
Abstract
Live-bearing fishes (Poeciliidae) are increasingly used as laboratory models, but their utility is hampered by high rates of filial cannibalism, which reduces fry recovery and increases husbandry demands. Traditional strategies, such as mesh tank inserts (TIs), rely on fry actively seeking refuge and are often ineffective. Here, we present a novel tank design called Baby Catchers (BCs) that use water flow to passively and automatically separate newborn fry from their mothers, fitting seamlessly into standard recirculating rack systems. We tested the efficacy of BC tanks compared to open tanks (OT) and traditional mesh TIs using Poecilia parae, a live-bearing species that is notorious for intense filial cannibalism. In short-term trials, BC tanks yielded significantly higher initial fry counts on day 0 and day 2, while mesh inserts performed no better than OT. Over 10 days, negative binomial generalized linear models predicted substantial progressive fry loss in OT (∼60% decline) and inserts (∼68% decline), but no significant change in BC, consistent with high fry survival. These results provide the first quantitative evidence that automated water-flow separation of fry dramatically reduces filial cannibalism in live-bearing fishes. The BC design offers a scalable, low-cost, and humane method for fry recovery, reducing technician effort, minimizing animal stress, and lowering barriers for research and breeding programs involving live-bearing fishes (e.g., swordtails, mollies, Poeciliopsis, mosquitofish, halfbeaks, etc.).
Introduction
Naturally occurring differences across species have long been recognized to provide unique opportunities to study biological principles, yet species often come with logistical constraints to traditional lab protocols. In recent years many species of live-bearing fishes in the family Poeciliidae (e.g., guppies, mollies, and platies) have emerged as powerful models for studying diverse topics including sex determination, genome evolution, and plasticity.1–3 While their short generation times, small size, and hardy nature make them excellent research models, 4 the added complexity of live birth incurs significant costs to the research groups working with them since females will start to cannibalize their fry soon after birth. Such filial cannibalism is common in fishes,5,6 and has been recorded in nearly every group in the family Poeciliidae (e.g., swordtails, 5 mollies,7,8 guppies, 9 mosquitofish, 10 Poeciliopsis 11 ). This can dramatically reduce the number of offspring recovered, skewing breeding experiments and inhibiting or preventing the growth of stock populations. A high-throughput tank design that automatically separates fry from the mother after birth would simultaneously increase the number of fry recovered while decreasing the time needed to manually check for fry.
Currently, the most common method to decrease filial cannibalism when working with live-bearing fishes is to place mesh screen inserts in the tank with a gravid female. A variety of materials are used for this (e.g., wedding veil, plastic netting), which provide holes that are small enough for the fry to swim through but block access to the female. However, this method requires fry to recognize the area of safety, make it through the mesh, and stay safely in the area until researchers identify them and remove them from their mother’s tank. As a result, fry recovery is still likely to be affected by cannibalism. This method has two major downsides: (1) the number of fry recovered is a function of how frequently tanks are being manually checked throughout the day, and (2) the number of tanks required can grow rapidly since each cross requires one tank for the mother and one to place the isolated fry into. To overcome these issues, we developed a tank design that uses water flow to automatically isolate newborn fry from the mother and can be stacked in the footprint of a single traditional on-rack tank.
The species Poecilia parae (aka Micropoecilia parae) provides a prime system to test the efficacy of our “Baby Catcher” tanks. P. parae is a live-bearing fish in the family Poeciliidae, subgenus Micropoecilia, which is native to shallow brackish streams in northeast South America. 12 Despite having similar water parameter requirements to guppies (P. reticulata), maintaining long-term breeding stocks of P. parae has long been elusive to both the research and hobbyist communities. 13 In this species, gestation lasts ∼28 days, at which point free-swimming fry are born one at a time over 1–2 days. Our anecdotal observations in the lab suggest that cannibalism has played a major role in hampering the establishment of captive populations since P. parae are voracious filial cannibals.
We tested the efficacy of our new Baby Catcher (BC) tank design to increase the number of fry recovered in both the short term and long term, compared to open tanks (OT) and screen inserts, by monitoring how many fry could be counted by an observer each day. These tanks are easily constructed from acrylic sheets that can be cut either by a laser cutter or Computer Numerical Control (CNC) mill and fit on circulating rack systems with minimal modification or can even be used above a standard aquarium with a simple pump and minor plumbing. Additionally, after fry have been separated, they can remain in these modular tanks, which can be stacked to organize cohorts during early growth while requiring the footprint of only a single tank on the rack.
