
Abstract
Recent advances in the three-dimensional (3D) cancer models give rise to a plethora of new possibilities in the development of anti-cancer drug therapies and bring us closer to personalized medicine. Three-dimensional models are undoubtedly more authentic than traditional two-dimensional (2D) cell cultures. Nowadays, they are becoming preferentially used in most cancer research fields due to their more accurate biomimetic characteristics. On the contrary, they still lack the cellular and matrix complexity of the native tumor microenvironment (TME). This review focuses on the description of existing 3D models, the incorporation of TME and fluidics into these models, and their perspective in the future research. It is clear that such an improvement would need not only biological but also technical progress. Therefore, the modern approach to anti-cancer drug discovery should involve various fields.
Impact Statement
This review provides insights into the field of three-dimensional (3D) in vitro models and their use in cancer research and therapies development. A special focus is put on the incorporation of the tumor microenvironment and non-cancerous cells in such models as a crucial part of tumorigenesis. Tumor microenvironment has been recognized as a hallmark of cancer. Therefore, ignoring its role in cancer development and progression in in vitro models means that they cannot provide a fully relevant platform for basic and translational cancer research, nor for anti-cancer drug development. The authors are discussing whether current 3D tissue models are capable of replicating all aspects of cancer biology and the directions which should be taken in developing them further into relevant in vitro platforms fully mimicking the tumor–stroma interactions as in native tumor tissue.
Introduction
Cancer is a major public health problem and one of the leading causes of death across the world with increasing incidence every year. 1 Despite decades of research on novel therapeutics, multiple tumors, such as glioblastoma, pancreatic cancer, and triple-negative breast cancer, still have limited clinical treatment options. Only about 10% of potential anti-cancer drugs succeed during their clinical testing. 2 Many of them are failing due to the disparities between results obtained in preclinical studies and outcomes in clinical settings. One of the main reasons behind this obstacle is the use of preclinical models that lack the ability to authentically represent all conditions and physiological processes in the human body.
There is a long journey before an anti-cancer drug progresses from laboratory to clinical testing stage. Usually, the initial in vitro studies are followed by the animal experiments—frequently on more than one animal species. However, there are many reasons that put a pressure on the need to use appropriate human in vitro models to their maximum potential. Those reasons include ethical problems of animal experimentation, legislative ones—such as 3Rs rules (Replacement, Reduction, and Refinement of the inclusion of animals in research) as well as cost. Yet another important set of factors that causes disconnection between preclinical and clinical stages are interspecies differences. Therefore, many of the promising therapies successfully tested on animals often fail when it comes to human treatment.
A very important step in improving and/or streamlining the preclinical stage is the newly published US Food and Drug Administration (FDA) decision from early 2023 that makes animal testing non-mandatory before human trials. 3 This law doesn’t ban the testing of new drugs on animals outright because of the many limitations of in vitro systems. Companies simply cannot switch from animals to in vitro systems that might not be able to capture all of the types of toxicities, such as systemic effects. This decision simply creates possibilities to avoid animal testing in areas where suitable models and methods exist or will be created in the future. Therefore, this puts even more possibilities and pressure on the development of accurate in vitro models.
Unfortunately, much of the in vitro cancer research is still performed on two-dimensional (2D) cell cultures. While it is a straightforward approach with low-cost maintenance, these two factors may be also the only advantages of such approach nowadays. 4 Limitations of using 2D in vitro models have been described many times proving their inadequacy as fully reliable preclinical cancer models that fail to address many pathological aspects.2,5,6 For example, it has been reported that 2D cultures of healthy cells do not conserve the original shape and polarization of the cells, 7 which could affect many properties through non-physiological cell signaling, even though the loss of cancer cells’ polarity is often observed. 8 Cell cultures growing in monolayers on the plastic dishes also do not have proper spatial interactions with other cell types that are normally present in the native tissues. Lack of such intercellular communication, in turn, results in further skewing of not only signaling but also transport of nutrients and other molecules, including the therapeutic ones. There is no doubt about the contributions of 2D cultures in cancer biology, but to move forward more deeply into the problems that are not solved yet, they are simply not enough. On the contrary, the internal environment of animal models has a limited ability to mimic the complex process of pathophysiological conditions in humans. This may, according to Mak et al., 9 lead to a low rate of successful translation from animal models to clinical trials, which is at present not more than 8%. There is no doubt that animal models contributed greatly to many important discoveries, but still, most of the promising therapies successfully tested on animals fail when it comes to human treatment.
Since Mina Bissell with her colleagues pointed out the importance of the extracellular matrix (ECM) in the cell biology,10–13 three-dimensional (3D) cell culture models are generally accepted as more accurate cancer research models than the 2D models that have been in use by then, since they allow studying not only the tumor cells but also to include various components of their microenvironment. The tumor microenvironment (TME) has an inseparable role in processes such as tumor proliferation, dissemination, immune evasion, angiogenesis, epigenetics, and many others associated with cancer progression. The composition of the TME varies between tumor types, but hallmark features include the presence of non-malignant epithelial cells, immune cells, stromal cells, blood and lymphatic vessels, microbiome, and ECM.14,15 All of these types of cells interact with cancer cells, each other, and also with ECM by secreting different types of molecules, such as cytokines, chemokines, extracellular vesicles, and miRNA. All these direct cell-to-cell interactions together with indirect communication through secreted molecules can influence various signaling pathways that help cancer cells to proliferate, migrate, and spread throughout the organism. 16 Therefore, an appropriate cancer model should be able to recapitulate all the aspects of the TME and its interactions with tumor, in order to properly study crucial cancer-related processes.
In the future, bioengineered humanized 3D models of tumors could eventually replace the requirement for testing on animals. In fact, it would be a desirable outcome for all the aforementioned ethical and even more importantly biological reasons. The development of such models would provide great ethical and economic benefits for the prediction of tumor response to treatment (e.g. chemotherapy, radiotherapy), and reduce the number of animals sacrificed in preclinical studies. Below, we discuss the currently available 3D in vitro models including the tumor cells with their TME and bring insight into the future of drug testing in oncology.
