
Abstract
The EWSR1 (EWS RNA binding protein 1) was originally discovered in Ewing sarcoma, a pediatric bone cancer. In a previous study, we identified zebrafish ewsr1a, the homologue of human EWSR1. Using a zebrafish mutant for ewsr1a, we demonstrated that Ewsr1a promotes differentiation of chondrogenesis in Meckel’s cartilage, a component of the facial skeletal elements. Furthermore, we demonstrated that Ewsr1a interacts with the transcription factor SRY-BOX transcription factor 9 (SOX9) and modulates the transcription of its target genes. In this study, the role of Ewsr1a was further investigated in axial skeletogenesis. The maternal zygotic (MZ) ewsr1a/ewsr1a zebrafish display curved spines derived from irregularly spaced intervertebral discs (IVDs) and aberrant differentiation of nucleus pulposus cells. Using an in vitro cell culture system, we further show that the N-terminus of Ewsr1a is required for the interaction with Sox9. In zebrafish, the Ewsr1a also interacts with a Sox9 target gene, specifically the first intron of col2a1a. Zebrafish with the MZ ewsr1a/ewsr1a genotype display an increased level of collagen type II protein in the notochord starting at 36 h post fertilization. We propose that Ewsr1a contributes to IVD formation by regulating the expression of col2a1a.
Introduction
The EWSR1 (EWS RNA binding protein 1) was originally discovered as part of the EWSR1/FLI1 fusion gene, which results from chromosomal translocation in a pediatric bone cancer, Ewing sarcoma. 1 The major function of the EWSR1/FLI1 is known as an aberrant transcription factor, while EWSR1 itself is a multifunctional protein.2–4 EWSR1 regulates numerous biological activities. For example, it regulates transcription through interacting with RNA polymerase II and TFIID, facilitates DNA damage repair by inhibiting R-loop formation, and maintains centromere and RNA splicing.5–9 We previously reported that EWSR1 interacts with Aurora B kinase and regulates midzone formation during mitosis and prevents induction of aneuploidy.10–14 Using the zebrafish ewsr1a (a homologue of human EWSR1) mutant, we also demonstrated that Ewsr1a promotes chondrogenesis of Mekel’s cartilage, prevents the induction of aneuploidy, and prevents tumorigenesis in the presence of a wildtype tp53.13,15
In this study, we expanded our research to characterize the role of Ewsr1a in the axial skeleton using ewsr1a zebrafish mutants. The formation of the axial skeleton starts with the notochord, an axial midline structure derived from the mesoderm. 16 During this process, the notochord undergoes vacuolization and secretes extracellular matrix proteins (e.g., collagens and aggrecan) to form the notochord sheath, and this process is necessary for spine formation.16–23 It is known that human vertebrae are formed through endochondral ossification, whereas zebrafish vertebrae are developed through intramembranous ossification. In mammals, the notochord sheath forms both condensed and non-condensed areas that are evenly distributed throughout the axis. The condensed area gives rise to the intervertebral disc (IVD), which is composed of nucleus pulposus cells and annulus fibrosus cells.24,25 In contrast, the non-condensed area generates the cartilaginous primordia of the vertebral bodies (VB), which are derived from sclerotomal cells. 25 In teleost, vacuolation of notochordal cells generates hydrostatic pressure, allowing for the elongation of the axis. This region becomes distinct when the vacuoles undergo fragmentation in the subset of notochordal cells. Then, the segmentation of the notochord sheath, known as chordacentra, undergoes mineralization along the anterior-posterior (A-P) axis and will become IVD.22,23 The remaining areas containing vacuolated cells become VB. 26
Collagen Type II is a protein that is essential for forming fibrils in the notochord sheath, and it plays a significant role in the development of the axial skeleton. Col2a1 null mice displayed high numbers of notochordal cells and fewer nucleus pulposus cell in the IVD. 20 This observation highlights the significance of the expression of collagen type II at the notochord sheath, suggesting that the proper organization of the extracellular matrix at the notochord sheath is required for the differentiation of the nucleus pulposus cells. Previously, we discovered that the Ewsr1a interacts with the chondrocyte master regulator Sox9, and this interaction modulates the expression of Sox9 target genes, including Col2a1a mRNA. 15
Here, we demonstrate that Ewsr1a is required for the formation of IVD through regulating the expression of collagen type II. We show that the N-terminus of Ewsr1a is essential for its interaction with Sox9 in vitro, and it interacts with the first intron of col2a1a, which is a major target gene of Sox9 in zebrafish embryos. Our study uncovers a unique role of Ewsr1a in the development of the axial skeleton.
