
Abstract
Glycine from sow’s milk only meets 20% of the requirement of suckling piglets. However, how glycine is synthesized endogenously in neonates is not known. This study determined glycine synthesis from 4-hydroxyproline (an abundant amino acid in milk and neonatal blood) in tissues of sow-reared piglets with normal birth weights. Piglets were euthanized at 0, 7, 14 and 21 days of age, and their tissues were used to determine glycine synthesis from 0 to 5 mM 4-hydroxyproline, activities and mRNA expression of key glycine-synthetic enzymes, and their cell-specific localization. Activities of 4-hydroxyproline oxidase (OH-POX), proline oxidase (POX), serine hydroxymethyltransferase (SHMT), threonine dehydrogenase (TDH), alanine:glyoxylate transaminase (AGT), and 4-hydroxy-2-oxoglutarate aldolase (HOA) occurred in the kidneys and liver from all age groups of piglets, and in the pancreas of 7- to 21-day-old piglets. Activities of OH-POX and HOA were absent from the small intestine of newborn pigs but present in the small intestine of 7- to 21-day-old piglets and in the skeletal muscle of 14- to 21-day-old piglets. Between days 0 and 21 of age, the enzymatic activities of OH-POX, AGT, and HOA decreased in the liver and kidneys but increased in the pancreas and small intestine with age. The mRNA levels of these three enzymes changed in a manner similar to their enzymatic activities. In contrast to OH-POX, AGT, and HOA, the enzymatic activities of POX, SHMT, and TDH were present in the kidneys, liver, and intestine of all age groups of piglets. Glycine was synthesized from 0.1 to 5 mM 4-hydroxyproline in the liver and kidney from 0- to 21-day-old piglets, as well as the pancreas, small intestine, and skeletal muscle from 14- to 21-day-old piglets in a concentration-dependent manner. Collectively, our findings indicate that 4-hydroxyproline is used for the synthesis of glycine in tissues of piglets to compensate for the deficiency of glycine in milk.
Impact statement
Glycine is the most abundant amino acid in mammals and is essential for the synthesis of proteins and many low-molecular-weight substances (including glutathione, creatine, purines, and heme) with enormous physiological significance. The milk of mammals (e.g. humans, swine, and cattle) is remarkably deficient in glycine (e.g. providing 20% of the requirements of suckling piglets). To date, it is unknown how glycine is synthesized in neonates. Using the piglet model, we found that glycine was synthesized from 4-hydroxyproline (an abundant amino acid in milk and neonatal blood) via the 4-hydroxyproline oxidase (OH-POX) pathway primarily in the liver and kidneys based on rates and, to a lesser extent, in the pancreas, small intestine, and skeletal muscle in an age-dependent manner. This metabolic pathway helps to compensate for glycine deficiency in milk to support neonatal growth, development, and survival. These results have broad and important implications for the nutrition of human infants and other mammalian species.
Introduction
Glycine comprises about 30% of the collagen and elastin in the body and participates in multiple essential biochemical reactions, including the formation of glutathione, creatine, heme, purines, and bile salts. 1 Thus, young animals have particularly high requirements for glycine. 2 However, the milk of mammals studied, including swine, humans, cattle, sheep, and rats, is severely deficient in glycine.3–5 Results of recent studies have shown that glycine from sow’s milk only meets 20% of the requirements of suckling piglets and that the neonates must synthesize 80% of the needed glycine. 1 We have estimated that serine, threonine, and choline in milk provide 12% of the piglet’s total glycine requirement. 1 Clearly, additional substrate(s) must serve as precursors for glycine synthesis in piglets but are currently unknown.
There are reports that dietary trans-4-hydroxy-L-proline (4-hydroxyproline) is oxidized to glyoxylate and oxalate in the liver and kidneys of rats6,7 and humans. 8 Glyoxylate can be converted into glycine by alanine:glyoxylate transaminase (AGT) in these two tissues.6,7,9,10 Thus, 4-hydroxyproline can be converted to glycine in animals. In support of this view, results of limited research indicate that glycine is released from rat kidneys perfused with 4-hydroxyproline 11 and that 14 C-glycine is formed by HepG2 cells incubated with 14 C-glycolate. 12 At present, little is known about 4-hydroxyproline metabolism or glycine synthesis in the neonates of any species. Therefore, this study was conducted to determine if 4-hydroxyproline (an abundant amino acid in sow’s milk and neonatal blood) 13 is used for glycine synthesis via the proposed OH-POX pathway in tissues of sow-reared piglets (Figure 1). To achieve this goal, a series of experiments were performed to determine enzymatic activities, abundances of mRNAs and proteins for key enzymes, the cell-specific localization of enzymes, and the synthesis of glycine from 4-hydroxyproline by tissues of 0- to 21-day-old piglets with normal birth weights. For comparison with the expression of OH-POX, we also determine the activities of proline oxidase (POX) as well as serine hydroxymethyltransferase (SHMT) and threonine dehydrogenase (TDH) in the same tissues.14–17
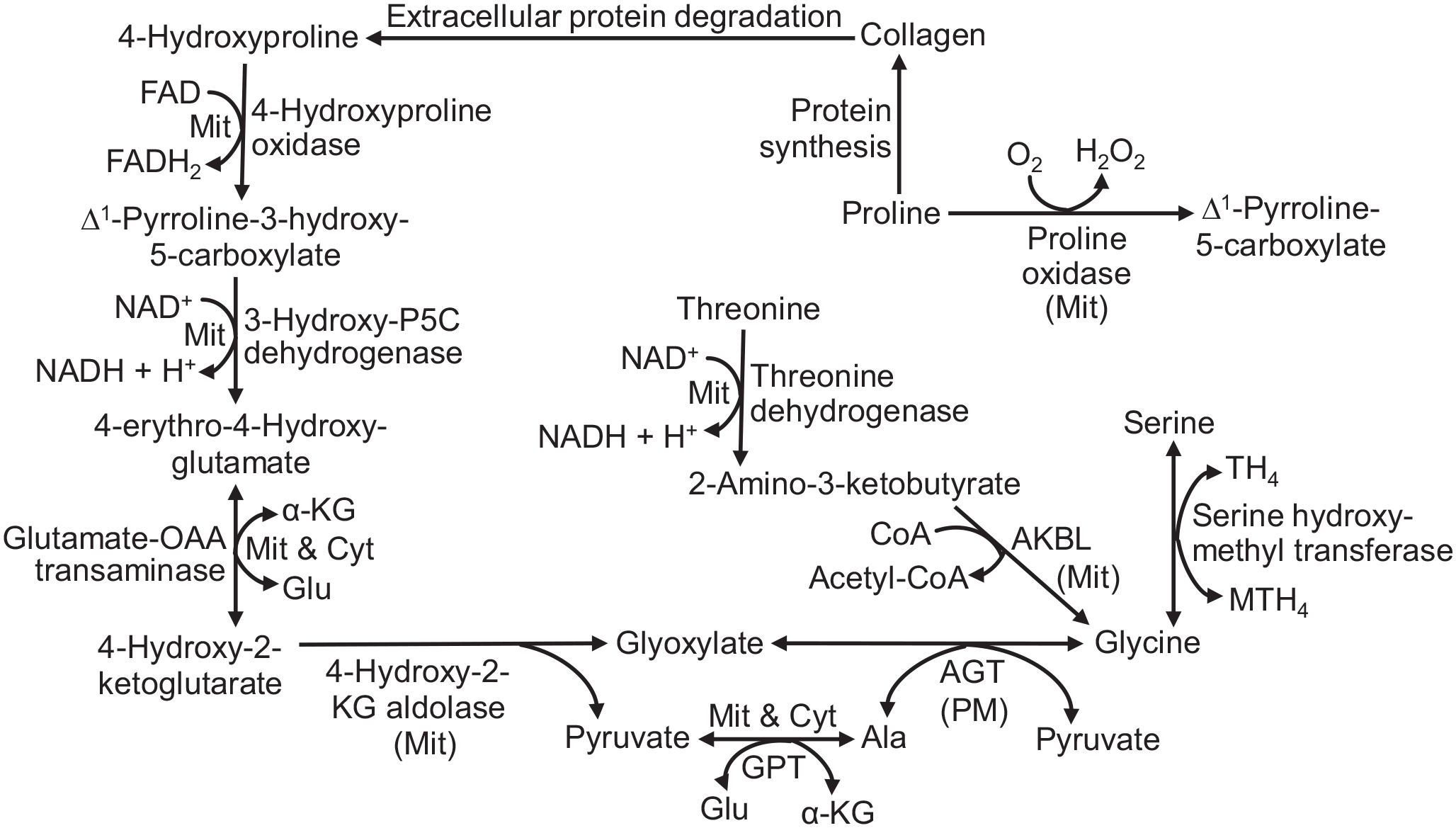
The proposed 4-hydroxyproline-glycine pathway in tissues of neonatal pigs. 4-Hydroxyproline is oxidized to glycine via this series of enzymes. For comparison, the formation of glycine from serine and threonine is shown. The gene name for 3-hydroxy-pyrroline-5-carboxylate dehydrogenase is ALDH4A1. Ala, alanine; AGT, alanine:glyoxylate transaminase; AKBL, 2-Amino-3-ketobutyrate lyase; Cyt, cytosol; α-KG, α-ketoglutarate; Glu, Glutamate; GPT, glutamate: pyruvate transaminase; Mit, mitochondria; MTH4, N5-N10-methylenetetrahydrofolate; OAA, oxaloacetate; P5C, Δ1-pyrroline-5-carboxylate; PM, peroxisome; Pyr, pyruvate; TH4, tetrahydrofolate.
Materials and methods
This study was approved by the Texas A&M University Institutional Animal Care and Use Committee.
Chemicals
High-performance liquid chromatography (HPLC)-grade methanol and water were obtained from Fisher Scientific (Houston, TX). Amino acid standards [including glycine and trans-4-hydroxy-L-proline (4-hydroxyproline)], and other chemicals were purchased from Sigma-Aldrich (St Louis, MO).
