
Abstract
Objective:
The skin barrier function is essential for maintaining skin health. Consequently, it is crucial to evaluate the protective and restorative effects of topical medications and cosmetics on the skin barrier. However, methods that can directly assess the skin barrier function in zebrafish beyond their regenerative capabilities are limited. Therefore, we aimed to develop an in vivo, in situ, high-throughput screening model for assessing skin barrier function in zebrafish embryos.
Methods:
Lactic acid was used to induce skin barrier damage and crystal violet was used for staining to observe skin barrier damage. Histopathology was assessed using H&E, Masson, and Victoria blue staining. Cellular apoptosis was evaluated using in situ acridine orange (AO) staining and flow cytometry with propidium iodide (PI) staining.
Results:
A significant increase was noted in fluorescence in the trunk and tail regions of zebrafish embryos following lactic acid stimulation compared with those in the control group, whereas exposure to the positive control for 4 h significantly reduced the fluorescence area, showing 35%–45% inhibition. Histological sections revealed that lactic acid caused noticeable damage to the epidermis of zebrafish embryos, characterized by thinning of the epidermal layer and the presence of inflammatory cell infiltrates. An increase in apoptotic cells was observed through in situ AO and PI staining using flow cytometry, and RT-qPCR revealed elevated expression levels of apoptosis-related genes, including fas, bax, and caspase 3. Furthermore, six additional samples from various sources were screened using this model, which yielded reasonable outcomes for both positive and negative samples, indicating its potential application value in evaluating the skin barrier protective efficacy of drugs or compounds.
Conclusion:
Our study supports the use of the lactic-acid-induced zebrafish skin injury model as a reliable in vivo and in situ method for evaluating skin barrier function.
Introduction
The skin serves as a vital immune organ. It’s not only a physical barrier and chemical barrier, but also the body’s first line of defense. The basic function of the skin barrier is to regulate the exchange of substances between the body and environment in an orderly manner, thereby maintaining hydration, preventing and eliminating microbial infections, reducing damage from ultraviolet exposure, and mitigating oxidative stress damage. 1 A decline in skin barrier function is associated with the occurrence and progression of various skin diseases, including infections, inflammation, allergies, and skin cancer. For example, atopic dermatitis (AD) is a multifactorial disease associated with barrier disruption and systemic inflammation.2–4
Given the importance of skin barrier function, many studies have suggested that protecting or repairing the integrity of the skin barrier can help improve or alleviate various skin diseases, including rosacea and AD. This strategy is beneficial to both healthy and compromised skin. Therefore, many topical medications and skincare products have been designed to maintain the integrity of the skin barrier.5–7 Ceramides, hydroxypropyl tetrahydropyrantriol (Proxylne), (S)-2-methyl-1,4,5,6-tetrahydropyrimidine-4-carboxylic acid (Ectoine), and some natural product extracts can enhance the function of the skin barrier.7–12 Therefore, assessing skin barrier function is crucial for these studies. Although clinical skin examinations offer valuable insights, quantitative models of skin barrier function are essential for assessing the efficacy of medications and skincare products in repairing the skin barrier.13–16 In clinical testing, transepidermal water loss (TEWL) is commonly used as an indicator of barrier function and some studies use skin sensitivity to lactate stimulation as a basis for evaluation.8,13,17–20 However, human testing has high ethical and cost requirements. Therefore, laboratory methods are required to supplement these tests. In vitro cell models, including keratinocytes, fibroblasts, and mixed cell co-culture models, are frequently used in barrier function research.21–25 The skin barrier is composed of multiple factors, therefore, a single-cell model cannot fully represent the overall state. It is necessary to explore a model that can directly evaluate skin barrier function to meet current research needs.
Zebrafish have been used as model organisms for drug screening and skin science research. Compared with traditional mammalian models, zebrafish have high fertility and rapid development, and can be used for high-throughput screening.26–31 Although lacking a stratum corneum, zebrafish have intact epidermal and dermal layers, and their skin barrier functions are similar to those of mammals. Currently, there are methods that use the zebrafish caudal fin regeneration model to assess skin regeneration capacity.32–35 Although skin regeneration capacity is related to skin barrier function and provides some insight into the self-repair mechanisms of the skin, it does not fully encompass the complete range of the skin barrier roles and functions. Therefore, there is a need for methods that can directly assess the skin barrier function in zebrafish beyond their regenerative capabilities. However, the research throughput on mammalian barrier function remains low; further, to the best of our knowledge, a fully mature model for studying zebrafish skin barrier function is yet to be established.