Materials and Methods
Zebrafish maintenance and ethics statement
The ewsr1a mutant line was obtained from Znomics, Inc. All embryos were maintained and staged as previously described.
27
Experiments were performed according to protocols approved by the
Alizarin red staining
Adult zebrafish were stained with Alizarin Red using a method described in the previous study. 28 The images were visualized using a Leica MZ FLIII dissecting microscope that was equipped with a Leica DFC320 camera.
Immunohistochemistry
Immunohistochemistry was performed on zebrafish embryos following the previously described method with minor modifications. 10 The following antibodies were used in this study: anti-collagen II primary antibody (Developmental Studies Hybridoma Bank at the University of Iowa, IA) (Invitrogen, diluted at 1:250), and anti-mouse Alexa 594 antibody (Invitrogen, dilution at 1:250). The images were documented with an Exi Aqua camera (Q Imaging) mounted on a Nikon Eclipse Ti microscope.
Co-immunoprecipitation
Co-immunoprecipitation (Co-IP) was performed as previously described with minor modifications. 12 HeLa cells were transfected with 14 ug of pSG5-Sox9 and pCS2-ewsr1a or pCS2-ewsr1a-C using Lipofectamine 2000 (LifeTechnologies, MA). Lysates were immunoprecipitated with either anti-mouse IgG or anti-FLAG antibodies (Agilent Technologies) conjugated to DYNA-protein G beads, followed by western blot using anti-Sox9 (AbCam, MA, diluted at 1/500). To generate an antibody against Ewsr1a, the C-terminus of ewsr1a (763–1878 bp) was cloned into the pET28a vector. The Ewr1a recombinant protein was purified from Escherichia coli (E. coli) and used as an antigen to generate a polyclonal antibody in rabbits (Pacific Immunology, CA).
Chromatin immunoprecipitation
Thirty-five wt/wt embryos at 27 hour post fertilization (hpf) were euthanized and subjected to a chromatin immunoprecipitation (ChIP) assay following a previously reported protocol. 29 The following PCR primers were designed for the col2a1a gene (4620–4865) and were used in this study.
col2a1a: F (5′-TTGCTTAGCACTGTACAGCC-3′)
col2a1a: R (5′-GTACCAAATCAGCATCATTCC-3′)
Results
Ewsr1a regulates the development of IVD in zebrafish
We previously reported that the maternal zygotic (MZ) ewsr1a/ewsr1a mutant, a line established by intercrossing zygotic ewsr1a/ewsr1a homozygous mutants, displays craniofacial defects in Meckel’s cartilage. 15 During the analysis, we discovered that the mutant also displays curved spines, prompting a study of the function of Ewsr1a during axial skeleton formation.
Alizarin red staining revealed that the neural arches were typically bent twice and projected dorsally in wt/wt zebrafish, while the trunk neural arches and spines were straight in the MZ ewsr1a/ewsr1a mutant zebrafish (Fig. 1A and B). One or more hemal spines were irregularly shaped with bifurcations or kinks in 75% (n = 20) of the MZ ewsr1a/ewsr1a zebrafish (Fig. 1B). In contrast, 9% of the wt/wt fish displayed either a bifurcated hemal spine or one bent hemal spine, resulting in a significant difference (n = 11, p = 0.0006). In addition, the MZ ewsr1a/ewsr1a mutants displayed curved spines (40%, n = 20). On the contrary, wt/wt fish did not display curved spines (0%, n = 11, p = 0.03) (Fig. 1A). In addition to the asymmetrical centrum of the caudal vertebrae and the curved spines, the IVD width was reduced in the MZ ewsr1a/ewsr1a (Fig. 1D and E). The average IVD width at the caudal vertebrae in the MZ ewsr1a/ewsr1a was 63 ± 16 mm (n = 19 fish), which was significantly less compared to 78 ± 22 mm in the wt/wt (n = 10 fish) (Fig. 1C and E). Note that between 6 and 15 IVDs were scored per fish, and the average score was presented as a single circle on the graph. The results suggest that the Ewsr1a plays an important role in IVD formation.
