
Abstract
This study aimed to investigate the association among genetic variants of the complement pathway CFB R32Q (rs641153), C3 R102G (rs2230199), and CFH (rs1410996) with age-related macular degeneration (AMD) in a sample of the Brazilian population. In a case-control study, 484 AMD patients were classified according to the clinical age-related maculopathy grading system (CARMS) and compared to 479 unrelated controls. The genetic variants rs1410996 of complement H (CFH), rs641153 of complement factor B (CFB), and rs2230199 of complement 3 (C3) were evaluated through polymerase chain reaction (PCR) and direct sequencing. The associations between single nucleotide polymorphisms (SNPs) and AMD, adjusted by age, were assessed by using logistic regression models. A statistically significant association was observed between AMD risk and rs2230199 variant with an OR of 2.01 (P = 0.0002) for CG individuals compared to CC individuals. Regarding the comparison of advanced AMD versus the control group, the OR was 2.12 (P = 0.0036) for GG versus AA genotypes for rs1410996 variant. Similarly, the OR for rs2230199 polymorphism was 2.3034 (P = 5.47e-05) when comparing CG individuals to CC carriers. In contrast, the rs641153 variant showed a significant protective effect against advanced AMD for GA versus GG genotype (OR = 0.4406; P = 0.0019). When comparing wet AMD versus controls, a significant association was detected for rs1410996 variant (OR = 2.16; P = 0.0039) comparing carriers of the homozygous GG versus AA genotype, as well as in the comparisons of GG (OR = 3.0713; P = 0.0046) and CG genotypes (OR = 2.2249; P = 0.0002) versus CC genotype for rs2230199 variant, respectively. The rs641153 variant granted a significant protective effect against wet AMD for GA versus GG genotypes (OR = 0.4601; P = 0.0044). Our study confirmed the risk association between rs2230199 and rs1410996 variants and AMD, and the protective role against AMD for rs641153 variant.
Impact statement
This is the first Brazilian study to show the association among genetic variants of the complement pathway CFB R32Q (rs641153), C3 R102G (rs2230199), and CFH (rs1410996) with age-related macular degeneration etiology and severity. The Brazilian population has unique characteristics, with ethnic peculiarities and miscegenation, therefore the evaluation of patients from other regions would strengthen the results. This study is also relevant since it analyzes not only the risk for general AMD, as most of published studies have done, but also the risk for different disease subphenotypes, allowing an association with AMD severity and possibly with disease progression.
Introduction
Age-related macular degeneration (AMD) is a degenerative and progressive retinal disorder and the leading cause of blindness in the elderly in Western World, 1 affecting approximately 8.7% of the blind population. 2 The number of affected individuals is expected to increase to 288 million by 2040. 3 Among the Brazilian elderly, it is estimated that approximately three million suffer from AMD, which is responsible for 4.9% of blindness. 4
Increasing prevalence of AMD as a result of increasing life expectancy highlights its deleterious effect on the quality of life in the elderly worldwide, 5 , 6 becoming a major public health problem. Visual impairment due to advanced AMD is significantly associated with low quality of life compared to patients with chronic disease 7 and also an increased incidence of falls 8 and cognitive impairment. 9
This complex disease is derived from inherited and environmental exposures. 10 The US twin study of AMD quantified the proportions of variance due to genetic and environmental influences as 46% to 71% and 19% to 37%, respectively. 11 Common risk variants near the complement genes CFH,12–14 CFB, 15 , 16 C2, 15 , 16 C3,17,18 and CFI19,20 account for nearly 57% of the contribution of known variants to disease risk, 21 emphasizing the critical role of the complement pathway in AMD pathogenesis. However, the associations between these genes and AMD vary due to genetic variation within a population in a country and across different populations (different ethnicities) worldwide. 22