Methods
Stocks and husbandry
All P. parae individuals used to evaluate the efficacy of the new tank designs were from our laboratory stock populations, which were established from wild-caught individuals in 2022. Each of the stock tanks held 200 gallons and housed ∼100 individuals at a 50:50 sex ratio. All the stock and treatment tanks were plumbed together as one large recirculating rack system (∼1200 gal) that was custom-designed by Aquaneering; this ensured the same water parameters across all tank designs. The system was kept on a 12:12 h light:dark cycle, with water temperature between 25 and 27°C, pH of 7.5 and conductivity at 7500 µS (DI water and Instant Ocean Sea Salt). All fish were fed ad libitum twice a day on a mixed diet of live brine shrimp and pellets (Fancy Guppy, Kyorin Co., Himeji, Japan). All work was approved by the Institutional Animal Care and Use Committee of Cornell (IACUC protocol #2021-0067).
Treatment tank designs
The three tank designs being compared were “Open Tanks,” “Tank Inserts,” and “Baby Catchers”:
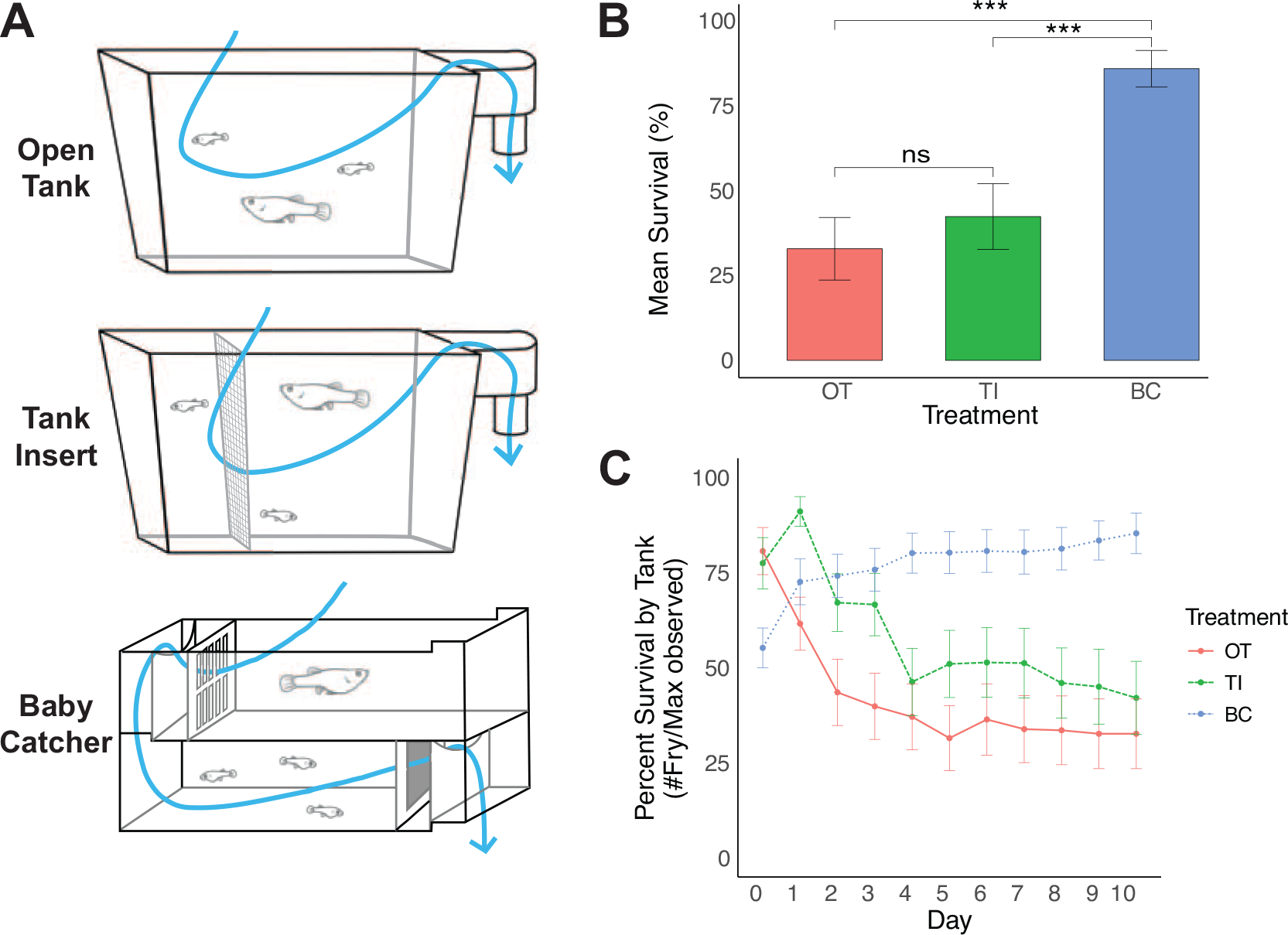
The key to isolating babies was the use of two alternative screens that slide into fixed notches in the chambers. The first insert was constructed by cutting 2.5 mm slits, which are large enough for newborn fry to pass through but too small for adult females to pass through, while the second insert was constructed by gluing 1010 micron polyester mesh (Component Supply, part U-CMY-1010) to a laser-cut acrylic frame. By placing an insert with slits in the upper chamber and an insert with mesh in the lower chamber, the water flow gently washes newborn fry through the insert and over the baffle to the lower chamber, where they remain since they are too big to pass through the 1010-micron mesh. The small volume of the tanks keeps the weight low, allowing the upper tank to be easily lifted off and the fry collected by net, turkey baster, or by gently pouring the lower tank into a new housing tank. This stacked flow-through design has the added benefit of only needing to feed the top chamber, since food flows through to the lower chamber. A supplies list, design files, and instructions for constructing these tanks have been included in the Supplementary Materials. The width of the slits that allow fry to wash to the lower chamber are wide enough for newborn fry of most Poeciliid species (2.5 mm). However, for species with especially large newborn fry (such as halfbeaks), the slit width can easily be widened either in the design files using a program such as Adobe Illustrator or by sanding the already cut piece.
Evaluating tank efficacy