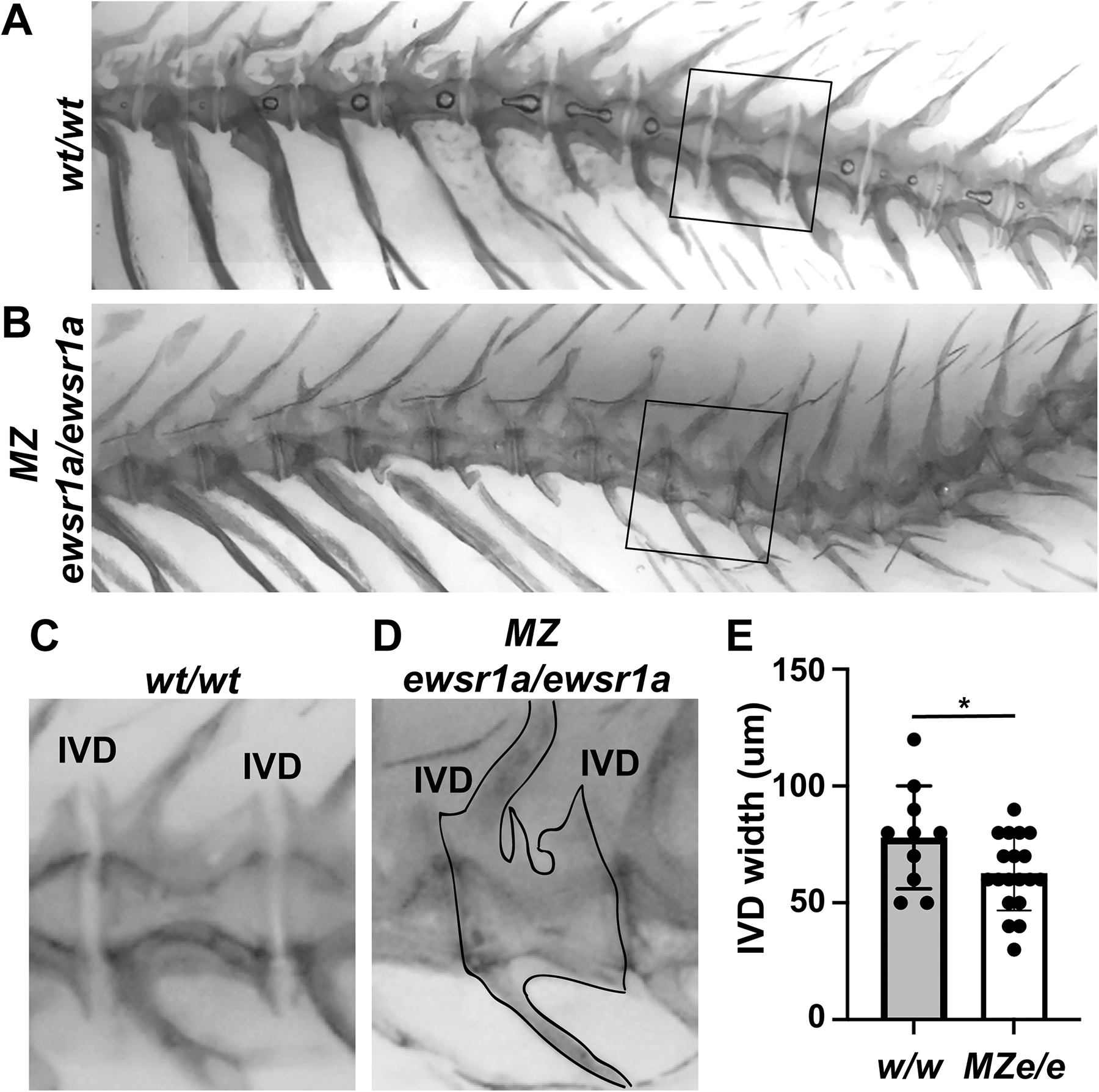
MZ ewsr1a/ewsr1a zebrafish have a reduced IVD width and curved spines.
