
Abstract
Understanding the immune response to SARS-CoV-2 is important for development of effective diagnostics and vaccines. We report here a broad antibody response to SARS-CoV-2 spike protein receptor binding domain (RBD) in 100 convalescent patient plasma samples. Antibody isotypes IgA, IgM, and IgG exhibited significantly higher anti-RBD titers when compared to SARS-CoV-2 negative controls. IgG subtyping indicated IgG1 and IgG3 to be most abundant. Greater than 90 % of SARS-CoV-2 positive plasma samples tested exhibited significant neutralization capacity using a surrogate virus neutralization assay. Of the IgG subclasses, IgG1 and IgG3 exhibited the highest viral neutralization capacity; whereas, IgG2 and IgG4 viral neutralization was not observed. Comparison of SARS-CoV-2 elicited total IgG binding to emerging variant (alpha, beta, and delta) RBDs indicated decreased binding. Furthermore, neutralization by SARS-CoV-2 convalescent plasma of delta and omicron variant RBDs was significantly decreased suggesting that neutralizing antibodies in convalescent plasma are less effective in inhibiting variants currently in circulation.
Impact Statement
The COVID-19 pandemic has spread rapidly across all continents, and has dramatically impacted both society and the respective health systems due to its high rate of infectivity, morbidity and mortality. The focus of this study was to profile the quality (isotypes) and quantity (titers) of anti-receptor binding domain (RBD) antibodies generated in response to COVID-19 in order to gain insight into their respective potential viral neutralization activity. Results indicate robust production of anti-RBD IgG and IgM antibodies in humans following COVID-19 infection. IgG1 and IgG3 neutralization resulted in blocking binding of the virus to the Ace2 receptor required for viral entry and replication. Immunotherapy using purified/enriched anti-RBD IgG1 and IgG3 from convalescent plasma may improve disease outcome by (1) affording protection and (2) minimizing patient encounters with potentially harmful components present in whole plasma currently used in transfusion therapy.
Introduction
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is the causative infectious agent responsible for the 2019 (COVID-19) pandemic. Entry of the virus into human cells is mediated by the spike protein S1 subunit receptor binding domain (RBD) located on the surface of the virus which binds to the Angiotensin-Converting Enzyme 2 (ACE2) receptor present on multiple human cell membrane surfaces.1–3 Convalescent plasma (CP) containing neutralizing antibodies (nAbs) has been used therapeutically to treat severe cases in which patients often presented with excessive levels of cytokines and ineffective T and B cell responses. Passive immunity acquired from the nAbs found in CP has been shown to bind to SARS-CoV-2 preventing viral entry, thus reducing viral load.
Generally, IgM is the first isotype detected following initial infection with levels peaking about 2 weeks post-infection, and declining 1–3 weeks later. Secretory IgA, the most abundant mucosal antibody, plays an important role in control of pathogen insult at the mucosal surface, for example, lungs and gut. However, it appears that the predominant antibody generated in response to SARS-CoV-2 is IgG with levels peaking at about 3–5 weeks post-infection, remaining in circulation for approximately 8 weeks after which levels begin to gradually decline. These neutralizing antibodies not only provide protection against SARS-CoV-2 re-infection but their titer and neutralization capacity constitute a predictor of survival. 4
In this study, blood donor plasma from the San Antonio area (South Texas, USA) was procured when the prototype SARS-CoV-2 was the dominant circulating infectious strain, and was assessed as to (1) isotype and quantitation, that is, titer of elicited antibody to the SARS-CoV-2 S1 subunit RBD, and (2) determination of the presence of neutralizing antibodies in the plasma. With subsequent emergence of additional variants, that is, delta and omicron, the study was expanded to include the assessment of variant neutralization capacity of this convalescent plasma.