Spheroids
Multicellular tumor spheroids are considered to be the first and also simplest 3D in vitro model frequently used in cancer research. 17 Spheroids are defined as cellular aggregates obtained in suspension or embedded within 3D matrix. 18 In general, spheroids can be described as rounded clusters of cancer cells (usually cancer cell lines) grown on low-attachment or agarose-coated culture plates. Spheroid media are usually non-defined and serum-containing media. Another approach is a “hanging drop” technique, where the cells are cultured in a drop of culture medium hanging on the lid of Petri dishes and incubated under physiological conditions until they create 3D structures. 19 The 3D formation allows cancer cells to be in close proximity to one another and form a “mass,” which provides great conditions for the accumulation of cell-generated collagen. 20 In majority of current models, spheroids are embedded in a gel 21 or encapsulated into the alginate shells 22 to more accurately recapitulate TME architecture in an ECM (different types of gels and scaffolds are discussed below). However, within spheroid formations, significant aspects of the tumor environment are not replicated because of the lack of stromal cells and their associated effects. On the contrary, there is a possibility to co-culture spheroids with immune cells,23,24 stromal cells, 25 or even both as a triple co-culture 26 in order to increase the validity of the model. There is even an option to co-culture spheroids with the tumor-colonizing bacteria, 27 which are an important part of the environment, especially in colorectal tumors. This strategy would be a promising one to study such modalities as microbial cancer therapies, which are lately occurring, thanks to the ability to combine the discovery of tumor-colonizing bacteria with advances in synthetic biology.
Undisputable advantage of spheroid models is their ability to simulate oxygen gradient. In the tumor, the abnormal vasculature leads to the defects in mass transport, especially in the case of oxygen. 28 This usually causes a formation of a necrotic core consisting of dead cancer cells due to an inadequate oxygen supply. As a result, creation of hypoxic regions occurs and these are present in the majority of solid tumors. The cancer cells on the periphery of the spheroid have, on the contrary, higher access to the nutrients and oxygen. The availability of oxygen in the spheroids is critical for metabolism and to the responsiveness to drug treatments. 29 Hypoxic cells usually switch to a glycolytic metabolism, which increases glucose uptake, decreases oxygen consumption, and alters the pH of the microenvironment. 30 The formation of a microenvironment within spheroid is determined by the balance between oxygen diffusion from the growth medium and its consumption by cells within the spheroid. In 2016, a simple non-invasive technique was modified for the quantitative measurement and subsequent evaluation of oxygen gradients in spheroids using electron paramagnetic resonance (EPR) oximetry. 29 Therefore, tumor spheroids can be used to model some characteristics of avascular tumors or micro-metastases of large solid tumors. In addition, this method enables to better replicate the barrier for drug penetration and thus allows to study the drug/therapeutics delivery in the pathophysiological context of the native tumor tissue.
The main benefits of spheroids are their low cost, high throughput, reproducibility, and ease of use, 31 but they are still not considered to be an ideal model for cancer therapies testing.
Tumor organoids
Tumor heterogeneity is known to be the major problem in the development of effective patient-specific therapies. Diverse tumor phenotypes usually change with the progression of disease and also as a reaction to different types of treatment. In addition to the intratumor heterogeneity, there is also a divergency across the patients that explains various patient-specific therapeutic responses 32 (Figure 1). Therefore, spheroids that are usually formed from cancer cell lines cannot provide sufficient conditions to circumvent this problem. The development of successful personalized therapies relies on the ability of laboratory models to accurately recapitulate not only microenvironment but also the heterogeneity of a given tumor. In the last few years, 3D organoid cultures established from patient tumor tissues are considered to become relatively representative model preserving the heterogeneity of the tissue and allowing the interactions with the TME. 33 It began in 2009, when Sato et al. established 3D epithelial organoids from a single leucine-rich repeat-containing G protein-coupled receptor 5 (LGR5) + intestinal stem cell. These highly polarized epithelial structures with proliferative crypt and differentiated villus compartments grow embedded in the MatrigelTM in a serum-free medium with several growth factors and inhibitors. 34 In the same year, Kuo’s laboratory introduced different organoid culture system using an air–liquid interface (ALI) with stromal support cells as a source of essential growth factors 35 that simplified used culture medium.