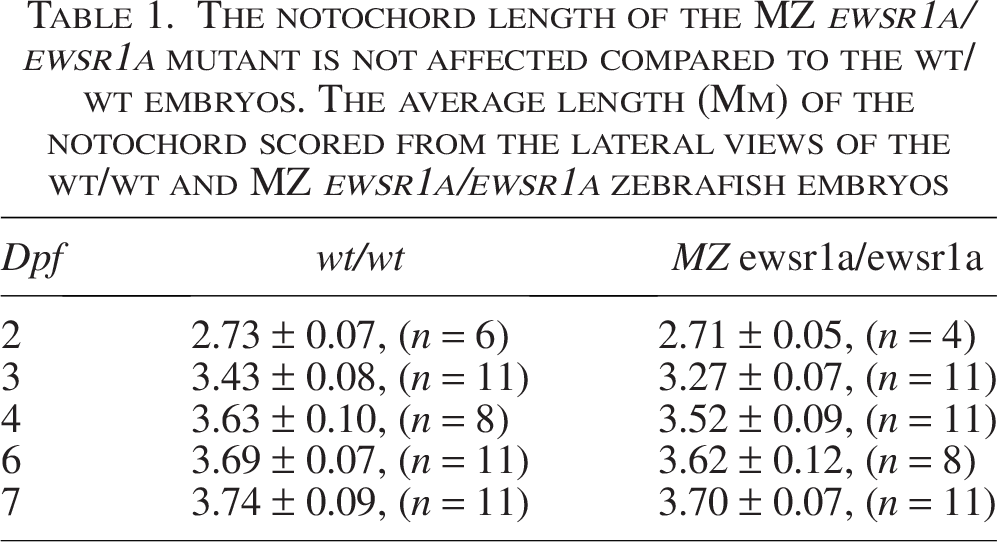
The notochordal cells differentiate into nucleus pulposus cells, which make up the IVD.30,31 This differentiation process is initiated by the formation of the notochord sheath, a layer of extracellular matrix proteins surrounding the notochord. 26 Since the notochord plays a crucial role in the development of IVD, we first investigated whether notochord specification was impacted. In situ hybridization using the notochord marker ntl as a probe revealed that the expression along the notochord was unaffected in the mutant at 27 hpf (wt/wt; n = 23, MZ ewsr1a/ewsr1a; n = 15) (p = 0.35) (Fig. 2A and B). In addition, the growth of the notochord of embryos at 2 and 7 dpf was not significantly affected in the mutant compared to the wt/wt zebrafish (Table 1). To visualize the cell morphology in the spines, we performed H&E staining on 5 dpf embryos, juvenile fish (1 cm total body length), and adult zebrafish. The morphology of the notochordal cells in the 5 dpf MZ ewsr1a/ewsr1a mutants (n = 6) was similar to that of the 5 dpf wt/wt zebrafish (n = 6) (p = 0.001) (Fig. 2C and D). The cell margins of the notochordal cells in the wt/wt zebrafish were clear and distinct, while some cells in the MZ ewsr1a/ewsr1a mutants had thicker and less clear cell margins (Fig. 2D, indicated with an asterisk). The spines of the wt/wt juvenile fish (1 cm total body length) displayed major reorganization, including in the IVD, with a partially calcified VB (Fig. 2E). In the wt/wt fish, the original notochordal cells with large vacuole morphology were no longer present, but instead, the nucleus pulposus cells with smaller cell morphology clustered in the IVD region (n = 3) (Fig. 2E). In contrast, the notochordal cells were retained in the MZ ewsr1a/ewsr1a mutants (100%, n = 3) (p = 0.05) (Fig. 2F). This result suggests that notochordal cells failed to differentiate to nucleus pulposus cells in the MZ ewsr1a/ewsr1a mutants. Furthermore, the adult IVD region in the adult MZ ewsr1a/ewsr1a fish (100%, n = 3) displayed an irregular and asymmetrical shape compared with the symmetrically shaped IVD in the wt/wt zebrafish (0%, n = 3) (p = 0.05) (Fig. 2G and H). Together, Ewsr1a is required for vertebral column formation by promoting the differentiation of notochordal cells into nucleus pulposus cells.
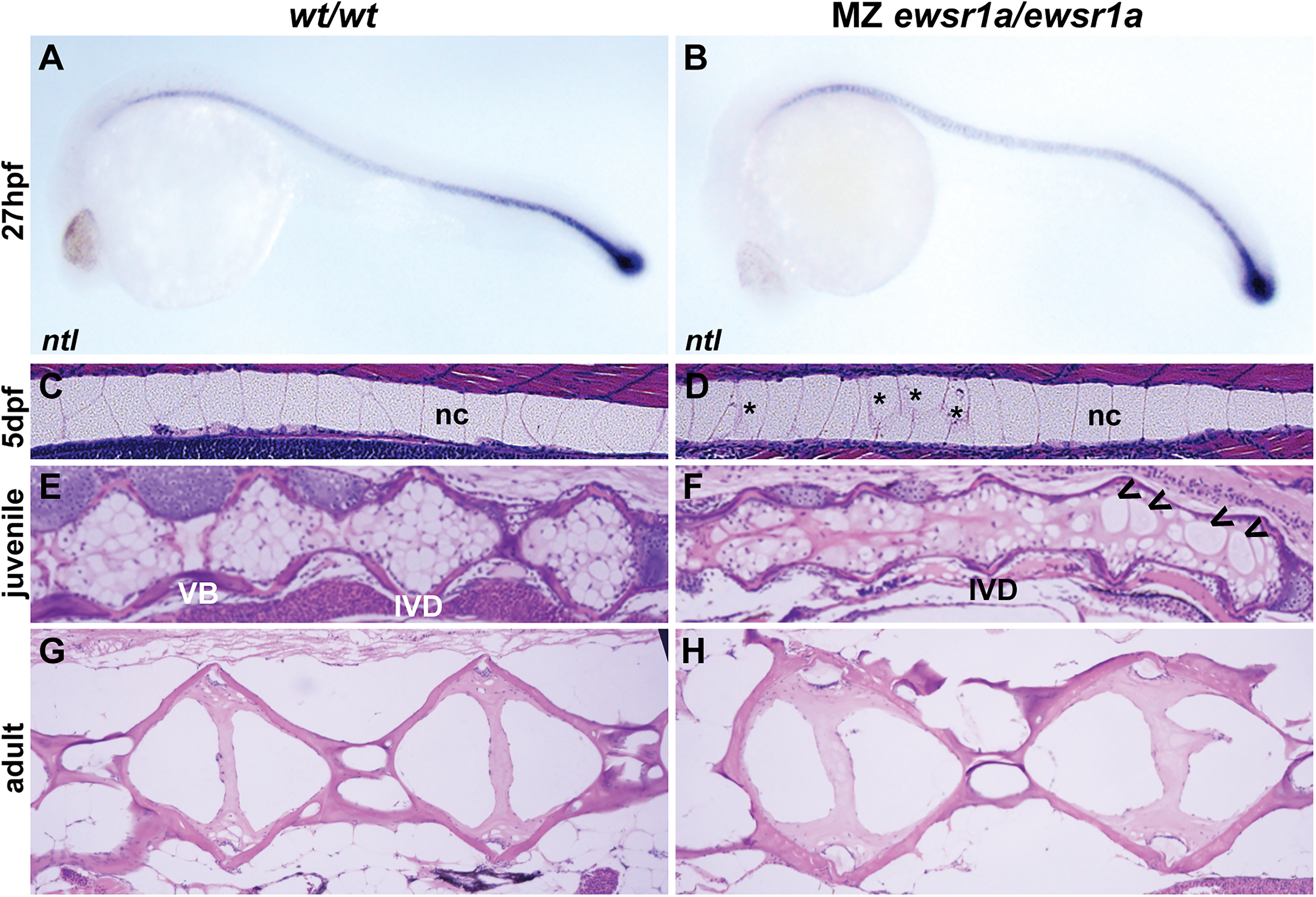
MZ ewsr1a/ewsr1a zebrafish display aberrant differentiation of nucleus pulposus cells.