Materials and methods
Plasma
Samples (120) were obtained from the Food and Drug Administration (FDA)-registered QualTex Laboratories, a subsidiary of BioBridge Global, in San Antonio, Texas, United States. COVID-19 positive (100 samples numbered 1–100) and negative (20 samples numbered 101–120) status was determined by the Abbott Alinity I assay which detects IgG antibodies to the nucleocapsid protein of the SARS-CoV-2 virus. These human plasma samples were de-identified and personal information (gender, age, ethnicity, blood type, etc.) is not available for inclusion.
Determination of antibody isotype and titer
A standard indirect Enzyme-Linked Immunosorbent Assay (ELISA) was used to determine antibody titers. Specifically, microplates (96-well) were coated with glycosylated recombinant SARS-CoV-2 RBD (Acrobiosystems, 10 ng/100 µL per well) or coating buffer (sodium bicarbonate pH 9.5) alone as the no antigen control. Plasma samples (primary antibody source) were serially diluted two-fold starting at 1:100 for determination of total IgG titer, and 1:50 for all other isotype titers. Antibody detection was performed using horse radish peroxidase (HRP)-conjugated goat anti-human anti IgM, IgA, IgG(H + L), IgG1, IgG2, IgG3, IgG4 secondary antibodies (SouthernBiotech, Birmingham, AL , USA) at a dilution of 1:5000. Substrate (3,3’, 5,5’-tetramethylbenzidine peroxidase, TMB) was added to each well, and incubated for 15 min at room temperature. Reactions were terminated by addition of 50 µL 2N H2SO4, and the HRP redox driven color change was monitored at 450 nm. Endpoint titers were determined according to the method of Frey and coworkers. 5 In addition, plasma samples were diluted 1:400 and reacted with glycosylated variant RBDs (GenScript) including B.1.1.7 (United Kingdom, UK “alpha” strain), B.1.351 (South African, SA “beta” strain), and B.1.617.2 (Indian, “delta” strain) to assess SARS-CoV-2 elicited total IgG binding to emerging variant RBDs using the ELISA described.
Antibody neutralization
Neutralization capacity was determined using a commercially available SARS-CoV-2 surrogate virus neutralization test (sVNT) kit (GenScript, Piscataway, NJ, USA) per manufacturer’s instructions. Briefly, plasma (10-fold diluted) or control (positive and negative control provided in the assay kit) was incubated with HRP-conjugated RBD at 37°C for 15 min and then added to the microplate well pre-coated with the hACE2 protein. After 15 min of incubation, hACE2-bound HRP-RBD was detected colorimetrically by addition of TMB substrate. Results of neutralization capacity are expressed as percent inhibition and samples with greater than 30% inhibition are considered positive for neutralization activity according to manufacturer’s setting. Emergent variant strain antibody neutralization was assessed similarly by substituting prototype HRP-RBD with HRP-conjugated B.1.617.2 (Indian, “delta” strain) and B.1.1.529 (South African “omicron” strain) glycosylated recombinant RBDs (GenScript) in the assay. Correlation of percent inhibition to each isotype endpoint titer was determined according to the procedure of Akoglu. 6
Results
COVID-19 positive plasma exhibits high anti-RBD titer
In this study, the COVID-19 positive sample is defined by positive detection of SARS-CoV-2 nucleocapsid protein. As shown in Figure 1(a), COVID-19 positive plasma samples consistently exhibited higher endpoint anti-RBD IgG, IgA, and IgM titers when compared to negative control samples. A small number of control samples also exhibited antibody titers comparable to that observed for COVID-19 positive samples. Further analysis of IgG subclasses revealed that anti-RBD IgG antibody was predominantly IgG1 and IgG3 subclasses and minimally IgG2 and IgG4 (Figure 1(b)).