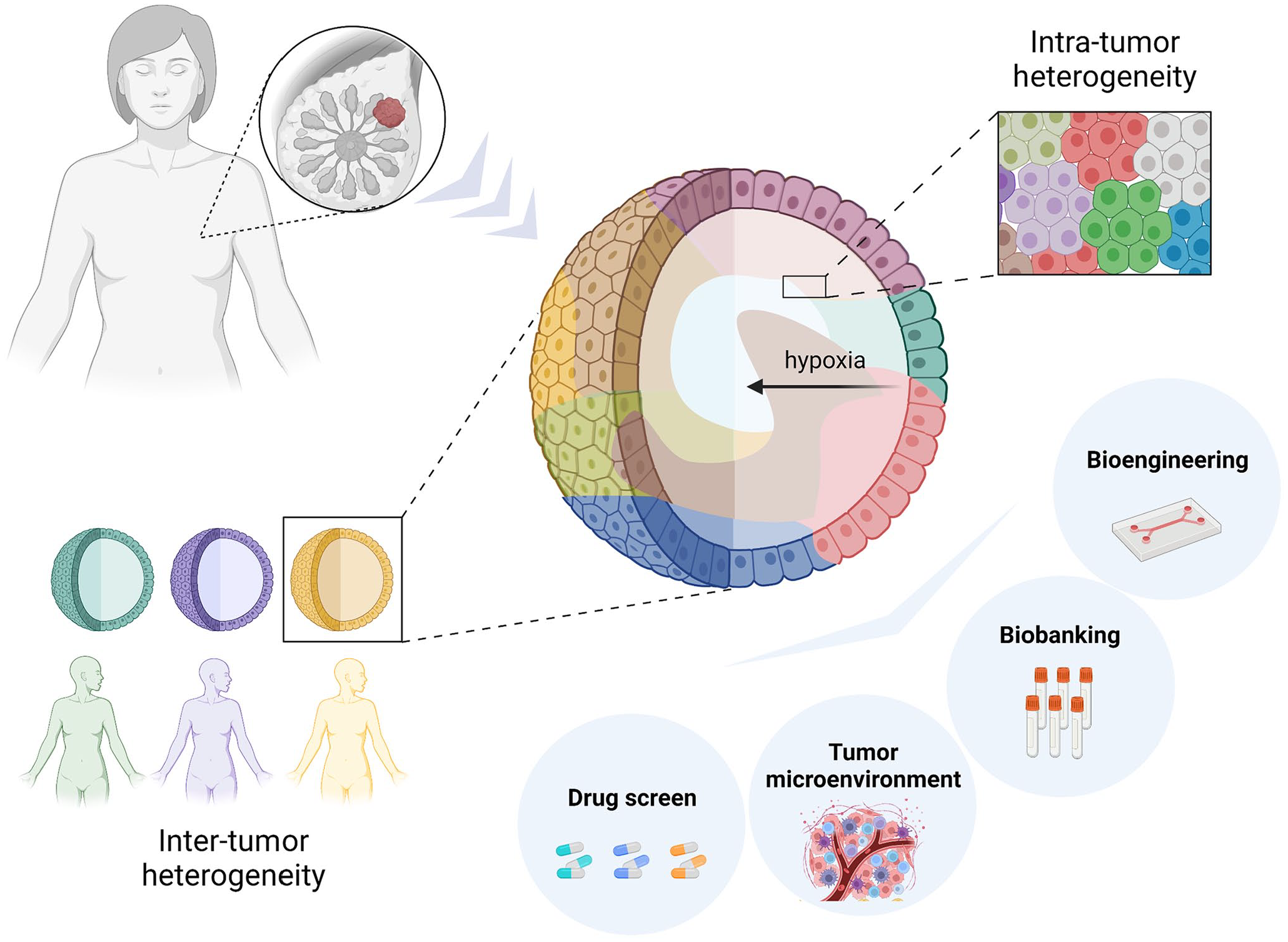
Tumor organoids established from patients’ tumor tissue represent inter- and intra-tumor heterogeneity. The hypoxic gradient is created similarly as in native tumors, once they reach a certain size. There is an opportunity to incorporate a tumor microenvironment, use the samples for biobanking, and drug screening, or incorporate them in bioreactors. Created with BioRender.com.
Subsequently, organoids culture protocol for other epithelial tissues, of mouse as well as human origin, were published following a great number of protocols for organoids establishment from tumor tissue and created so-called “living biobanks” (reviewed in LeSavage et al. 36 ). Each tumor has unique composition on cellular and environmental level, which contributes to the heterogeneity within and across tumors. Many publications showed that organoid models could accurately recapitulate this biological heterogeneity and maintain phenotypic and genetic characteristics of original tumor epithelium. Moreover, tumor organoid cultures seem to be suitable for modeling the TME heterogeneity and cell interactions using co-culture with different type of cells, such as cancer-associated fibroblasts (CAFs)37,38 or immune cells.39,40 As mentioned above, hypoxia, which greatly contributes to malignant behavior and chemo- and radio-resistance, does develop in organoids once they reach a certain size. 41 Therefore, tumor organoids established from primary tumors as well as from metastatic tissues have become a promising and eligible high-throughput platform for patient-specific testing.
Despite all the promising features mentioned above, organoids, as all other existing preclinical models, have their own limitations. The main disadvantage is the lack of standardized culture protocol. There is a great technical variability across the tumor organoid studies, including non-standardized tissue sources, variable protocols for their processing, different types of media formulation, and the use of heterogeneous animal-derived 3D matrices that are not able to properly mimic native tumor ECM. 36 It was shown that tumor organoids often grow slower than their matching normal organoid counterparts due to the higher rates of mitotic failures and subsequent cell death. 42 Unfortunately, there is a great chance of the overgrowth of healthy epithelial tissue organoids derived from normal tissue in tumor biopsies. 33 This could be avoided by removing as much normal tissue as possible from tumor biopsies by skilled pathologist or by using selective culture medium. For example, tumors with mutations in the epidermal growth factor receptor (EGFR) signaling pathway can be selected by EGF withdrawal. 43 Another obstacle is linked to the efficacy of organoid derivation and subsequent in vitro expansion. There is a great deal of evidence that many tumor types have an efficacy of organoid derivation higher than 80%, but for some of them, for example non-small lung cancer 44 , it is extremely low and unpredictable. Another major pitfall is the fact that the derivation time of most cancer organoids is currently weeks to months, which is unacceptable if they are supposed to be utilized as a tool of personalized medicine. Derivation time might be shortened if they were used as co-clinical models. This would have added value for the patient treatment setting.
Overall, given the great potential of cancer organoids to accurately recapitulate the intra- and inter-tumoral heterogeneity associated with patient-specific cancers, elimination of the technical variability in all steps of organoid derivation and in vitro expansion is necessary to establish reproducible robust platforms that can be translated into the patient care.
Assembloids
To look a little bit further into the future, organoids generated by spatially organizing multiple cell types, so-called assembloids, could allow more profound insight into the different processes and functions of the various types of tissues. One of such approaches, a simple mixing of different cell types into the organoid structures, was previously used, for example, to create brain organoids with vasculature-like structures. 45 These vascularized human cortical organoids acquired several blood–brain barrier characteristics, such as an increased expression of tight junctions, nutrient transporters, and trans-endothelial electrical resistance. Assembloids might help to look deeper into multiple biological processes and therefore be useful in research of several different complex diseases, including cancer. Even though aggregating different cell types into organoids is technically possible, it is hardly a model with defined spatial organization. This requires a much more advanced approach than just creating an organoid from a simple mixture of cell types. As an example, neuronal migration during development is a spatially organized process, which is not easy to recapitulate in vitro. Recently, a partial progress was made in this area when the researchers managed to create a model by fusing brain organoids of different regional identities. Moreover, they observed migration of interneurons from ventral to dorsal forebrain regions within the assembloids (shortly reviewed in Vogt 46 ).
