
Abstract
Hemolytic diseases such as Sickle Cell Disease (SCD) are characterized by a natural propensity for both arterial and venous thrombosis. The ability of heme to induce tissue factor (TF) activation has been shown both in animal models of SCD, and in human endothelial cells and monocytes. Moreover, it was recently demonstrated that heme can induce coagulation activation in the whole blood of healthy volunteers in a TF-dependent fashion. Herein, we aim to further explore the cellular mechanisms by which heme induces TF-coagulation activation, using human mononuclear cells, which have been shown to be relevant to in vivo hemostasis. TF mRNA expression was evaluated by qPCR and TF procoagulant activity was evaluated using a 2-stage assay based on the generation of activated factor X (FXa). Heme was capable of inducing both TF expression and activation in a TLR4-dependent pathway. This activity was further amplified after TNF-α-priming. Our results provide additional details on the mechanisms by which heme is involved in the pathogenesis of hypercoagulability in hemolytic diseases.
Impact Statement
Hemolysis is associated with an increased risk of thromboembolism. Heme can induce tissue factor expression in endothelial cells and monocytes and induce coagulation activation in the whole blood of healthy volunteers in a TF-dependent fashion. However, little data is available in the peer-reviewed literature about the mechanisms by which heme activates TF. In this study, we confirm that heme-induced coagulation activity is dependent on mRNA expression and on activation of hematopoietic TF in human mononuclear cells, and we provide additional details about these mechanisms, including their dependency on TLR-4 and crosstalk with other inflammatory pathways.
Introduction
Clinical and experimental data clearly demonstrate that hemolytic anemias such as Sickle Cell Disease (SCD), β thalassemia, paroxysmal nocturnal hemoglobinuria, and autoimmune hemolytic anemia are associated with hypercoagulability.1–3 Heme is an essential molecule present in almost all forms of life. However, evidence gathered in the last two decades shows that free heme, which can be released from hemoglobin during intravascular hemolysis, can be toxic to cells by both direct (via Fenton reaction and reactive oxygen species generation) and indirect (innate immunity-mediated) mechanisms. 4 The effects of heme on several compartments of innate immunity have been described over the last two decades, leading to the concept that free heme can behave as an erythrocyte danger associated molecular pattern (DAMP), through the activation of TLR4 receptors.5,6 Since hemostasis is currently regarded as part of innate immunity,7,8 heme has also been studied as a potential mediator of hypercoagulability.
In this regard, heme has been shown to induce the expression of tissue factor (TF) in cultured endothelial cells, 9 and was subsequently characterized as a mediator of coagulation activation in vivo in mouse models of SCD. 10 In the same study, Sparkenbaugh et al. 10 also demonstrated that heme can induce TF activation in human monocytes. Moreover, heme has also been shown to induce the formation of neutrophil extracellular traps, which facilitate coagulation activation during thromboinflammation.3,11 Recently, we demonstrated that incubation of heme in whole blood from healthy volunteers triggers coagulation activation in a TF-dependent fashion. 12 Together, these results support the concept that free heme is involved in the initiation and/or maintenance of hypercoagulability in SCD and other hemolytic conditions.
Yet, the mechanisms by which heme activates TF are not entirely clear. Here we characterized in more detail the effect of heme on TF mRNA expression and activation in human peripheral blood mononuclear cells (PBMCs) and explored the effects of TNF-α and TLR-4 in this process.
Materials and methods
Obtention of PBMC from blood donors
This study was performed in accordance with the Declaration of Helsinki and approved by the Ethics Committee of the University of Campinas (CAAE: 46853115.4.0000.5404). Peripheral blood mononuclear cells (PBMCs) were obtained from buffy coats as previously described,13,14 using de-identified blood bags provided after whole blood fractionation from the blood center of the University of Campinas (Hemocentro). Bags containing residual buffy coats were conserved at 4°C overnight and quickly preprocessed the next day. PBMCs were isolated with 1.077 density of ficoll (Ficoll-paque Plus, GE Healthcare) after dilution of the obtained buffy coat with PBS 1× (1:3 dilution volume:volume). After separation of PBMCs, cells were seeded in culture plates at a quantity equivalent to 5*106 and 2*106 monocytes (based on automated cell counts in a hematology analyzer) for quantitative PCR (qPCR) and functional coagulation assays, respectively. Cells were then incubated at 37°C for 4 h with heme or vehicle at different concentrations. When necessary, cells were primed with 2 ng/mL of TNF-α at 37°C for 1 h or pre-treated with 5 ng/mL of TLR4 specific inhibitor Tak-242. After incubation, supernatants were removed to harvest only adherent cells, thereby enriching the monocyte content of the sample.15–18 Cells were washed with PBS and re-suspended in HBSA buffer to measure TF pro-coagulant activity (TF PCA) with a two-stage assay based on FXa generation or were lysed with Trizol for TF mRNA expression by qPCR. Pre-processed samples were stored at −80°C until total RNA extraction or TF PCA analysis.