In this study, we aimed to establish an in vivo, in situ method for evaluating skin barrier function using zebrafish, which involves damage to the skin of zebrafish embryos with lactic acid and crystal violet as a tracer to evaluate the functional changes in the skin barrier. We also examined the pathological changes and levels of cellular apoptosis in the skin of zebrafish embryos after lactic acid injury. Finally, we used this method to screen for skin barrier protective function in a batch of different samples.
Materials and Methods
Reagents and assay kits
Lactic acid was purchased from Shanghai Aladdin Biochemical Technology Co. Ltd. (Shanghai, China). Crystal violet and ectoine were purchased from Shanghai Macklin Biochemical Technology Co. Ltd. (Shanghai, China). The FastPure Cell/Tissue Total RNA Isolation Kit V2, HiScript III RT SuperMix for qPCR (+gDNA wiper), and PerfectStart Green qPCR Super Mix were purchased from Nanjing Vazyme Biotech Co. Ltd. (Nanjing, China). Propidium iodide, acridine orange, and collagenase type IV were obtained from BioFroxx (Einhausen, Germany). Trypsin was provided by Thermo Fisher Scientific (Guangzhou, China), and Fetal Bovine Serum was supplied by Beijing Solarbio Science & Technology Co. Ltd. (Beijing, China).
Sample collection
Hydroxypropyl tetrahydropyrantriol, keratin, Amaranthus caudatus fermented extract, notoginseng total saponins, Gentiana extract, and glucose were obtained from the Chinese market. The samples were sequentially numbered (e.g., i–vi).
Zebrafish husbandry
Adult wild-type AB lines and Tg (lyz:EGFP) zebrafish were purchased from the China Zebrafish Resource Center. All the zebrafish experiments complied with the Organization for Economic Co-operation and Development Test Guidelines (TG203 and TG236). All animal experiments were conducted in accordance with the ethical standards of the Guangdong Provincial People’s Hospital Ethics Review Committee (S2024-1140-01).
All zebrafish were maintained in a recirculating aquaculture system (Shanghai Haisheng Biological Equipment Co. Ltd., Shanghai, China), under a 14-h light/10-h dark photoperiod. The room and tank temperatures were regulated to remain between 26.0°C and 29.0°C, and the water quality parameters, including pH and conductivity, were controlled at 7.0–8.0 and 500–550 μS/cm, respectively. The fish were fed live brine shrimp larvae twice daily to achieve apparent satiation. At night, prior to fertilization, zebrafish breeding tanks were arranged with dividers to separate male and female fish. The following morning, dividers were removed to facilitate mating. Subsequently, fertilized eggs that had fallen through the grid net at the bottom of the tanks were collected after 1 h, selected for viability, and then placed in an incubator at a temperature setting of 28.0°C–29.0°C for optimal development.
Establishment of a zebrafish skin barrier model
At 48 h postfertilization (hpf), embryos were washed and randomly selected zebrafish embryos were divided into several groups (with 20 embryos transferred to each six-well plate), and then incubated at a constant temperature of 28 ± 0.5°C with a pH of 7.4 ± 0.2. Embryos were first exposed to the tested samples for 1, 2, or 4 h. The embryos were exposed to 0.07%–0.11% lactic acid and 0.02 mg/mL of crystal violet for 20 min, and each concentration was tested in triplicate. Red fluorescence in the tail fin area was observed as an indicator of injury. Embryo phenotypes at 48 hpf were photographed individually using an Upright Fluorescence Microscope (Optec, Chongqing, China at 546 nm).
Histopathological sections
Zebrafish embryos were fixed in 4% paraformaldehyde for 24 h. Subsequently, the embryos were subjected to a series of treatments including gradient ethanol dehydration, paraffin embedding, sectioning, drying, and dewaxing. The sections were then stained with hematoxylin and eosin (HE), Masson’s trichrome, or Victoria blue. After staining, pathological changes between the groups were observed under a light microscope.
Acridine orange staining
Acridine orange reagent was used to visualize apoptotic cells, according to the manufacturer’s instructions. Zebrafish embryos were randomly selected and divided into several groups (10 per group), washed in phosphate-buffered saline (PBS), and then stained with 10 mg/L of acridine orange at 25.0°C–28.0°C for 30 min. The staining solution was aspirated and the embryos were washed in PBS. Zebrafish were anesthetized by immersion in 0.0025% tricaine solution. The embryos were observed under a confocal microscope (Carl Zeiss, Jena, Germany) at 485 nm. Each group underwent three independent biological replicates.
Reverse transcription quantitative real-time PCR
For qPCR analysis, three independent biological replicates were performed. For each replicate, 20 randomly selected zebrafish embryos from each group were pooled, and total RNA was extracted using the FastPure Cell/Tissue Total RNA Isolation Kit V2, followed by reverse transcription using the HiScript III RT SuperMix for qPCR (+gDNA wiper). Real-time qPCR analysis was carried out using the PerfectStart Green qPCR SuperMix on the Archimed X4 detection system (Bio-Rad). β-actin was used as the internal reference gene. The primer sequences are listed in Table 1. These embryos were separate from those used for live imaging or crystal violet staining.
