
Abstract
The capsid has a central role in viruses’ life cycle. Although one of its major functions is to protect the viral genome, the capsid may be composed of elements that, at some point, promote interaction with host cells and trigger infection. Considering the scenario of multiple origins of viruses along the viral evolution, a substantial number of capsid shapes, sizes, and symmetries have been described. In this context, capsids of giant viruses (GV) that infect protists have drawn the attention of the scientific community, especially in the last 20 years, specifically for having bacterial-like dimensions with hundreds of different proteins and exclusive features. For instance, the surface fibrils present on the mimivirus capsid are one of the most intriguing features of the known virosphere. They are 150-nm-long structures attached to a 450-nm capsid, resulting in a particle with a hairy appearance. Surface fibrils have also been described in the capsids of other nucleocytoviruses, although they may differ substantially among them. In this mini review for non-experts, we compile the most important available information on surface fibrils of nucleocytoviruses, discussing their putative functions, composition, length, organization, and origins.
Impact Statement
Considering the scenario of multiple origins of viruses along the viral evolution, many peculiar structural features have been reported and described for nucleocytoviruses. In this context, capsids of some of these so-called giant viruses (GV) are highlighted by surface fibrils, which are responsible for adhesion and interaction with their host membranes, such as those in amoeba. Knowing the singularities of these structures promotes the deepening of fundamental knowledge on the biology of these members within the virosphere, ultimately shedding light on their adaptive convergence in evolution, virus–host interactions, and their biological cycle, as well as possibilities for their proteins and viral assembly.
Introduction
A glimmer of microbial evolution: giants among viruses
The term “giant viruses” has been used to designate a putative monophyletic group of viruses belonging to the phylum Nucleocytoviricota, infecting uni- and pluricellular, hetero- and autotrophic organisms, from protists to animals. 1 Some authors consider bona-fide giant viruses those with capsids larger than 500 nm, which are easily visible by optical microscopy. However, the term “giant viruses” has also been used to refer to smaller nucleocytoviruses, with particles ranging from 150 to 500 nm. Giant viruses are ubiquitous and have already been isolated from different countries,2–5 from different environmental6,7 and clinical samples. 8 The first virus to be called a giant virus was Paramecium bursaria chlorella virus 1 (PBCV-1), which infects algae and which has particles up to approximately 190 nm in diameter, 9 and a dsDNA genome of approximately 330 kb. 10 Although nucleocytoviruses include historically important representatives, such as poxviruses, 11 the discovery of mimivirus in 2003 12 highlighted the remarkable structural complexity of the virions of this group.
Among mimiviruses, the Acanthamoeba polyphaga mimivirus (APMV) 12 isolate was the first amoebae-associated GV to be described. APMV was isolated from water samples collected from a hospital cooling tower during a pneumonia outbreak in Bradford, England. After analyzing these samples by Gram stain, researchers noticed small purple-stained dots inside amoebae, similar to gram-positive bacteria. However, after the failure of several techniques used in the identification and characterization of bacteria, questions about the nature of these organisms remained for years. After almost a decade, new techniques were used to study the mysterious microorganism, such as genome sequencing and electron microscopy, which led to the surprising discovery of an actual virus, and not bacteria. 12 This virus attracted the attention of the scientific community due to the large size of its particle and its genome. The APMV particle is approximately 750 nm in diameter, and its genome reaches the million mark, about 1.2 megabases (Mb). Since then, several other amoeba GV groups have been discovered, such as marseillevirus, 13 pandoravirus, 14 pithovirus, 15 and cedratvirus, 16 among others.