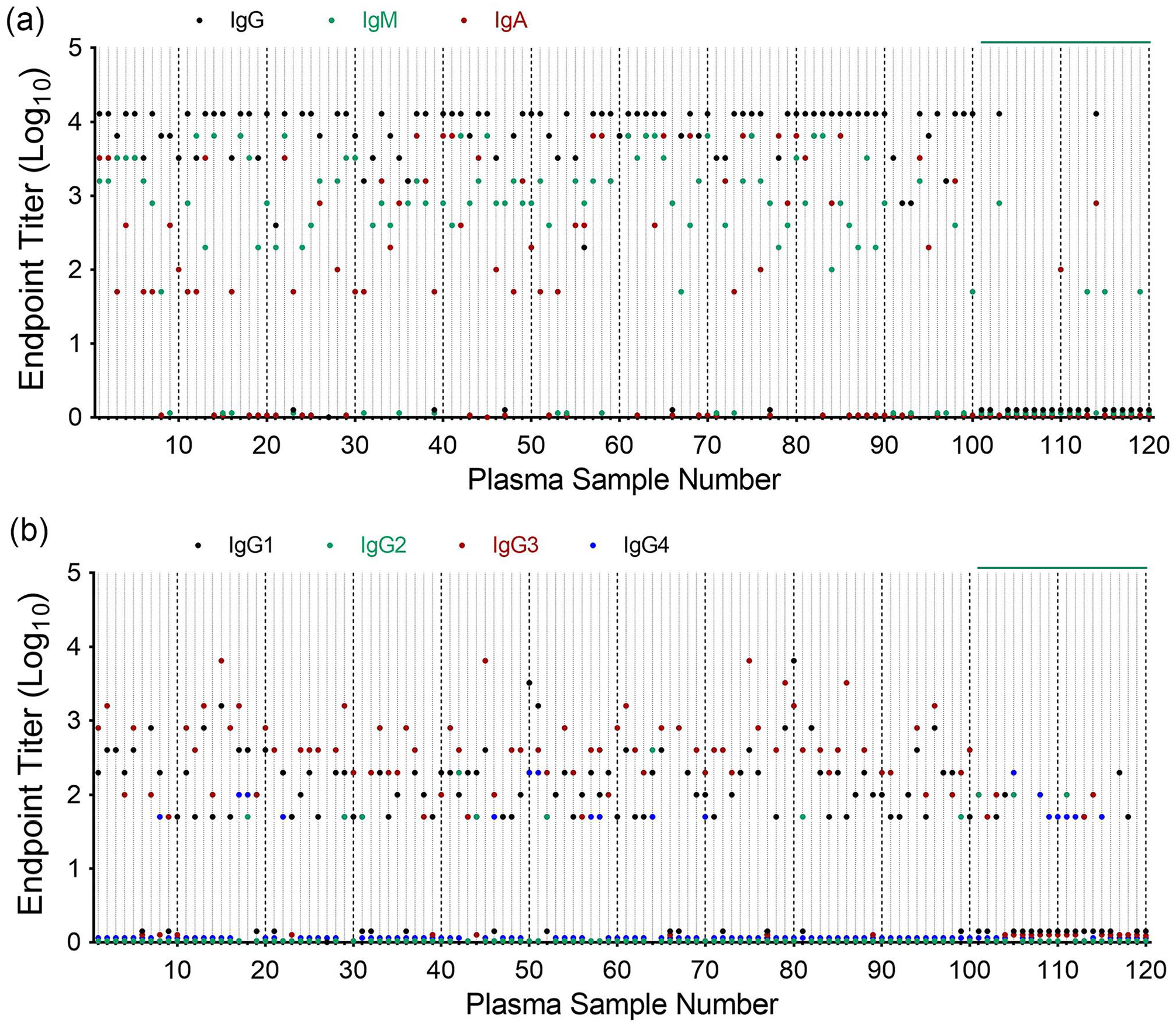
Anti-SARS-CoV-2 RBD titers in COVID-19 positive plasma. Plasma samples from COVID-19 positive (No. 1–100) and negative (No. 101–120) donors were assessed for anti-RBD immunoglobulin titer: (a) IgG, IgM, and IgA. (b) IgG subclasses, IgG1, IgG2, IgG3, and IgG4.