List of Primers Used in Real-Time Polymerase Chain Reaction

Flow cytometric analysis of apoptotic cells
Zebrafish embryos were randomly selected and divided into several groups (40 per group), washed with PBS, homogenized, and centrifuged at 4816 xg for 3 min. The pellets were resuspended and treated with 0.25% trypsin and 1 mg/mL of collagenase at 37°C for 5 min. After digestion, 5% FBS in PBS was added to stop the reaction. Cells were counted using a Corning CytoSmart Cell Counter, resuspended in 1× Binding Buffer, and stained with Annexin V-FITC and PI. After incubation, the cells were analyzed using flow cytometry, and annexin V-FITC and PI were detected in their respective channels. Each group underwent three independent biological replicates.
Statistical analysis
All data collection was performed using ImageJ, and all analyses of the collected data were performed using Excel software. Differences between two groups were evaluated using Student’s t-test. Differences among multiple groups were analyzed using a one-way analysis of variance and post hoc tests. Differences were considered statistically significant at p < 0.05.
Results
Lactic-acid-induced skin barrier damage in zebrafish embryos
To evaluate the effect of lactic acid on the skin barrier of zebrafish, we stained the zebrafish skin with crystal violet and observed the penetration. First, the safety effects of lactic acid and crystal violet on zebrafish were evaluated separately by treating zebrafish embryos with 0.07%–0.15% lactic acid or 0.004–0.5 mg/mL crystal violet individually. After 10–20 min of exposure, lactic acid concentrations above 0.11% led to a significant increase in mortality (Table 2). In addition, significantly elevated mortality was also observed at crystal violet concentrations above 0.1 mg/mL (Fig. 1A, B). Therefore, the preliminary test concentration ranges were determined as 0.07%–0.11% for lactic acid and 0.004–0.1 mg/mL for crystal violet.
Dose- and Time-Dependent Mortality by Lactic Acid
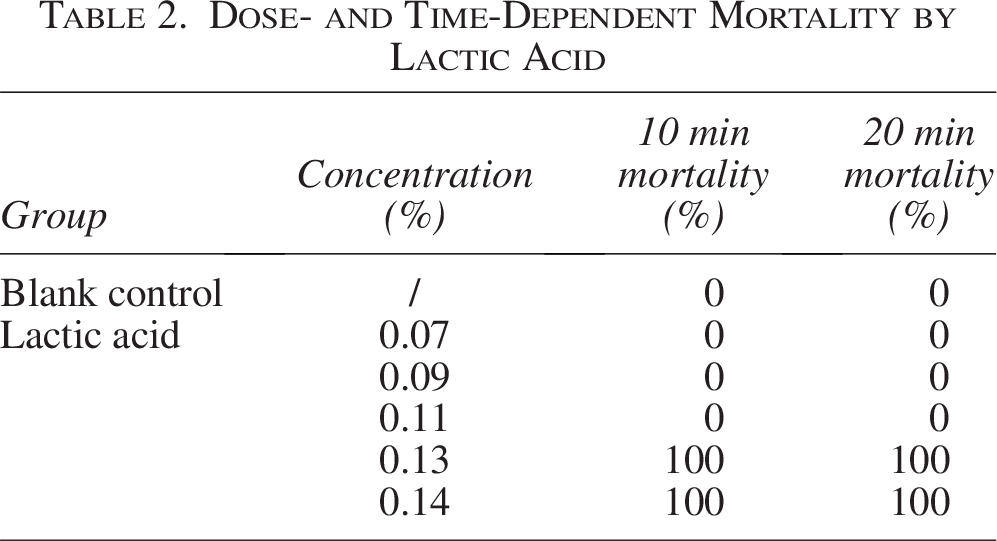
To further determine the treatment time for lactic acid, zebrafish embryos were exposed to 0.07%–0.11% lactic acid together with 0.02 mg/mL crystal violet for 10–20 min. The results showed that under 10 min of combined treatment, no visible increase in fluorescence was observed in zebrafish treated with 0.07%–0.11% lactic acid compared with the blank control group (Fig. 1C). Under 20 min of combined treatment, a visible increase in fluorescence was observed compared with the blank control group (Fig. 1D). Thus, it was determined that the established modeling condition—20 min of combined treatment with 0.11% lactic acid and 0.02 mg/mL crystal violet—is feasible and stable (Fig. 1E, F).