Animals
Piglets were the offspring of Yorkshire × Landrace sows and Duroc boars. All pigs were maintained at the Texas A&M University Swine Center. The average litter size of sows was 12.4 piglets at birth. Throughout the gestation and lactation periods, sows had free access to water and corn-soybean meal-based diets 18 that met the National Research Council (NRC) 19 requirements. On postnatal days 0, 7, 14, and 21, six piglets for each day of age with average body weights of 1.41 ± 0.12 (normal birth weight), 2.57 ± 0.09, 4.13 ± 0.16, and 5.51 ± 0.34 kg, respectively, were selected randomly for tissue collection.
Collection of tissues from piglets
At 0, 7, 14, and 21 days of age, after blood was sampled from piglets, they were anesthetized with an intramuscular injection of Telazol (10 mg/kg body weight) and then euthanized by the intracardiac administration of saturated KCl, as we described previously.20,21 After the abdomen was opened, the heart, liver, lung, stomach (with luminal contents being removed), pancreas, spleen, jejunum (proximal half of the remaining small intestine below the duodenum, with luminal contents being removed), kidneys, gallbladder, skeletal muscle, skin, and brain were quickly isolated and weighted. Tissues were cut into small portions, and some were fixed in freshly prepared 4% (wt/vol) paraformaldehyde in phosphate-buffered saline (PBS) (pH 7.2) for 24 h, followed by storage in 70% ethanol for 24 h. 22 The fixed tissues were dehydrated through a graded series of alcohol to xylene and embedded in Paraplast-Plus (Oxford Labware, St. Louis, MO) for immunohistochemical analyses.
Slices of fresh tissues were placed in ice-cold Krebs-Henseleit bicarbonate (KHB) buffer before being used for incubation (see below). The remaining portions of the tissues were frozen in liquid nitrogen and stored at −80°C.
Production of glycine by tissues incubated with 4-hydroxyproline
The production of glycine by fresh tissues incubated with 4-hydroxyproline was determined using established methods as reported for studies of other metabolic pathways.23,24 Slices (~100 mg) of freshly isolated tissues from piglets were incubated at 37°C for 2 h in 2 mL of oxygenated (95% O2/5% CO2) KHB buffer containing 5 mM D-glucose, 20 mM HEPES (pH 7.4), and either 0, 0.1, 0.25, 0.5, 2, or 5 mM 4-hydroxyproline. The reaction was terminated by addition of 200 µL of 1.5 M HClO4. The tissue plus the incubation medium was homogenized for 2 min on ice, and the solution was then neutralized with 100 µL of 2 M K2CO3. After centrifugation at 600g for 5 min at 4°C, the supernatant fluid was used for HPLC analysis of amino acids. 25 In parallel experiments, concentrations of amino acids in tissues without incubation were also determined as previously described. 25 Results of our preliminary studies indicated that the rates of production of glycine from 4-hydroxyproline (e.g. 5 mM) by tissue slices of liver, pancreas, kidney, small intestine, and skeletal muscle from piglets (e.g. 21-day-old pigs) were linear with 2 h incubation. We did not use tissue homogenates to determine the formation of glycine from 4-hydroxyproline, because we found in our preliminary studies that its rates per gram tissue were only 30% to 40% of those obtained for tissue slices.
Analysis of enzymatic activities of tissues
Tissue homogenate
Frozen tissue (0.5 g) was homogenized in 2 mL of freshly prepared buffer consisting of 300 mM sucrose, 5 mM HEPES (pH 7.4), 1 mM ethylenediaminetetraacetic acid (EDTA), 3 mM dithiothreitol, 0.5% (vol/vol) Triton X-100, and 0.1% (wt/vol) protease inhibitor (aprotinin, chymostatin, pepstatin A, and phenylmethylsulfonyl fluoride; 5 µg/mL each) for 2 min on ice. 11 The whole homogenization mixture was transferred to a tube, which was centrifuged at 600g for 10 min at 4°C. The supernatant fluid was subjected to three cycles of freezing in liquid nitrogen and thawing in a 37°C water bath before use for assays of POX and the following enzymes.10,11,15 Maximum activities of all enzymes were measured under optimum conditions.11,15 Results of our preliminary studies with piglet tissues indicated that the assays of activities of all enzymes were linear with the time and amount of protein used in this study (Supplemental Table 1).
Activity of OH-POX
The assay for OH-POX, which is a rate-controlling enzyme for the 4-hydroxyproline-glycine pathway,26,27 was performed as described by Carnie et al. 10 with modifications. Briefly, the assay solution (3 mL) consisted of 40 mM potassium phosphate (pH 8.0), 150 mM 4-hydroxyproline, 8 µM cytochrome C (C2506; Sigma-Aldrich), and 0.3 mL of tissue homogenate. The solution was incubated at 37°C. After 0 and 30 min, 0.8 mL of the mixture was added to 1.6 mL of 40% ethanol and 6% trichloroacetic acid (TCA) which contained 1 mg/mL O-aminobenzaldehyde (A9628; Sigma-Aldrich). The mixture was allowed to stand for 30 min at room temperature and was then centrifuged at 600g for 10 min to acquire the supernatant fluid for measurement of optical density (OD) at the wavelength of 443 nm.
Activity of 4-hydroxy-2-oxoglutarate aldolase (HOA)
The activity of HOA in tissues was determined as previously described. 10 Briefly, the reaction solution (1 mL) contained 0.2 mL of homogenate, 0.2 mL of H2O or 100 mM DL-4-hydroxy-2-ketoglutarate (disodium salt), and 0.6 mL of 100 mM potassium phosphate buffer (pH 7.8). The mixture was incubated at 37°C for 0 or 20 min, and 1 mL of 10% TCA was added to stop the reaction. After the solution was centrifuged at 600g for 10 min, the supernatant fluid was analyzed for pyruvate as previously described. 28
Activity of AGT
The activity of AGT was determined by quantifying pyruvate production, as described by Rowsell et al. 9 Briefly, the assay solution (0.5 mL) contained 0.05 mL of tissue homogenate, 0.1 mL of H2O or 100 mM glyoxylic acid, 0.1 mL of 100 mM L-alanine, and 0.25 mL of 100 mM potassium phosphate buffer (pH 7.4). The mixture was incubated at 37°C for 0 or 20 min, and 1 mL of 10% TCA was added to stop the reaction. The solution was centrifuged at 600g for 10 min. Pyruvate in the supernatant fluid was analyzed using L-lactate dehydrogenase, as previously described. 28
SHMT
SHMT activity was measured as described by Lowry et al., 15 except that 20 mM serine rather than 14C-serine was used, and glycine was determined as an end-product of the enzymatic reaction. Briefly, the assay mixture (1 mL) contained 20 mM L-serine, 0.25 mM pyridoxal phosphate, 0.25 mM tetrahydrofolate, 10 mM β-mercaptoethanol, 75 mM potassium phosphate, and tissue extract (0.25 or 0.5 mg protein). The same assay mixture without tetrahydrofolate served as the blank. After a 15-min incubation at 37°C, the reaction was terminated by the addition of 0.2 mL of 1.5 M HClO4. The solution was neutralized by the addition of 0.1 mL of 2 M K2CO3. The extract was analyzed for glycine by HPLC. 25
Threonine dehydrogenase
TDH activity was measured as described by Chen et al. 29 The assay mixture (0.2 mL) for threonine dehydratase contained 30 mM potassium phosphate buffer (pH 8.2), 0.3 mM EDTA, 0.5 mM nicotinamide adenine dinucleotide phosphate (NADPH), 10 mM α-ketoglutarate, glutamate dehydrogenase, 0.1 mM pyridoxal phosphate, 0 or 25 mM L-threonine, and tissue extract (0.25 or 0.5 mg protein). Absorbance at 340 nm was determined at 0, 5, 10, and 15 min using a Molecular Devices Microplate Reader (Sunnyvale, CA).
Possibility of 4-hydroxyproline as a substrate for POX
Mitochondria were isolated from the fresh jejunal mucosae, kidney and liver of 21-day-old pigs and used for POX activity in the presence of 15 mM L-proline or 4-hydroxyproline, 20 µM ferricytochrome C, as we described previously. 17 The absorbance of assay solution (1 mL) after the sequential addition of 0.5 mL of 10% TCA and 0.1 mL of O-aminobenzaldehyde was measured at 440 nm.
RNA isolation and quantitative real-time PCR analyses
Total RNA was isolated from piglet tissues using Trizol (15596026; Invitrogen) according to Jobgen et al. 30 The quantity and quality of the total RNA were determined by spectrophotometry (wavelength 230 nm). The expression of mRNAs for OH-POX (PRODH2), 16 HOA (HOGA), and AGT (AGXT2) in different tissues was determined by quantitative real-time polymerase chain reaction. Primers were designed using Primer Express Software Version 1.5 (Applied Biosystems). Primer pairs for selected genes are summarized in Supplemental Table 2. The quantitative polymerase chain reaction (PCR) was performed using the ABI prism 7900HT system (Applied Biosystems) with the Power SYBR Green PCR Master Mix (4309155; Applied Biosystems) as instructed by the manufacturer. Swine 18 S ribosomal RNA (RN18S, GenBank No. NR_046261) was used as the reference gene (Supplemental Table 2). The relative expression of mRNAs was calculated using the comparative ∆∆Cq method. 31
Immunohistochemical analyses
Immunohistochemical localization of OH-POX and POX proteins in tissue sections (∼5 μm) of piglets was performed to investigate their cell-specific and temporal expression, as we described previously for other proteins in animal tissues. 32 The immunohistochmical analyses were conducted using human anti-OH-POX immunoglobulin G (IgG) (0.5 mg protein/mL; developed and kindly provided by Dr. James M. Phang, National Cancer Institute, Frederick, MD) and rabbit anti-POX polyclonal IgG (HPA020361; Sigma-Aldrich, St Louis, MO) at dilutions of 1:500 and 1:200, respectively. Antigen retrieval was performed by boiling the samples in 10 mM sodium citrate buffer (pH 6.0). Purified non-relevant rabbit IgG was used as a negative control to replace the primary antibody at the same final concentration. Immunoreactive proteins were visualized in sections using the VECTASTAIN ABC Kit (PK-6101 for rabbit IgG; Vector Laboratories) and 3,3′-diaminobenzidine tetrahydrochloride (D5637; Sigma-Aldrich) as the color substrate.
Statistical analysis
Results are expressed as means ± SEM. Statistical analyses of data were performed by one-way analysis of variance using the General Linear Models procedures. 33 Differences among treatment means were determined using the Student-Newman-Keuls multiple comparison method. 33 A probability value of ⩽0.05 indicated statistical significance.