The CFH gene, located on chromosome 1, region q32, was the first significant gene to be implicated in the pathogenesis of AMD. CFH encodes complement factor H (fH), a key regulatory glycoprotein that acts as a negative regulator of the complement system, inhibiting complement alternative pathway by promoting factor I (C3b inactivator) and preventing local formation of more C3b. 23 A common variant of CFH, in intron 14 (−543 G > A, rs1410996) has been reported to be associated with a higher risk of AMD.24–26 Similarly, the rs2230199 (R102G) in exon 3 of the C3 gene, a common missense variant that causes replacement of the arginine residue at codon 102 for a glycine (p.Arg102Gly), 27 has been strongly associated to the disease in studies on Caucasian populations. 17 , 28 However, this correlation has not been confirmed in Asian populations. 29
Nevertheless, there are some protective variants in complement genes. The CFB gene is located at chromosome 6p21, and its several transcript variants, such as rs641153, rs4151667, rs2072633, rs12614, rs1048709, have been related to the pathogenesis of AMD. 30 The CFB R32Q (rs641153) is a non-synonymous variant in exon 2 characterized by the exchange of the amino acid glutamine for arginine at codon 32. Former studies have shown its protective effect against AMD, 15 , 30 , 31 since it decreases the potential to form convertase and to amplify complement activation. 31
Although several studies from different populations have suggested the implication of these gene variants on the risk of developing AMD, there have been no genetic studies confirming these associations in the Brazilian population. Therefore, our study aimed to assess the association between genetic variants of the complement pathway CFB R32Q (rs641153), C3 R102G (rs2230199), and CFH (rs1410996) with AMD in a sample of the Brazilian population, from the Southeast region.
Materials and methods
A case–control study was conducted, encompassing 484 patients with AMD and 479 patients without AMD selected at the Clinical Hospital of the University of Campinas (UNICAMP), Campinas, São Paulo, located in the Southeast of Brazil. The study adhered to the tenets of the Declaration of Helsinki and was approved by the institution’s Ethics Committee. The written informed consent was obtained from all participants.
The study subjects aged older than 50 years were classified according to the Clinical Age-Related Maculopathy Grading System (CARMS). 32
Case group formation
Affected individuals were defined as those having AMD (CARMS stages 2–5) and were stratified in advanced AMD with either geographic atrophy or neovascular disease (CARMS stages 4 and 5) and wet/neovascular AMD (CARMS stage 5), considering the status of the worse eye.
Control group formation
The control group included individuals who did not show any evidence of AMD (CARMS 1). These individuals consisted of non-family companions of patients examined in the Ophthalmology Department or patients with initial or intermediate cataracts.
Clinical assessment of cases
All participants underwent a standard ophthalmic examination, including visual acuity, refraction, biomicroscopy, and fundoscopy through a dilated pupil performed by a retinal specialist. Subsequently, fluorescein angiography, indocyanine green angiography (ICG), and optical coherence tomography (OCT) were performed and the diagnosis of AMD was confirmed following CARMS criteria as well.
Exclusion criteria
The exclusion criteria used for both case and control groups were: presence of any opacity of media that would prevent good quality of fundus biomicroscopy and/or retinography; patients with other diseases that could be related to the development of macular neovascularization, such as: polypoid choroidal vasculopathy, angioid streaks, high myopia (spherical equivalent less than −6 diopters), infectious, inflammatory or hereditary chorioretinal disease or trauma; presence of macular hemorrhage from any other cause and consanguinity among subjects.
DNA extraction and genotyping
Genomic DNA was extracted from peripheral blood using QIAamp® DNA Blood Midi Kit (Qiagen, Wertheim, Germany), according to the manufacturer’s instructions. The quantification and purity of the DNA were measured using a Nanodrop 2000 spectrophotometer (260/280 absorbance) (Waltham, MA, USA). Genotyping was undertaken by PCR and direct sequencing using custom-designed primers presented in Table 1. Primers were selected based on the sequences of the specified genes available at https://www.ncbi.nlm.nih.gov/genbank/.
Primers used for PCR amplification and sequencing.

SNP: single nucleotide polymorphism.