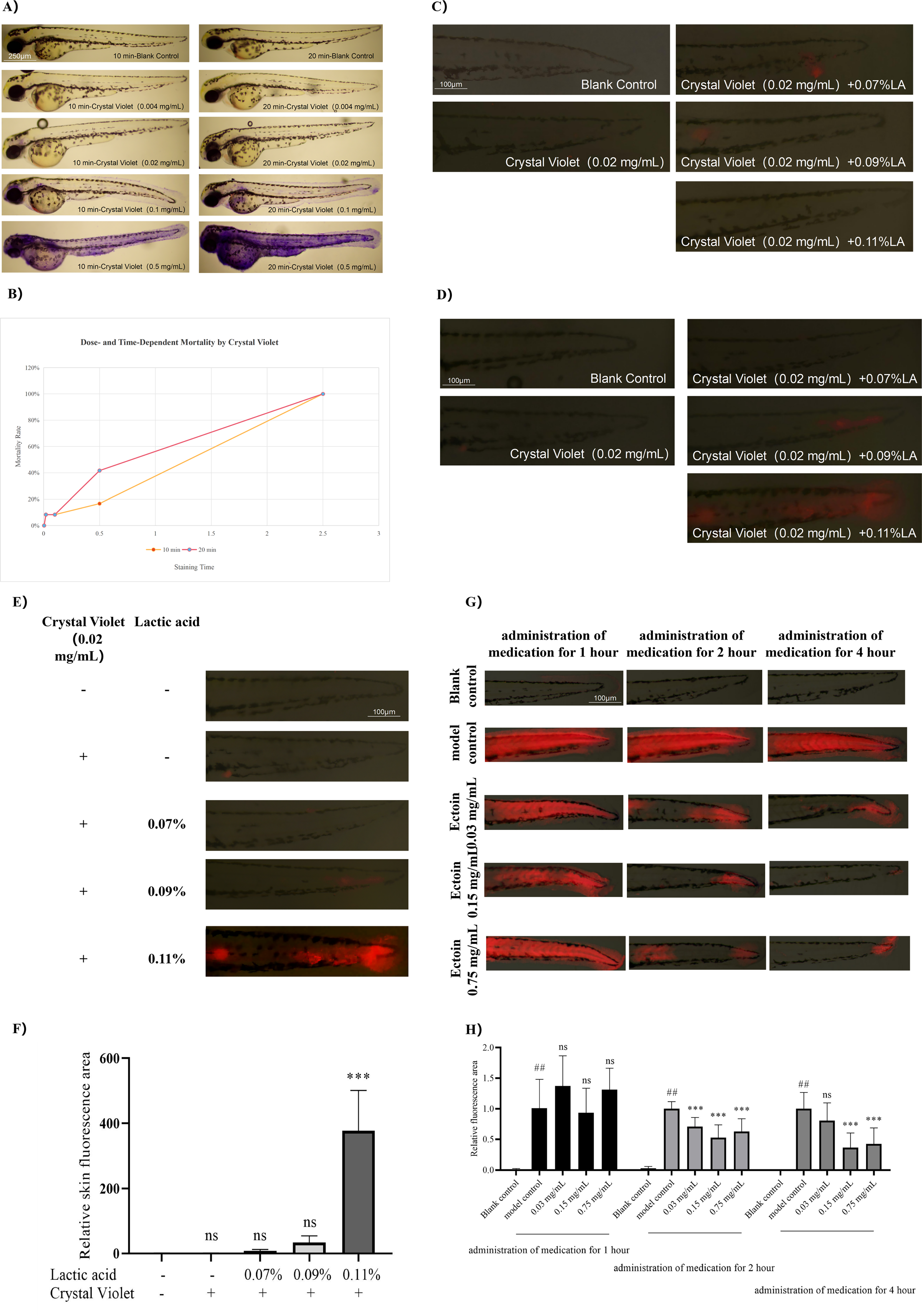
Lactic acid and crystal violet exposure leads to impaired skin barrier function in zebrafish.
Despite ectoine has been widely used to enhance the skin barrier, its specific effects on the zebrafish epidermal barrier have yet to be elucidated. To assess the effect of ectoine on the skin barrier damage model, zebrafish embryos were immersed in solutions containing 0.03, 0.15, or 0.75 mg/mL ectoine for 1, 2, or 4 h prior to lactic acid induction. These concentrations were selected based on practical application considerations. The protective effect of ectoine was positively correlated with the duration of treatment, with the most pronounced effect observed at a concentration of 0.15 mg/mL after 4 h (Fig. 1G, H). Based on these optimization results, the subsequent formal experiment was conducted using a mixture of 0.11% lactic acid and 0.02 mg/mL crystal violet to establish the model, while 0.15 mg/mL ectoine was used as a positive control, with a treatment duration of 4 h.