Whole blood collection and plasma separation
Whole blood samples were collected from healthy volunteers by venipuncture in vacuum tubes containing 3.2% sodium citrate (BD Vacutainer Coagulation Tube, 2.7 mL, 0.109 M buffered sodium citrate). These samples were also treated with heme or vehicles. Samples were then incubated at 37°C for 4 h. Plasma samples were immediately processed to perform a one-stage coagulation assay.
Heme dilution
Heme (Ref H651-9; Frontier Scientific, USA) was diluted with 100 μL of NaOH 0.1M at a pH of ~11 to minimize the formation of heme aggregates. Then this solution was adjusted with 900 μL of ultra-filtered water to obtain an initial working solution at a concentration of 5 mM. This solution was then filtrated through a 0.22 μm filter and freshly used according to the scenario. It is worth noting that very small volumes of this concentrated heme solution were used in each experimental condition (6 μL for 1 mL of final volume of culture medium at a concentration of 30 μM heme, for example), preventing any change in the culture medium pH.
Gene expression quantification by qPCR
Total RNA was extracted using Trizol using classical methods. For each sample, 1 μg of RNA was transcribed using the High-capacity cDNA Reverse Transcription Kit (Applied Biosystems™). After transcription, cDNA was diluted with nuclease-free water, and qPCR experiments were performed in duplicate using an amplification mixture (12 µL) that contained 60 ng of template cDNA (3 µL of total volume), 6 µL of SYBR Green I Master Mix buffer (Applied Biosystems), and 150 nM forward and reverse primer. No-template controls were included in each plate. Reactions were run on an StepOnePlus™ System (Applied Biosystems, Life Technologies, Foster City, EUA). The cycling conditions comprised 10 min polymerase activation at 95°C, 40 cycles at 95°C for 15 s and 60°C for 1 min, and a 20 min step (60°C–95°C) for melting curve analysis. A standard curve of five serial dilution points composed of a mixture of all analyzed cDNA was analyzed to determine primer pair efficiency. All qPCR efficiencies were above 95%. TF gene (Gene symbol: F3; transcript targeted: ENST00000334047) expression was evaluated by delta-delta Cq method 19 using the geometric mean of two reference transcripts EEF2 (ENST00000309311) and MYH9 (ENST00000216181) provided by HT Atlas database. 20 These transcripts were shown to be among the most stably expressed in PBMCs and have been validated in our experiment. The stability of these reference transcripts was determined using the geNorm method 21 and outperformed two commonly used genes (GAPDH and ACTB) in this experiment. Primer sequences and concentration are described in Table 1. Primer pair specificity was confirmed using Blast and post-qPCR analysis of the melting curve.
Information of primer pairs.

F: forward primer; R: Reverse primer; bp: base pairs.
Two-stage TF PCA assay
TF PCA of adherent fractions of PBMCs were evaluated by a two-stage chromogenic assay based on the kinetics of FXa generation, as previously described. Briefly, 30 µL of treated cells suspended in HBSA were incubated with 20 µL of HBSA containing 150 nM FX, 1 nM FVIIa, and 3 mM CaCl2 for 2 h at 37°C. 22 In these experiments, an IgG control antibody, and an anti-TF antibody (HTF-1; Ref 16-1429-85 eBioscience) treatment (15 min of incubation at room temperature with 400 µg/mL final concentration) were used to measure the fraction of Xa generation that is dependent on TF. After incubation, FXa generation was stopped by 25 μL of EDTA buffer (25 mM) for 10 min, and then 25 μL of 0.3 mM FXa substrate Pefachrome sFXa (Pefachrome®) was added for absorbance reading at 405 nm every 30 s, for 30 min. Absorbance was referred to an Innovin (2.5–0.02 nM) standard curve. Results were expressed as the equivalent of TF (in pg/mL) to the procoagulant activity from IgG treated samples subtracted from procoagulant activity in anti-TF treated samples.