Plasma neutralization capacity correlates with anti-RBD IgG1 and IgG3 titers
Using a surrogate virus neutralization assay, the SARS-CoV-2 positive samples exhibited significant neutralization capacity (mean = 76.7%), that is, prevented binding of SARS-CoV-2 HRP-RBD to immobilized ACE-2 receptor when compared to SARS-CoV-2 negative control plasma (mean = 27.1%; Figure 2(a)). Among major antibody isotypes, anti-RBD IgG titer exhibits a “very strong” correlation (interpreted according to the report by Akoglu) 6 with SARS-CoV-2 neutralization capacity (i.e. % Ace2 binding inhibition) showing a Pearson’s correlation coefficient (r) of 0.83. SARS-CoV-2 neutralization was also positively correlated with anti-RBD IgM (r = 0.58, moderate) and IgA (r = 0.43, fair) titer (Figure 2(b)). Further assessment of IgG subclass revealed that correlation is moderate for both IgG1 (r = 0.74) and IgG3 (r = 0.71) while IgG2 (r = −0.06) and IgG4 (r = −0.14) did not correlate with SARS-CoV-2 neutralization (Figure 2(c)).
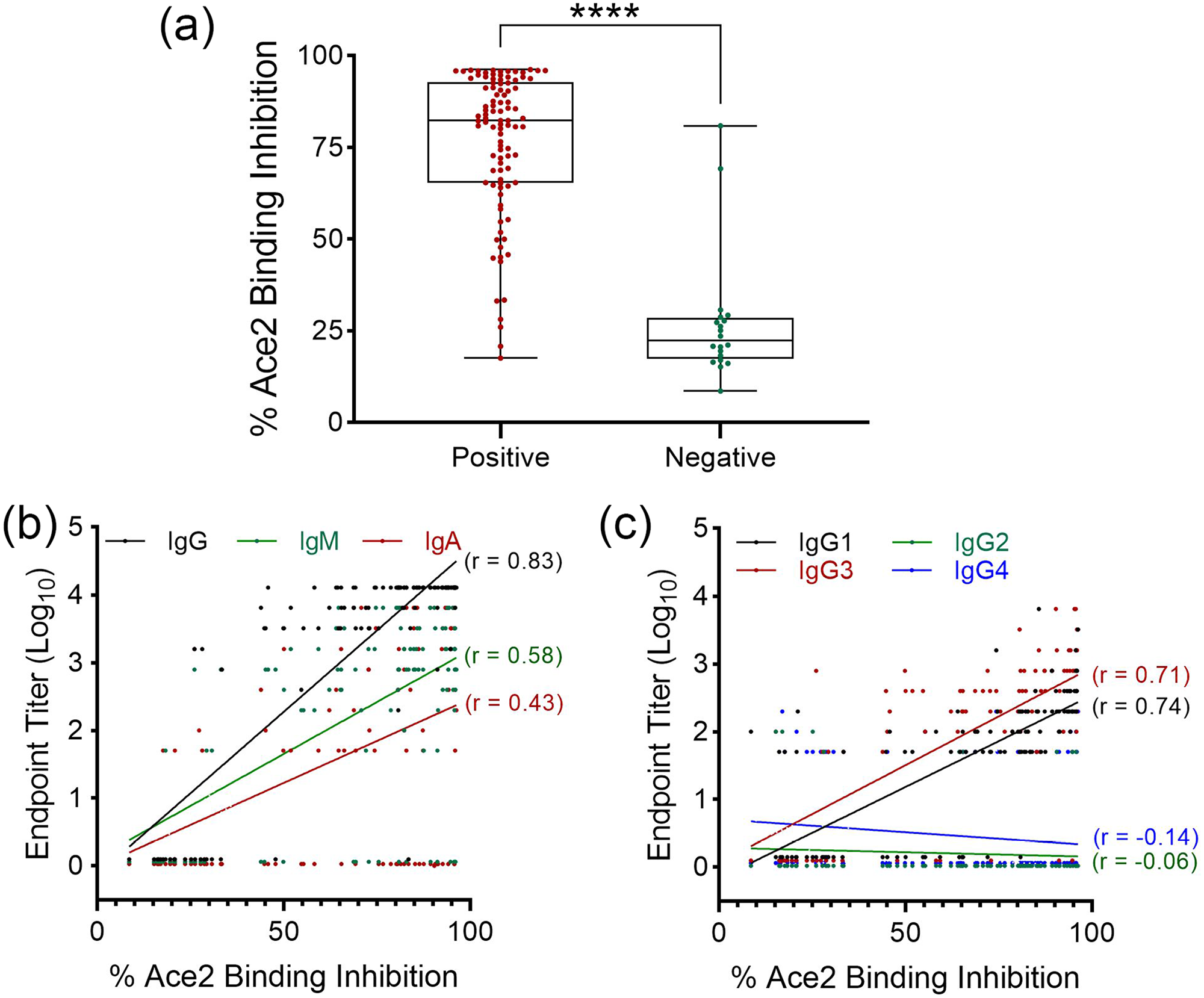
Correlation of SARS-CoV-2 neutralization with anti-RBD antibody titer. (a) A SARS-CoV-2 surrogate virus neutralization test was used to measure the capacity of COVID-19 positive and negative plasma to inhibit viral RBD binding to host Ace2 receptor. ****P < 0.0001, t-test. Correlation of plasma neutralization capacity (% Ace2 binding inhibition) with (b) major anti-RBD antibody isotypes, and (c) IgG subclasses was determined by Pearson’s Correlation Test. Correlation coefficient (r) is indicated for each test.
Plasma neutralization capacity against variant RBD binding to hACE2 is reduced