Histological analysis of lactic-acid-induced skin barrier disruption
To further validate the effects of lactic acid on the skin barrier of zebrafish embryos, histological sections were obtained post-lactic acid treatment and subjected to HE, Masson’s, and Victoria Blue staining to assess skin tissue pathology. Compared with the blank control group, the model control group exhibited compromised epidermal integrity. Initially, the epidermal layer thickened in response to lactic acid damage; with further damage, the outer epidermis eroded, resulting in epidermal thinning, and severe damage led to tissue loss in the zebrafish skin (Fig. 2A–C). Inflammatory cell infiltration was noted at the sites of epidermal injury upon HE staining (Fig. 2A), whereas no elastic fibers were detected with Victoria Blue staining (Fig. 2C), indicating that substantial elastic fibers had not developed in zebrafish at 48 hpf. Epidermal thickness measurements were performed on sections with minor injuries. Quantitative analysis revealed that lactic acid induced skin damage and inflammation (Fig. 2D), causing a significant increase in epidermal thickness (p < 0.001) and culminating in tissue loss.
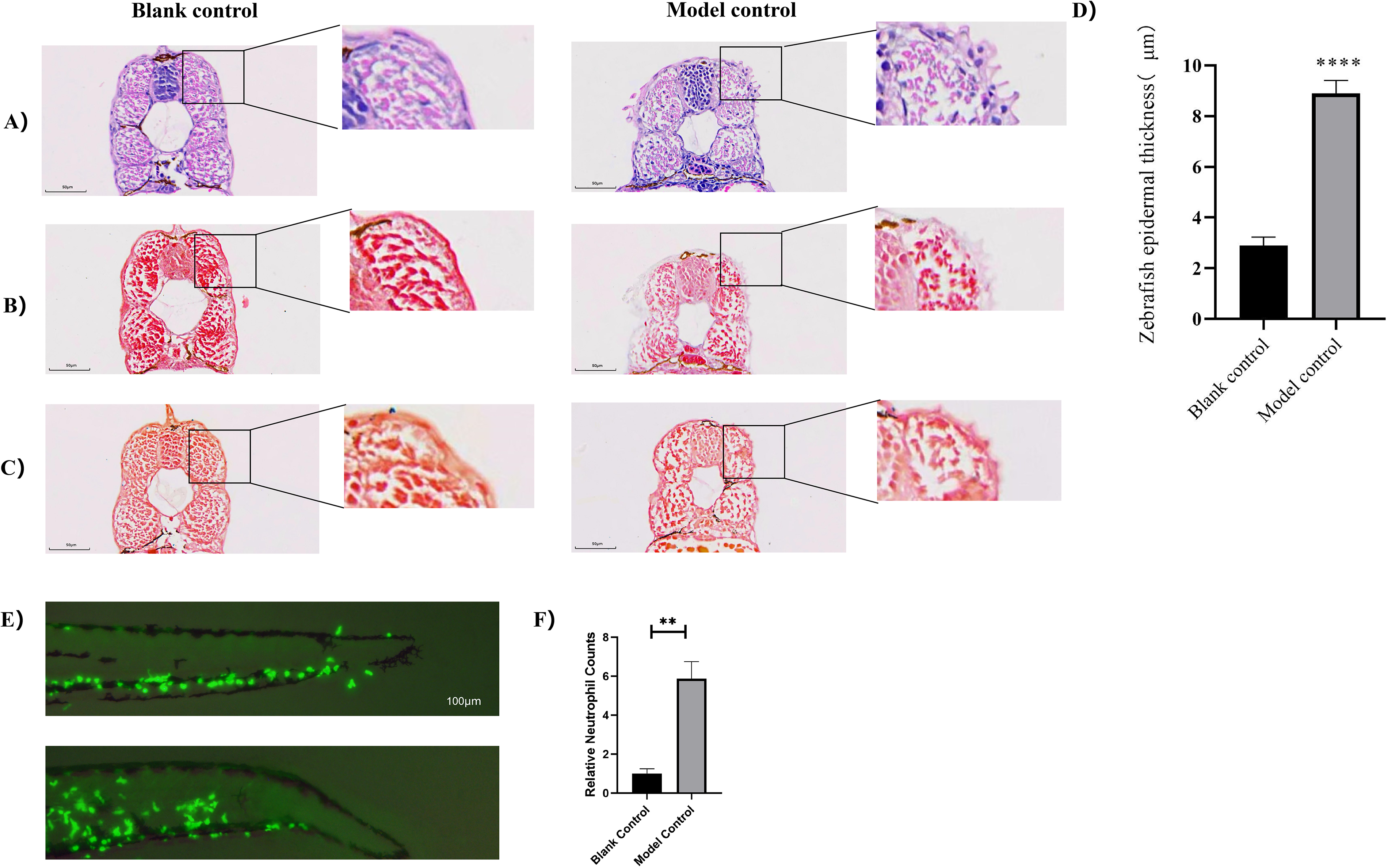
Lactic acid exposure leads to tissue damage in zebrafish skin.