There is great diversity within giant viruses, considering their morphological characteristics. In general, the capsids of giant viruses do not present an external envelope. Instead, the capsids surround an inner lipid sac, which contains the viral genome and proteins related to the early phases of the replication cycle. Particles can have icosahedral symmetry (e.g. marseillevirus 13 and faustovirus 17 ), or pseudoicosahedral symmetry, as in mimiviruses, due to the presence of a stargate, that is, a vertex at the capsid that allows for DNA release. 18 They may also be oval (e.g. pithovirus 15 and pandoravirus 14 ) or even round-shaped (e.g. mollivirus 4 ). For instance, one of the largest viruses ever described, Tupanvirus, has a capsid attached to a tail variable in size, allowing the particle to reach up to 2.3 µm. 19 In some GV, we can find some structures decorating their capsids, such as the spherical-headed spikes that cover medusaviruses’ surfaces and mimiviruses’ fibrils. 20
Viral fibrils: an intriguing structural feature among giants
As aforementioned, mimiviruses exhibit particles with a capsid of approximately 750 nm, being 450 nm in diameter and covered by a dense layer of fibrils (~150 nm), 21 which were suggested to resemble gram-positive bacteria. 12 This intriguing misunderstanding would be feasible due to its structural fibrils’ composition, which is morphologically unique among these viruses, 22 and not being fully elucidated yet, 23 with all that is currently known being related to mimiviruses. The fibrils are embedded in a dense layer of peptido glycan-like structures 24 that stain crystal violet in a Gram stain, as those peptidoglycans that are present in the walls of gram-positive bacteria. 25 Important to the adhesion onto the amoeba surface (Figure 1(A)), as for APMV, fibrils are often found with one of their ends free while the other end is attached to the viral capsid. 26 Moreover, scanning electron microscopy (SEM) and transmission electron microscopy (TEM) analyses have shown different densities of fibrils on GV capsid surfaces, which are simultaneously acquired with the genome acquisition during morphogenesis in the fibril acquisition area, located at the periphery of the viral factories 27 (Figure 1(B)). In addition, fibrils’ peptidoglycan-like structures exhibit successive so-called rings of density (Figure 1(C)), which supports their key role in viral entry, 26 as exemplified for APMV (Figure 1(D)).