Plasma samples used in this study were collected at a time when SARS-CoV-2 was the dominant viral strain circulating in the United States. However, since the initial procurement of plasma, SARS-CoV-2 variants have emerged with various amino acid mutations within the RBD (Figure 3(a)). Assessment of anti-RBD reactivity in our plasma samples revealed a significant reduction in binding to variant RBDs (including alpha, beta, and delta strains) as shown in Figure 3(b) and (c) for individual and collective plasma, respectively. The median (and mean) plasma antibody binding values for SARS-CoV-2, alpha, beta, and delta RBDs were 0.67 (0.71), 0.46 (0.53), 0.10 (0.24), 0.24 (0.38), respectively. Consistent with decreased total IgG binding to variant RBDs was decreased neutralization by SARS-CoV-2 convalescent plasma of the delta variant RBD (Figure 4(a)). The median (and mean) percent inhibition for SARS-CoV-2 and delta RBD binding with Ace2 is 75 (71) and 63% (59%), respectively. With the recent surge of omicron variant infection, we further compared the neutralization activity of plasma against SARS-CoV-2 with that of the omicron variant. As shown in Figure 4(b), there are 15 mutations in the omicron RBD compared to SARS-CoV-2, and prevention of omicron RBD binding to Ace2 by SARS-CoV-2 plasma is greatly reduced from a median of 60 to 4.8%, and mean of 63 to 5.7% for SARS-CoV-2 and omicron, respectively. Furthermore, using a 30% inhibition as the cutoff value (defined by manufacturer), 91, 81, and 0% of SARS-CoV-2 positive plasma are considered positive for neutralizing CoV-2 prototype, delta, and omicron variant, respectively.