One-stage clotting test
Heme-induced procoagulant activity of plasma was measured using a one-stage clotting test performed using a semi-automated coagulometer (Max Start 4, Diagnostica Stago) after pre-incubation of heme at 37°C with normal human plasma, followed by re-calcification as previously described. 23 The procoagulant activity was determined using standard curve generated with a recombinant human relipidated TF (Innovin, Dade Behring). The results were reported in units of TF activity. In 50 µL of test sample, one unit of TF activity was arbitrarily defined as the amount of TF which yields a 50 s clotting time with pooled normal plasma. 24
Statistical analysis
Experiments were repeated at least 3 times, and results were expressed as the median of each variable. Statistical analysis was performed using GraphPadPrism 6.0 (GraphPadPrism Software Inc., San Diego, California, USA). Wilcoxon two-side or Friedman tests were used to compare quantitative parameters obtained from the same samples exposed to vehicle or heme, with or without TNF-α. A correction for multiple comparisons was performed using Dunn`s test. A P-value of < 0.05 was considered significant.
Results and discussion
Heme induces TF gene expression in PBMCs
Heme has been previously shown to induce procoagulant activity in whole blood, 12 and to activate TF activity in monocytes. 10 Here we explored whether this effect was dependent on hematopoietic TF gene expression. When compared to vehicle, heme significantly induced TF gene expression measured by qPCR in PBMC after 4 h of incubation (Figure 1). The induction was evident with heme concentrations above 10 μM, suggesting a dose-dependent effect.
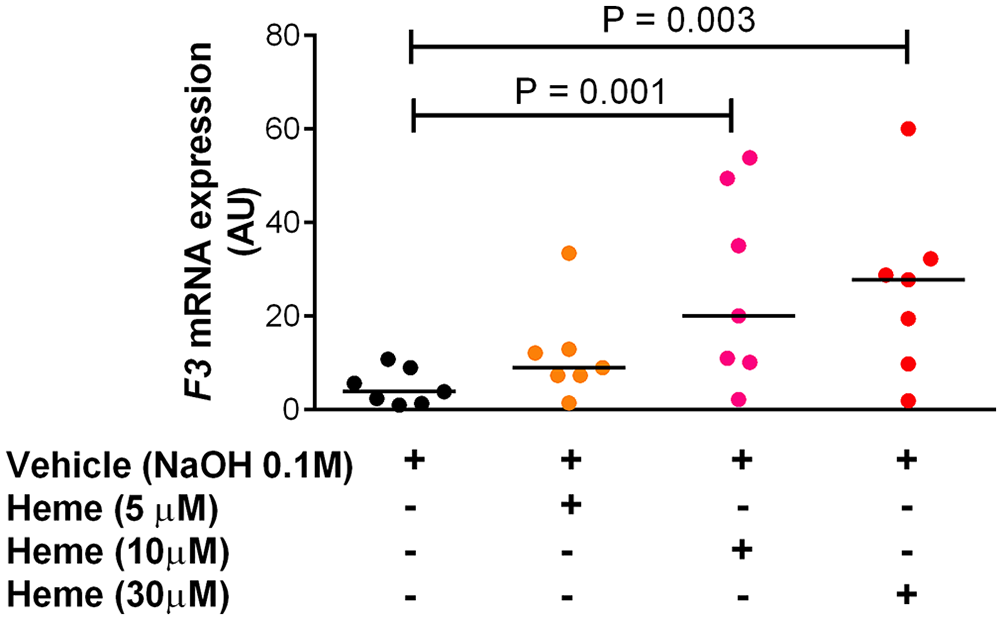
Evaluation of F3 mRNA expression in mononuclear cells by qPCR. PBMCs were seeded in culture plates at a quantity equivalent to 5 × 106 monocytes. After 4 h of incubation with vehicle and heme at different concentrations ranging from 5 µM to 30 µM, only adherent cells were used to perform qPCR. Results were expressed in arbitrary units (AU) as median of normalized F3 expression using two stably expressed reference transcripts EEF2 (ENST00000309311) and MYH9 (ENST00000216181). Comparisons were made between vehicle and increasing concentrations of heme and showed that heme induces TF mRNA expression in a dose response fashion. Friedman test was performed, and multiple comparisons were corrected using the Dunn`s test; a P-value of < 0.05 was considered significant. Sample from 7 different participants were analyzed. (A color version of this figure is available in the online journal.)
Heme triggers TF procoagulant activity
We next explored the effects of heme on TF activation. Heme 30 μM induced a significant increase in TF procoagulant activity of PBMCs when compared to vehicle, and this effect was enhanced by priming cells with TNF-α prior to heme stimulation (Figure 2(A)). We also demonstrated that heme-induced TF activation seems to be dose dependent (Figure 2(B)). TNF-α alone at a concentration of 2 ng/mL was not able to induce TF procoagulant activity.