Statistical analysis
Statistical analysis was performed using the R software (Foundation for Statistical Computing, Vienna, Austria). Mann–Whitney nonparametric test and Fisher’s exact test were used to compare age and gender distribution, respectively, between affected and not affected cohorts. Chi-square statistics were employed to test Hardy–Weinberg equilibrium (HWE) in the studied groups. Associations and odds ratios (OR) estimates were modeled through logistic regression models adjusted by age.
To evaluate the role of the genetic variants of the complement pathway in AMD susceptibility, a general comparison (cases vs. controls) and comparisons after disease stratification in advanced AMD (advanced AMD vs. controls) and wet form (wet AMD vs. controls) were performed considering all genotypes as independent co-variables in comparison to the reference genotype.
To reduce the possibility of type I error due to multiple testing, an adjusted P-value for Bonferroni correction was used in each comparison as a threshold to confirm statistical significance (P = 0.05/9 = 0.0055; since we tested three dependent variables for three genetic variants).
Results
This case–control association study comprised 484 AMD patients (54.96% females) including 281 wet AMD (58.06%) and 333 advanced AMD (68.80%), and 479 healthy individuals (53.65% females). No statistically significant difference was observed for gender between cases and controls (P = 0.6843). The mean ± standard deviation (SD) age was 73.05 ± 9.12 years for the AMD group and 66.07 ± 9.79 years in the control group (P < 0.0001). As the difference in age distribution between the AMD group and controls was statistically significant, age was adjusted as a common confounder in the analysis of the association between SNPs and AMD. Table 2 describes age and gender distributions among controls, cases, and subgroups of AMD.
