
Abstract
Chemokines critically orchestrate the tumorigenesis, metastasis, and stemness features of cancer cells that lead to poor outcomes. High plasma levels of transforming growth factor-β1 (TGFβ1) correlate with poor prognostic features in advanced lung cancer patients, thus suggesting the importance of TGFβ1 in the lung tumor microenvironment. However, the role of chemokines in TGFβ1-induced tumor stemness features remains unclear. Here, we clarify the previously undocumented role of CXCL1 in TGFβ1-induced lung cancer stemness features. CXCL1 and its receptor CXCR2 were significantly upregulated in TGFβ1-induced lung cancer stem cells (CSCs). CXCL1 silencing (shCXCL1) suppressed stemness gene expression, tumorsphere formation, colony formation, drug resistance, and in vivo tumorigenicity in TGFβ1-induced lung tumorspheres. Immunohistochemistry staining showed that patients with stage II/III lung cancer had higher expression levels of CXCL1. The levels of CXCL1 were positively associated with lymph node metastasis and correlated with the expression of the CSC transcription factor Oct-4. Furthermore, online database analysis revealed that CXCL1 expression was negatively correlated with lung cancer survival in patients. Patients with high TGFβ1/CXCL1/CD44 co-expression had a worse survival rate. We suggest that CXCL1 serves as a crucial factor in TGFβ1-induced stemness features of lung cancer.
Impact Statement
Chemokines are intricately involved in cancer malignant progression and the potential targets for therapy in recent years. A high plasma level of transforming growth factor-β1 (TGFβ1) was found in lung cancer patients and worsened the prognosis, while the role of chemokines in TGFβ1-induced stemness properties remains largely unknown. With A549 and EKVX-derived lung tumorspheres, we found that CXCL1 knockdown inhibited TGFβ1-promoted stemness gene expression, chemo drug resistance, and tumorigenicity. Therefore, the critical role of CXCL1 is demonstrated as a signal hub in TGFβ1-promoted lung tumor stemness features and further uncovered its clinical relevance. Our findings will provide a theoretical basis for the TGFβ1–CXCL1 axis as a therapeutic target or diagnostic marker for lung CSCs.
Introduction
Lung cancer is the leading cause of cancer death, and 85% of patients are diagnosed with non-small cell lung cancer (NSCLC).1,2 Most NSCLC patients develop metastatic disease and eventually die. Recent studies have demonstrated that cancer stem cells (CSCs) critically contribute to metastasis and tumor progression, 3 thus urging the identification of regulatory factors for CSC formation in NSCLC.
CSCs are a small subpopulation of tumor cells that harbor self-renewal capacity, tumor initiation, drug resistance, and metastatic potential. These characteristics can cause a relapse and failure of therapies.4,5 Chemokine receptors on cancer cells and CSCs enable navigation via corresponding chemokine gradients to the target organs, resulting in metastasis and tumorigenesis. 6 Thus, targeting chemokines and their receptors have potential in the development of therapeutic strategies. 7 Jung et al. 8 illustrated a connection between chemokine receptors and CSCs in lung cancer and found that upregulation of CXCR4 on CSCs is crucial for maintaining stemness and drug resistance in NSCLC. Currently, the CXCL12–CXCR4 axis is the best-understood chemokine–receptor interaction involved in cancer stem-like properties. Moreover, CXCR4 serves as a CSC marker for several types of cancer.9,10 Emerging evidence has highlighted the critical role and relevance of chemokines in cancer stem-like properties and CSC targeting, while molecular crosstalk still requires further illustration.
Cytokines within the tumor microenvironment intricately orchestrate the fate of CSCs, thereby necessitating the mechanistic illustration of component interactions. 11 The transforming growth factor-β1 (TGFβ1) is highly expressed and correlated with poor prognosis in lung cancer. TGFβ1 sculpts the tumor niche by modulating cytokines and consequently promotes angiogenesis, metastasis, drug resistance, immune escape, and cancer stem-like properties.12–15 The TGFβ/interleukin (IL)-6 paracrine loop between cancer-associated fibroblasts (CAFs) and cancer cells conferred epithelial-to-mesenchymal transition (EMT), chemoresistance, and stemness features in NSCLC. 16 Long-term exposure to TGFβ promoted invasion, chemoresistance, tumor formation, and recruited neutrophil infiltration via CXCL5 in hepatocellular carcinoma (HCC). 17 We also found that CXCR7 critically mediates the TGFβ1-promoted stemness properties and tumor initiation features of NSCLC. 18 However, research regarding the role of chemokine in TGFβ1-regulated lung cancer stem-like properties is comparatively rare.
In this study, we found that the expressions of CXCL1 and its receptor CXCR2 were significantly increased in TGFβ1-induced lung tumorspheres. A suppression of CXCL1 dramatically inhibited the expression of stemness-related genes, tumorsphere formation, and drug resistance induced by TGFβ1 treatment in CSCs. In addition, patients with high TGFβ1/CXCL1/CD44 co-expression had a worse survival rate. Findings from this study not only provide novel insight into the key role of CXCL1 in TGFβ1-promoted stem-like properties but may also lead to new approaches for the development of therapeutic strategies and molecular diagnosis for lung cancer.
Materials and methods
Cell culture
Human lung adenocarcinoma A549 (ATCC no. CCL-185) and H1299 (ATCC no. CRL-5803) cell lines were purchased from the Bioresource Collection and Research Center (BCRC, Hsinchu, Taiwan). Human lung cancer cell lines, including adenocarcinoma cells (EKVX, NCI-H522, NCI-H23), and large cell lung cancer cell lines (NCI-H460) were kindly provided by Dr. Sung-Liang Yu (National Taiwan University, Taiwan) and purchased from the National Cancer Institute (Bethesda, MD, USA). A549, EKVX, NCI-H522, and H23 cell lines were routinely maintained in a complete RPMI-1640 medium (Thermo Scientific Hyclone, South Logan, UT, USA) containing 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin (P-S; Gibco BRL, Life Technologies, Carlsbad, CA, USA). An H1299 cell line was grown in a DMEM (Thermo scientific Hyclone) medium supplemented with 10% FBS and 1% P-S.
Lentivirus-mediated knockdown of CXCL1
pCMV-ΔR8.91, pMD.G, and pLKO.1 plasmid with CXCL1 short hairpin RNA (shRNA) lentiviral knockdown vectors was packaged as per the procedures outlined by the National RNAi Core Facility (Academia Sinica, Taipei, Taiwan). The cells were infected with a shLuc or shCXCL1 lentivirus in the presence of 8 μg/mL of protamine sulfate (Merck Millipore, Billerica, MA, USA). Stable clones were selected using 2 μg/mL of puromycin (Sigma-Aldrich, St Louis, MO, USA).