The notochord length of the MZ ewsr1a/ewsr1a mutant is not affected compared to the wt/wt embryos. The average length (Mm) of the notochord scored from the lateral views of the wt/wt and MZ ewsr1a/ewsr1a zebrafish embryos
Ewsr1a regulates the expression of collagen type II
A previous study demonstrated that the expression of the collagen type II in the notochord is essential for the differentiation of nucleus pulposus cells.20,32 We previously showed that the MZ ewsr1a/ewsr1a mutant embryos (27 hpf) express higher levels of col2a1a and col2a1b mRNA compared to wt/wt using qPCR. 15 In the same study, we showed that the col2a1a and col2a1b mRNA are upregulated in the axis of the MZ ewsr1a/ewsr1a mutant (27 hpf) compared to wt/wt (27 hpf) using in situ hybridization. 15 Note that there were no signals in the axis of the 3 dpf MZ ewsr1a/ewsr1a and wt/wt embryos (data not shown). The study demonstrated that Ewsr1a regulates col2a by regulating Sox9, the master regulator of chondrocyte formation. 15 To determine if the expression level and localization of collagen type II protein are altered in this mutant, we conducted immunohistochemistry with an anti-collagen type II antibody. At 36 hpf, the MZ ewsr1a/ewsr1a mutant exhibited a higher intensity of collagen II signals in the notochord extension (anterior end of the notochord) compared to the wt/wt (73%, n = 30 vs. 0%, n = 16, respectively; p < 0.0001) (Supplementary Fig. S1A and B). The mutants also displayed a higher intensity and number of collagen type II signals (93%; n = 55) compared to the wt/wt (16%, n = 25) (p < 0.0001) (Supplementary Fig. S1C and D). It is worth noting that there was unexpected ectopic expression of collagen type II in the yolk extension of the mutant, for reasons unknown (Supplementary Fig. S1D). The data suggest that Ewsr1a may play a role in inhibiting the expression of collagen II in the notochord of normal embryos. Furthermore, the collagen type II expressing cells cluster with a regularly spaced vertical stripe pattern in wt/wt (9% displayed a disorganized pattern, n = 11), whereas this pattern is disorganized in MZ ewsr1a/ewsr1a mutants (100%, n = 8) at 3 dpf (p < 0.0001) (Fig. 3). The results suggest that Ewsr1a is required for maintaining a low expression level of collagen type II and for the formation of the notochord sheath in normal embryos. It is noteworthy that the correlation between the high expression of collagen II and its pattern disorganization in the MZ ewsr1a/ewsr1a mutants remains unknown.