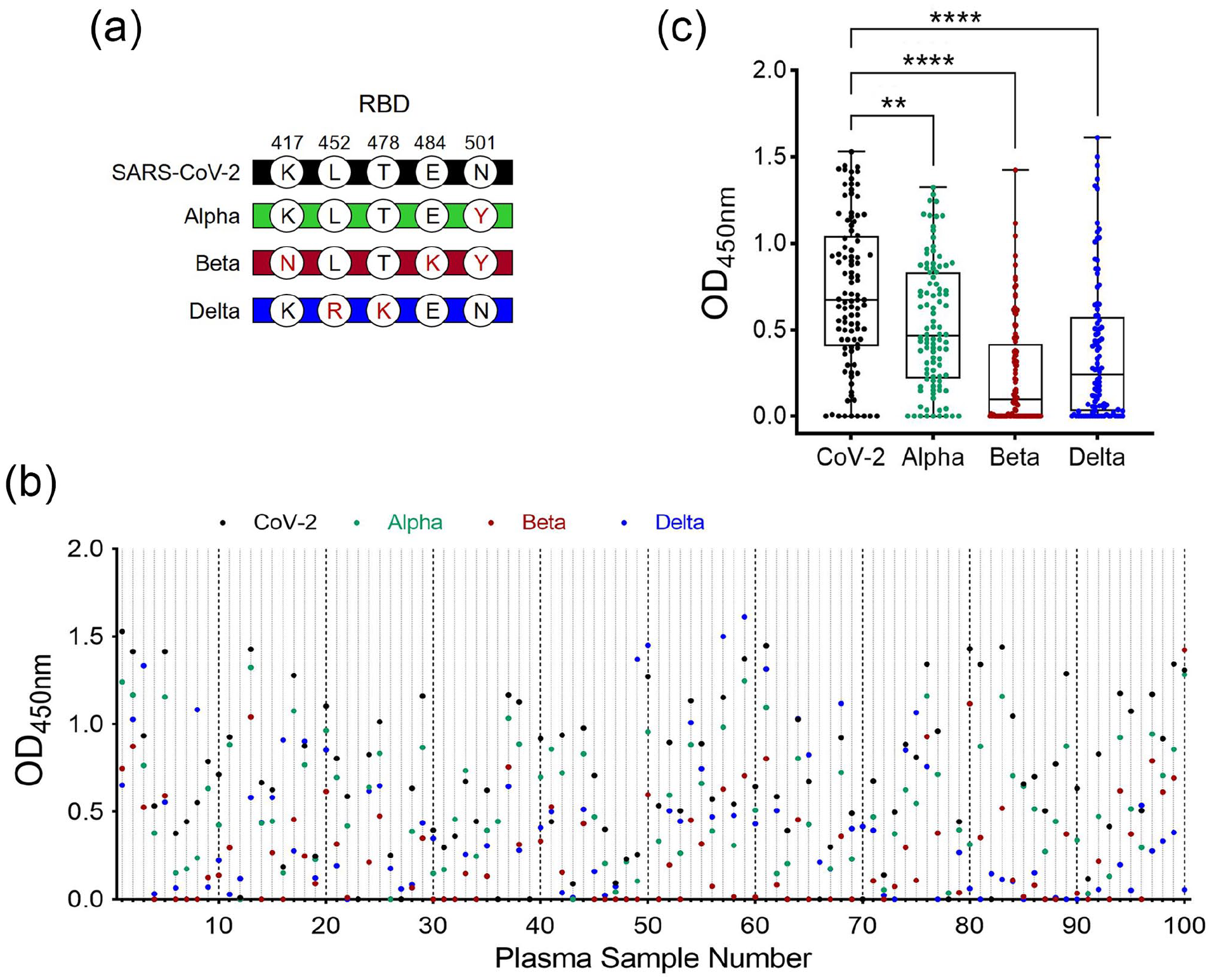
Comparison of SARS-CoV-2 elicited total IgG binding to emerging variant RBDs. (a) Amino acid substitutions (position indicated) within the spike protein receptor binding domain (RBD) in alpha, beta, and delta variants of the original SARS-CoV-2 strain. (b) Individual and (c), collective anti-RBD IgG binding was assayed using a standard indirect ELISA, and reported as optical density at 450 nm. COVID-19 positive plasma samples were diluted 1:400 and reacted with indicated variant RBD. Differences between indicated groups were analyzed by one-way ANOVA with Tukey’s multiple comparison test. **P < 0.01, and ****P < 0.0001.