Brood size in P. parae is highly variable, making it impossible to predict exactly how many fry are expected from any particular female. This makes it difficult to distinguish between small brood sizes and rapid filial cannibalism that occurs before observation. To overcome this issue, we used visibly gravid female P. parae from our laboratory stock populations and randomly assigned individuals to treatment groups. For each of the treatment groups we set up 30 individual females (N = 90 total). Tanks were checked daily until fry were observed. This was recorded as day 0, and daily recordings of the numbers of fry were taken until day 10. To prevent unnecessary stress, which could impact survival, fish were not removed from tanks until after the 10 days had concluded to verify final counts. If no fry were observed after 45 days, females were returned to stock tanks and considered to have 0 fry, as this was long past the 28-day gestation.
To confirm the females from different stock tanks were evenly distributed across treatments, we ran a Fisher’s exact test and found no differences (p = 0.99), indicating sufficient randomization. Fecundity is often associated with female size. To ensure that female size did not differ between tank treatments, we compared their standard lengths using a one-way analysis of variance (ANOVA) and found no difference between groups (F(2, 87) = 1.372, p = 0.26).
Designing the BC tanks to match the footprint of the 1.8 L tanks resulted in each chamber holding 1.1 L (Fig. 1A). This smaller volume would be expected to increase cannibalism rather than decrease it; therefore, we did not factor tank size into our final comparisons.
Statistical analysis
All statistical analyses were performed in R and plotted with ggplot2.14,15 We specifically used these data to ask two questions about the efficacy of the tank designs: (1) does BC increase initial fry survival? (2) does BC minimize fry death over time?
We defined “initial survival” as the survival of fry within the first 2 days after birth (day 0–2). This is the most valuable time frame to use when comparing to other methods as most caretakers would notice fry in tanks and remove them by this point. However, determining initial survival is challenging, as fry are not always born on the same day, and fry death from cannibalism can occur before they are even noticed in the tank. Thus, we gauged initial survival on days 0–2 as well as the maximum number of fry recorded in each treatment type to control for miscounts in tanks with numerous fry.
Numbers of fry between each group were compared. A Shapiro-Wilk test was conducted to assess the normality of the data from each treatment group on day 0. The test of OTs on day 0 revealed a W statistic of 0.7315, with n = 30, and a p value <0.001, indicating the data were not normally distributed. This was repeated for all treatments across days 0, 1, and 2 as well as the max fry counts over the entire 10 days, which confirmed a pattern of non-normality, consequently requiring the use of a non-parametric test. Thus, all the treatment groups were compared using a Kruskal-Wallis H test, as well as a post-hoc Dunn test to determine the significance when comparing specific groups in the short term.