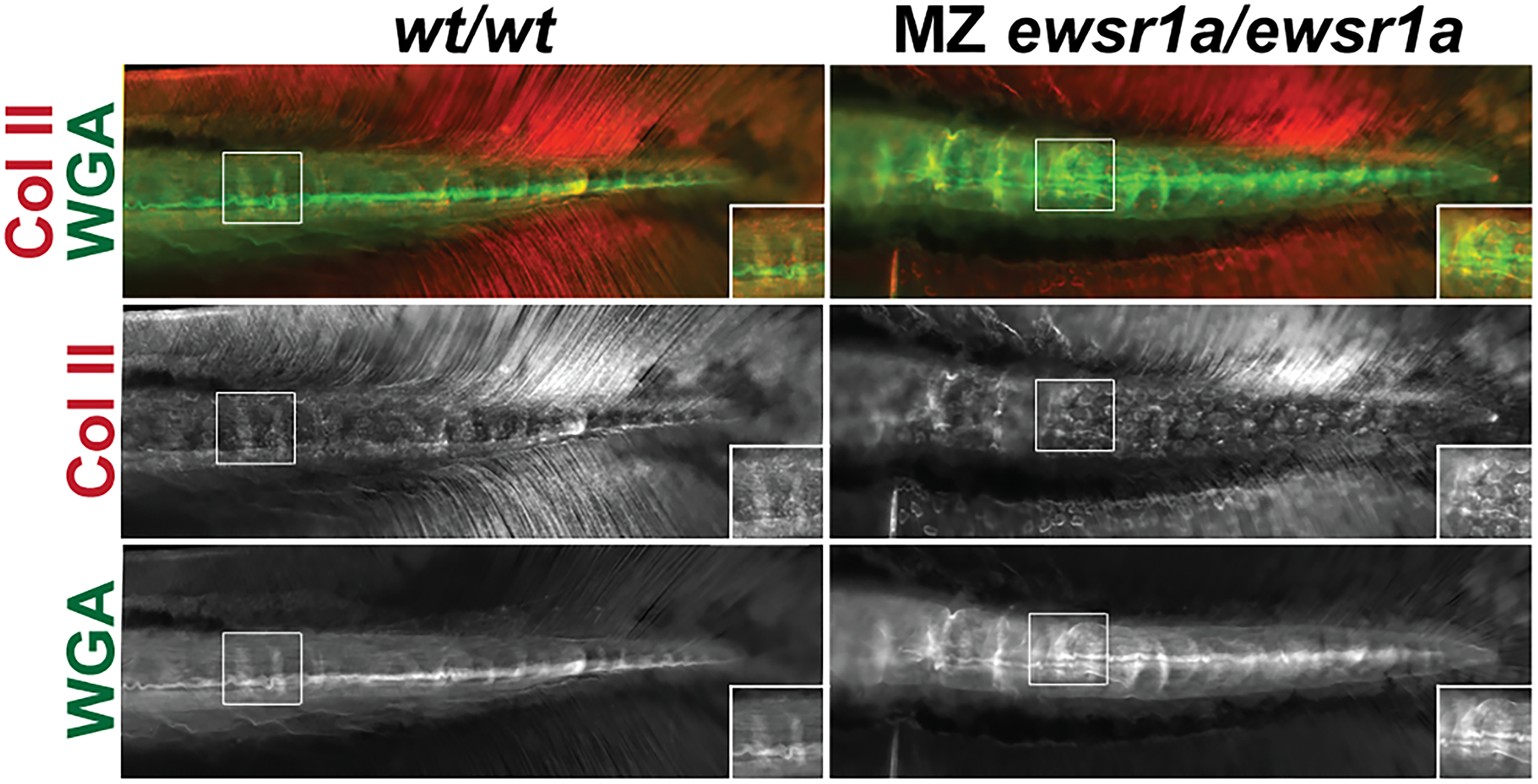
The MZ ewsr1a/ewsr1a mutants display increased expression of collagen II in the notochord. The trunk of wt/wt (left panel) and MZ ewsr1a/ewsr1a (right panel) 36 hpf zebrafish embryos at views (anterior to the left). All embryos were visualized with immunohistochemistry using an anti-collagen type II antibody (red), co-stained with wheat germ agglutinin (green). Col II, collagen type II; WGA, wheat germ agglutinin.