Results
Activities of glycine-synthetic enzymes in tissues of postnatal piglets per gram tissue
The activities of key enzymes (OH-POX, HOA, and AGT) in tissues from piglets, expressed per gram tissue, are summarized in Table 1. The activities of OH-POX in the liver and kidney decreased (P < 0.05) gradually during the first 3 weeks of postnatal life. The enzymatic activity of OH-POX in kidneys at day 21 of age was less (P < 0.05) than at day 14 of age, but the hepatic activity of OH-POX did not differ (P > 0.05) between days 14 and 21 of age. OH-POX activity was not detected in the stomach of 0- to 21-day-old piglets, the heart of 0- to 14-day-old piglets, the skeletal muscle of 0- to 7-day-old piglets, and the small intestine of newborn (0-day-old) piglets. OH-POX activity was present in the pancreas of 0- to 21-day-old piglets and increased (P < 0.05) gradually with age. In the small intestine, OH-POX activity was greater (P < 0.05) in 21-day-old piglets than that in 7- and 14-day-old piglets.
Activities of glycine-synthetic enzymes and related enzymes in tissues of neonatal pigs expressed per gram tissue.
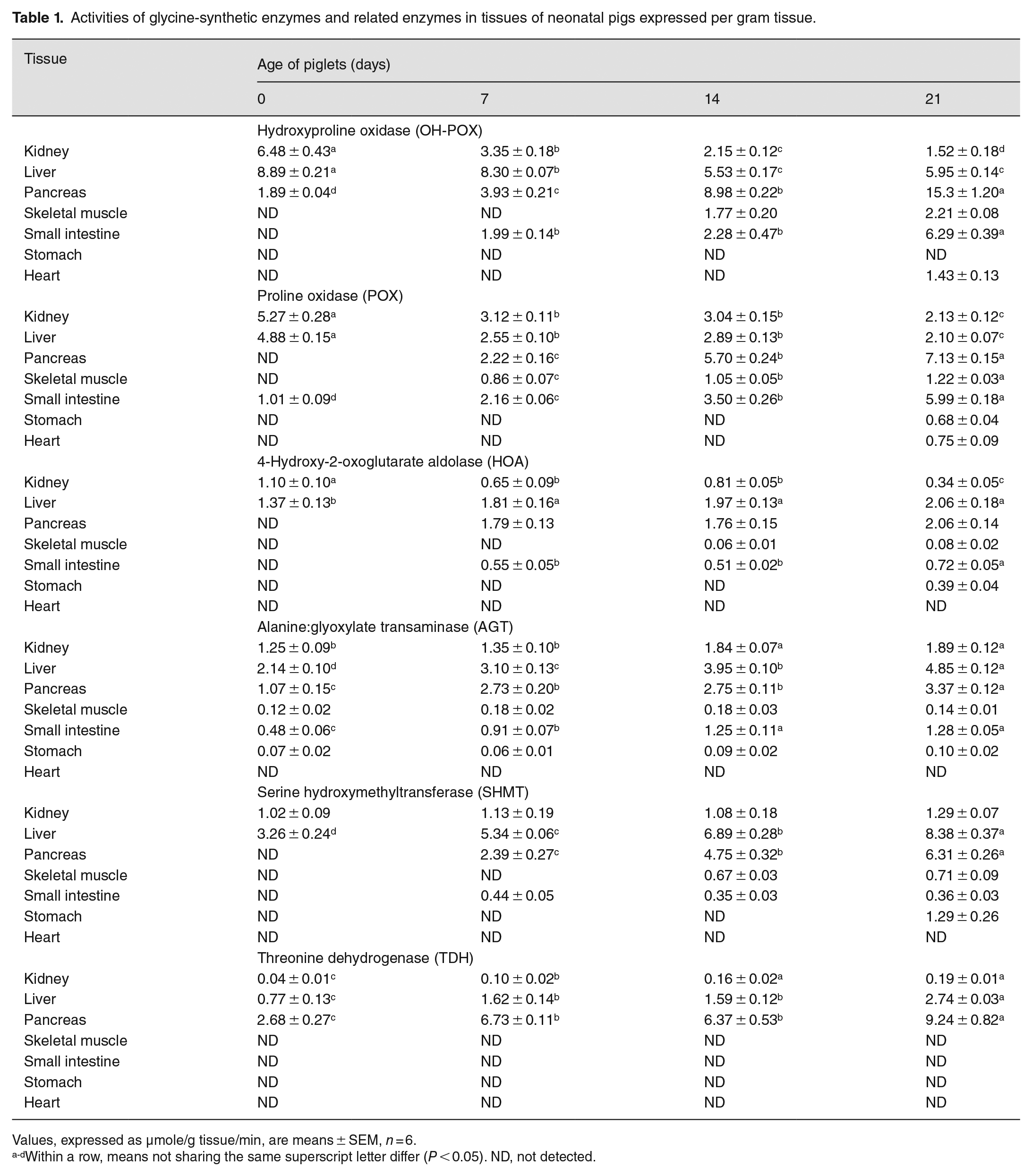
Values, expressed as µmole/g tissue/min, are means ± SEM, n = 6.
Within a row, means not sharing the same superscript letter differ (P < 0.05). ND, not detected.
Like OH-POX, HOA and AGT activities were present in the liver and kidneys of 0- to 21-day-old piglets (Table 1). In contrast to OH-POX, HOA and AGT activities in these two tissues increased (P < 0.05) gradually with increasing the age of piglets. The patterns of HOA and AGT activities in other tissues of piglets also differed from those of OH-POX. Specifically, HOA and AGT activities were absent from the heart of 0- to 21-day-old piglets. HOA activity was also absent from the skeletal muscle of 0- to 7-day-old piglets, the pancreas and small intestine of newborn (day 0) piglets, the stomach of 0- to 14-day-old piglets, and the heart of 0- to 21-day-old piglets. AGT activity in the skeletal muscle and stomach of piglets did not change (P > 0.05) between 0- and 21-day-old piglets, but AGT activities in both the pancreas and the small intestine increased (P < 0.05) between 0 and 21 days of age.
The patterns of postnatal changes of POX enzymatic activities in tissues of piglets were generally similar to those for OH-POX except that POX enzymatic activity occurred in the small intestine of newborn pigs and the stomach of 21-day-old piglets but was absent from the pancreas (Table 1). In the presence of 15 mM L-proline, the enzymatic activities of POX in the mitochondria of the jejunal mucosa, kidney, and liver were 12.6, 3.0, 1.9 nmol P5C/mg protein/min, respectively. By contrast, in the presence of 15 mM 4-hydroxyproline, production of ∆1-pyrroline-3-hydroxy-5-carboxylate (a product of potential oxidation of 4-hydroxyproline) was not detected, as measured with O-aminobenzaldehyde. The limit of detection for ∆1-pyrroline-3-hydroxy-5-carboxylate was 0.03 nmol/mg protein/min.
SHMT and TDH enzymatic activities were restricted mainly to the kidney and liver of 0- to 21-day-old piglets and to the pancreas of 7- to 21-day-old piglets. In the kidneys, SHMT enzymatic activity did not change between days 0 and 21 of age, but TDH activity increased (P < 0.05) during this period of postnatal life. In the liver and pancreas, both SHMT and TDH enzymatic activities increased (P < 0.05) between days 0 and 21 of age.
Total activities of glycine-synthetic enzymes per whole tissue in postnatal piglets
We estimated the total activity of each glycine-synthetic enzyme per whole tissue in piglets as enzyme activity per gram tissue (Table 1) × total weight of tissue (Supplemental Table 3). The results are summarized in Table 2. In 0- to 7- and 14- to 21-day-old pigs, the liver and skeletal muscle contained the most total activity of OH-POX, respectively. The total activity of OH-POX in the small intestine increased gradually (P < 0.05) between 7 and 21 days of age and in 21-day-old pigs, the total activity of OH-POX was similar between the gut and the liver. The liver also had the most activity of POX in newborn pigs, but skeletal muscle had the most total activity of POX in 7- to 21-day-old pigs, followed by the small intestine in both 14- and 21-day-old pigs. The total activity of HOA increased gradually (P < 0.05) in the liver between 0 and 21 days of age, and in the pancreas, skeletal muscle, and small intestine between 7 and 21 days of age. Thus, in 14- and 21-day-old pigs, skeletal muscle and the small intestine had the second and third most total activity of HOA, respectively. Similar results were obtained for the tissue distribution of total activity of AGT. Interestingly, in 0- to 7- and 14- to 21-day-old pigs, the liver and skeletal muscle contained the most total activity of SHMT, respectively. In all ages of piglets studied, the liver had the most TDH activity.
Activities of glycine-synthetic enzymes and related enzymes per whole tissue in neonatal pigs.