For analysis over the long term, we observed the survival rates over a period of 10 days. This would remove the frequency at which fry need to be detected and transferred by technicians. This was done using a negative binomial generalized linear model (GLM) to determine the effect of treatment group on fry count and account for overdispersion (θ = 0.813).
Results
Initial survival
The maximum number of fry observed differed substantially between the different tank designs [H(2) = 25.45, p < 0.001], indicating that tank design had a major effect on filial cannibalism, with the BC tanks having a higher maximum number of fry observed than either the OTs (Z = 4.00, p < 0.001) or the tank inserts (TIs) (Z = 4.66, p < 0.001). To determine if the tank designs were affecting immediate filial cannibalism, we examined the number of fry detected on day 0 and found large differences between tank designs [H(2) = 15.813, p < 0.001], with substantially more fry detected in BC tanks than in either the OTs (Z = 2.90, p = 0.011) or the TIs (Z = 3.81, p < 0.001). Surprisingly, there was no difference between OTs and TIs in the number of fry on day 0 (Z = 0.908, p = 1.00), suggesting TIs do not help decrease immediate filial cannibalism. Since females were randomly assigned to tank designs, and they did not differ in size, differences between tank designs were unlikely to be due to differences in fecundity and were instead the effects of tank designs on rates of filial cannibalism.
Two-day survival
To determine if tank design was affecting the number of fry that could be removed within 2 days (a common interval for fry monitoring by technicians), we examined the number of fry detected on day 2 and again found large differences between tank designs [H(2) = 26.874, p < 0.001]. The BC tanks again had more fry than either the OTs (Z = 4.85, p < 0.001) or the TIs (Z = 4.01, p < 0.001). Again, there was no difference in the number of fry between the TIs and the OTs (Z = −0.848, p = 1.00).
Long-term survival
The negative binomial GLM indicated that fry counts declined at a rate of ∼8.8% per day in OTs (β = −0.092 ± 0.0242, p < 0.001) and ∼10.5% per day in the TIs (β = −0.0192 ± 0.0340, p < 0.001), consistent with progressive cannibalism. By day 10, the model predicts that only ∼39.7% of fry would remain in the OTs and only ∼32.8% of fry would remain in the TIs. In contrast, fry counts in the BC tanks showed no significant decrease over time (β = 0.128 ± 0.0314, p < 0.001), indicating fry survival was maintained in this treatment.
Discussion
In this study, we developed a novel technique for housing gravid female livebearers. Our results demonstrate that BC tanks substantially improve fry recovery by reducing both immediate and progressive filial cannibalism. In contrast, traditional insert-based designs performed no better than OTs in the short term and showed substantial fry loss over time. These findings provide the first quantitative evidence that automated separation using water flow is an effective husbandry tool for preventing filial cannibalism in live-bearing fishes.
Short-term
Use of BC tanks resulted in substantially more fry being observed on day 0 compared to OTs and TIs (Fig. 2). After confirming this difference could not be explained by differences in either the female size or stock tank of origin, the most likely explanation is that filial cannibalism occurs rapidly after birth. This interpretation is consistent with the number of tanks for which fry were never observed: 8 of 30 females in inserts, 8 of 30 in OTs, but only 1 of 30 in BC tanks. Given that all females appeared gravid and were housed beyond the length of gestation, the higher incidence of tanks with zero fry in the open and insert treatments likely reflects complete cannibalism prior to observation rather than reproductive failure. Interestingly, there was no significant difference between the OTs and inserts, highlighting that this common method is not effective at preventing immediate cannibalism (Fig. 2).