Because our previous study showed that Ewsr1a interacts with Sox9a, a co-IP experiment was performed to identify the essential domain of Ewsr1a required for interaction with Sox9a. A FLAG-tagged sox9a and either full-length or 3′-terminus of ewsr1a DNA constructs were co-transfected into HeLa cells, respectively (Fig. 4A). The lysates were immunoprecipitated using an anti-FLAG antibody and an anti-mouse IgG antibody as a negative control, followed by Western blot analysis using an anti-Ewsr1a antibody (Fig. 4A). Consistent with a previous report, the full-length Ewsr1a was co-precipitated with Sox9a. 15 However, there was a significantly reduced interaction between Sox9a and the deletion construct containing only the C-terminus of Ewsr1a. This result suggests that the N-terminus of Ewsr1a is the domain required for interaction with Sox9a (Fig. 4A). Our previous data showed that Ewsr1a inhibits the expression of col2a1a mRNA at 27 hpf embryos. 15 To further elucidate whether Ewsr1a interacts with the col2a1a gene, a ChIP assay was performed. From 27 hpf wt/wt zebrafish embryo lysates, Ewsr1a was immunoprecipitated, and the co-precipitated genomic DNA was amplified by PCR primers that were designed at the Sox9 consensus sequence of col2a1a located in the first intron (Fig. 4B). As a result, we discovered that Ewsr1a interacts with the first intron of the col2a1a gene (Fig. 4B). Together, Ewsr1a regulates the expression of col2a1a by interacting with the first intron of the col2a1a gene.
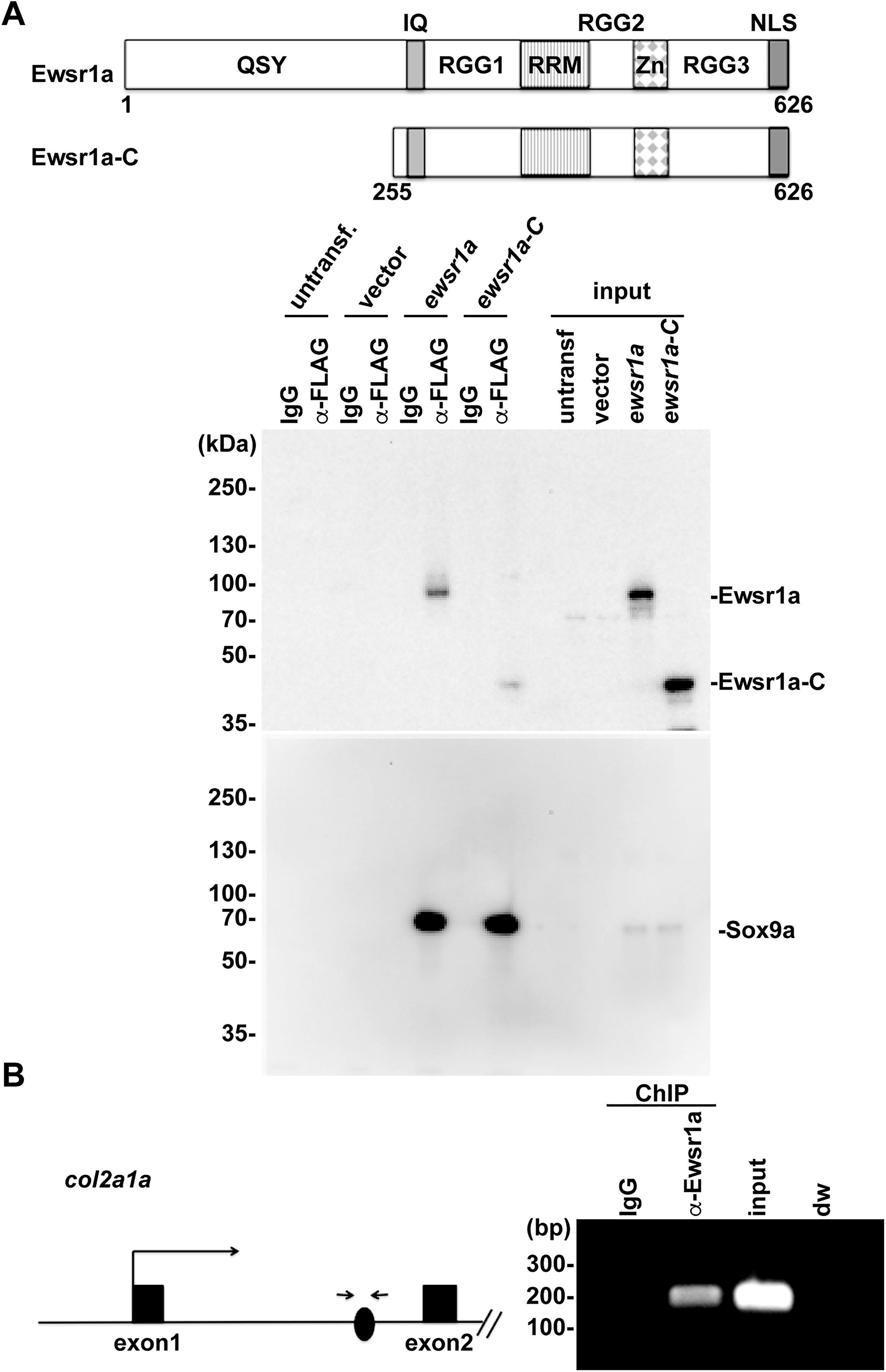
Ewsr1a interacts with col1a2a gene.