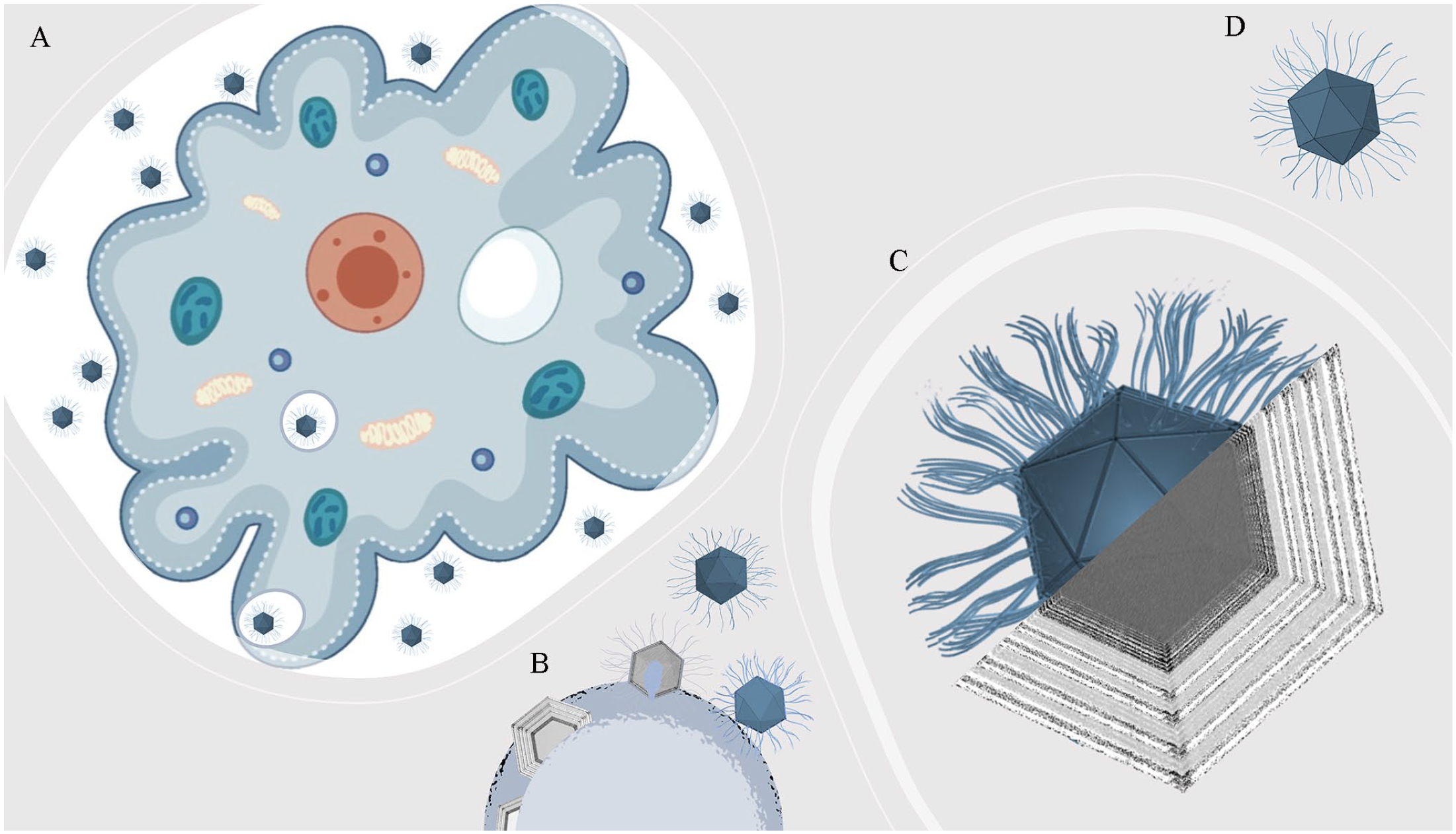
Fibrils’ role for giant viruses. (A) Fibrils (~150 nm) are important to the adhesion of mimiviruses onto amoebas, such as APMV. (B) During the morphogenesis, fibrils seem to be acquired in the fibril acquisition area, located at the periphery of the viral factory, concomitantly with the genome acquisition. (C) Fibrils’ peptidoglycan-like structures exhibit successive rings of density under electronic microscopy. (D) Schematic representation of a mature APMV particle. Amoeba image was generated from free vectors available at Vecteezy (https://www.vecteezy.com).
Many GV also present fibrils (Figure 2) as APMV, such as: (1) tupanviruses, with a capsid of approximately 450 nm in diameter and a tail attached to it, both covered in fibrils, resulting in particles varying from 1.2 up to 2.3 µm in length; 19 (2) Cotonvirus japonicus, 28 a mimivirus isolate, with approximately 400 nm in diameter and also surrounded by surface fibrils of approximately 100 nm; (3) Marseillevirus, 13 with approximately 250 nm of diameter, and 12 nm fibrils surrounding its particles’ surface; (4) PBCV-1, an algae infecting virus with approximately 190 nm in length and small fibrils around the capsid, of nearly 19 nm; 9 (5) orpheovirus, with particles larger in length than APMV (up to 1.1 µm), but presenting smaller fibrils when compared to these mimiviruses, as well as two tegument layers between the fibrils and the inner membrane; 29 (6) Mollivirus sibericum, 4 an almost spherical GV with 500–600 nm in diameter and surrounded by a fibrillar tegument as a mesh of fibrils; (7) cedratviruses, around 500 nm up to 1 µm in length,16,30 whose viral factory can be divided in two, one being associated with its fibrils acquisition; and finally (8) Yasminevirus, 31 the first Klosneuvirus to be isolated and that presents a particle with approximately 330 nm covered by a thin layer of fibrils.