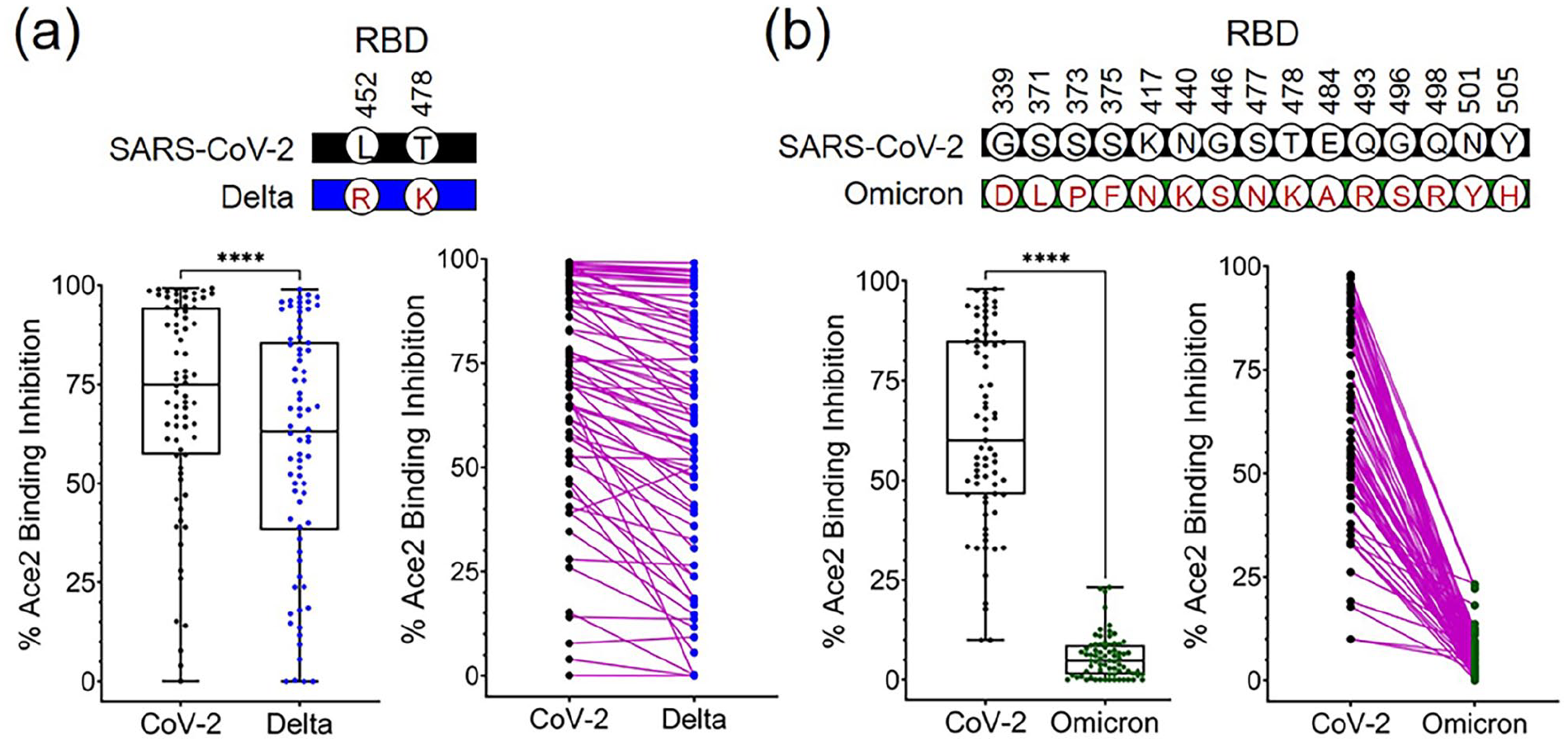
Reduction of delta and omicron variant neutralization was observed in SARS-CoV-2 positive plasma. A SARS-CoV-2 surrogate virus neutralization test was used to measure the capacity of COVID-19 positive plasma to inhibit SARS-CoV-2 and variant (a) Delta and (b) Omicron RBD binding with host Ace2 receptor. Percent inhibition of RBD binding to Ace2 receptor is presented as collective plasma (left panel) and paired individual plasma (right panel). Amino acid substitutions within RBD of delta and omicron strains are indicated, respectively. ****P < 0.0001, t-test. (A color version of this figure is available in the online journal.)