Speaking of cancer research, only a few laboratories have succeeded to create a useful model of tumor assembloids yet. Multilayer bladder assembloids were developed by reconstituting tissue stem cells with stromal components to represent an organized architecture with an epithelium surrounding stroma and an outer muscle layer. Malignant tumor counterparts to these assembloids were developed to recapitulate the in vivo pathophysiological features of urothelial carcinoma. Here, they used connection of organoids with a connective tissue layer (patient-derived CAFs and endothelial HULECs), and the outer muscle layer (induced pluripotent stem cell (iPSC)-derived smooth muscle cells). 47 Pancreatic ductal adenocarcinoma (PDAC) assembloids were established from PDAC organoids, endothelial cells, and autologous immune cells to study cancer cell plasticity. In PDAC, the ability of tumor cells to dynamically switch between cancer-initiating and non-initiating cell states has been suggested as a reason for poor drug efficacy. Classic organoid model fails to recreate such cancer cells’ plasticity but assembloids containing plasticity-inducing endothelial and immune cells can. This model allowed to identify JAG1 as an essential player in this process and as a potential therapeutic target. 48
To reflect the complexity of processes in the human tissues, the complexity of organoids must be increased and assembloids are considered to be the nearest next step in this long journey. Different approaches in their assembly should be mixed and used to create highly complex, tissue-mimetic organoid models that could serve as very realistic laboratory models. These could be applied for studying the basic biology of human tissues and organs as well as for studying pathological processes.
Bioreactors
In the last few years, bioreactors and various types of flow systems have become indispensable tools in cell-based therapy research. They are used to maintain microenvironment and regulate cell growth, differentiation or tissue development, and therefore, are essential for providing standardized cell-based products for in vitro testing of various treatment agents. The bioreactors could be divided into three major groups: cell expansion bioreactors, tissue engineering bioreactors, and lab-on-a-chip systems. 49 There are a great number of companies with different modifications and improvements of bioreactors and microfluidic systems. Many of them are widely used in cancer research.
Microfluidics has greatly improved our ability to mimic the natural biophysical and chemical conditions of cell cultivation under in vitro conditions. There are different types of microfluidic platforms, so-called organ-on-chip or lab-on-chip, or in case of cancer tumor-on-chip devices, that can model several physiological functions of tissues and organs. These platforms combine the advantages of microfluidic technology and 3D cell culture models with improved recapitulation of the native microenvironment. 50 They allow performing high-throughput tests with decreased cost because of the usage of microscale volumes; they have the same ethical advantages as other in vitro systems with added bonus of enhanced reproducibility. 50 Their main advantage is the ability to modify and control various parameters independently—from the types and localization of cells or the orientation of tissue interface, to application of mechanical forces or chemical gradients. 51 Last but not least, the microfluidic channels allow the combining of different types of tissues into logical units, thus simulating the relationships between different distant organs and/or tissues.
Microfluidic chips are usually fabricated by technique called “soft lithography”—method, where the desired patterning is printed on silicon wafers by photolithography, followed by replica molding of the silicon wafer with a liquid polymer, usually polydimethylsiloxane (PDMS).50,51 PDMS is commonly used because of its numerous positive features such as, low price, gas-permeability, and transparency, allowing a high-resolution optical imaging. 52 On the contrary, the main disadvantage of this material is the unspecific absorption of small molecules,52,53 which is the main demand for the intensive search for a new material to replace PDMS in the manufacture of biochips. Other approaches for microchip fabrication include micromolding, microetching, laser etching, injection molding, photopolymerization, and 3D printing. 51 Fabrication method depends on the specific microchip application.
Most microfluidics platforms designed for cancer research are coated with ECM or even involve scaffold-based materials to better mimic the native TME. Multiple cell types can be cultured in a microfluidic device to analyze specific interactions between cancer cells and their stroma.54–56 For example, ECM remodeling is a typical consequence of tumor–stroma communication and subsequent activation, necessary for cancer progression. A model allowing to study this process was constructed by combining fibroblast-assembled ECM, mimicking a stromal compartment, and breast cancer cells (Figure 2, left). Authors observed that invasion of cancer cells led to the activation of CAFs and overexpression of both fibronectin and hyaluronic acid in the ECM. 57 Therefore, this is very unique in vitro platform allowing monitoring of the key factors in the switch between healthy and pathological stroma in cancer.