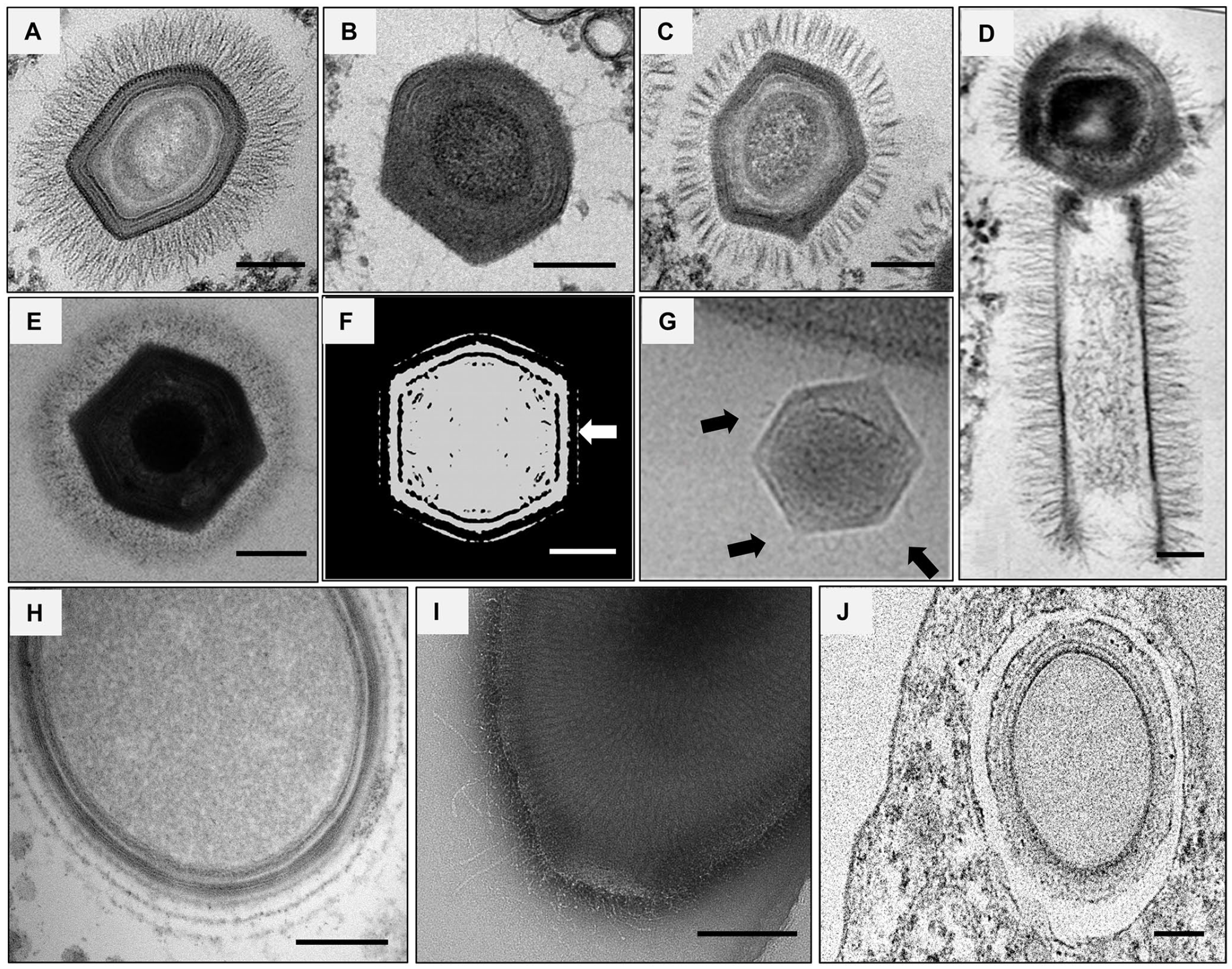
Giant viruses that possess surface fibrils. Electron microscopy images and shaded-surface representation of different GV isolates and their fibrils. (A–C) Lineage A, B, and C mimiviruses: mimivirus puntaullmanensis, moumouvirus crenensis, and megavirus caiporensis, respectively. (D) Tupanvirus particle, with its tail, a trademark of this group (adapted from DOI: 10.1038/s41467-018-03168-1). (E) Cotonvirus japonicus (adapted from DOI: 10.1128/JVI.00919-21). (F) Marseillevirus shaded representation illustrating the globular portions of its fibrils indicated by the white arrow (adapted from DOI: 10.1038/s41467-018-03168-1). (G) PBCV-1 (adapted from DOI: 10.1073/pnas.1107847108) fibrils are pointed by the black arrows. (H) Mollivirus sibericum (credits: provided by Dr Chantal Abergel, IGS UMR7256 CNRS-AMU). (I) Cedratvirus pambiensis. (J) Orpheovirus IHUMI-LCC2. Scale bars: 150 nm.