Demographic characteristics of AMD patients and controls.

n: number of subjects.
*P < 0.0001 when cases were compared to controls (Mann–Whitney test).
+P = 0.6843 when gender distribution was compared between cases and controls (Fischer’s exact test).
The frequencies of genotype distributions of all studied samples were in Hardy–Weinberg equilibrium (P > 0.05), with the exception of the control group for the rs2230199 variant in the C3 gene (P = 0.0116).
The genotype and allele frequencies of the genetic variants of CFB R32Q (rs641153), C3 R102G (rs2230199), and CFH (rs1410996) are depicted in Table 3.
Genotype and allele distributions of CFB R32Q (rs641153), C3 R102G (rs2230199), and CFH (rs1410996) polymorphisms among the control group, AMD cases, and after disease stratification in advanced and wet AMD subgroups.
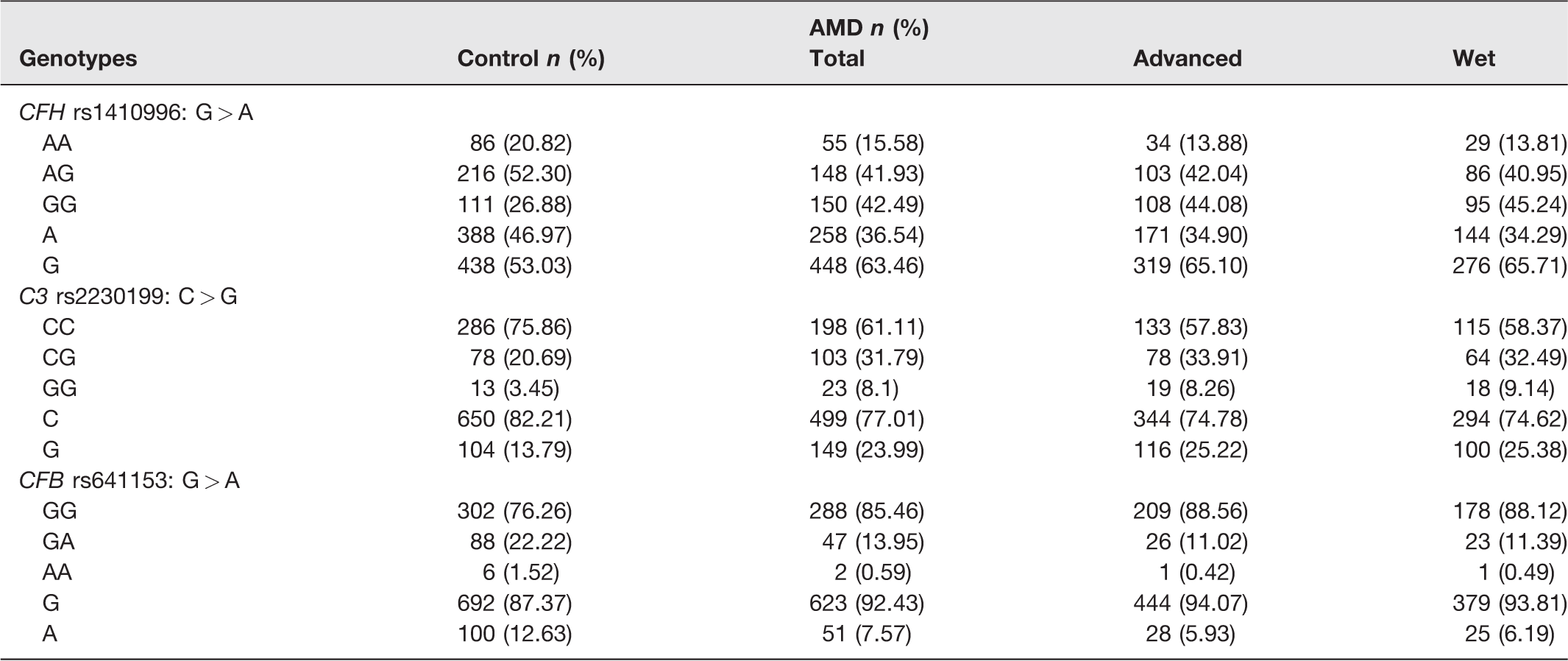
When evaluating cases versus controls, a significant association with AMD was detected for SNP rs2230199 in the C3 gene for the comparison between CG versus CC genotypes (P = 0.0002). However, our data did not show significant association with the risk of AMD for the variants rs1410996 in the CFH gene and rs641153 in the CFB gene, after Bonferroni correction (Table 4).
Comparisons of CFB R32Q (rs641153), C3 R102G (rs2230199), and CFH (rs1410996) polymorphisms between AMD cases and controls through logistic regression analysis adjusted by age.
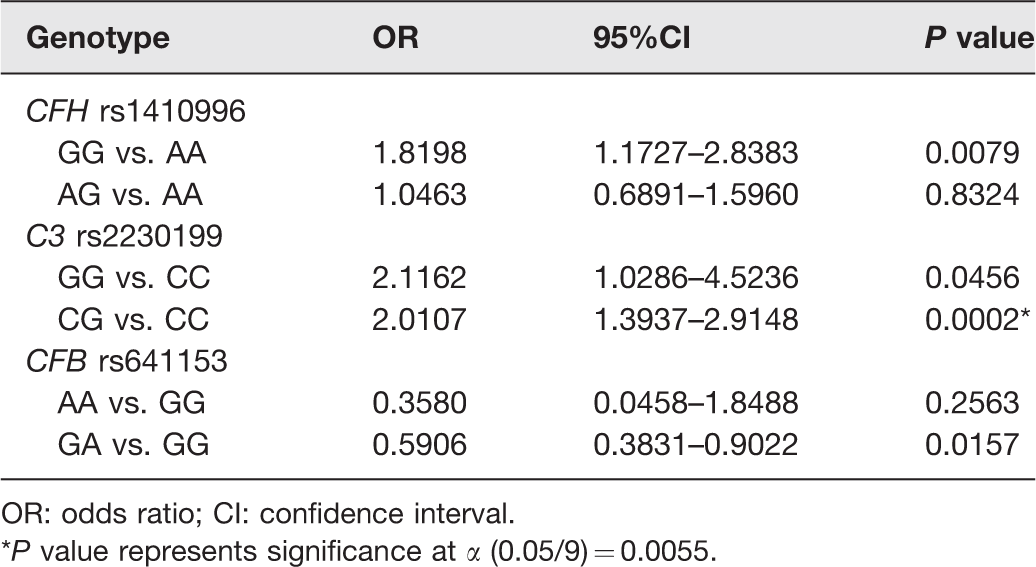
OR: odds ratio; CI: confidence interval.