Furthermore, the level of in vivo inflammation was observed using transgenic zebrafish Tg(lyz:EGFP) with fluorescently labeled neutrophils under an upright fluorescence microscope following lactic acid treatment (Fig. 2E, F). Compared to the blank control group, a significant increase in neutrophil migration was exhibited in the model control group, indicating the occurrence of inflammatory stress.
Validation of lactic-acid-induced apoptosis in zebrafish embryos
Acridine orange staining, RT-qPCR for apoptosis-related genes, and flow cytometry were performed to assess the effects of lactic acid on apoptosis in a zebrafish skin barrier damage model.
Acridine orange, which fluoresces in apoptotic cells, was used for visualization. Poststaining, a notable increase in dense green fluorescence was observed in the lactic-acid-treated group compared with that in the control, suggesting that apoptosis was induced (Fig. 3A, B). RT-qPCR analysis showed upregulation of apoptosis-associated genes, such as fas, baxa, and casp3a in the lactic-acid-treated group (Fig. 3C; p < 0.05). Flow cytometry indicated a higher proportion of early apoptotic cells in the treated group, with late apoptotic and dead cell nuclear debris percentages comparable to those in the control group (Fig. 3D, E).
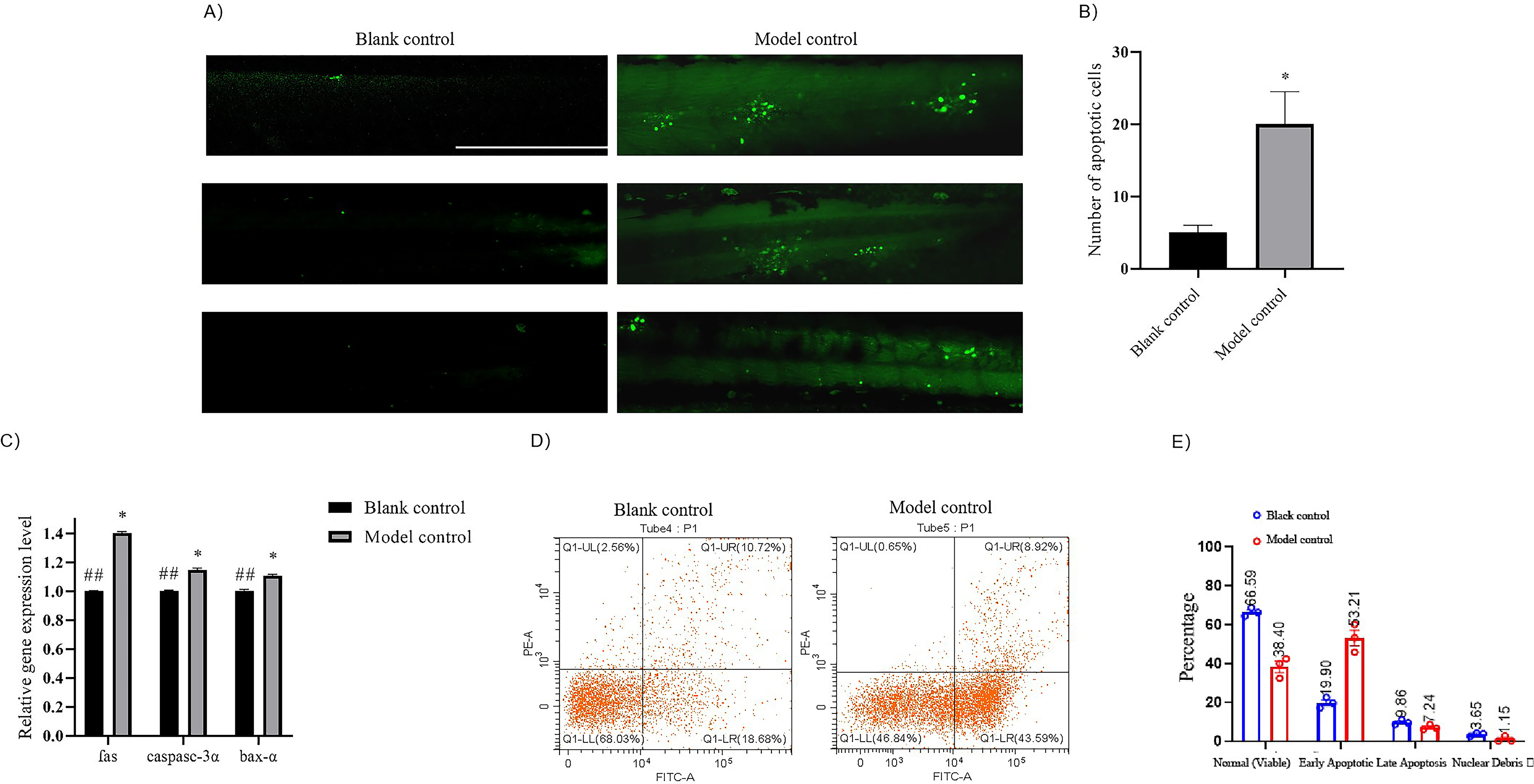
Lactic acid exposure induces apoptosis in zebrafish.