For instance, Notaro et al. demonstrated that fibrils are glycosylated with two different polysaccharides in their composition, whose structures have been elucidated by nuclear magnetic resonance (NMR) analysis, one being
Notaro et al. also showed rare amino-acid sugars to be synthesized from different GV of the subfamily Megamimivirinae, which are absent from their hosts and usually found only in bacteria: (1)
Harboring giants’ structural repertoire: genes associated with fibrils
Initially, it was speculated that the mimiviruses’ hairy-like appearance could be related to the large number of open reading frame exhibiting the characteristic collagen triple-helix repeat. Thus, fibrils could be linked to collagen and glycosylated proteins, consisting of cross-linked glycosylated collagen. 34 However, the fibrils are resistant to collagenases, even after pre-treatment with lysozyme, suggesting that the fibril surfaces are not collagen-linked. 35 For instance, the L829 and L725 genes, which codify proteins with unknown functions, and the R135 gene, which codifies a putative GMC-oxidoreductase, were also associated with mimivirus fibrils in previous works.36,37 Boyer et al. obtained an artificial mimivirus strain called M4 after several blind passages of APMV in axenic amoebae culture. M4 has a reduction of ~200,000 bp in its genome, and its particles have a bald appearance with fewer fibrils. After analysis of purified viral fibers by 2D gel electrophoresis coupled with matrix-assisted laser desorption/ionization mass spectrometry, it was reported that there was an absence of R135, L829, and L725 proteins in M4 particles when compared to the original mimivirus M1. They also compared the protein glycosylation patterns in the M1 and M4 viruses and observed that L829 and R135 proteins were glycosylated only in M1. 36 Furthermore, L725, which is encoded by an ORFan gene, that is, genes with no detectable sequence similarities in databases, 38 was suggested to be associated with fibrils by RNA silencing experiments. 39
In this sense, the R135 protein, APMV’s putative GMC-oxidoreductase, was pointed to be part of the fibrils composition and of the helical protein shell that encases mimivirus genomic material by cryo-electron microscopy, cryo-electron tomography, and proteomics,32,37 with the only difference between the fibrils and the genomic fiber composition being the presence of a Cys-Pro rich N-terminal domain only in the fibrils. In addition, Aquino et al. also discussed those genes in contrast to their previous functionalities or relationships with fibrils, as isolates with fewer fibrils (e.g. Borely moumouvirus) are morphologically similar to mimivirus M4 and present R135 and L829 genes in their genomes. 40 Interestingly, these three predicted proteins can also be found only in some GV that have fibrils when analyzing search results using the BLASTp tool 41 (Table 1) from the National Center for Biotechnology Information (NCBI). Herein, the best hits were found for all the proteins among mimiviruses from lineages A, B, and C, and for Cotonvirus japonicus, even though lineage B mimiviruses are almost fiberless. As for tupanviruses, a best hit for R135 was not found, and only a best hit for L829 was found for molliviruses, marseillevirus, and cedratvirus. No best hits were found for any of the proteins in PBCV-1 or yasminevirus, which also raises questions about the relationship of these proteins with GV’s fibrils. Furthermore, it is also important to highlight that the best hits were also found for R135 and L829 in viruses that do not have surface fibrils described as part of their particles, like pandoravirus and Pithovirus. 40
BLASTp best hits for the three genes associated with fibrils: L725, L829, and R135.