Reverse transcription and quantitative polymerase chain reaction test
Total RNAs were extracted using a TRIzol reagent (Ambion, Life Technologies). RNAs were reverse transcribed in a 20-μL volume using SuperScript III First-Strand Synthesis System (Invitrogen, Carlsbad, CA, USA) as per the manufacturer’s protocol. The cDNA was subjected to quantitative real-time reverse-transcription polymerase chain reaction test (qRT-PCR) conducted in triplicate on a StepOnePlus™ Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) using the Fast SYBR Green Master Mix (Applied Biosystems). The results were averaged and normalized to the expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The primers used in quantitative polymerase chain reaction test (qPCR) are listed in Supplementary Table 2.
Spheroid formation assay
A single-cell suspension from trypsinization was plated at a low density in the tumor sphere medium comprising a serum-free DMEM/F12 (Gibco BRL, Life Technologies, Grand Island, NY, USA) medium, an N2 supplement or B27 supplement (Gibco BRL, Life Technologies), 20 ng/mL human recombinant basic fibroblast growth factor (FGF) (Gibco, BRL, Life Technologies), and 20 ng/mL epidermal growth factor (EGF) (Gibco BRL, Life Technologies) in the presence or absence of TGFβ1 (10 ng/mL). Fresh aliquots of TGFβ1 (10 ng/mL) were added every four days. After a four-week cultivation period, the primary spheres were gently trypsinized, counted, and re-seeded under the same culture conditions as the initial growth of the spheres. The numbers of any primary and secondary spheres larger than 100 μm were counted in each well.
Western blot
The A549 cells were cultured for two days and TGFβ1-treated A549 tumor spheres were generated as aforementioned. Total cellular proteins were extracted in the radioimmunoprecipitation assay (RIPA) Lysis buffer (Millipore, Billerica, MA, USA), supplemented with a protease inhibitor cocktail (Thermo Fisher Scientific). The cell lysates were resolved by a 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred to a polyvinylidene difluoride membrane (Millipore). The membrane was blotted with the indicated primary antibodies (CXCR2, ab89254; GAPDH, NB300-221) and then with horseradish peroxidase–conjugated secondary antibodies (Jackson ImmunoResearch, 115-035-003). The immunoblots were revealed using an ECL Kit (Millipore).
Flow cytometry
The cells were permeabilized and fixed using the BD Cytofix/Cytoperm solution (BD Biosciences, San Jose, CA, USA). After 20 min, the cells were centrifuged and washed with 1 × BD Perm/Wash buffer. The cells were then incubated on ice for 1 h with indicated primary antibodies, followed by fluorescein isothiocyanate (FITC)-conjugated secondary antibodies. After the final wash, the cells were suspended in phosphate-buffered saline (PBS) and analyzed using the FACS Calibur (BD Biosciences).
Anchorage-independent growth assay
The spheres (5 × 102) were suspended in a medium containing 0.33% Bacto-agar (Sigma-Aldrich) and layered on top of the base layer of 0.5% Bacto-agar. After 30 days, the colonies were fixed and stained with a Giemsa stain to examine the colony formation.
Cell viability
The spheres (5 × 103) were seeded in 96 well plates and incubated with the indicated concentrations of cisplatin (Sigma-Aldrich) or etoposide (Sigma-Aldrich). After 48 h, the MTS assay was conducted as per the manufacturer’s instructions. Absorbance at 490 nm was measured using a TECAN Sunrise enzyme-linked immunosorbent assay (ELISA) reader (Tecan Trading AG, Switzerland).
ELISA
The A549 and EKVX cells (2 × 105) were cultured in an RPMI medium for 48 h. The supernatant of each cell was collected and centrifuged for further detection. A 96-well plate was precoated with the CXCL1 capture antibody overnight and then blocked for 2 h for CXCL1 detection. Afterward, the cell supernatant was diluted and analyzed by Human CXCL1/GRO alpha Quantikine ELISA Kit (R&D Systems, Inc., Minneapolis, MN, USA) as per the manufacturer’s instructions.
Immunohistochemistry
Tissue microarray slides were commercially obtained from Biomax (US Biomax Inc., Rockville, MD, USA) and the company provided certified documents that all human tissues were collected with the informed consent of the donors. A waiver was granted by the institutional review board (IRB) of the Taipei City Hospital for the usage of human tissues for this study. The slides embedded in paraffin were deparaffinized through two changes of xylene and rehydrated through serial dilutions of alcohol. The slides were subjected to antigen retrieval by heating them in a 10-mM citrate buffer (pH 6.0) in a microwave for two 5-min sections, followed by blocking with 3% H2O2. The slides were then incubated with CXCL1 (R&D system, Minneapolis, MN, USA) or Oct-4 (GeneTex Inc., Hsinchu City, Taiwan) at 4°C overnight. The slides were washed and subsequently processed using a Dako EnVision™ + Dual Link System-HRP (DAB+) kit (Dako North America Inc., Carpinteria, CA, USA) as per the manufacturer’s instructions. The slides were incubated with an horseradish peroxidase (HRP)-labeled polymer for 40 min at room temperature and developed using a DAB, then counterstained with hematoxylin, and rinsed and mounted. They were then examined and scored using the digital pathology system Aperio ImageScope (Aperio Technologies Inc., Vista, CA, USA). Every tissue core was given a score as per the intensity of the staining (0, 1, 2, and 3 were designated as negative expression, weak expression, intermediate expression, and strong expression, respectively) and the extent of the stained cells (scored as 0, 1+, 2+, 3+, and 4+). The total score was determined by adding the intensity scores and the extent of the positivity scores. The clinicopathologic characteristics of individual tissues in the microarray are listed in Supplementary Table 1.
Xenograft model
NOD/SCID mice (NOD.CB17-Prkdcscid/IcrCrlBltw; 6–8 weeks of age, male) were purchased from BioLASCO (Taipei, Taiwan) and maintained at the Animal Center of National Yang Ming Chiao Tung University in specific pathogen-free conditions. The experiments were conducted as per the regulations of the Animal Care and Use Committee of National Yang Ming Chiao Tung University. The enriched secondary spheres were subcutaneously injected into NOD/SCID mice at doses of 103, 104, 105, and 106, respectively. The length and width of the tumors were measured with a caliper every week. The mice used in this experiment were randomly separated into each group.
Statistical analysis
Statistical analyses were performed using the SPSS software program (Version 22.0; SPSS Inc., Chicago, IL, USA). The data were expressed as mean ± SD. The statistical significance of the data was examined by the two-tailed Student’s t-test. The relationship between the protein expression and the lung cancer stage was assessed using the Kaplan–Meier method. The two-tailed χ2 test was conducted to identify the association between protein expression and clinicopathological characteristics. The Pearson correlation test was used to assess the correlation between CXCL1 and Oct-4 expression. A P value of <0.05 was considered statistically significant.