*P value represents significance at α (0.05/9) = 0.0055.
Regarding the comparison of advanced AMD versus controls, the analysis of rs1410996 variant showed that carriers of the homozygous GG genotype had more than two-fold increased risk of developing advanced AMD (P = 0.0036) compared to those carrying the AA genotype. Similarly, the OR for the rs2230199 polymorphism for advanced AMD was 2.3034 (P = 5.47e-05) for heterozygous carriers (CG) compared to the CC genotype. In contrast, the rs641153 variant showed a significant protective effect against advanced AMD for the comparison between GA versus GG genotypes (P = 0.0019) (Table 5).
Comparisons of CFB R32Q (rs641153), C3 R102G (rs2230199), and CFH (rs1410996) polymorphisms between Advanced AMD cases and controls through logistic regression analysis adjusted by age.
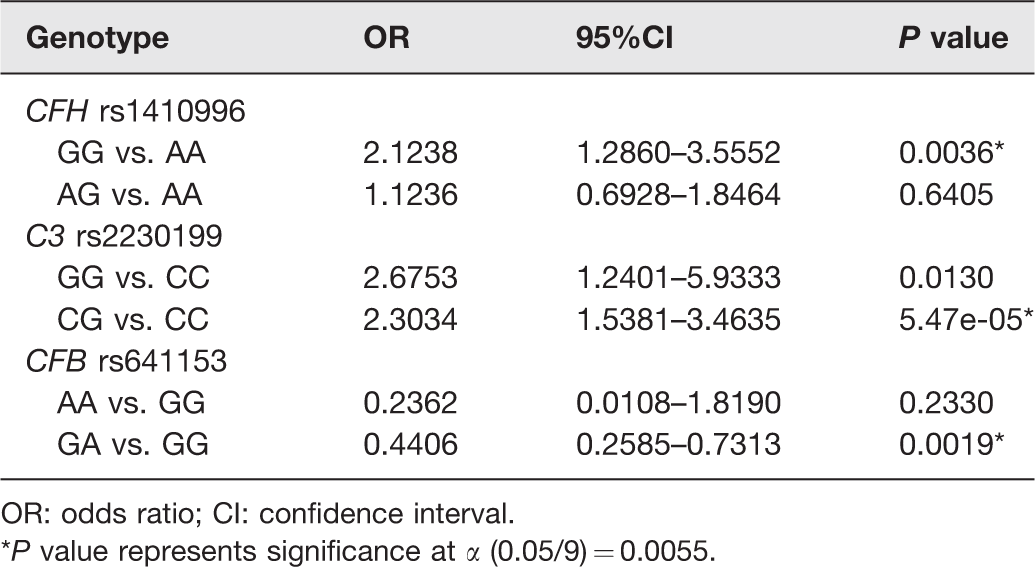
OR: odds ratio; CI: confidence interval.
*P value represents significance at α (0.05/9) = 0.0055.
The comparison of wet AMD and controls was statistically significant for the SNP rs1410996, showing that individuals carrying the GG genotype have more than two-fold increased risk of developing wet AMD (P = 0.0039) compared to AA individuals. The same pattern is observed for the comparisons of genotypes GG (OR = 3.0713; P = 0.0046) and CG (OR = 2.2249; P = 0.0002) versus CC for the rs2230199 variant, conferring a higher risk of developing this disease form. On the other hand, the rs641153 variant granted a significant protective effect against wet AMD for carriers of the GA genotype compared to the GG genotype (P = 0.0044) (Table 6).
Comparisons of CFB R32Q (rs641153), C3 R102G (rs2230199), and CFH (rs1410996) polymorphisms between wet AMD cases and controls through logistic regression analysis adjusted by age.

OR: odds ratio; CI: confidence interval.
*P value represents significance at α (0.05/9) = 0.0055.