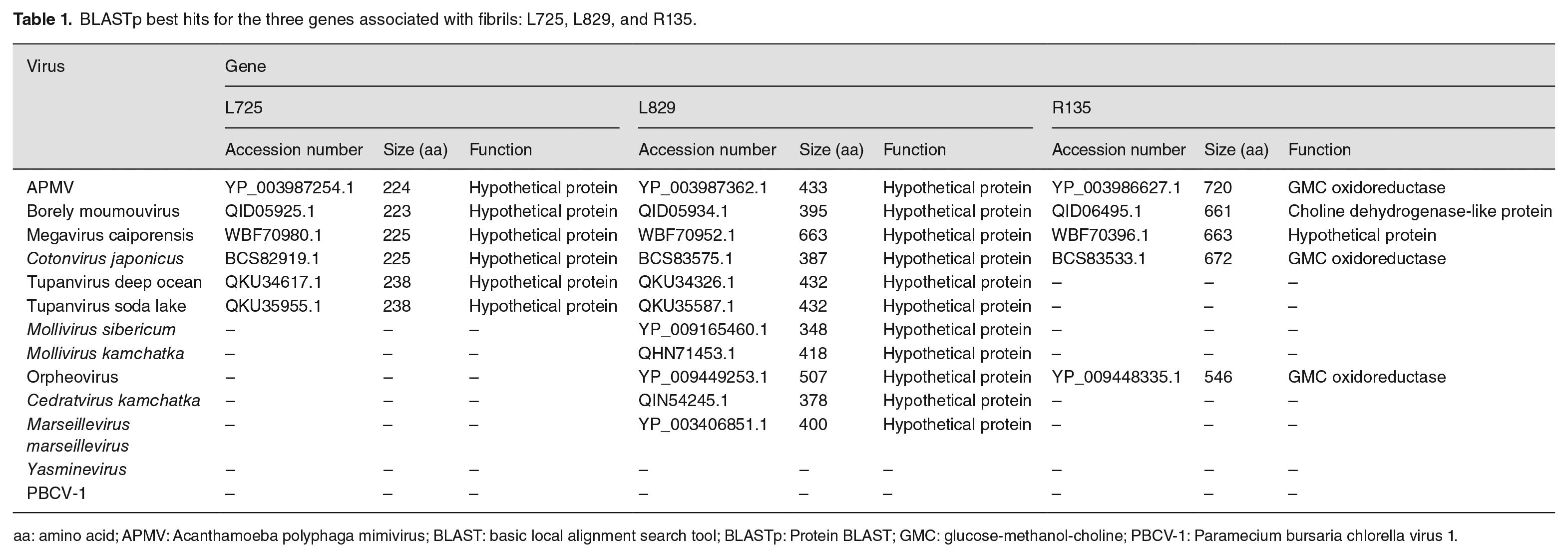
aa: amino acid; APMV: Acanthamoeba polyphaga mimivirus; BLAST: basic local alignment search tool; BLASTp: Protein BLAST; GMC: glucose-methanol-choline; PBCV-1: Paramecium bursaria chlorella virus 1.