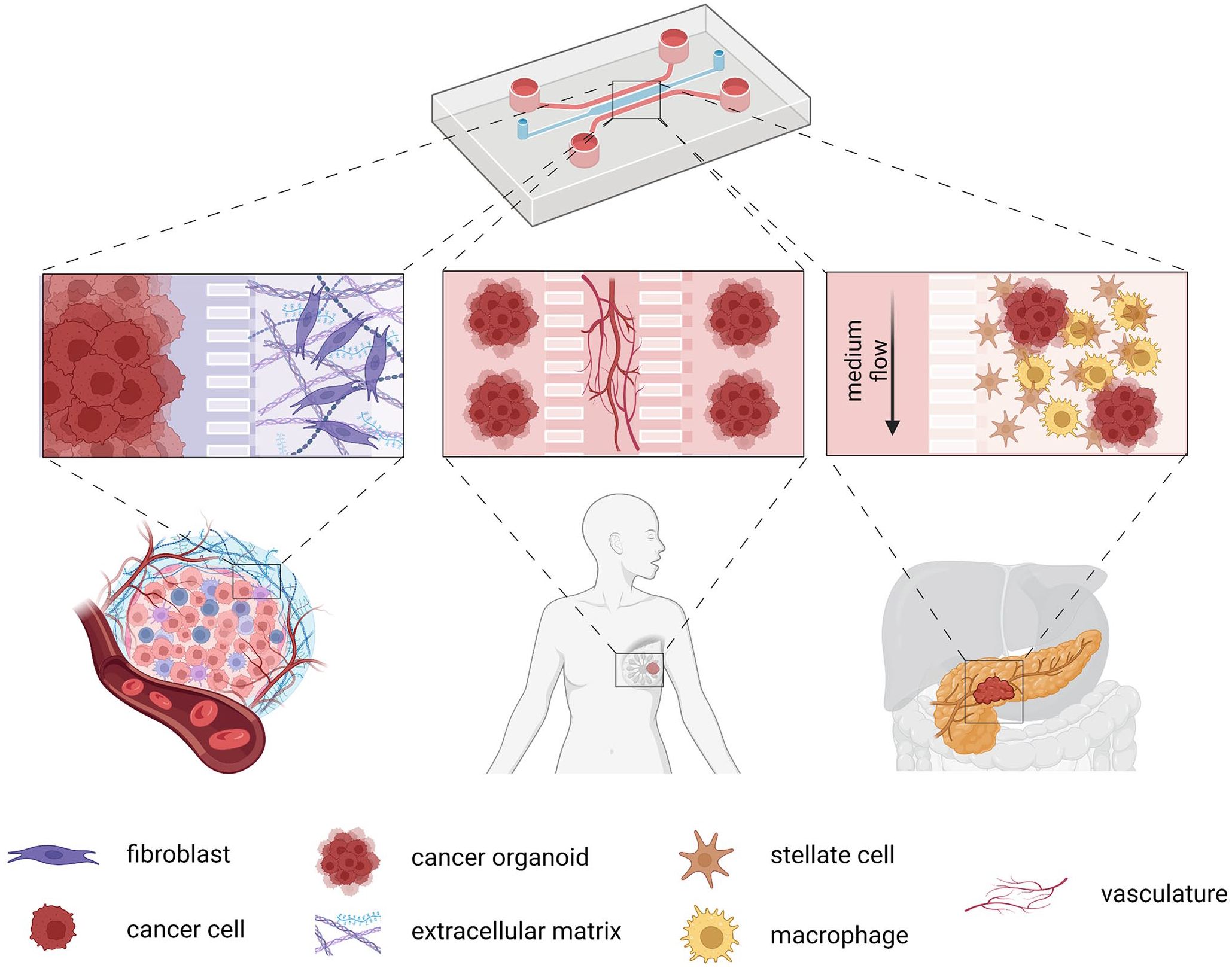
Schematic presentation of different approaches of lab-on-chip devices in cancer research: ECM remodeling processes, microvasculature-related processes such as angiogenesis or metastasis, and TME incorporation for different fields of cancer research. Created with BioRender.com.
Even more innovative models than a standard co-culture of cell lines in fluidic devices have recently been developed. Microfluidic chip systems combined with organoid models, called organoids-on-chip, combine benefits of both innovative approaches in cancer research and allow us to study tumor heterogeneity and tumor–stroma interaction on a much different level. For instance, a state-of-the-art multicellular chip device was engineered to mimic the PDAC TME by using patient-derived organoids (PDOs), pancreatic stellate cells, and macrophages (Figure 2, right). This tissue-chip model improved long-term cell survival of primary PDOs and significantly increased the chemotherapy effect on cancer cells, thus validating the use of the device for drug testing. 58 In addition, their system allows extending its complexity by incorporating vasculature and multiple types of immune cells.
Furthermore, the microfluidic chip devices incorporating a microvascular compartment have been developed to recapitulate the crucial features of tumors, such as tumor angiogenesis and metastasis. 59 This self-assembly microvascular approach can be used to reconstruct tumor angiogenesis and vasculature for analysis of patient-derived tumor cellular behavior and drug testing, or metastatic cascade where extravasation of tumor cells plays a crucial role (reviewed in Lim et al. 60 ). Also, in this case, multiplex microfluidic systems combining microvasculature and organoids have been developed (Figure 2, middle). The microvascularized 3D tissues retain their functions representing the original in vivo tissues. For example, there is a platform, where a central feature is a quiescent perfused 3D microvascular network created prior to loading of breast patient-derived tumor organoids in an adjacent compartment, which provides the opportunity to simultaneously and dynamically observe hallmark features of tumor progression, including cell proliferation, angiogenesis, cell migration, and tumor cell intravasation. Moreover, organoids are viable for several weeks and induce robust sprouting of angiogenesis. 61 Another research team combined PDAC organoids, human fibroblasts, and endothelial cells with the bio-scaffold that mimics perfusable vascularized vessels. Remodeling of tumor stiffness through myofibroblast contraction and collagen deposition was observed as well as decreased efficacy of a drug applied through the vasculature when compared with static organoid culture. 62
Modern approaches using combination of patient-derived tumor organoids and vasculature on microchip devices are the key to better understanding of cancer biology obstacles mentioned in this review. Representation of an “in vivo-like microenvironment” using platforms like these allows the development of new anti-cancer therapeutics and biological studies. As mentioned above the technology still has many imperfections and we are certainly not at the end of a journey. However, the number of options these technologies open for studying physiologically relevant interactions between different cell types (between tumor and TME), as well as the new ways to examine mechanisms of actions of candidate drugs in such constructs are certainly exciting. Further improvements of the limitations mentioned above should lead to increased efficiency of the preclinical stage of drug development and eventually to true customized patient-specific therapy.