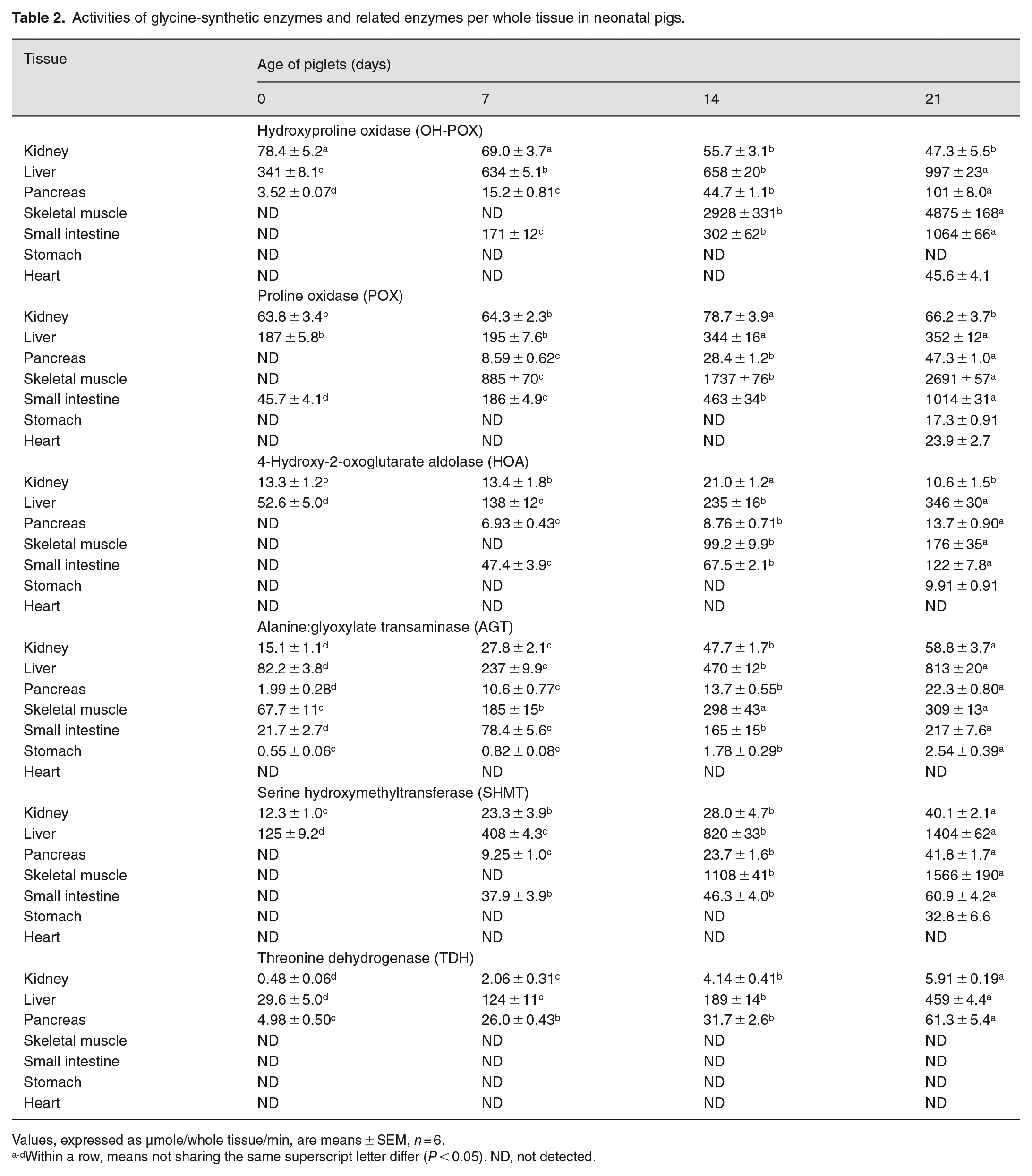
Values, expressed as µmole/whole tissue/min, are means ± SEM, n = 6.
Within a row, means not sharing the same superscript letter differ (P < 0.05). ND, not detected.
Expression of mRNA for enzymes in tissues of postnatal piglets
The abundance of the OH-POX (PRODH2) mRNA exhibited a similar pattern of changes in the liver and kidneys to their enzymatic activities; namely, expression levels were high at birth and decreased (P < 0.05) with age (Table 3). Of particular note, the expression of OH-POX mRNA in the liver and kidneys of newborn piglets was 2.7 and 4.6 times greater (P < 0.05), respectively, than that for 21 day-old piglets, whereas the expression of OH-POX mRNA in the pancreas of 21 day-old piglets was 5.8 times greater than that for newborn piglets. With increasing age from day 0 to day 21, the expression of mRNAs for HOGA and AGXT2 increased (P < 0.05) in the pancreas, small intestine, and stomach, but the expression of HOGA decreased (P < 0.05) in the liver. The expression of AGXT2 mRNA in the liver was not affected by the age of piglets (P > 0.05).
Expression of mRNAs for glycine-synthetic enzymes and related enzymes in tissues of neonatal pigs.
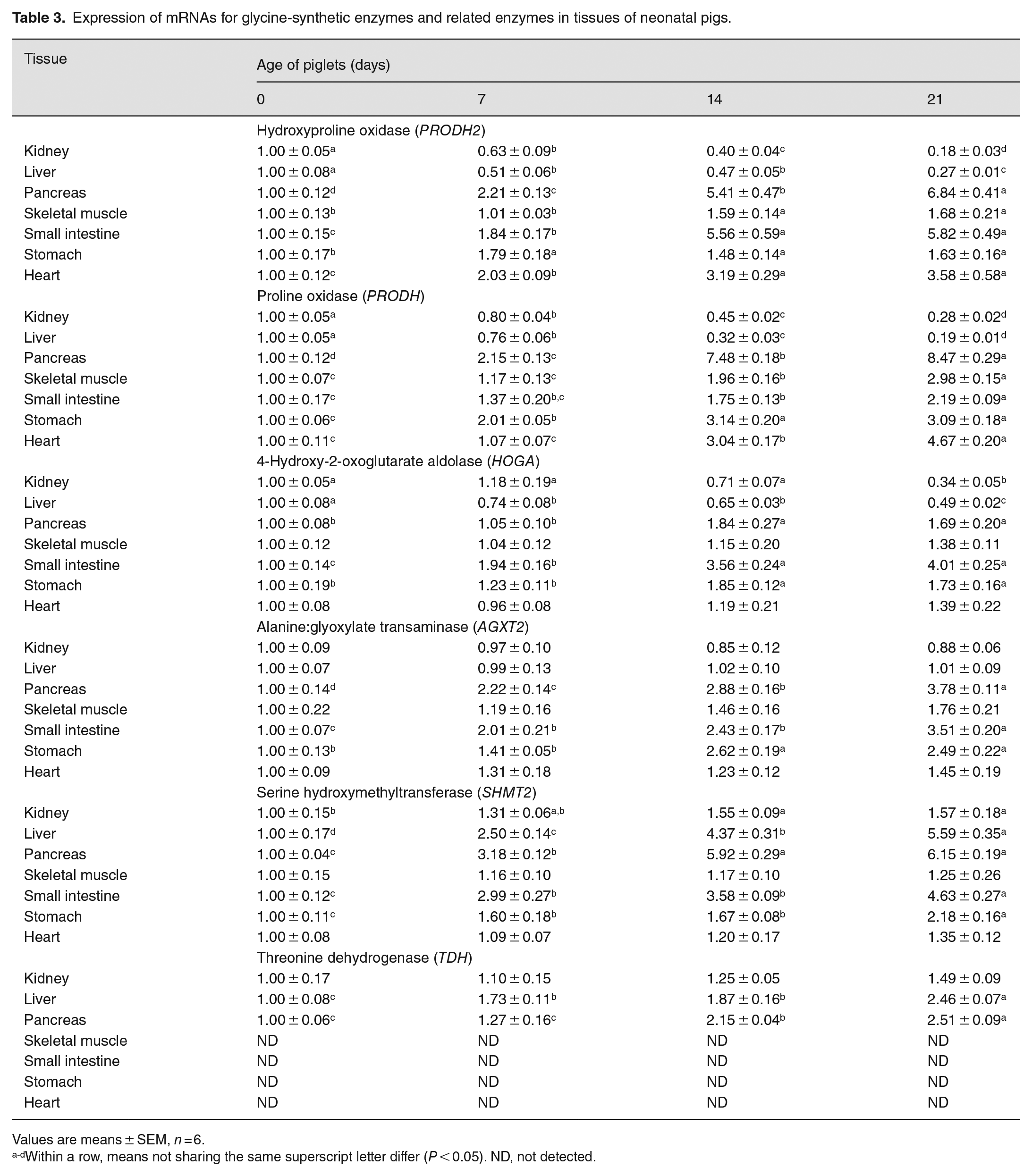
Values are means ± SEM, n = 6.
Within a row, means not sharing the same superscript letter differ (P < 0.05). ND, not detected.
PRODH mRNA was expressed in all tissues of 0- to 21-day-old piglets. Patterns of postnatal changes in the expression of PRODH mRNA (Table 3) were generally similar to those for changes in the enzymatic activities of POX (Table 1), except that the expression of PRODH mRNA increased (P < 0.05) gradually in the skeletal muscle, stomach, and heart with increasing the age of piglets between days 0 and 21. Except for skeletal and cardiac muscles, the expression of SHMT2 mRNA in other tissues increased (P < 0.05) gradually between days 0 and 21 of age. TDH mRNA was not detected in the skeletal muscle, small intestine, stomach, and heart of 0- to 21-day-old piglets. As the age of piglets increased from 0 to 21 days, the expression of TDH mRNA increased (P < 0.05) gradually in the liver and pancreas but did not change (P > 0.05) in the kidneys (Table 3).
Localization of OH-POX and POX proteins in pig tissues
In the kidneys, OH-POX protein was localized mainly to the convoluted tubules, primarily the proximal tubules, whereas immunostaining for OH-POX was low to undetectable in the renal corpuscles (Figure 2). Renal OH-POX protein was highly expressed at birth and decreased to day 21 of age, as was observed for its enzymatic activity and mRNA abundance. POX protein was localized primarily to the renal corpuscles and a subset of tubules (medullary rays) within the cortex, and expression appeared to decrease between days 0 and 7 of age.