Results
CSCs express high levels of CXCL1
The accumulated evidence has identified the vital role of TGFβ1 in CSCs. A previous study by our group showed that a TGFβ1 treatment under a serum-free sphere formation medium promoted a tumorsphere formation of lung cancer cells. 18 Therefore, we utilized TGFβ1 to generate lung tumorspheres, which possess similar characteristics to CSCs. First, to investigate the difference in expression patterns of chemokines and chemokine receptors in CSCs, TGFβ1-induced tumorspheres were analyzed by qRT-PCR. We found that the expressions of CXCL1, CXCL12, and CXCR7 increased more than 20-fold in tumorspheres compared to the TGFβ1-untreated control group (Figure 1(A)). Furthermore, CXCR2, which is a receptor of CXCL1, was also upregulated approximately five-fold in the tumorspheres. Since the importance of the CXCL12–CXCR7 axis in CSCs has been clearly demonstrated, 18 we aimed to illustrate the role of CXCL1 in lung CSCs. To examine whether induction of CXCL1 by TGFβ1 was detectable in different types of lung tumorspheres, a panel of lung cancer cell lines was cultured with TGFβ1 under a serum-free sphere formation medium to generate tumorspheres. Except for some types of lung cancer cell lines that were unable to generate tumorspheres (data not shown), the spheres generated by different lung cancer cells expressed higher levels of CXCL1 (Figure 1(B)). To further validate whether CXCL1 was positively correlated with the stemness of tumorspheres, the expression of CXCL1 in primary and secondary spheres was analyzed using qRT-PCR and ELISA. As shown in Figure 1(C), the expression of CXCL1 was elevated in primary spheres when compared with parental A549 cells. Notably, we found a more dramatic induction of CXCL1 in the secondary spheres than in the primary spheres (Figure 1(C), left). We also detected an increased secretion of CXCL1 in the cell-conditioned medium through the serial passage of spheres (Figure 1(C), right). CXCR2 expression was also upregulated in TGFβ1-promoted tumorspheres (Figure 1(D)). These results revealed that a high CXCL1 expression may correlate with the high level of stemness in lung tumorspheres.
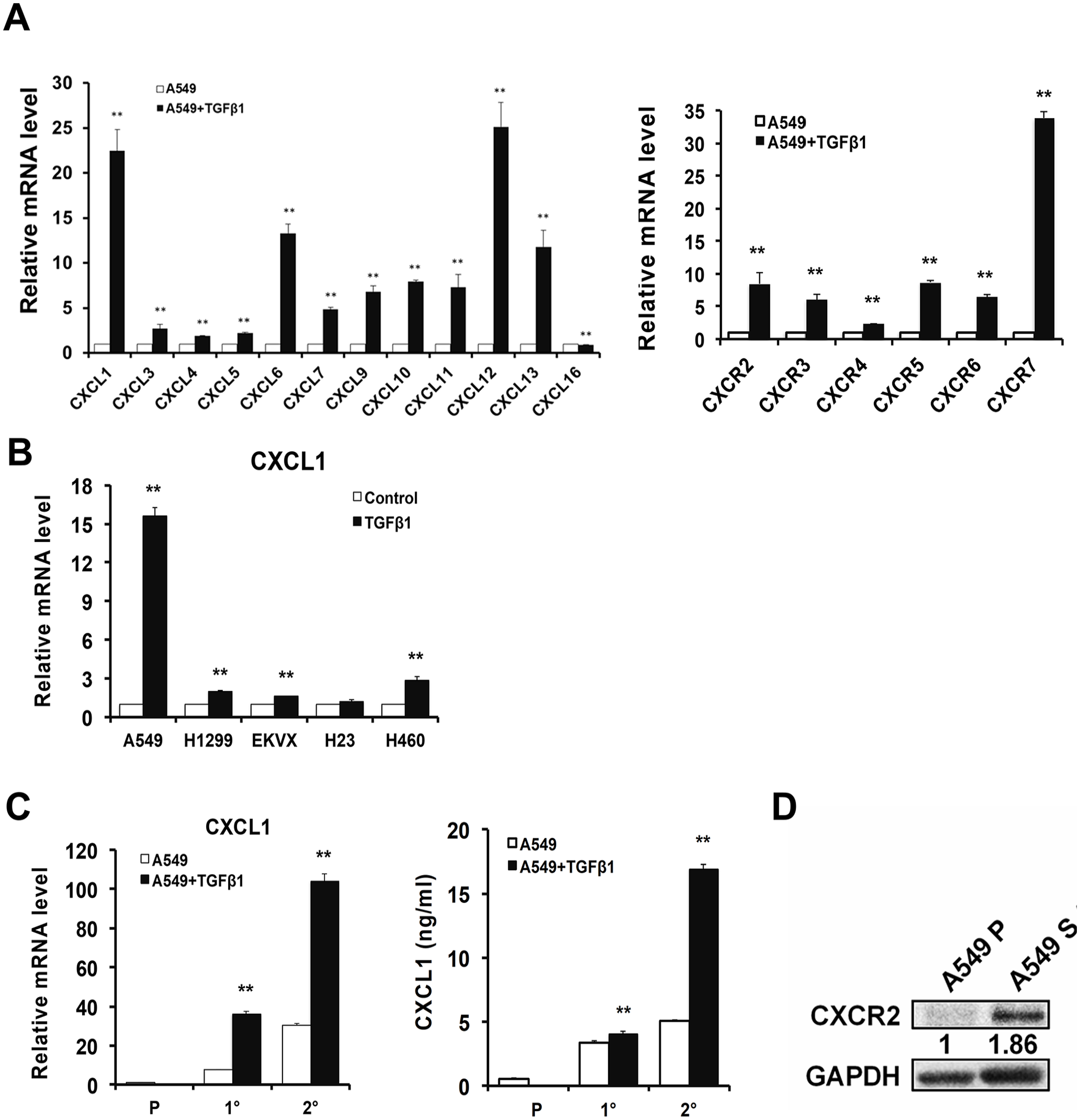
Lung CSCs express high levels of CXCL1. (A) and (B) A panel of lung cancer cells (2 × 104) was seeded in a six-well plate and cultured in a defined serum-free DMEM/F12 tumorsphere medium in the presence or the absence of TGFβ1 (10 ng/mL). After 28 days, the spheres were serially passaged to generate secondary spheres and analyzed by qRT-PCR. C (left) The A549 cells were cultured under a sphere medium with or without TGFβ1 (10 ng/mL). The CXCL1 expressions of the parental cells and the primary and secondary spheres were determined by qRT-PCR. (Right) The parental cells and the primary and secondary spheres (2 × 104) were seeded in a 6-cm plate for 48 h. The supernatant of the cells was collected and an ELISA was conducted. (D) The CXCR2 expression in the parental cells (A549 P) and the TGFβ1-treated tumor spheres (A549 S) were examined by the western blot.
CXCL1 downregulation inhibits the expression of stemness-related genes
To illustrate whether CXCL1 is associated with stemness features of lung cancer, we used a lentiviral shRNA system to knockdown CXCL1 in the A549 and EKVX cells. After a puromycin selection, the efficiencies of gene silencing were analyzed in both the mRNA and protein levels. As shown in Figure 2(A), the CXCL1 expression was significantly reduced in the shCXCL1-infected A549 and EKVX cells compared to the shLuc-infected controls. Next, the CXCL1 knockdown cells were cultured under a serum-free sphere formation medium in the presence or absence of TGFβ1 to generate tumorspheres. A qRT-PCR (Figure 2(B)) and flow cytometry (Figure 2(C)) analysis were conducted to evaluate the expressions of stemness-related genes, including Nanog, SOX-2, and Oct-4. We found that TGFβ1-induced Nanog, SOX-2, and Oct-4 expressions were markedly decreased when the CXCL1 was silenced (Figure 2(B) and 2(C)). These results indicate that the CXCL1 plays a role in the regulation of TGFβ1-induced stemness genes in lung cancer.