Discussion
AMD is a multifactorial disease, in which complement system plays a key role. Previous studies in different ethnicities have shown that the genetic variants studied in this pathway (rs1410996; rs2230199; rs641153) are associated with AMD. However, these associations are not consistent and vary according to the geographic region and environmental factors to which these populations are subjected. Considering the high miscegenation content of the Brazilian population, this study evaluates the genetic profile of a sample of AMD patients from the Southeast regarding three variants in genes from the complement system.
Our data showed significant association of the risk of advanced and wet AMD with the noncoding variant rs1410996 in the CFH gene. This study in a sample of the Brazilian population demonstrated a higher risk of AMD with the G allele of rs1410996 in accordance with findings in Caucasian, 25 Japanese, 33 and Chinese 34 populations. The comparison of AMD cases and controls was not statistically significant, since we used an adjusted P-value for Bonferroni correction, in order to reduce the possibility of type I error due to multiple testing (P = 0.05/9).
When analyzing patients with wet AMD, the frequencies of the risk G allele were 65.71% in cases and 53.03% in the controls, similar to those reported in Japanese (66.8% in cases and 50.4% in controls), 33 Chinese (72% in cases and 55.6% in controls), 34 and Caucasian (80.8% in cases and 57.1% in controls) 25 populations. Compared to individuals with the AA genotype of rs1410996, individuals with the homozygous GG genotype had a 2.16-fold increased risk of developing wet AMD, while in an Asian meta-analysis the OR was 2.49. The same was observed for the advanced form of the disease, in which GG individuals had a 2.12-fold increased risk compared to AA individuals.
CFH inhibits the alternative complement cascade by regulating the spontaneous activation and amplification of C3 cleavage and assisting in the degradation of C3 cleavage products. 23 The mechanism of the relationship between AMD and the rs1410996 is unclear. It is suggested that the variant modulates the risk of AMD by regulating the expression of CFH, rather than disrupting the CFH protein function. 24
Despite the fact that the control group of the rs2230199 variant in the C3 gene was not in Hardy–Weinberg equilibrium (HWE) (P = 0.0116), our results were consistent with previously published findings. Although statistical tests play a key role in identifying the departures, when sample sizes are relatively large, significant departures from HWE can occur having little biological importance. Therefore, departures can be ignored since the downstream results are robust to the nature and magnitude of the deviations. 35 , 36
The prevalence and phenotypic spectrum of AMD vary among different ethnic groups as well as allele and genotype distributions. Frequencies of the risk G allele of rs2230199 were 23.99% in AMD cases and 13.79% in controls in this Brazilian sample, while the frequencies of the G allele were 25% to 31% in AMD patients and 19% to 21% in controls in Caucasians. 17 , 18 , 37 However, this allele was absent in Japanese and rare (< 1%) in Chinese populations. 38 Therefore, the rs2230199 variant is not associated with AMD due to its low frequency in Asians.
Our results showed significant association between the rs2230199 SNP and AMD susceptibility for the CG genotype compared to the CC genotype (OR = 2.01, P = 0.0002) consistent with the meta-analysis conducted by Zhang 29 in 2015, in overall populations (OR = 1.571, P = 0.000) and in the stratified analysis by ethnicity, as well as in Caucasians (OR = 1.576, P = 0.000). There was also a significant positive correlation between the rs2230199 variant and advanced AMD when comparing CG vs. CC genotypes (OR = 2.3034; P = 5.47e-05*), similar to another meta-analysis, also conducted by Zhang 39 in 2018, in overall (OR = 1.53; P < 0.001) and in the Caucasian population (OR = 1.55; P < 0.001). Significant associations with wet AMD were detected and were stronger than those observed for advanced AMD susceptibility for GG (OR = 3.0713, P = 0.0046) and GC individuals (OR = 2.2249, P = 0.0002).