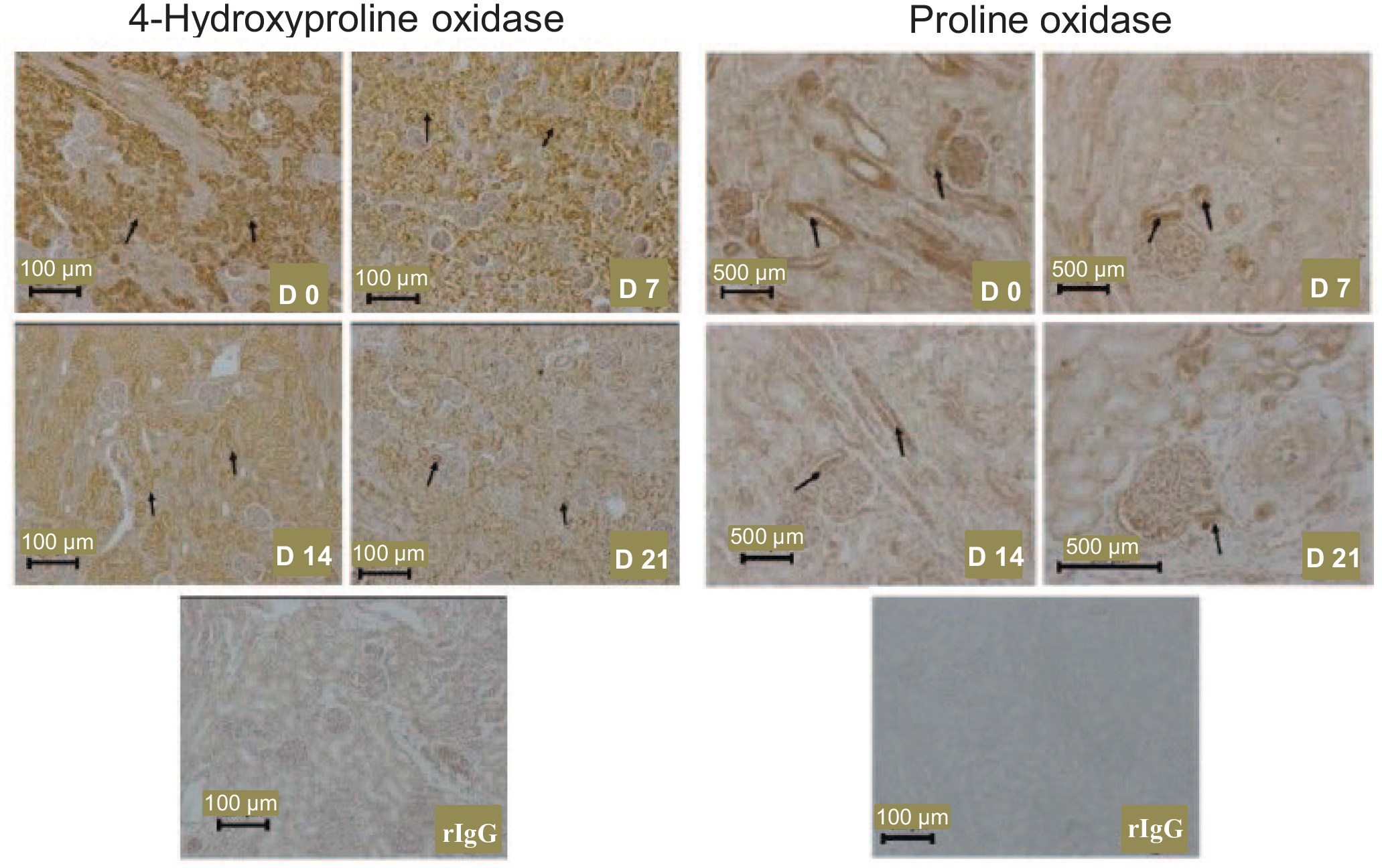
The localization of 4-hydroxyproline oxidase and proline oxidase proteins in the kidneys of 0-, 7-, 14-, and 21-day-old piglets. Arrows indicate expression of 4-hydroxyproline oxidase in convoluted tubules and proline oxidase is a subset of kidney tubules. D, day; rIgG, rabbit immunoglobulin G, served as the negative control.
In the liver of piglets, there were cell-specific changes in the expression of immnoreactive OH-POX and POX proteins with age (Figure 3). Specifically, OH-POX protein was present mainly in the periportal part of the liver lobule at day 0 of age. The expression of OH-POX protein shifted to the transitional part of the liver lobule by days 7 and 14 of age, and this protein was localized to cells in the perivenous zone around the central vein in the liver at day 21 of age. In contrast, POX protein was localized mainly to the periportal hepatocytes in the liver lobules of 0- to 21-day-old piglets.
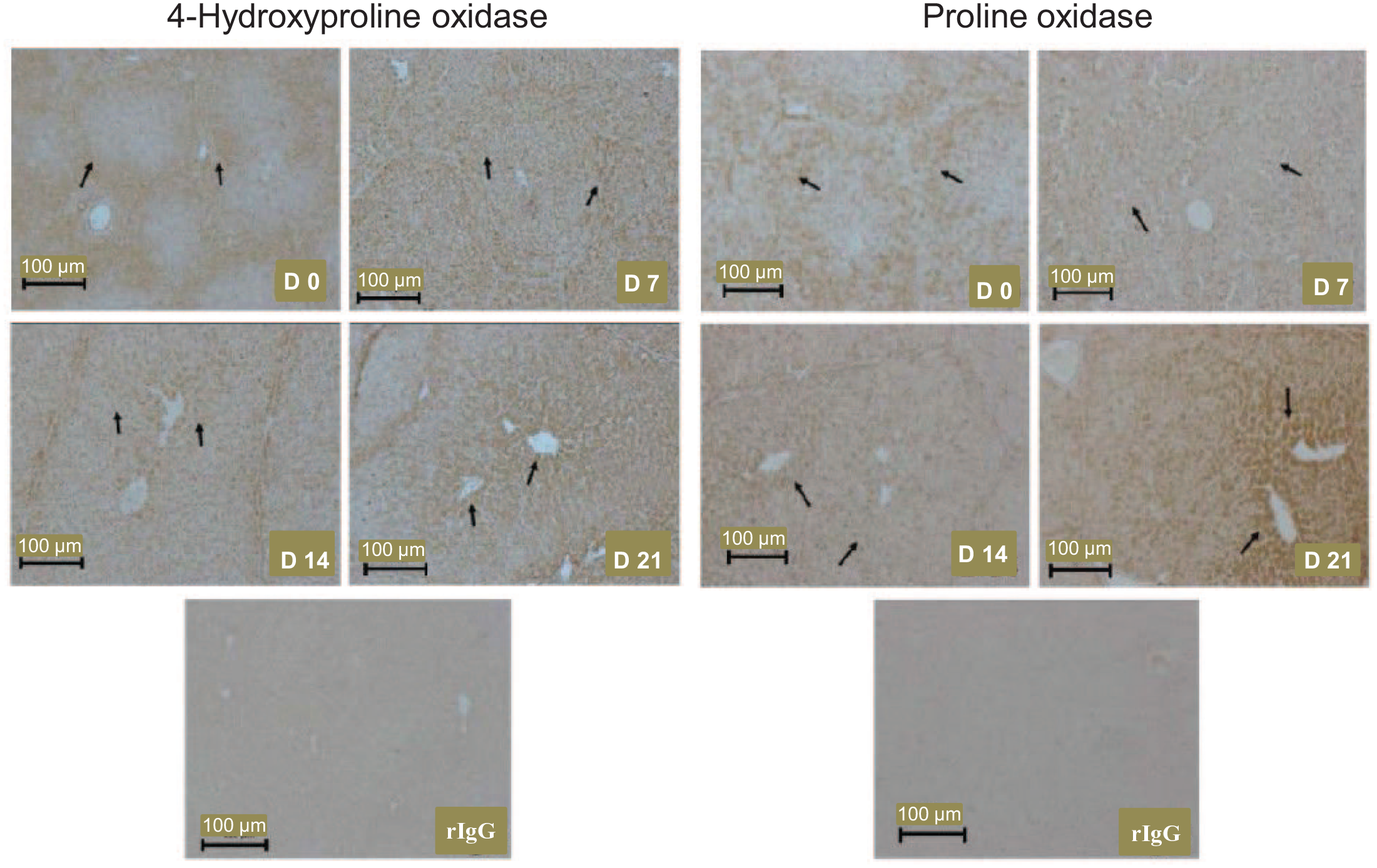
The localization of 4-hydroxyproline oxidase and proline oxidase proteins in the liver of 0-, 7-, 14-, and 21-day-old piglets. Arrows indicate the progression of 4-hydroxyproline oxidase expression from the periportal region to the central venous drainage region of the hepatic lobule between days 0 and 21 of age, and increasing expression of proline oxidase. D, day; rIgG, rabbit immunoglobulin G, served as the negative control.
In the pancreas, OH-POX was localized to the epithelial cells of the acini of the exocrine component of the gland and was not detected in the islets of the endocrine portion of the gland (Figure 4). The abundance of OH-POX protein appeared to increase after 7 days of age. In contrast, POX protein was localized to both the acini and ducts of the exocrine pancreas of 7- to 21-day-old piglets (Figure 4), but not in the pancreas of newborn piglets (data not shown).
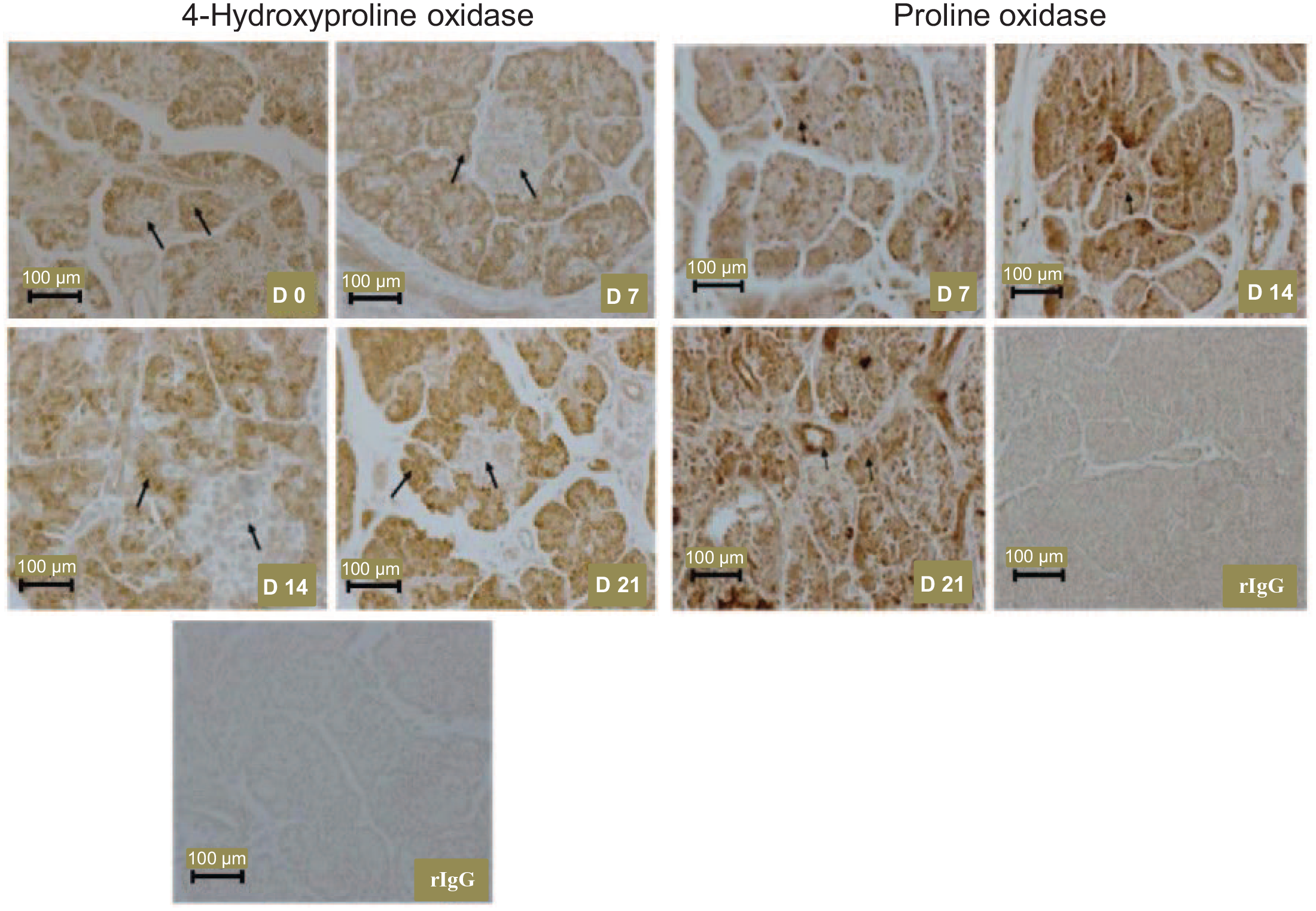
The localization of the 4-hydroxyproline oxidase protein in the pancreas of 0-, 7-, 14-, and 21-day-old piglets and the localization of the proline oxidase protein in the pancreas of 7-, 14-, and 21-day-old piglets. Proline oxidase protein was not detected in the pancreas of newborn pigs. Arrows indicate expression of 4-hydroxyproline oxidase in the acini of the exocrine pancreas but not in the islets of the endocrine pancreas, and indicate expression of proline oxidase in the acini and ducts of the exocrine pancreas. D, day; rIgG, rabbit immunoglobulin G, served as the negative control.