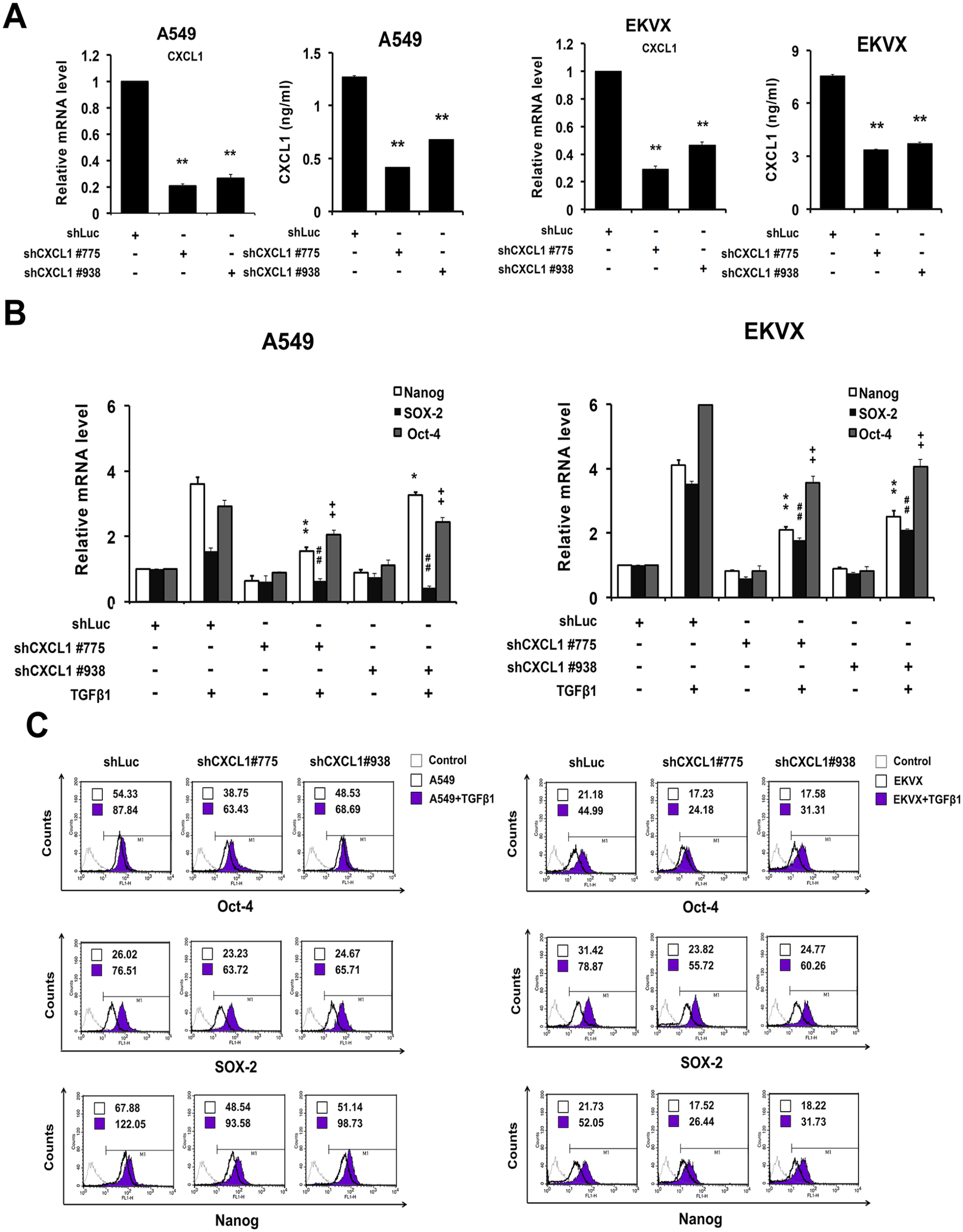
Suppression of CXCL1 reduced expressions of stemness genes. A lentiviral-mediated shRNA expression was used to knockdown the expression of CXCL1 in A549 and EKVX cells. The cells (2 × 105) were infected with the shCXCL1 lentivirus and selected with puromycin (2 μg/mL). (A) qRT-PCR and ELISA were conducted to validate the CXCL1 knockdown efficiency in the A549 and EKVX cells. (B) and (C) The CXCL1-knockdown cells were cultured with a defined sphere medium in the presence or absence of TGFβ1 (10 ng/mL). The expressions of the stemness-associated genes of the secondary spheres were analyzed by qRT-PCR (B) and flow cytometry (C). The Mean Fluorescence Intensity (MFI) was exhibited in the figures.
CXCL1 inhibition impairs stemness properties of lung CSCs
We next determined the effects of CXCL1 knockdown on self-renewal by evaluating the ability of sphere formation. Control cells or CXCL1-silenced cells were cultured in the sphere formation medium to form primary spheres and were propagated to generate secondary sphere cells. By adding TGFβ1 to the sphere formation medium, the number and size of the spheres were dramatically elevated. In addition, the TGFβ1-treated cells displayed more compact and round spheres compared with the non-treated group (Figure 3(A)). We observed a decreased number of spheres in the CXCL1-silenced A549 cells compared to the TGFβ1-treated shLuc cells (Figure 3(A), left). This result was consistent with the data from the EKVX cells (Figure 3(A), right). These data indicates that the inhibition of CXCL1 results in a lower frequency of lung cancer cells forming offspring spheres.