C3 is the central component of the complement activation cascade and initiates the alternative complement pathway in order to form the membrane attack complex. Defective alternative pathway regulation and genetic alterations in the C3 gene is associated with AMD, increasing retinal cell damage and degeneration. 27 Variants in this gene, such as rs2230199, can affect the expression level and binding affinity of complement factors to the corresponding ligands. 40
Regarding the CFB polymorphism (rs641153), in the comparison of cases versus controls, although there was a suggestive protection for GA individuals, this result did not survive Bonferroni correction. We believe that this finding does not invalidate the protective role of this variant described in literature,41–43 since Bonferroni’s correction may increase the chance of type II error. Data from Caucasian, Asian, and Indian populations describe a significant protection for both GA and AA genotypes in general AMD risk.
In this study, a statistically significant protection GA versus GG genotype when evaluating advanced and wet AMD forms compared to controls was demonstrated, resulting in a risk reduction of 56% and 54%, respectively. These results are in accordance with what was described in literature, mainly in studies conducted in Caucasian populations. 43 , 44 Comparisons of AA versus GG genotype did not reach statistical significance in any of the evaluations. The low frequency of homozygous AA in the studied sample may have limited these associations, especially after disease stratification. Despite this, data in literature agree that AA genotype is also protective for advanced AMD, with an OR of 0.27. 43
The rs641153 variant of the CFB gene has a well-established protective role against AMD, and our results also confirm this finding in the Brazilian population. A functional study by Montes et al. 31 was performed to assess the biological role of the polymorphic variant. The authors demonstrated that CFB R32Q variant has lower hemolytic activity and about four times less binding affinity for C3, with reduced formation of convertase following plasma purification from AA homozygous individuals. Thus, the protective effect of CFB R32Q would be a consequence of a lower potential for activation of the alternative complement pathway, which is directly related to immune response in the pathophysiology of AMD.
Despite the association observed, the studied group comprises a sample of patients from the Southeast of Brazil. Due to the high ethnic heterogeneity observed in Brazil, the evaluation of patients from other regions would strengthen the results. Besides that, the effect of environmental factors, including smoking habits, was not assessed due to insufficient data.
In summary, our data suggest that the CFH noncoding variant rs1410996 and the C3 coding variant rs2230199 increase the risk of AMD in a sample of the Brazilian population. The rs1410996 variant showed a weaker risk association with AMD than that observed in Caucasians, but similar to the Asian population. In contrast, the rs2230199 variant was strongly associated with the disease, with more robust findings than those seen in Caucasians. The rs641153 variant of CFB showed a protective effect comparable to results in Caucasians. As a whole, our data support previous observations that genetic susceptibility to AMD varies among different ethnicities.
Since this is the first analysis of these polymorphisms in a Brazilian cohort, studies with a larger sample size are required to validate our findings. The identification of genetic risk factors in various ethnic groups will lead to a better understanding of the pathogenesis of AMD and subsequently will help to identify people at risk for the disease.
Footnotes
AUTHORS’ CONTRIBUTIONS
JMN, MGMV, JPCV, GA, and MBM participated in the design, interpretation of the studies and analysis of the data; JMN, MGMV, JPCV, FFC, and MBM reviewed the manuscript; JMN, MGMV, ABR, PHHR, and FMM performed ophthalmological examination in the patients included in the study; JMN, MGMV, ABR, GFSB, and RFO collected clinical data and blood samples; JMN, FFB, and SMSC conducted the experiments; JMN, JPCV, and MBM wrote the manuscript.
ACKNOWLEDGMENTS
The authors are grateful to Daniela Stancato, Francisco Carenzi, and Thiago Rodrigues for the help with sample storage and laboratory analysis.
DECLARATION OF CONFLICTING INTERESTS
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
ETHICAL APPROVAL
This research was approved by the Ethics Research Committee of the University of Campinas. Informed consent was obtained from all participants included. The procedures used were in accordance with the tenets of the Declaration of Helsinki for research involving human beings.
FUNDING
This study was supported by the Fund for Support to Teaching, Research and Outreach Activities (FAEPEX) grants 1525/15 and 251/18 and by São Paulo Research Foundation (FAPESP) grant 2010/18353–9.