In the small intestine (jejunum), OH-POX protein was localized primarily to enterocytes, the epithelia of intestinal glands (crypts), and the smooth muscle of the muscularis mucosa, whereas POX protein was highly abundant in enterocytes (Figure 5). Both OH-POX and POX were expressed in the connective tissue of the jejunum. In contrast, the OH-POX protein was absent from the stomach of 0- to 21-day-old piglets (data not shown) and was present only in the stomach of 21-day-old piglets where OH-POX was expressed in all cell types but was most prominent in the glands (Supplemental Figure 1).
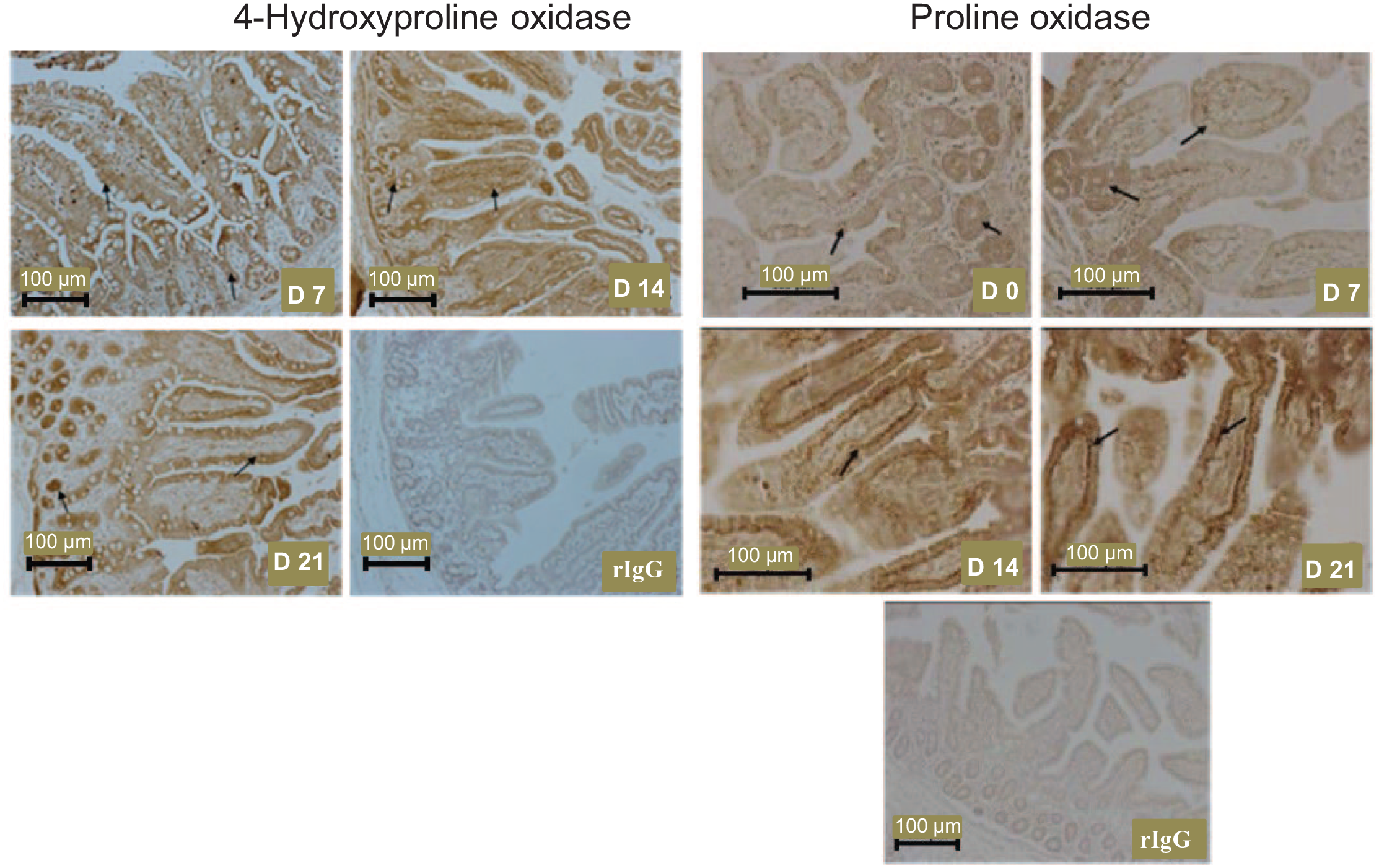
The localization of the 4-hydroxyproline oxidase protein in the small intestine (jejunum) of 7-, 14-, and 21-day-old piglets and the localization of the proline oxidase protein in the small intestine of 0-, 7-, 14-, and 21-day-old piglets. The 4-hydroxyproline oxidase protein was not detected in the small intestine of newborn pigs. Arrows indicate the expression of 4-hydroxyproline oxidase in enterocytes and crypts, and proline oxidase in enterocytes. D, day; rIgG, rabbit immunoglobulin G, served as the negative control.
OH-POX and POX proteins were expressed in cells of the bile ducts of the liver. These two proteins were expressed highly in epithelial cells of the gallbladder and, to a lesser extent, in the connective tissue of neonatal piglets (Figure 6). OH-POX and POX proteins were absent from the heart and skeletal muscle of 0- to 14-day-old piglets. However, both proteins were detected in the striated muscle fibers and the connective tissue fibroblasts of the heart and skeletal muscle of 21-day-old piglets (Figure 7). In the skeletal muscle of 21-day-old piglets, POX protein also appeared to be present in the endothelium (Figure 7).
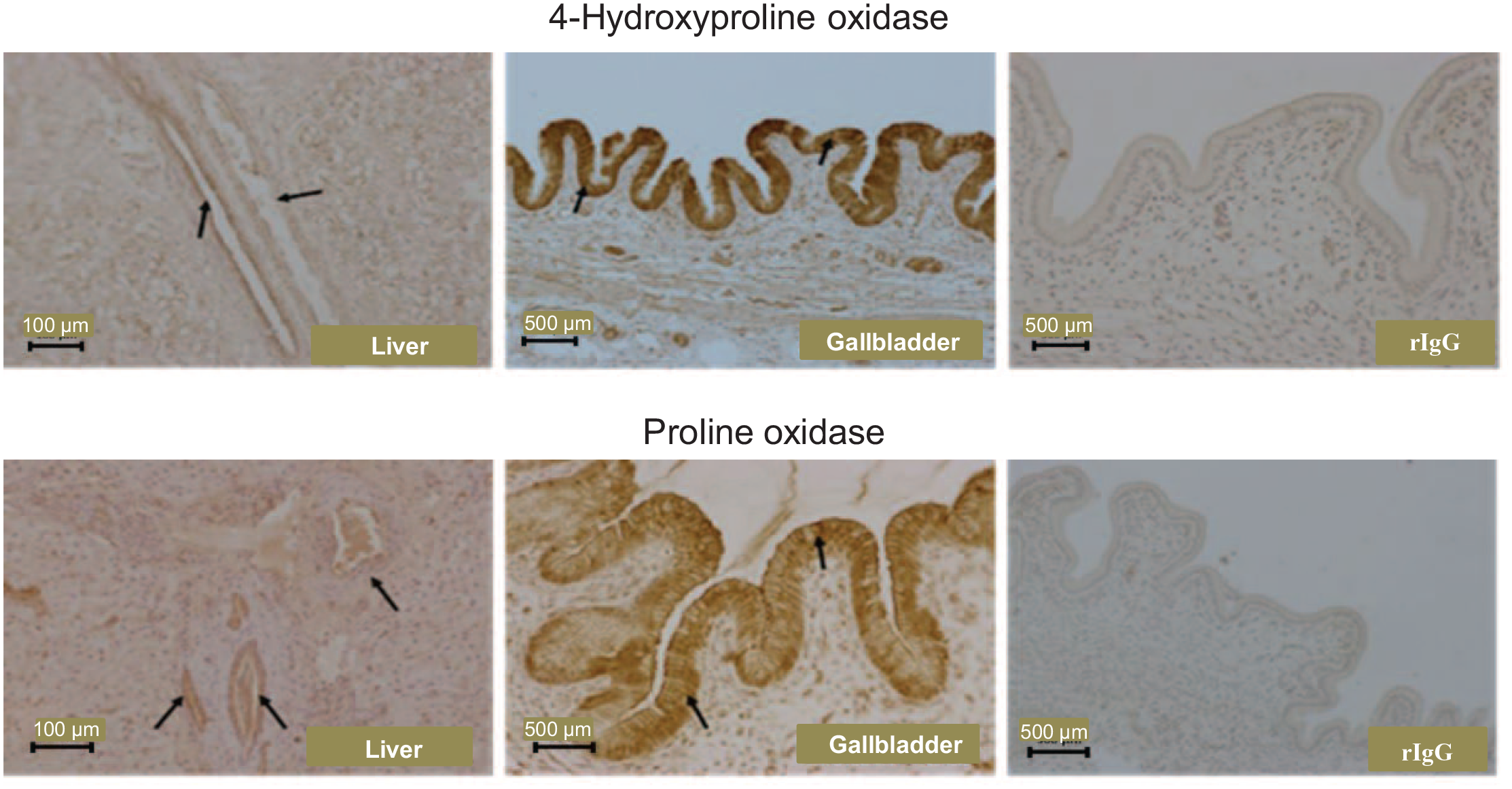
The localization of 4-hydroxyproline oxidase and proline oxidase proteins in the bile duct of the liver and the gallbladder of 21-day-old piglets. rIgG, rabbit immunoglobulin G, served as the negative control.