Heme-induced procoagulant activity is potentiated by TNF-α. PBMCs were seeded in culture plates at a quantity equivalent to 2 × 106 monocytes. After 4 h of incubation with vehicle or heme, adherent cells were harvested and used to perform the experiments. Comparisons were made for heme versus vehicle. When indicated cells were primed with TNF- α. Results were ’presented as median of procoagulant activity in ng/mL. Heme-induced TF procoagulant activity is potentiated by TNF-α-priming (A) as shown by the comparison between heme (30 μM) and heme (30 μM) + TNF-α; this heme effect is also dose dependent (B). Seven and eight samples were analyzed respectively in A and B. Wilcoxon two-tail test was performed to compare the variables and sample size was indicated in each panel. (A color version of this figure is available in the online journal.)
Since circulating TF and plasma components such as micro-vesicles carrying heme have been shown to be relevant to coagulation activation25,26 we hypothesized that heme might also be capable of inducing procoagulant activity in plasma. Accordingly, using a one-stage coagulometric assay, we were able to show that heme induces procoagulant activity in plasma separated from whole blood after 4 h of incubation with heme (Figure 3(A)).
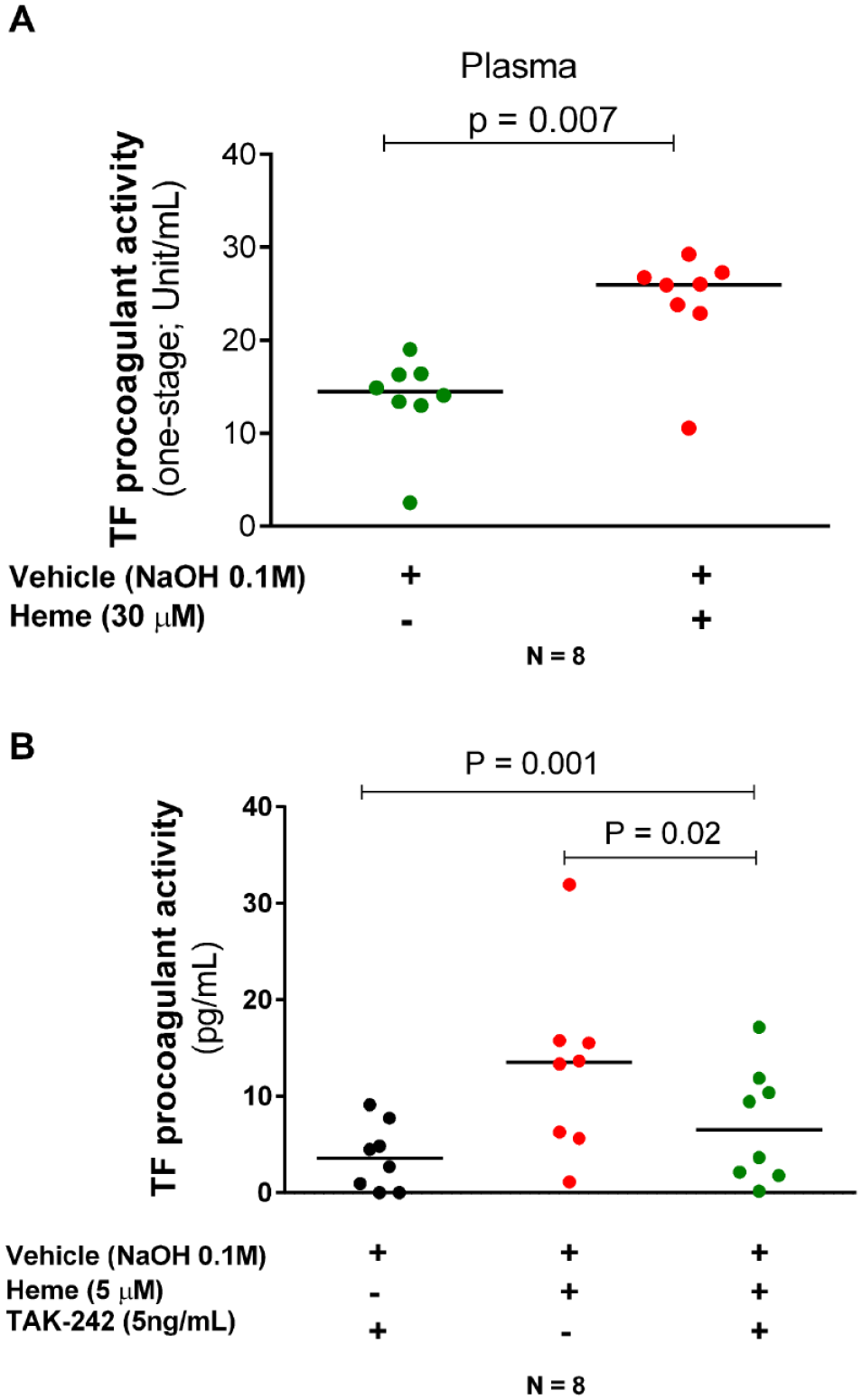
Heme-induced procoagulant activity is dependent on TLR4. Comparisons were made for heme versus vehicle. When indicated, cells were pre-incubated with Tak-242. The heme pro-coagulant effect was confirmed in plasma using a coagulometric assays (A) illustrating the relevance of circulating TF and plasma components. Tak-242, an inhibitor of TLR4, reverted heme-induced TF PCA (B). Data are presented as median units of TF activity (A) or TF procoagulant activity (B). A Wilcoxon two-tail test was performed to compare the variables. Eight samples were analyzed in each panel. (A color version of this figure is available in the online journal.)