Extracellular matrix, hydrogels, and scaffolds
As mentioned above, the 3D in vitro models were developed to recapitulate, at least partially, the complex architecture of real tissue/organ in a “Petri dish.” The principal environmental factors such as ECM composition affect greatly with the tumor development and metastasis. For instance, the ECM is directly involved in the regulation of tumor stemness properties, epithelial-to-mesenchymal transition (EMT), or drug response.63–65 In fact, the ECM composition changes with tumor progression. Higher concentration of some proteins, including laminin, fibronectin, or fibrillar collagens, is responsible for alterations in the ECM architecture and mechanical and physicochemical properties underlining the key role of ECM in tumor biology.66–69
The organoids establishment usually requires animal-derived ECM, which is commercially known under different names (e.g. Cultrex® or MatrigelTM). It has been considered a “gold standard” due to the spontaneous crosslinking at physiological temperature and biomolecular composition; it is derived from Engelbreth-Holm-Swarm mouse sarcoma tumors. This type of matrix is composed of a mixture of various basement membrane components, including laminin-1, collagen IV, entactin, heparan sulfate proteoglycans, and growth factors, and thus shows batch-to-batch variability in their composition. Apart from the remarkable tumor stroma mimicking characteristics and tissue-like physical features, usage of heterogeneous native ECM-obtained matrices generates discrepancies in organoid culture protocols and reduces the reproducibility and clinical compatibility by generating structures with variable cell-type composition, shape, and size. Therefore, enormous efforts have been put to develop bio-inspired products that might replace them and help to standardize the cultivation protocol and allow high-throughput screening.70–72
Hydrogels have emerged as novel supporting materials for 3D cell culture. They are able to form 3D polymeric networks and absorb and retain over 20% of their mass in water or other biological fluids. 73 In parameters such as viscoelasticity, mechanical features, and bioactivity, the hydrogels are akin to the native tissue ECM. Noteworthy, their microarchitecture (pore shape and size, fiber length, specific distribution of mechanical signals and motifs involved in cell adhesion, and chemical structure) allows them to be modeled to mimic the adequate microenvironments for cancer cells expansion as well as 3D culture development.74,75
Hydrogels are formed through physical and chemical crosslinking. In general, they are categorized according to their polymer composition. As an example, collagen, fibrin, gelatin, hyaluronic acid, and decellularized organ-derived ECM belong to natural hydrogels and are well known for their outstanding biocompatibility and bioactivity. These materials include native cell-binding epitopes (e.g. RGD motif) as well as degradation motifs for proteases that can cause uncontrolled hydrogel digestion. Moreover, the batch-to-batch variation of their natural sources may impair the hydrogels’ biochemical and biophysical tunability.76–79 Type I collagen hydrogels are frequently used as supportive scaffolds for growing cancer stem cells (CSCs), spheroids, and organoids. Some of the greatest benefits of collagen hydrogels are their ability to sustain the broad spectrum of linear elastic moduli (10–200 kPa), allowing the 3D culture to be treated with several different mechanical stimuli. 80 It was reported that human osteosarcoma and breast tumor spheroids, embedded in controlled pore size collagen hydrogels, can grow more efficiently, when the stiffness of the supporting matrix mimics authentically the cancer cell niche. Osteosarcoma spheroids proliferate more efficiently in stiffer hydrogels, while breast spheroids grow bigger in softer hydrogels. In addition, the breast cancer spheroids, grown in a proper mechanical environment, show ability to predict the in vivo response to chemotherapy more accurately. 21
Synthetic hydrogels based on polyacrylamide, polyethylene glycol (PEG), poly lactic acid (PLA), poly(lactic-co-glycolic acid) (PLGA), polycaprolactone (PCL), or polyurethane (PU) are commonly used as a scaffolds for tissue engineering. Their physicochemical characteristics are associated with lower batch-to-batch variance, higher stiffness, and tunability compared with hydrogels derived from natural sources. To fine-tune their features, the techniques for adjusting the polymer backbones and molecular weights of synthetic hydrogels are frequently used. Nonetheless, these hydrogels do not provide inherent principal biological stimuli and need to be conjugated with bioactive motifs and/or molecules (e.g. GFOGER, RGD, fibrinogen, and fibronectin) that mediate their biocompatibility.81,82
Semi-synthetic hydrogels, such as methacrylated hyaluronic acid (HAMA) or gelatin methacryloyl (GelMA), are formed by incorporation of crosslinking sites into the backbone of a natural polymer.83–85 These sites equip the semi-synthetic hydrogels with a stability and tunability that is lacking in natural hydrogels. Semi-synthetic hydrogels maintain biocompatibility and bioactive features to some extent since they are natural polymers derivates.86,87 The efficiency of GelMA hydrogels were tested in studying the invasiveness of breast cancer cells. GelMA hydrogels with stiffness of 4.8 kPa enabled MDA-MB-231 cells to form spheroids having an upregulated expression of stemness-related genes. Upon 5-day cultivation, cancer cells migrated out of the 3D spheroid structure showing an increased expression of genes involved in the breast cancer invasiveness compared to spheroids established under the standard scaffold-free and low-attachment conditions. In addition, GelMA hydrogels-embedded cancer cells exhibited enhanced tumorigenic ability in vivo. Six weeks after their intravenous injection, the tumor nodules could be identified in lungs and in the thoracic cavity. Such an invasive behavior was not observed in tumors generated from cells grown in 2D monolayers. 88
Therefore, ongoing extensive development of advanced hydrogels that lead to the generation of multifactorial tissue-specific signals, and the validation of the reliability of specific composite hydrogels for 3D cultures will be crucial to make practical differences across numerous downstream applications in the near future.