Discussion
This study aimed to characterize axis formation in the MZ ewsr1a/ewsr1a zebrafish mutant. We demonstrate that Ewsr1a regulates IVD formation, potentially by suppressing the expression of col2a1a during the differentiation from notochordal cells to nucleus pulposus cells. Because Ewsr1a regulates Sox 9 target genes through interaction, we further conducted a deletional analysis of Ewsr1a to identify the essential domain required for the interaction with Sox 9. We identified that the N-terminus of Ewsr1a is an essential domain required for the interaction with Sox 9 and showed that Ewsr1a regulates the first intron of the col2a1a gene.
Our results suggest that Ewsr1a plays a crucial role in the formation of the IVD by regulating the differentiation of notochordal cells into nucleus pulposus cells. Our previous study demonstrated that Ewsr1a interacts with Sox 9 and inhibits the expression of col2a1a during early embryogenesis. 15 Because collagen type II is crucial for the formation of collagen fibrils in the notochord sheath, the vertebral body, located outside the notochord sheath, exerts a mechanical force that contributes to the elimination of the notochordal cells. 20 Another possibility is that the collagen fibril creates a scaffold that allows the clustering of as-yet unknown signaling molecules, triggering notochordal cell elimination and promoting nucleus pulposus cell differentiation. The identification of this signal will be critical for a better understanding of the differentiation of the nucleus pulposus. It is noteworthy that polycystin1a (pkd1)a/b- and pkd2-deficient zebrafish embryos induce overexpression of collagen type II, leading to a curved axis. Therefore, the study supports the importance of the expression level of collagen type II. 33
In addition to inhibiting the differentiation of the nucleus pulposus in MZ ewsr1a/ewsr1a mutant, our previous study also demonstrated that the chondrogenesis of endochondral ossification in Mackel’s cartilage was delayed in the mutants. It has been suggested that both phenotypes stem from compromised Sox 9 activities, as these phenotypes resemble in Sox 9 knockdown mice.34,35 In this study, we discovered that the N-terminus of Ewsr1a is essential for its interaction with Sox9. The N-terminus of Ewsr1a contains a prion-like domain (PrLD).36,37 This domain has self-assemble activity, and it leads to phase separation. Therefore, it is possible that the self-assembled Ewsr1a may prevent Sox9 from accessing its target locus due to phase-separated compartments. Because differentiation of all cell types is a dynamic process, Ewsr1a may serve as a temporal hub through its self-assembly mechanism, and this complex may inhibit the activity of Sox 9. To test this concept, it is essential to further generate mutation constructsfor PrLD of Ewsr1 and study its activities of self-assembly, the expression of the col2a1a gene, and the differentiation of notochordal cells into nucleus pulposus cells. Spinal deformity induced during developmental stages (e.g., congenital kyphosis, congenital scoliosis, and congenital lordosis) often compromises quality of life due to fatigue, stiffness of muscle, neurological defects, and cerebrospinal fluid leakage.38,39 Given the phenotypes displayed in the MZ ewsr1a/ewsr1a zebrafish mutant, it is possible that the Ewsr1a (or human EWSR1) may play a critical role in spinal development. Here, we report a unique role of EWSR1 in axial development, and this zebrafish model should provide a platform for further dissecting the pathway.
Footnotes
Acknowledgments
Research reported in this publication was supported by the University of Kansas Graduate Research Fund (GRF).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.