TF activity induced by heme is dependent on TLR4
Heme is an erythrocyte DAMP that has been shown to be a potent inducer of inflammation and to trigger of vaso-occlusive events in animal models of SCD via TLR4 receptors.5,27,28 Accordingly, we explored whether heme triggers TF procoagulant activity through TLR4 by using Tak-242, which was capable of reverting heme-induced TF PCA (Figure 3(B)).
SCD is characterized by an increased risk of venous thromboembolism, 29 and the role of heme as a mediator of this risk was elegantly demonstrated in a study using an animal model of SCD and human monocytes, which demonstrated TF-dependent coagulation activation by heme. 10 Here we explored additional details of heme-induced coagulation activation in human mononuclear cells and demonstrated that this effect was dependent on both activation and mRNA expression and at least partially occurred in a TLR-dependent fashion. Since TLR4 inhibition only partially reduced TF PCA, this pathway is not likely the only mechanism by which heme could induce TF gene expression and/or PCA. Heme is known to regulate the expression of many genes. 4 Therefore, it wouldn’t be surprising if heme also induces TF expression by regulating its expression in a TLR4 independent manner involving another cell signaling. Furthermore, heme could also induce TF PCA by interacting with its decryption mechanism. These hypotheses must be further explored to gain additional insights into heme-induced TF expression and PCA.
Studies using endotoxemia models have previously shown that hematopoietic TF is more relevant for coagulation activation than endothelial TF. 30 Heme has been previously shown to be an activator of TF in human monocytes. 10 Here we provide additional details about the effects of heme on the expression and activation of hematopoietic TF, using two different functional assays and also demonstrating a dose-effect relationship.
The pathogenesis of hypercoagulability in SCD is multifactorial and involves crosstalk with other inflammatory pathways.3,31–33 In the present study, we also demonstrated that heme-induced TF PCA was potentiated by TNF-α. Of note, synergy of heme with other pro-inflammatory mediators have been previously described in experiments involving LPS stimulation 34 and in an in vitro model of NETosis in neutrophils pretreated with TNF-α. 11 However, as far as we are aware, there have been no reports of a priming effect of TNF-α in TF activation. Moreover, as described with other compartment of innate immunity,11,28 as well as reported in a meeting abstract, 35 we also demonstrated that heme-induced TF activation is TLR-dependent.
In conclusion, we provide additional details on the mechanisms by which heme triggers both TF mRNA expression and TF activation in human hematopoietic cells, reenforcing the interplay between hemostasis and inflammation in this process. Further studies are necessary to elucidate the mechanisms by which heme induces at low dose (5 μM) TF PCA but not its gene expression, as well as the synergistic mechanism of TNF-a.
Footnotes
Authors’ Contributions
BWH, CRPM, and CL performed experiments. BWH analyzed, interpreted data, and drafted the manuscript. MNNS interpreted data. FFC and EVDP contributed with reagents and laboratory infrastructure. EVDP designed the study, reviewed, and analyzed data and drafted the manuscript. All authors revised and approved all submitted versions of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
The study was performed in accordance with the Declaration of Helsinki and approved by the Ethics Committees of State University of Campinas (CAAE: 46853115.4.0000.5404).
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was financially supported by the Sao Paulo Research Foundation, grants # 2014/0984-3, 2015/24666-3, and 2016/14172-6; CNPq Brazil, grant # 309317/2016; and Coordenacao de Aperfeicoamento de Pessoal de Nivel Superior—Brasil (CAPES)—Finance Code 001.