These findings collectively suggest that the skin barrier of zebrafish embryos is damaged and cell apoptosis is promoted by lactic acid.
Assess the raw materials restorative effects on the zebrafish skin barrier damage model
The restorative activities of six natural products and small molecules were assessed. Phenotypes were observed in the blank, model, and positive control groups, as well as in the treatment groups after lactic acid induction (Fig. 4A). The fluorescence area measurements are shown in Figure 4B. The model control group exhibited a significant increase in tail fluorescence than did the blank control group (p < 0.001), indicating successful model construction. Except for the blank group, the negative control group with glucose showed no significant increase in the fluorescence area compared with that in the model control group, confirming the reliability of the model. Groups treated with bosewellic acid, collagen peptides at high concentrations, and low to medium concentrations of A. caudatus fermentation extract, Panax notoginseng total saponins, and decolorized Gentiana extract anti-irritant factor (GB), showed significantly reduced fluorescence areas than those of the model control group (p < 0.05), suggesting restorative activity (Fig. 4B). Table 3 summarizes the restorative effects of the six natural products or small molecules.
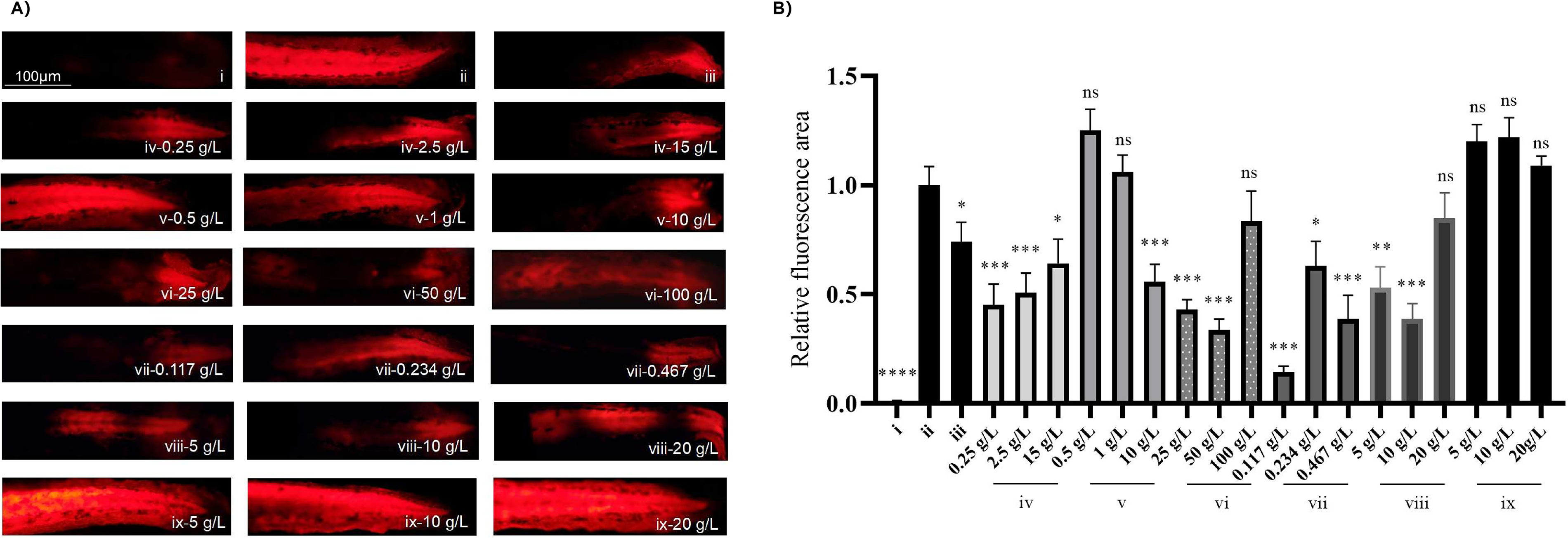
Screening of materials based on the zebrafish skin barrier injury model.