Viral attachment: depicting the role of fibrils
Considered intriguing structural features of viruses’ morphology until then, fibrils stood out in early studies, and their function remained unknown for more than 10 years after the discovery of APMV. As mentioned, it was described that they play an important role in triggering the host amoeba’s phagocytosis by promoting the adhesion of viral particles to the cell surface, mediated by glycans. 24 Interestingly, in the presence of high concentrations of certain glycans (e.g. GlcNAc), interaction of the viral particles with other molecules would be prevented, due to the fibrils being saturated by a given carbohydrate. In addition, a smaller number of fibrils does not alter APMV replication but decreases its attachment to Acanthamoeba castellanii cells. 24
Rodrigues et al. treated APMV particles with different enzymatic conditions, such as lysozyme and lysozyme followed by bromelain and proteinase K, both in comparison to APMV and the M4 mimivirus. Interestingly, the presence of more fibrils leads to an increase in viral attachment to the amoebal surface when mediated by mannose and GlcNAc, no changes in viral titers were observed in the presence of glucose or N-acetylgalactosamine (GalNAc) at any of the concentrations assessed. The presence of mannose and GlcNAc at different concentrations (>50 and >25 µg/mL, respectively) reduced the viral titer up to 1000-fold. In addition, APMV fibrils also attach differentially to distinct organisms or structures by their glycoside interactions, such as Aedes sp. legs (i.e. chitin), Aspergillus fumigatus (i.e. mannose and chitin), Staphylococcus aureus and Escherichia coli (i.e. peptidoglycan), resulting in 18-fold, 7.32-fold, and no differences of the viral particles compared with the control, respectively. Finally, particles were saturated with chitin and peptidoglycan, and the presence of both polymers interfered with APMV adhesion to A. castellanii cells, with up to a 100-fold reduction in viral titer at concentrations >25 and >75 µg/mL, respectively. 24
These results could suggest that the peptidoglycan-like fraction of fibrils would also be restricted to interactions with Acanthamoeba cellular plasma membranes, 42 which are rich in lipophosphoglycan, 43 as well as potentially guaranteeing their host specificity. Furthermore, other functions have been suggested for fibrils, such as: (1) optimizing phagocytosis by expanding particle size; 44 (2) stimulating phagocytosis in amoeba by partially mimicking bacteria with peptidoglycan-like compounds, which are “food” of amoebae; 45 (3) acting as a natural decoy for hosts; and (4) increasing resistance under adverse conditions, but more studies remain necessary 26 to better understand those propositions.
Uniqueness of an intriguing structure: diversity in surface fibrils pattern
A first glimpse of fibril-like structures was observed with SEM and TEM methods for chloroviruses termed “fibers” at the time. 46 Subsequent structural 26 studies of amoeba GV, such as mimiviruses, helped in the discovery of other GV 22 and their fibrils. As fibrils of mimiviruses are polysaccharide-containing structures likely built from their glycosylation machinery, which decorates the capsid as surrounding structures, 11 one could argue what the disposition, organization, and composition of different GV’s fibrils would impact their biological cycle and even host specificities. Cultivation-independent approaches (e.g. metagenomics) have enhanced the discovery of new genome sequences of potentially new GV, with the potential for structural and functional diversity of fibrils. Although there are some similarities in how these viruses enter host cells (e.g. fibrils triggering phagocytosis), much is yet to be found about GV and their fibrils, as they can be found nearly anywhere on Earth, and only a small fraction of GV’s genomes have been discovered so far.11,47 In this sense, experimental characterization and validation, as demonstrated by Aquino et al., help unravel these differences among different GV’s fibrils. Authors showed that mimiviruses from lineages A (e.g. APMV), B (e.g. Borely moumouvirus), and C (e.g. Megavirus caiporensis) present differences regarding their surface fibrils organization and disposition. SEM and TEM allowed authors to observe that fibrils are (1) long and abundantly surrounding the capsid; (2) fewer in number and less homogeneously distributed; or (3) similarly abundant to the first, but organized in small groups (i.e. clusters or clumps). 40
For instance, APMV fibrils are homogeneously distributed, whereas Megavirus caiporensis appears in clumps or clusters. Both were estimated for their relative abundance of surface fibrils, whereas Megavirus caiporensis exhibited a 552-fold relative average contrast increase when compared to Borely moumouvirus, in comparison to a 394-fold increase for APMV. In addition, different combinations of the three mimiviruses mixed with purified particles were also compared by TEM, disregarding a potential SEM preparation influencing fibrils appearance and conformation. In addition, adhesion and entry into A. castellanii were also quantitatively assessed at a multiplicity of infection (MOI) of 10, showing that APMV and Megavirus caiporensis particles bind to the host plasma membrane approximately 10 times more than Borely moumouvirus after 1 h post infection. Finally, more APMV-infected cells were observed (43/50), followed by Megavirus caiporensis (16/50), and Borely moumouvirus (4/50), both under the same experimental conditions. 40 These results could suggest that the pattern of fibrils may also affect adhesion and their incorporation by amoebas.
