted in male and female offspring with reduced plasma renin activity at six months. Prenatal programming mediated by a low-protein diet resulted in an increase in serum aldosterone in both males and females at four and eight weeks. We found an increase in aldosterone in only females at two months but not at other ages nor in males at any age. The difference in the study examining the effect of prenatal low protein diet and this study is likely due to the type and severity of the prenatal insult. None-the-less renin and aldosterone were not likely factors mediating the hypertension in prenatal programming seen here.
Previous studies have examined the effect of programming on transporter abundance in several nephron segments. Offspring of rats whose mothers were fed a 6% protein diet had an increase in NCC and NKCC2 protein abundance but no difference in NHE3 protein abundance nor increase in any of the ENaC subunits at four weeks of age compared to offspring whose mothers were fed a 20% protein diet. 32 There was a comparable increase in NCC and NKCC2 protein abundance in programmed males and females in rats programmed with a low-protein diet 32 . We have previously shown that in male offspring of mothers treated daily with prenatal dexamethasone on days 15–18 had an increase in NHE3, NCC, and NKCC2 protein abundance at two months of age.1–3 However, alpha, beta, or gamma ENaC protein abundance is not affected by prenatal programming by glucocorticoids. None-the-less prenatal programming by a low protein diet resulted in an increase in cortical collecting tubule sodium transport at least in part mediated by higher serum aldosterone levels. 3 Proteinuria seen in programmed animals may increase urinary proteases which has been shown to activate gamma ENaC by removal of a 43-mer domain which has not been examined.33–35
It should also be mentioned that a change in expression of claudins by a prenatal insult, which could affect paracellular passive transport, is possible but has never been examined.
The result of the present study demonstrates that the increase in transporter abundance cannot explain the higher blood pressure in male programmed rats. However, it is possible that prenatal dexamethasone could have affected the phosphorylation of NCC or NKCC2 activity without affecting transporter abundance which was not explored here. In the present study, we examined if a less intense prenatal insult would allow us to determine if an increase in transporter abundance was an important factor for the elevated blood pressure with programming. We find that there was a greater NCC protein abundance in female than male rat kidneys, and NKCC2 in female rat kidneys after six months of age, yet the female groups had lower blood pressure than the male dexamethasone group after two months of age. The only group where prenatal dexamethasone resulted in an increase in transporter protein abundance was for NKCC2 in males at two months, while the protein abundance was comparable in the male control and hypertensive dexamethasone groups at 6 and 14 months. In summary, our data are consistent with other factors besides renal transporter abundance, plasma renin activity, and aldosterone being the primary cause of the elevated blood pressure with prenatal programming in male rats. Female rats are protected from hypertension with prenatal progamming by dexamethasone.
Footnotes
AUTHORS’ CONTRIBUTIONS
All authors contributed to the design, interpretation and execution of the studies and analysis of the data and review of the manuscript. MB wrote the manuscript. All individuals who made contributions to this study are authors of the manuscript.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship and/or publication of the article.
FUNDING
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by National Institutes of Health Grants [DK078596] (MB) and [1P30DK079328-01] (Orson Moe, PI-MB Co PI Physiology Core) and a grant from Children’s Clinical Research Advisory Committee (CCRAC) from Children’s Health of Dallas.