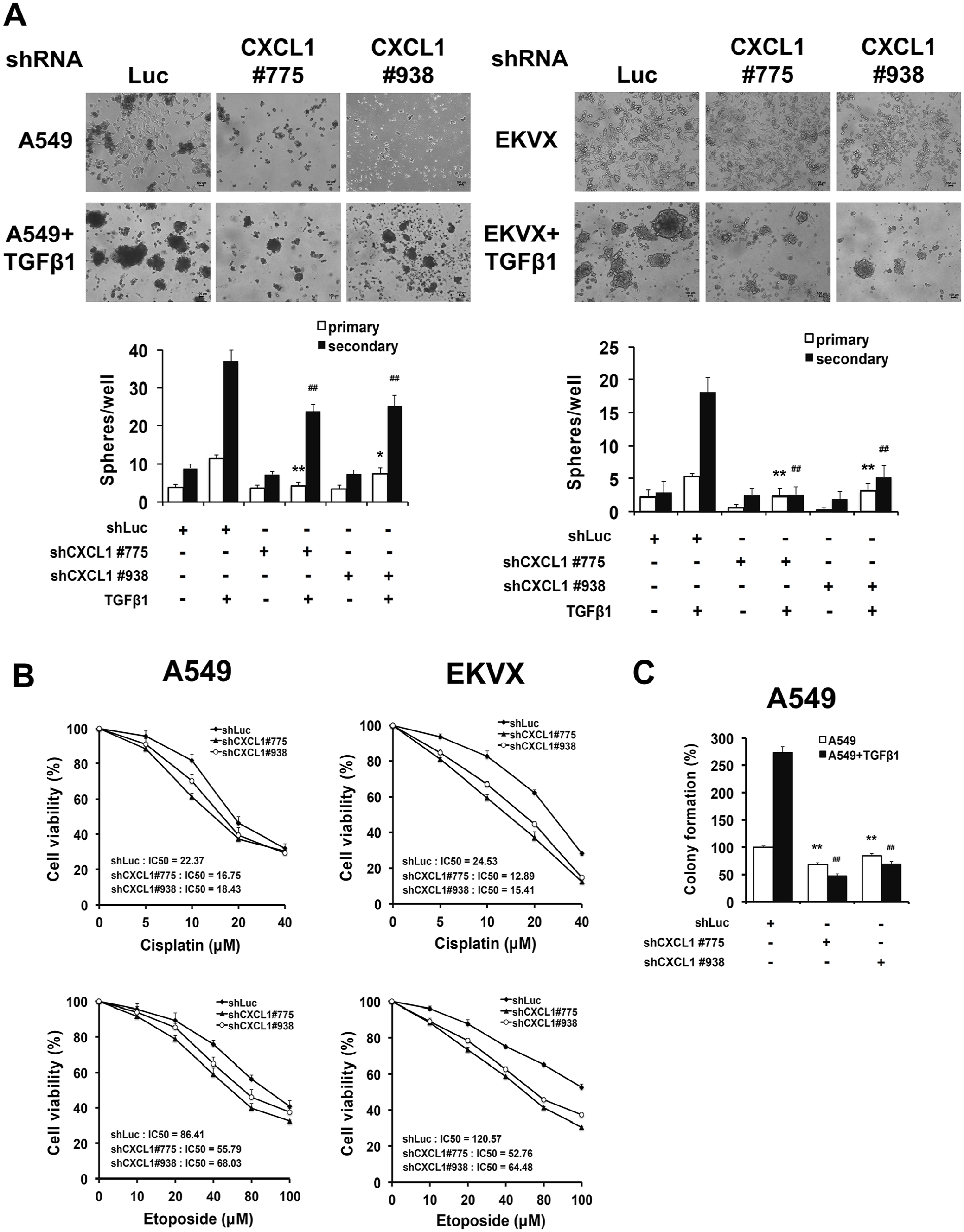
Inhibition of CXCL1 abrogated stemness properties of lung CSCs. The CXCL1 knockdown cells were grown under a defined sphere medium in the presence or absence of TGFβ1 (10 ng/mL). (A) The number of primary and secondary spheres larger than 100 μm were considered a sphere and calculated. (Top) Representative bright-field microscopy images of the spheres (magnification, ×40). (Bottom) Quantification of the spheres counted in a 96-well plate. (B) The secondary spheres (5 × 103) were seeded in a 96-well plate overnight and incubated with the indicated concentrations of cisplatin and etoposide for 48 h. Cell viability was then determined by an MTS assay. (C) The secondary spheres (5 × 102) were mixed with 0.33% agarose and layered as single cells on top of the base layer of 0.5% agarose. After 30 days, the cells were stained crystal violet and counted.
Since CSCs can escape the toxic effects of chemotherapy, we tested the chemoresistance of sphere cells by incubating them with cisplatin or etoposide. In the TGFβ1-treated spheres, CXCL1-silenced CSCs exhibited lower viability when compared with the control CSCs (Figure 3(B)). In A549 tumorspheres, the survival rate of shLuc control CSCs was 81.7%, whereas the two clones of the CXCL1 knockdown CSCs exhibited 61.0% and 70.1% survival in the presence of 10 μM cisplatin. Furthermore, etoposide susceptibility also increased in the CXCL1-knockdown CSCs (Figure 3(B), left). In addition, the half-maximal inhibitory concentration (IC50) of cisplatin for the two clones of the CXCL1-silenced EKVX CSCs (12.89 and 15.41 μM, respectively) demonstrated an approximately two-fold reduction compared to shLuc CSCs (24.53 μM). Similar results were obtained in etoposide treatment (Figure 3(B), right). To further clarify the effects of CXCL1 on the stemness properties of lung CSCs, we conducted a soft agar colony-forming assay of secondary spheres. The clonogenic potential of the spheres was markedly elevated in the TGFβ1-treated spheres. However, the increased colony number was significantly suppressed in CXCL1-silenced CSCs (Figure 3(C)). These observations in aggregate suggest that CXCL1 may be a critical modulator in regulating lung CSCs.
CXCL1 silencing reduces tumorigenicity in vivo
In vivo, tumorigenicity has been considered a critical cellular characteristic for evaluating the stemness of CSCs. CXCL1 knockdown cells and control cells were cultured and propagated into secondary tumorspheres followed by subcutaneous xenografts in NOD/SCID mice to evaluate the incidence of tumor growth. As shown in Figure 4(A), the knockdown of the CXCL1 completely abrogated the tumorigenicity of the CSCs when compared with the control CSCs. The tumorspheres cultured under the TGFβ1 treatment increased tumorigenicity when compared to untreated CSCs. Furthermore, the CXCL1 knockdown effectively decreased both tumor incidence (Figure 4(A)) and tumor size (Figure 4(B) and (C)) in the TGFβ1-induced sphere cells. These data are consistent with the findings that a CXCL1 knockdown inhibits stemness-related gene expression (Figure 2(B) and (C)) and sphere-forming capability (Figure 3(A)) in the presence of TGFβ1 and further validates the pivotal role of CXCL1 in regulating the tumorigenicity of lung cancer CSCs.