Three-dimensional bioprinting
Thanks to the development of laser technology, computer-aided design techniques, and digital microelectronic devices, 3D printing has developed significantly in recent decades. The ultimate goal of this technology is to provide tools or approaches capable to create 3D constructs with a structure and composition similar to native tissue and involve sophisticated patterning of the ECM, biomolecules, and even cells to study biological processes or create living structures/tissues. 89
These technologies could further bridge the space between 2D cell cultures and animal models. Although, in vitro and in vivo models are gold standard and indispensable in preliminary tests for safety, efficacy, and cytotoxicity, as mentioned above, those traditional models have many limitations, with the high costs and ethical concerns being just the basic ones. 90
As stated before, the leaps made in the development of 3D cultivation greatly improved the ability of in vitro models to reproduce in vivo situation. However, achieving proper spatial organization within these structures is still a major pitfall; an internal architecture of many of them can still be considered unorganized. 91
The 3D bioprinting techniques may be just a tool that is needed here. A 3D-printed construct can have a defined architecture and mechanics modulated by individual fibers’ orientation, density, and arrangement.92,93 These can also be modified in different ways to create multiple layers with different porosity and nutrient supply to support the cell migration, proliferation, and differentiation in the construct. Because of the possibility of easy customization, which could be a part of personalized medicine, the product could be tailored to a specific patient. 94
The birth of 3D printing is dated back to 1984 when stereolithography (SLA) was invented, printing 3D objects from digital data. 95 The first bioprinting was carried out in 1988 using a standard inkjet printer. 96 Since that time, thanks to advances in both material science and technology, many bioprinting products have been introduced. In 2002, the first extrusion-based bioprinter, a predecessor of the later commercialized 3D-bioplotter, was presented. 97 Since that year, many printers have been developed and delivered, such as the first ink-jet bioprinter, commercial SLA printer, 98 or integrated tissue-organ printer. 99 Various procedures have created many different constructs such as the first 3D tissue with only cells (not scaffold), 100 scaffold-free vascular construct, 101 3D printed tubular structure, 102 cartilage model, bioprinted cardioid structure, and collagen human heart,103,104 skin, 105 nerve, 106 bone,107,108 cartilage109,110 and so on.
The biggest challenge in producing a 3D bioprinted cancer model is to imitate the TME as faithfully as possible. Such complexity of the TME in vivo cannot be simulated using 2D monolayer cultivation or 3D co-cultivation models. It was proven in a study by Herrores-Pomres et al., 111 where they compared 2D culture and three different 3D printed platforms, rigid scaffold, hydrogel-based scaffold, and suspension spheroid culture enriched in CSCs from non-small-cell lung carcinoma. They observed different proliferation profiles of cancer cell lines and primary cancer cells on various scaffolds. Also, gene expression analysis confirmed that tumor spheres and cells seeded on hydrogel scaffolds significantly overexpress most of the stemness and invasive promoters tested compared to control cells grown in 2D culture. Their findings provide strong evidence of advantages of 3D-printed models, especially hydrogels as the primary scaffold material for studying CSCs, due to mimicking tumor complexity and regulating cancer cell behavior. Similarly, another scientific team compared cell proliferation, matrix metalloproteinase (MMP) protein expression, and chemoresistance in the printed 3D cervical tumor models and conventional 2D planar culture models, 112 where HeLa cells showed a higher proliferation rate in the bioprinted 3D environment and tended to form cellular spheroids, but in 2D culture they formed monolayer cell sheets. HeLa cells in 3D printed models also displayed higher MMP protein expression and chemoresistance than in the 2D culture. 112 Also, 3D bioprinted osteosarcoma model (3DBPO), containing osteosarcoma cells and photo-cross-linkable bioinks composed of gelatine methacrylamide and hyaluronic acid methacrylate, was compared with 2D models and tumor spheroids. The 3DBPO model showed significant changes in cell cycle, metabolism, adherens junctions, and other pathways associated with epigenetic regulation and was also more sensitive to therapies targeted to the autophagy pathway. 113 Currently, the studies involving 3D bioprinted tumor models are trending massively, as evidenced by the great number of works reporting the preparation of 3D models of various malignancies such as malignant melanoma, 114 multiple myeloma, 115 ovarian cancer, 116 brain tumor, 117 breast cancer, 118 and urological carcinomas. 119
Three-dimensional bioprinting techniques can also be used to study the metastatic process. Holzapfel et al. 120 aimed to engineer a morphologically and functionally intact humanized organ bone that could serve as a homing site for human prostate cancer (PCa) cells. Human mesenchymal progenitor cells were seeded on the bone structure, and recombinant human bone morphogenetic protein-7 (rhBMP-7) growth factor was used to induce the metabolic activity and production of ECM components. Upon further culture, the PCa cells demonstrated a preference for the engineered bone constructs, proliferated, and developed macro-metastases. Similarly, a microtissue model of osteoblastic bone metastases was prepared using cultured primary human osteoprogenitor cells on a polymer scaffold and created a mineralized osteoblast-derived microtissue containing viable osteoblastic cells, osteocytic cells, and appropriate expression of osteoblast-/osteocyte-derived mRNA and proteins and mineral content. 121 Co-cultured cancer cells showed increased affinity to the microtissue, upregulation of alkaline phosphatase and collagen-I, and sclerostin downregulation, consistent with the clinical marker profile of osteoblastic bone metastases. 121
The knowledge/understanding gained from the systematic and methodological study of the complexity and dynamics of the TME using 3D bioprinted constructs can speed up the translation of research into clinical practice by accelerating the drug development process, reducing costs, saving resources, limiting animal testing, or providing a higher level of personalization. There are still some challenges discussed here that must be overcome before we are able to translate research into clinical practice, but these advances in material science and technology could mean major progress in cancer research. One such example is a recent development of new “smart” materials that enable a shift even into the so-called fourth dimension. So-called four-dimensional (4D) printing of dynamic 3D objects that can change their morphology and/or characteristics in time by external stimuli, leading to shape-shifting abilities, seems to be very promising in drug testing or disease modeling for a patient-centered approach. 122
Conclusions and future direction
There is a great deal of evidence highlighting the importance of tumor–stroma communication in various processes related to cancer progression, including drug resistance. Therefore, creating better laboratory models relevantly mimicking the interplay between the tumor and stroma would lead to the development of more effective anti-cancer therapies. In general, tumors need to be cultured in a physiologically relevant environment resembling the processes and niche as in the native tissue. To achieve such progress in this field, patient-derived 3D structures preserving the tumor heterogeneity together with variable cells and molecules from the TME should be cultured together in a specifically organized manner in xeno-free ECM. This can be accomplished by the integration of state-of-the-art technologies such as polymeric scaffolds, 3D bioprinting, and organ-on-chip platforms. Such a combination should allow to better mimic disease and develop personalized medicine programs. On the contrary, the increased complexity of the models could also complicate their analysis. However, nowadays this aspect can be, at least partially, solved by incorporating in silico analyses and/or artificial intelligence algorithms into the whole process, but that is the theme for another review.