Discussion
The focus of this study was partial characterization of the immune response to SARS-CoV-2 using convalescent plasma. It has been shown that antibodies elicited in response to the SARS-CoV-2 receptor binding domain (RBD) are effective in preventing viral entry, thus halting viral replication and decreasing its propagation. We report here a diverse antibody response to the SARS-CoV-2 spike protein RBD in convalescent patient plasma indicating significantly higher anti-RBD titers for multiple antibody isotypes, that is, IgA, IgM, and IgG when compared to SARS-CoV-2 negative plasma controls which is consistent with the findings of Noval and coworkers. 7 Further subtyping of IgG indicated IgG1 and IgG3 subclasses to be most abundant. Greater than 90% of SARS-CoV-2 positive convalescent plasma samples exhibited significant IgG neutralization capacity using a surrogate virus neutralization assay. Antibody response to SARS-CoV-2 has been observed to exhibit variable neutralization capacities.8–11 Of the IgG subclasses, IgG1 and IgG3 exhibited highest viral neutralization capacity; whereas, IgG2 and IgG4 viral neutralization was not observed. Positive correlation between IgG titers and neutralization are consistent with previously published findings.9,12 It has been suggested that individuals who elicit IgG in combination with IgM and IgA antibodies against the RBD have greatest neutralization capacity. 7 Thus, the patient plasma examined here indicates a broad antibody repertoire, that is, IgG, IgA, and IgM following exposure to SARS-CoV-2 which may contribute to more effective neutralization and protection. It is noted that the two control plasma samples (number 103 and 114, Figure 1(a)) showing high anti-RBD IgG antibody titer also displayed high neutralization capacity. Individuals who have contracted seasonal human coronaviruses may generate circulating IgG antibodies that cross-react with SARS-CoV-2 antigens. Indeed, Ng et al. have reported that a small proportion of SARS-CoV-2 uninfected individuals did have IgG antibodies that reacted with the more conserved S2 subunit of the SARS-CoV-2 spike protein. However, COVID-19 patients elicit IgA, IgG, and IgM antibodies that recognized both the S1 (including RBD) and S2 subunits. 13
Several SARS-CoV-2 RBD variants, that is, alpha (N501Y), beta (K417N, E484K, and N501Y), delta (L452R and T478K), and omicron (15 mutations) have been reported causing considerable concern. Substituting commercially available glycosylated recombinant variant RBDs for that of the SARS-CoV-2 RBD, convalescent plasma was observed to bind to a lesser extent than that of SARS-CoV-2 RBD. Consistent with decreased variant RBD binding, decreased binding of the delta variant was associated with decreased neutralization. Antibody neutralization against omicron was further reduced due to the high number of amino acid substitutions within RBD. Although neutralizing capacity was determined using a surrogate-recombinant RBD viral format and may differ from that derived for viral particles, data reported here support the varied nature of the immune response to SARS-Cov-2 and is representative of a snapshot of antibody response to COVID-19 infection absent donor specifics. A study by Gaebler et al. provided insight into long term antibody immunity in COVID-19 patients revealing that titers of IgM and IgG antibodies against RBD decrease significantly over a 6.2-month period with neutralizing activity in plasma decreasing by five-fold. 14 However, antibodies secreted by RBD-specific memory B cells have greater somatic hypermutation, resistance to RBD mutations, and increased potency supporting the benefit of protection against SARS-CoV-2 by vaccination.
Footnotes
Authors’ Contributions
All authors participated in the design, interpretation of the studies and analysis of the data. JJY, AKM, JPC, and BPA conceived and designed the experiments. KAG and SBP carried out the experiments. KAG, JJY, JPC, and BPA analyzed the data. RB provided materials. KAG, JJY, JPC, and BPA wrote and edited the manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by The Jane and Roland Blumberg Endowment fund to BPA.