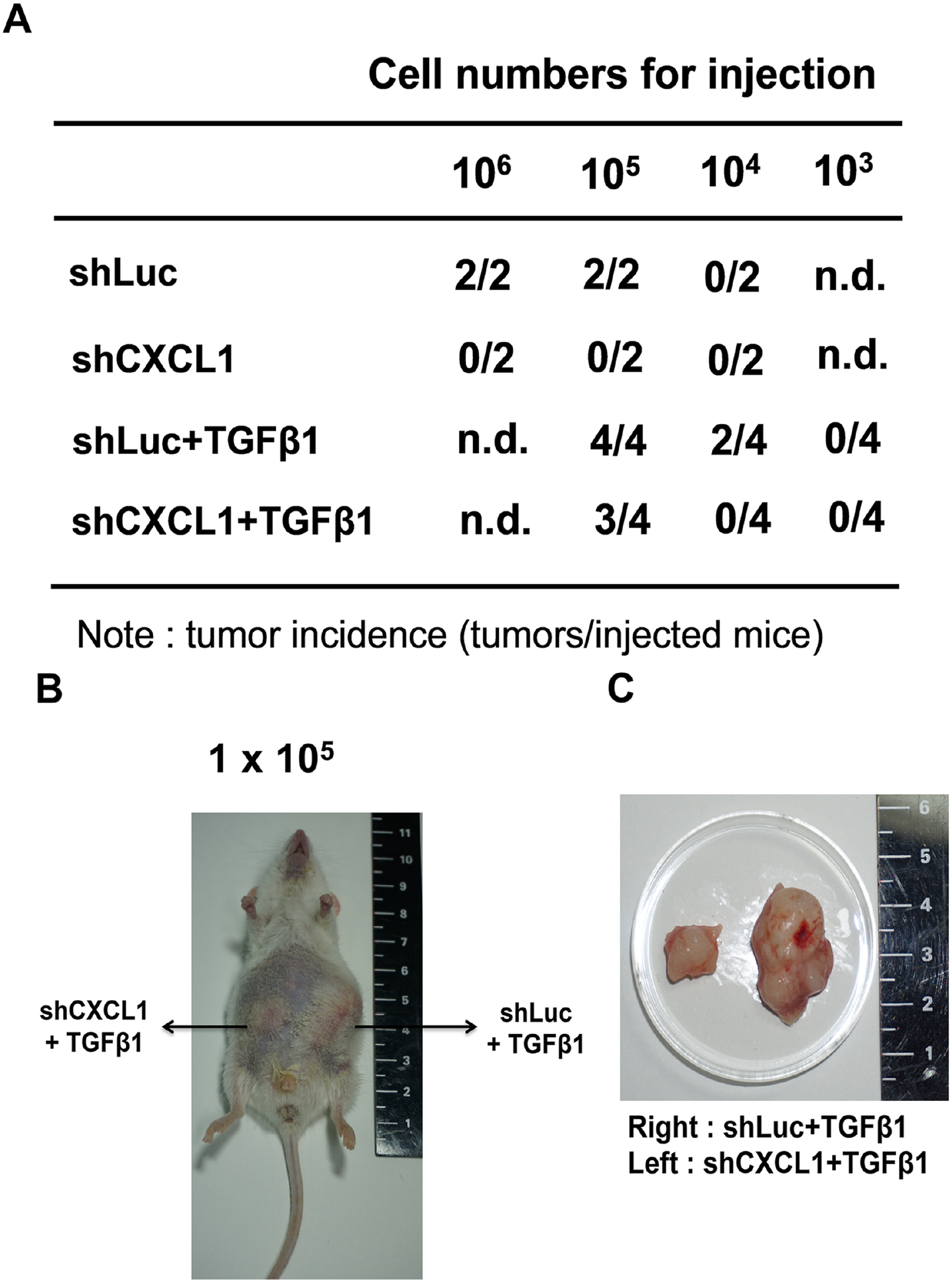
CXCL1 silencing reduces tumorigenesis in vivo. NOD/SCID mice were injected subcutaneously with a different number of secondary spheres. (A) The tumor-initiating potential of the shLuc, shCXCL1-infected spheres (treated with or without TGFβ1) was determined (n = 2 or n = 4 for each group). (B) and (C) The image of the tumors in NOD/SCID mice.
Elevated CXCL1 expression is positively correlated with advanced lung cancer
To investigate whether the CXCL1 expression was correlated with the progression of lung cancer, the CXCL1 expression was analyzed in the commercial tissue microarray using immunohistochemistry (IHC). As shown in Figure 5(A), the CXCL1 levels were significantly higher in the advanced stage (stage III) than in the early stage (stage I) of lung cancer tissues. Through further classification, we also found that the CXCL1 expression was positively associated with lung adenocarcinoma and squamous cell carcinoma stages (Figure 5(B) and (C)). Notably, lung cancer patients with lymph node metastasis expressed higher levels of CXCL1 (Figure 5(D)). These findings revealed that CXCL1 may play a vital role in lung cancer progression and metastasis. Our next aim was to clarify whether a CXCL1 expression was associated with CSC markers. The results showed that an Oct-4 expression was higher in stage III than in stage I of lung cancer and squamous cell carcinoma tissues (Figure 6(A) and (C)), while no significant difference in the Oct-4 levels was found in the different stages of the adenocarcinoma tissues (Figure 6(B)). Furthermore, a higher Oct-4 expression was observed in the tissues of metastatic lung cancer patients (Figure 6(D)). We also noticed that high levels of CXCL1 positively correlated with increased levels of Oct-4 (Figure 6(E)). To further identify the clinical relevance of a CXCL1 expression, we used the SurvExpress online database. The Kaplan–Meier analysis showed that the high co-expression of TGFβ1, CXCL1, and CD44, a well-known CSC marker, exhibited worse survival outcomes when compared to patients with a low expression (Figure 6(F)). A high CXCL1 expression was therefore positively associated with advanced lung cancer and poor outcomes.
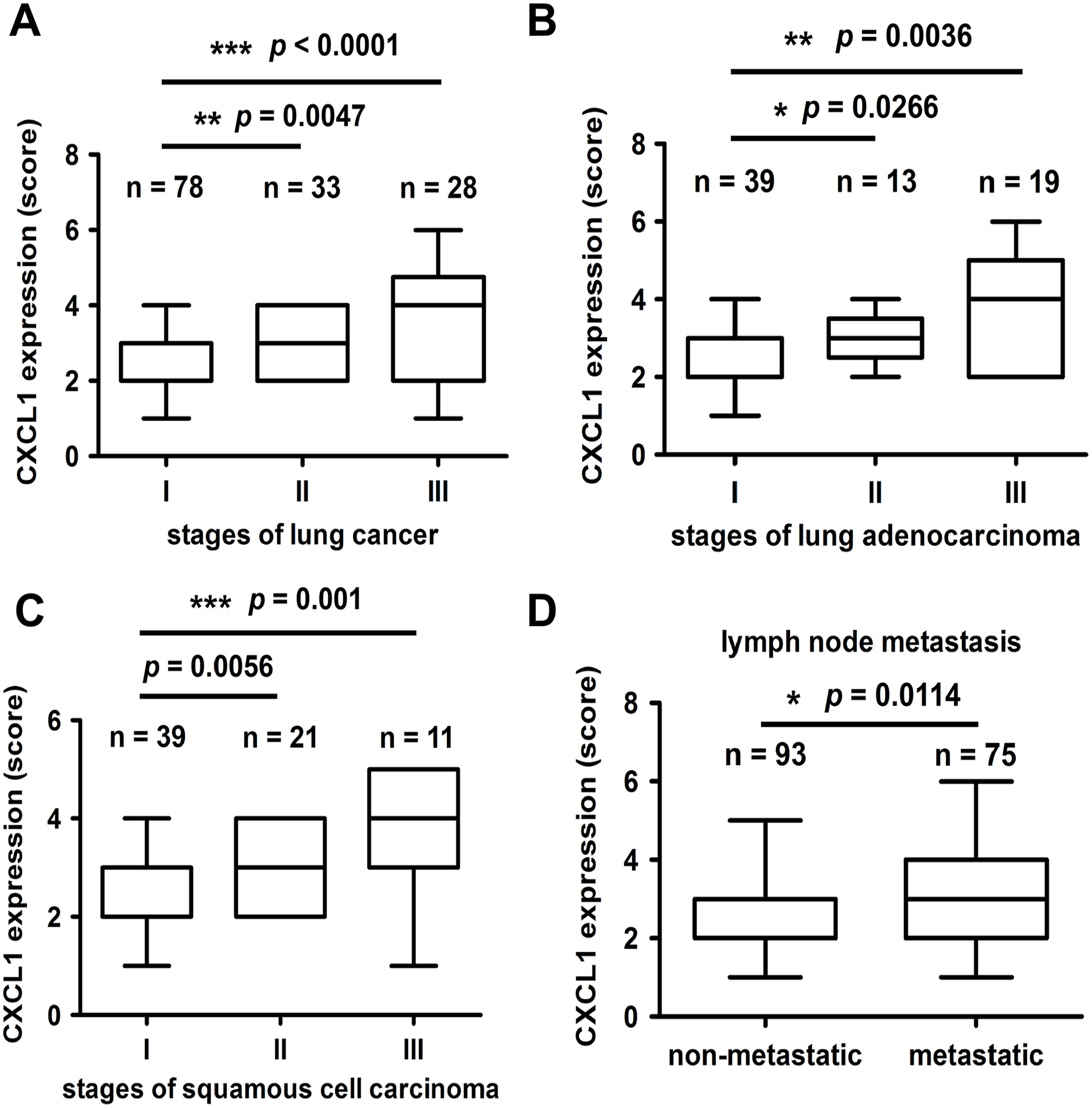
High CXCL1 expression is positively correlated with advanced lung cancer. Human lung cancer tissues were stained with IHC. (A) to (C) The expressions of CXCL1 in total lung cancer types (A) in lung adenocarcinoma (B) or in squamous cell carcinoma (C) were subdivided into clinical stages. (D) The comparison between the CXCL1 expression levels in the lymph node non-metastasis and metastasis groups.