To sum up, 3D culture models (available 3D cancer models with their advantages and limitations are summarized in Table 1) that combine primary tumor cells with their microenvironment together with various flow technologies have the potential not only to bridge the gap between 2D cell cultures and animal models, but become completely new testing platform for personalized medicine. In the future, the increasing complexity of these models and their integration with information technologies should allow us to achieve comparable outputs to those obtained in vivo without the involvement of experimental animals. Therefore, the future laboratory models in the oncology field, especially models used for personalized medicine, will rely on the interdisciplinary collaboration between various fields such as biology, medicine, physics, chemistry, mathematics, and bioinformatics. By using an increased number of 3D modeling as a preclinical tool, the number of animal studies may be reduced and lead to a more ethical approach in cancer research and better anti-cancer therapies.
Overview of currently used 3D cancer models and technologies with their advantages and limitations.
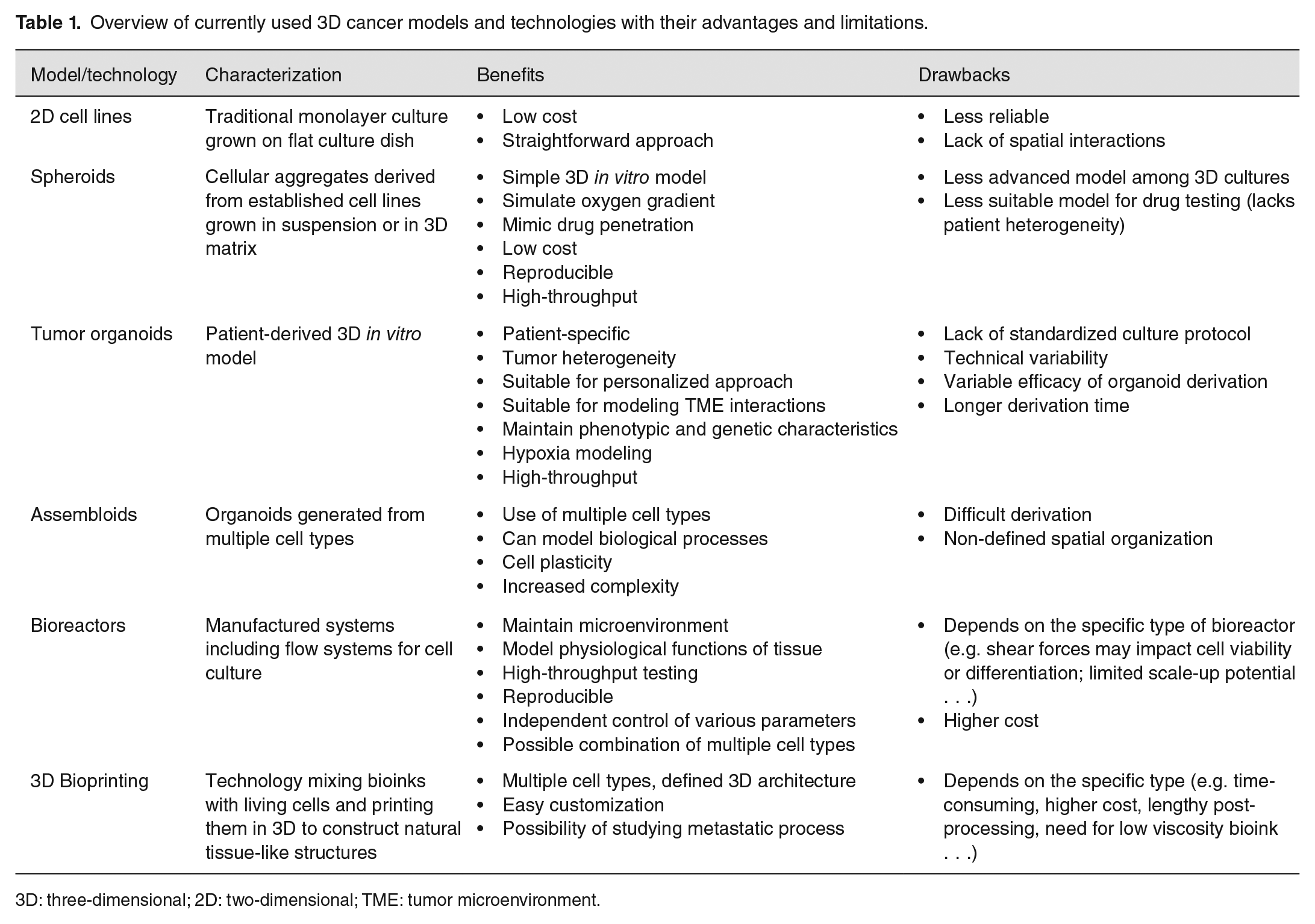
3D: three-dimensional; 2D: two-dimensional; TME: tumor microenvironment.
Footnotes
Authors’ Contributions
All authors contributed equally to the conceptualization and writing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This publication was created with the support of the Operational Program Integrated Infrastructure for the project BIOFORD [ITMS2014+ 313011AFG5], co-financed from the European Regional Development Fund and the state budget of the Slovak Republic, and Scientific Grant Agency (VEGA) (grant numbers 2/0067/22 and 2/0138/20).