Notwithstanding, other GV surface fibrils present some characteristics that distinguish them structurally from those of mimiviruses, such as Cotonvirus japonicus, 28 which has denser and shorter fibrils in its surface when observed by TEM analysis, which revealed a smoother arrangement when compared, for example, to lineage A mimiviruses’ particles. As aforementioned, artifacts caused by preparation procedures cannot be excluded, but these structural disposition and size variations are also observed for orpheoviruses, which have larger particles than other mimiviruses (e.g. APMV) but present shorter fibrils. 29 In PBCV-1, its external fibrils extend from some of its capsomers and potentially facilitate particle attachment to algae hosts, 48 similar to the mimiviruses’ mechanism initiating viral infection by attaching to these hosts’ cell walls.
Current knowledge about fibrils: issues and needed studies
The absence of a three-dimensional (3D) solved fibril structure is still a limiting factor for further structural studies, including its binding mechanism onto host membranes during viral entry. Expression, purification, and obtaining crystallized or cryo-EM structures would help in predicting and performing subsequent binding or enzymatic assays. For instance, near-atomic resolution structures of different nucleocytoviruses,49,50 yet one of the largest groups of viruses that infect eukaryotic hosts, could also help unravel possibilities for their proteins and assembly, 50 such as fibrils structural organization.
Moreover, in-depth studies about genes that have already been related to mimivirus fibrils 36 are also needed, as is the expansion in search of new genes, both in mimivirus and in other GV with fibrils, which remains a field to be explored. In addition, much of the existing information about these structures was obtained using mimiviruses as a study model, and it would be interesting if new studies with other giant viruses were carried out with the purpose of raising more data and knowledge about the possible diversity of fibrils. In this sense, it was shown that Megavirus chilensis has a gene cluster responsible for producing glycosylated proteins that can be associated with fibrils, and that this profile is shared with several other megaviruses, 51 still in need of more studies. Furthermore, if we consider L725, L829, and R135 as fibril-associated proteins,32,36,37,39 the absence of them in some GV that presents fibrils (Table 1) could mean that other genes are responsible for the formation of fibrils in other species of nucleocytoviruses, and it is important to consider that perhaps this characteristic is an adaptive convergence in the evolution of GV.
Footnotes
Authors’ contributions
ILMdA, MGB, TBM, MSMS, and JSA wrote the review.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (grant number 303680/2022-9); Coordenação de Aperfeiçoamento de Pessoal de Nível Superior/Ministério da Saúde (grant numbers 88882.348380/2010-1, 88887.595578/2020-00, and 88887.684031/2022-00); Fundação de Amparo à Pesquisa do Estado de Minas Gerais (PPM-00732-18); Pró-Reitoria de Pesquisa e de Pós-Graduação from Universidade Federal de Minas Gerais (04/2022); and Centro de Microscopia from Universidade Federal de Minas Gerais (1099). JSA is a Conselho Nacional de Desenvolvimento Científico e Tecnológico researcher (303680/2022-9).