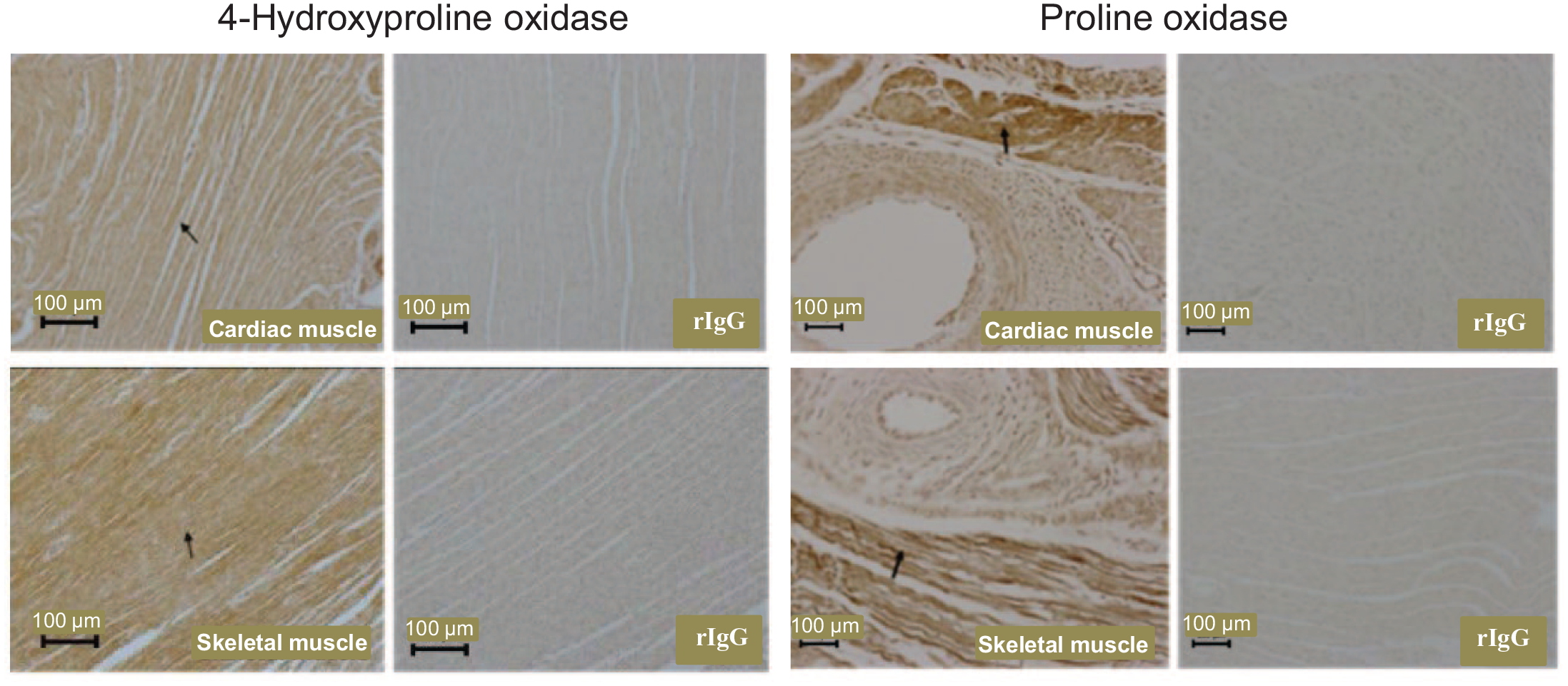
The localization of 4-hydroxyproline oxidase and proline oxidase proteins in the heart and skeletal muscle of 21-day-old piglets. Neither of these proteins were detected in the heart and skeletal muscle of 0- to 14-day-old piglets. Arrows indicate expression of protein in the striated muscle fibers of cardiac and skeletal muscles. rIgG, rabbit immunoglobulin G, served as the negative control.
Glycine synthesis from tissues of postnatal pigs
Except for the liver, kidney, and skeletal muscle of 0- to 7-day-old piglets, concentrations of glycine in the extracts of tissues after a 2 h incubation were greater (P < 0.05) than those for tissues without incubation due to net protein degradation (Table 4). As the addition of 4-hydroxyproline to the incubation medium increased from 0 to 5 mM, concentrations of glycine in the extracts of the liver and kidneys of 0- to 21-day-old piglets after a 2 h incubation increased (P < 0.05) in a dose-dependent manner (Table 4). Except for newborn piglets, concentrations of glycine in the extracts of the small intestine of 7 to 21-day-old piglets after 2 h incubation increased (P < 0.05) with increasing the concentrations of 4-hydroxyproline in the incubation medium from 0 to 5 mM. Similar results were obtained for skeletal muscle from 14- and 21-day-old piglets.
Concentrations of glycine in incubation medium plus tissue from neonatal pigs.
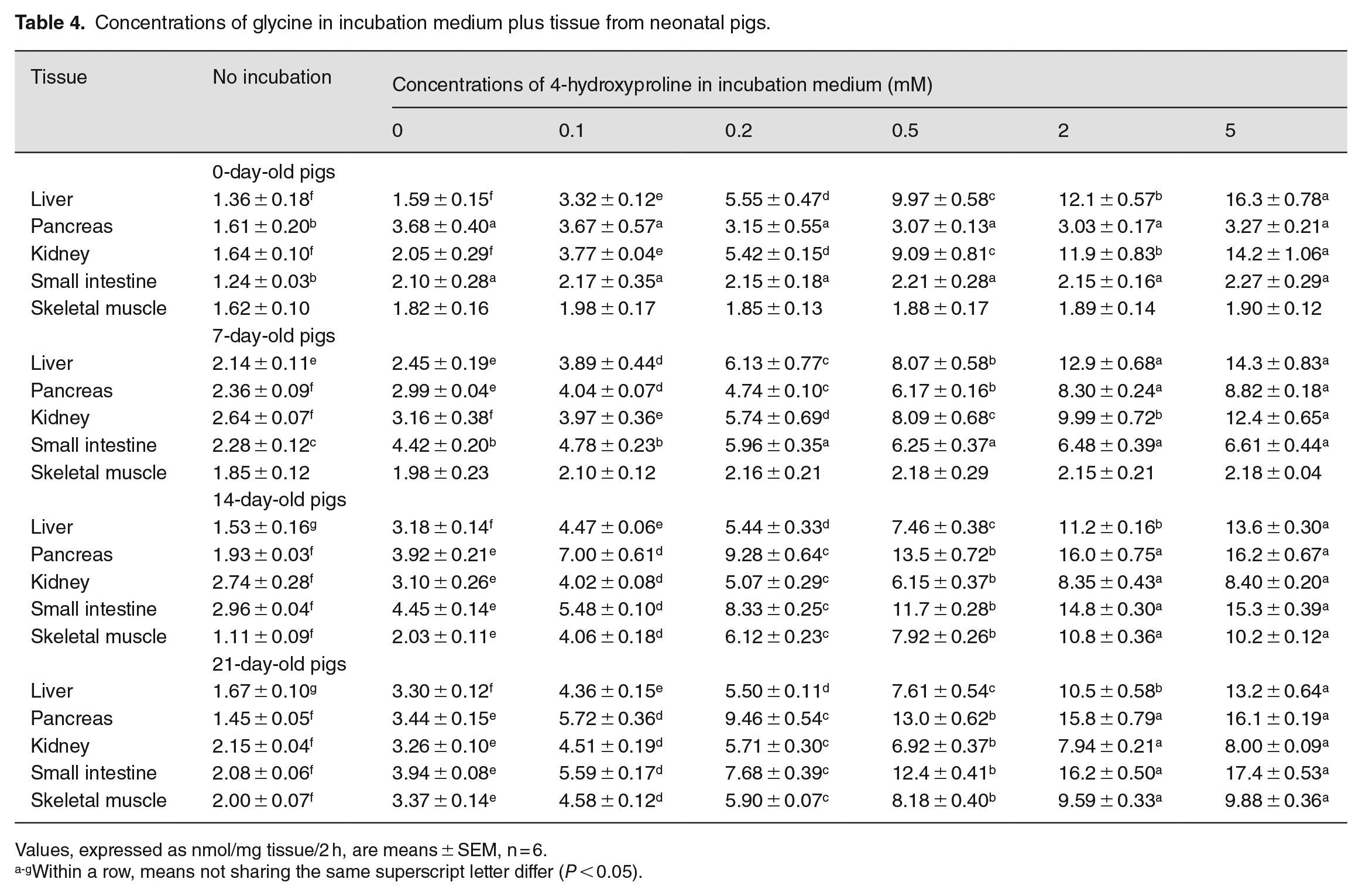
Values, expressed as nmol/mg tissue/2 h, are means ± SEM, n = 6.
Within a row, means not sharing the same superscript letter differ (P < 0.05).