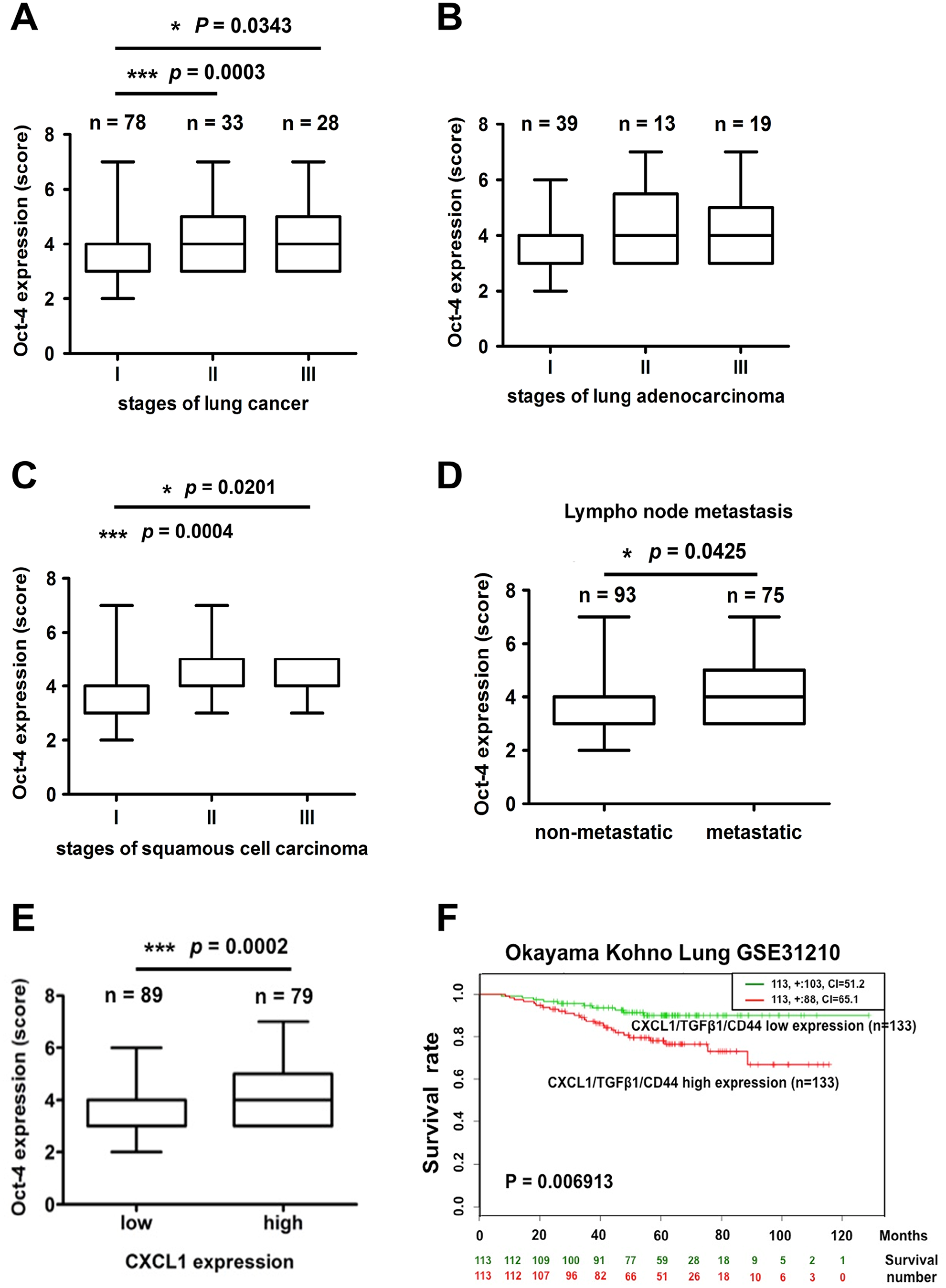
High Oct-4 expression is positively correlated with advanced lung cancer. Human lung cancer tissues were stained with IHC. (A) to (C) Expressions of Oct-4 in the total lung cancer types (A) in lung adenocarcinoma (B) or squamous cell carcinoma (C) and were subdivided in the clinical stages. (D) The comparison between the Oct-4 expression levels in the lymph node non-metastasis and metastasis groups. (E) The correlation analysis of the tissue content of CXCL1 and Oct-4 in lung tumor biopsies. (F) The Kaplan–Meier curves showing survival stratified by the CXCL1, TGFβ1, and CD44 co-expression using SurvExpress online software. The red and green curves are represented as high and low co-expression groups of patient death based on the Okayama Kohno Lung GSE31210 database.
Discussion
The TGFβ has been considered a double-edged sword in cancer progression. The TGFβ induces growth arrest and apoptosis in early progression but promotes EMT and immune escape in the later stage of malignancy. 12 Numerous studies have demonstrated the inhibitory effect of TGFβ signaling on CXCL1 expression in inflammatory diseases and cancers. Short-term exposure to the TGFβ may elicit a TGFβ-mediated immunosuppressive activity and downregulate the pro-inflammatory CXCL1 expression in the cells.19–21 Abrogation of TGFβ signaling (Tgfbr2 knockout) caused CXCL1 secretion from the tumor and stroma cells in breast and prostate cancer.22–24 Although the inhibitory regulation of the TGFβ signaling on the CXCL1 has been reported, this study is the first to illustrate that TGFβ1 promotes a CXCL1 expression in a long-term serum-free tumorsphere culture to enhance the stem-like properties of lung cancer.