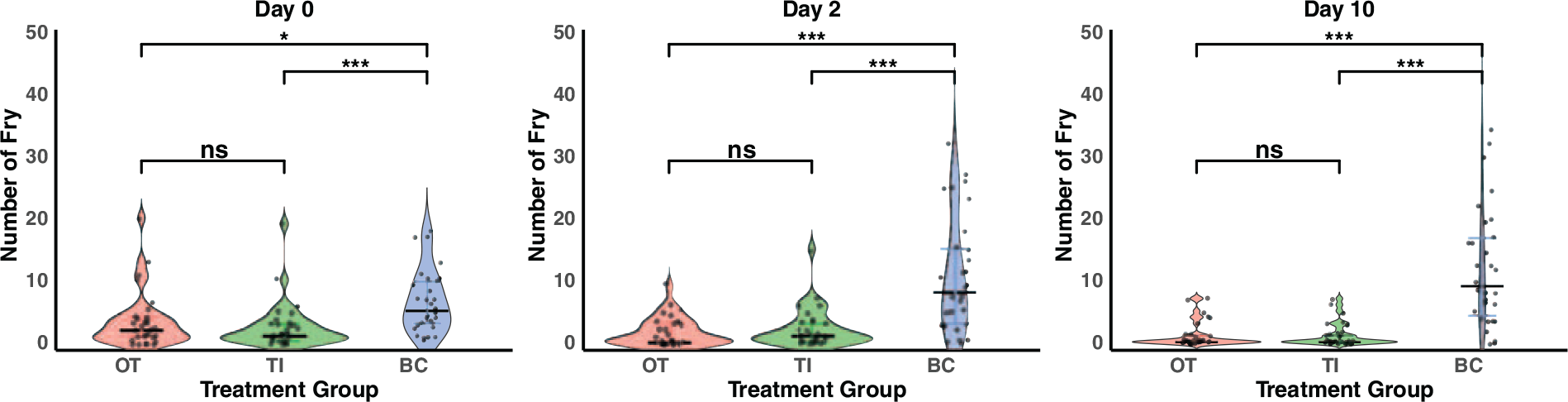
Number of fry observed by tank design at day 0, 2 and 10. Open tanks (OT), tank inserts (TI) and Baby Catchers (BC). The Baby Catcher design consistently had more fry than both the open tank and tank insert designs. The open tank and tank insert groups did not differ in fry number significantly on any of the days. (*p < 0.05, **p < 0.01, ***p < 0.001).
Long-term
The BC tanks also prevented progressive cannibalism substantially better than the other treatments. Negative binomial GLMs indicated fry counts declined at a rate of ∼8.8% per day in OTs and ∼10.5% per day in TIs. By day 10 the models predicted ∼60 and 68% decline in fry numbers observed in the OTs and TIs, respectively. In contrast, BC tanks showed no significant changes in fry counts over the same period, consistent with near-complete survival. While 10 days is likely longer than necessary for most husbandry applications, this result underscores the strength of the design in maintaining fry during periods of absence and ensuring separation without human intervention.
Why this work matters
Traditional insert-based methods rely on fry actively finding safe zones, which our results show is ineffective at preventing both immediate and progressive cannibalism. In contrast, BC tanks use passive water flow to automatically separate fry, eliminating reliance on fry behavior.
Both research and breeding facilities rely on the health and stability of their fry population; yet the manual transfer of fry into new tanks is often required to prevent cannibalism, which is time-intensive, stresses both mother and fry, and creates trade-offs: removing fry earlier improves survival but requires more frequent checks, whereas waiting reduces technician effort and stress but sacrifices fry recovery. Stress can negatively impact fish physiology as well as rates of reproduction, leading to a negative feedback loop of lower reproductive output. 16 When fry are being used for research, especially when specific crosses are required, the number of recoverable fry can determine feasibility; missing a brood may mean an additional month of waiting for the female to produce another brood. This problem can be even more serious for genetic studies, where random loss of even a small fraction of newborns can shift the interpretation of segregation frequencies.
The BC system provides a passive, scalable alternative that reduces effort and stress, integrates easily into rack systems, and requires minimal costs beyond construction. The reduction in immediate cannibalism also improves the accuracy of birth-date tracking, and fry in the lower tanks can be removed without disturbing the female. Beyond fry collection, these modular tanks can also serve for individual housing or grow-out tanks while taking a smaller footprint on rack systems. Where large recirculating racks are not available, they can also be used on top of an external tank with a small pump and minor plumbing.
While this initial proof-of-concept study was conducted on P. parae, we anticipate these tanks can benefit breeding efforts in nearly any small live-bearing fish, such as swordtails, mosquitofish, Poeciliopsis, and halfbeaks. By lowering the logistical and financial barriers to breeding livebearers, BC improves animal welfare and research efficiency. They offer a humane, cost-effective solution that minimizes technician effort, reduces stress on animals, and maximizes fry recovery. While future work should test performance across additional live-bearing species, BC tanks represent a significant advance in poeciliid husbandry.
Authors’ Contributions
Project design: B.A.S.; Tank design: B.A.S. and B.B.; Fry quantification: H.S., M.Z.-F., E.J.B., M.D.T., J.V., L.C., L.C.-C., and B.A.S.; Statistics: A.M.Z.; First draft: A.M.Z. and B.A.S.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Research reported in this publication was supported by the National Institute of General Medical Sciences of the National Institutes of Health under Award Number R35GM155164 to BAS. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. Support was also provided to JV and LC through the NEXUS Scholars Program at Cornell University.
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.