Discussion
Glycine, the most abundant amino acid in the animal body, 34 has now been recognized to be nutritionally essential for the maximal growth and optimal development of young pigs.1,2,35 Paradoxically, sow-reared piglets continue to grow despite insufficient amounts of glycine in milk, 36 indicating a role for endogenous synthesis in glycine provision. We have estimated that threonine, serine, and choline (traditionally classified substrates) in milk contribute to 12% of endogenous glycine synthesis in sow-reared piglets. 1 Major precursor(s) of glycine in the neonates remain unknown. The main finding of this study is that primary tissues of piglets actively convert 4-hydroxyproline to glycine. The results of this work significantly advance our knowledge of amino acid biochemistry and nutrition in animals.
Glycine, proline, and 4-hydroxyproline are the major constituents of the extracellular matrix proteins, collagen and elastin.37,38 The degradation of those proteins releases a glycine-proline-hydroxyproline tripeptide into physiological fluids, such as milk and the blood of neonates. 13 Thus, this tripeptide is abundant in sow’s milk (up to 10 mM) and the plasma of piglets (up to 3 mM). 13 For comparison, sow’s milk and the plasma of piglets contain 4 mM glutamine 39 and 1 mM glycine, 36 respectively. The unusual abundance of the Gly-Pro-4-hydroxyproline tripeptide in milk and blood raises an important question about its nutritional and physiological significance. 2 The major finding of this study is that these three amino acids are closely related not only as components of collagen but also in metabolism. Namely, proline is incorporated into collagen, where some proline residues are hydroxylated into 4-hydroxyprolines. With the degradation of collagen, there is the release of 4-hydroxyproline, which is then used by multiple tissues (e.g. the liver, kidney, skeletal muscle, pancreas, and small intestine) for the synthesis of glycine. This proline-hydroxyproline-glycine pathway provides endogenous glycine to maintain the whole-body homeostasis, growth, and development of piglets.
The conversion of 4-hydroxyproline into glycine requires several enzymes, including OH-POX, HOA, and AGT (Figure 1). Interestingly, the distribution of these enzymes is highly tissue- and age-specific (Table 1). OH-POX plays a key role in degrading 4-hydroxyproline in animals. 40 Interestingly, this study revealed that OH-POX protein was highly expressed in the kidneys (Figure 2) and liver (Figure 3) of 0- to 21-day-old piglets but was absent from the small intestine of newborn piglets (Figure 5). Furthermore, OH-POX activity in the pancreas increased gradually after birth and reached its greatest level at 21 days of age among all the tissues examined. This pattern of the OH-POX activity differed from that of POX in the tissues of piglets, as POX activity was greatest in the small intestine but was absent from the pancreas and skeletal muscle at birth (Table 1) and during the suckling period.41,42 Although 4-hydroxyproline and proline are structurally similar, they are metabolized in different tissues via distinctly different enzymes. 43 We found that POX in pig tissues (e.g. the small intestine, kidney, and liver) did not use 4-hydroxyproline as a substrate and, therefore, would not contribute to the 4-hydroxyproline-glycine pathway. In this work, we did not determine mRNA levels or enzymatic activities of 3-hydroxy-pyrroline-5-carboxylate dehydrogenase. The latter is identical to pyrroline-5-carboxylate dehydrogenase in mammalian tissues. Future studies are warranted to determine whether this enzyme may potentially limit glycine production from 4-hydroxyproline in pig tissues.
Consistent with our results regarding OH-POX in the tissues of piglets, Bengtsson and Hakkarainen 44 reported that [ 14 C]4-hydroxyproline administered into newborn piglets were rapidly detected in their liver and kidneys to a much greater extent than in other organs. Thus, the liver and kidneys actively take up 4-hydroxyproline in vivo. 4-Hydroxyproline is then metabolized by tissues to form glycine during the suckling perid (Table 4). In addition, the skeletal muscle and the small intestine of 14- to 21-day-old piglets, as well as the pancreas of 7- to 21-day-old piglets also synthesized glycine from 4-hydroxyproline. Based on the tissue distrubition of glycine-synthetic-enzyme activities (Table 2) and metabolic rates (Table 4), the liver and kidney were the primary tissues for converting 4-hydroxyproline into glycine in 0- to 7-day-old piglets. However, given the large amount of skeletal muscle in young animals (40% of the body weight), 45 it is the main site for the 4-hydroxyproline-glycine pathway in 14- to 21-day-old piglets (Table 2), further supporting the notion that skeletal muscle plays a vital role in amino acid metabolism and nutriiton. This helps to explain high concentrations of glycine in the plasma of piglets. 36 Sufficient concentrations of glycine are needed to activate the mechanistic target of rapamycin cell signaling for the synthesis of proteins and nucleotides (purines) in skeletal muscle.1,46 We estimated that the piglet must be able to synthesize 1.05 g glycine/kg body weight/day from precursors other than serine, threonine, and choline to meet its demands for growth and development. 1 It appears that this amount of glycine is generated mainly from 4-hydroxyproline present in milk and derived from the catabolism of collagen in vivo.2,13
The synthesis of glycine from 4-hydroxyproline is an adenosine triphosphate (ATP)-producing process. 45 Particularly, 1 mole of FADH2 and NADH and 2 moles of pyruvate are generated from 1 mole of 4-hydroxyproline during this process, which suggests that the conversion of 4-hydroxyproline to glycine is preferred by animals for glycine synthesis due to the formation of both ATP and glycine. Thus, the 4-hydroxyproline-Gly pathway is energetically efficient for glycine production in the body. The re-use of 4-hydroxyproline released from collagen degradation for the production of glycine helps to maximize the efficiency of utilization of dietary and endogenously synthesized proline and minimize the urinary excretion of 4-hydroxyproline from animals. 13 This is of nutritional and physiological importance for young piglets that have high demands for both ATP and glycine. 2
The liver of piglets exhibits cell-specific expression of OH-POX (Figure 3). There are two different types of hepatocytes (periportal and perivenous) and three distinct zones in the liver. 47 The periportal and perivenous hepatocytes have very different metabolic activities.48,49 The hepatic zonation includes the periportal (zone I), transitional (zone II), and perivenous (zone III) zones. A change in the localization of the OH-POX protein within those zones (Figure 3) may indicate a shift in its function between birth and weaning. The periportal hepatocyte contains abundant mitochondria and participates in amino acid synthesis and catabolism.47–49 However, the perivenous hepatocytes contain fewer mitochondria and are involved in fatty acid synthesis.49,50 The high abundance of the OH-POX protein in the periportal hepatocytes at birth suggest that those cells have a greater capacity to catabolize 4-hydroxyproline to generate glycine than perivenous hepatocytes. As neonatal piglets grow, the small intestine and skeletal muscle express OH-POX protein and become metabolically active tissues to convert milk- and blood-borne 4-hydroxyproline into glycine. Thus, lesser amounts of 4-hydroxyproline would enter the liver via the portal vein in 14- to 21-day-old piglets than in 0- to 7-day-old piglets. The 4-hydroxyproline that enters the liver would largely bypass the periportal hepatocytes and be converted to glycine in the perivenous hepatocytes where both glycine oxidation and urea cycle activity are limited. 51 Thus, this cell-specific expression of enzymes for the 4-hydroxyproline-dependent glycine-metabolic pathway is important for physiologically maximizing the synthesis of glycine by the liver.
The cell-specific expression of OH-POX protein in extrahepatic tissues is also physiologically relevant. Any dietary 4-hydroxyproline that escapes the liver and 4-hydroxyproline produced from the degradation of collagen and elastin in the connective tissue are taken up by the kidneys, skeletal muscle and possibly other tissues for glycine synthesis. 52 In the kidneys, the proximal tubules of the cortex are the main site of OH-POX expression, 4-hydroxyproline degradation, and glycine synthesis (Figure 2). Of interest, the proximal tubules of the kidney express argininosuccinate synthase and lyase for synthesis of arginine from citrulline, as well as arginine: glycine amidinotransferase that converts arginine and glycine into guanidinoacetate, the immediate precursor of creatine. 53 The colocalization of the enzymes for guanidinoacetate production in the same renal site can increase the efficiency of creatine synthesis, which is crucial for the growth of milk-fed piglets. 54 Likewise, the presence of glycine synthesis in skeletal muscle, which lacks the glycine cleavage system for glycine degradation, one can maximize the production of glycine locally to improve the health and growth of piglets.
Conclusion
Results of this study indicate that 4-hydroxyproline is a major precursor for glycine synthesis in sow-reared pigs. OH-POX, the key enzyme for the degradation of 4-hydroxyproline to generate glycine, is highly expressed in the liver and kidneys of neonatal pigs. The pancreas, small intestine, and skeletal muscle of 14- to 21-day-old piglets can also actively synthesize glycine from 4-hydroxyproline. Based on the tissue distribution of glycine-synthetic enzymes, metabolic rate, and tissue mass of piglets, the 4-hydroxyproline-glycine pathway is quantitatively most important in the liver and kidneys during the first week of life and in skeletal muscle thereafter. The cell-specific and temporal changes in the expression of glycine-synthetic enzymes maximize the net production of glycine to compensate for a severe deficiency of glycine in sow’s milk, thereby supporting the growth and development of piglets.
Supplemental Material
sj-pdf-1-ebm-10.1177_15353702231181360 – Supplemental material for Synthesis of glycine from 4-hydroxyproline in tissues of neonatal pigs
Supplemental material, sj-pdf-1-ebm-10.1177_15353702231181360 for Synthesis of glycine from 4-hydroxyproline in tissues of neonatal pigs by Shengdi Hu, Wenliang He, Fuller W Bazer, Gregory A Johnson and Guoyao Wu in Experimental Biology and Medicine
Footnotes
Acknowledgements
The authors thank Dr. James M. Phang, National Cancer Institute (Frederick, MD 21702, USA) for kindly providing hydroxyproline oxidase antibody; Drs. Gayan Nawaratna, Xiaoqiu Wang, and Barry D. Long for technical assistance; and Drs. John T. Brosnan and Margaret E. Brosnan for helpful discussion.
Authors’ Contributions
GW designed and supervised the study. SH, WH, and GW performed the experiment. SH and GW statistically analyzed experimental data, summarized results, and wrote the manuscript. FBW and GAJ contributed to data interpretation and manuscript revisions. All authors read and approved the final manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by Agriculture and Food Research Initiative Competitive Grant (2014-67015-21770) from the USDA National Institute of Food and Agriculture.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.