CSCs are currently considered tumor-initiating cells (TICs), capable of driving tumor population expansion and relapse after treatment. However, the regulatory mechanisms and precise roles of a CXCL1 expression in TGF-β1-induced CSCs formation and stemness-related characteristics require further scrutinization. Tang et al. 25 first observed that CXCL1 was highly induced by the neurontensin/IL-8 axis via the MAPK pathway in CD133+ liver TICs. Ciummo et al. 26 found that the CXCL1–CXCR2 autocrine axis sustained a mammosphere formation in breast cancer. Furthermore, Agarwal et al. 27 reported that tumor necrosis factor (TNF)-α expanded CXCL1-secreted chronic myeloid leukemia (CML) stromal progenitor cells to enhance the proliferation and self-renewal of leukemic stem cells (LSCs) via CXCR2 signaling, and demonstrated the therapeutic potential of the CXCR2 inhibitor (SB225002) in depleting CML LSCs. The aforementioned studies consistently suggest the pivotal role of CXCL1 in CSCs, but without further characterizing the CXCL1 axis-regulated stemness properties. We found that Wnt/β-catenin and nuclear factor (NF)-κB pathways were activated in TGFβ1-promoted lung CSCs (data not shown). Besides, bioinformatic analysis displayed the existence of β-catenin, NF-κB, Oct4, and Sox2 binding sites in the CXCL1 promoter region, suggesting potential regulatory mechanisms of TGFβ1-induced CXCL1 in lung CSCs. By evaluating the stemness markers and performing functional assays in lung CSCs under TGFβ1 treatment, we provide a novel insight into CXCL1 in TGFβ1-induced lung cancer stem-like properties and suggest the clinical feasibility of CXCL1-targeting in lung CSCs abrogation.
CXCL1, known as growth-regulated oncogene-α (GROα) or melanoma growth-stimulating activity (MGSA-α), is a type of ELR+ chemokine. CXCL1 presents a 90% identical peptide sequence with CXCL2 and navigates circulating leukocytes to the inflammation site via CXCR2. 28 CXCR2 is expressed on neutrophils, monocytes, and mast cells and also regulates the self-renewal of hematopoietic stem cells and cancer progression.29–32 A CXCL1 overexpression has been observed in gastric, colon, liver, and bladder cancers and is critically associated with patients’ poor prognosis.33–36 One meta-analysis further demonstrated the positive correlation between CXCL1 and a shorter overall survival, advanced TNM stages, and lymph node metastasis in cancer. 37 The positive correlation between the TGF-β and CXCL1 serum levels was also associated with a poor prognosis, higher detection of circulating tumor cells (CTCs), and the propensity of lung metastases in breast cancer patients. 38 In our study, the IHC staining of the lung cancer tissue microarray showed a positive correlation between CXCL1 and advanced cancer stages (Figure 5). The high co-expression of the TGF-β/CXCL1/CD44 further leads to worse survival rates in lung cancer patients (Figure 6(F)). This result is consistent with reports that CXCL1 upregulation is an independent poor prognostic factor for cancer patient survival.
Several studies have revealed that CXCL1 signaling participates in tumor progression, including tumor growth, angiogenesis, chemo-/radio resistance, and metastasis.39–43 CXCL1 conferred malignant features by mediating cellular crosstalk among pro-tumoral cell lineages in the tumor niche, thus suggesting the potential of CXCL1 signaling-targeted therapy.44–47 The CXCL1 was induced by a vascular endothelial growth factor (VEGF) and a TNF from the cancer cells or tumor stroma in lung carcinoma.48,49 Yuan et al. 50 found that highly expressed CXCL1 in the serum of tumor-bearing mice directly promoted tumor growth and increased the infiltration of tumor-associated neutrophils (TANs), which consequently inhibited the T cell-mediated antitumor function. Tumor-associated macrophages (TAMs) have been elucidated for their immune suppressive and pro-tumoral role in cancer progression. 51 TAM-derived CXCL1 was discovered to have a pro-metastatic role in breast cancer through NF-κB/SOX4 signaling. 52 Miyake et al. further displayed the pro-tumoral role of CXCL1-mediated crosstalk within cancer cells, TAMs, and CAFs in bladder cancer. CXCL1 enhanced multicellular spheroid formation in the three-dimensional co-culture of cancer cells and TAMs/CAFs. 53 Recently, SenGupta et al. 54 found that triple-negative breast cancer (TNBC) spheroids recruited neutrophils by secreting TGFβ and GRO chemokines, including CXCL1. Monoclonal antibodies against CXCL1 suppressed tumor growth and angiogenesis via an IL-6 inhibition. 46 These findings reveal the pro-tumoral role of CXCL1 signaling in the tumor microenvironment and suggest the promising applicability of TGFβ1/CXCL1 for developing therapeutic strategies and molecular diagnosis.
To the best of our knowledge, our study is the first to scrutinize the undocumented role of CXCL1 in the TGFβ1-promoted stemness features of lung cancer and suggest that targeting the critical TGFβ1/CXCL1 signaling axis may offer a novel strategy to therapeutically eradicate lung CSCs.
Supplemental Material
sj-pdf-1-ebm-10.1177_15353702231220662 – Supplemental material for TGFβ1 induces CXCL1 to promote stemness features in lung cancer
Supplemental material, sj-pdf-1-ebm-10.1177_15353702231220662 for TGFβ1 induces CXCL1 to promote stemness features in lung cancer by Ta-Jung Peng, Yi-Ching Wu, Shye-Jye Tang, Guang-Huan Sun and Kuang-Hui Sun in Experimental Biology and Medicine
Footnotes
Authors’ Contributions
All the authors participated in the interpretation of the study, formal analysis of the data, and preparation of the manuscript. K-HS administrated, conceptualized, and supervised the project, acquired funding, conducted formal analysis, and reviewed and edited the manuscript; T-JP wrote the original draft, and conducted formal analysis and data visualization. Y-CW conducted the experimental investigation and validation, wrote the original draft, and conducted formal analysis and data visualization; S-JT conceptualized the project and conducted formal analysis. G-HS conceptualized the project, and reviewed and edited the manuscript.
Declaration Of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Ministry of Science and Technology, Taiwan (grant nos 109-2320-B-010-038, 110-2320-B-A49A-514-MY3); the Cancer Progression Research Center, National Yang Ming Chiao Tung University from The Featured Areas Research Center Program within the framework of the Higher Education Sprout Project by the Ministry of Education in Taiwan (grant nos 110CRC-T210, 111W31210); and the Tri-Service General Hospital and National Defense Medical Center, and Taipei City Hospital, Taiwan.
Ethical Approval
A waiver was granted by the IRB of Taipei City Hospital for the usage of human tissues for this study.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.