Quantitative Analysis of the Fluorescent Area in the Tail of Zebrafish
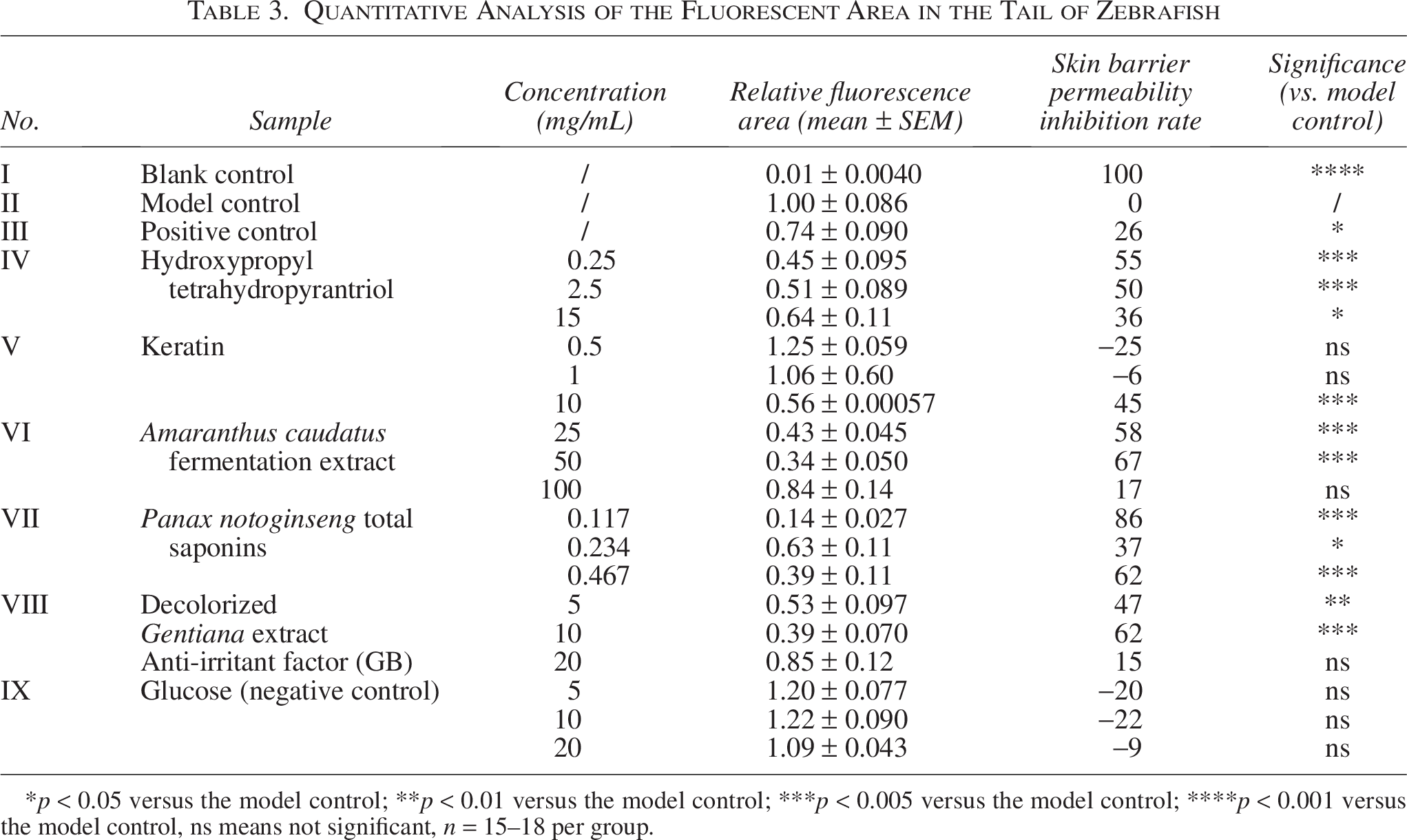
*p < 0.05 versus the model control; **p < 0.01 versus the model control; ***p < 0.005 versus the model control; ****p < 0.001 versus the model control, ns means not significant, n = 15–18 per group.
In addition to collagen peptides and glucose, no dose-dependent effects were observed for other compounds. A negative correlation between concentration and effectiveness was noted for hydroxypropyl tetrahydropyrantriol, A. caudatus fermentation extract, P. notoginseng total saponins, and decolorized Gentiana extract anti-irritant factor (GB), highlighting the need to determine the optimal concentration for maximum restorative activity in future material applications.
Discussion
The skin barrier is crucial for maintaining overall skin health and providing vital defense against environmental stressors, infections, and dehydration. Its key functions include preventing water loss, protecting against microbial invasion, and mitigating the effects of ultraviolet radiation and oxidative stress.8,36 Impairment or disruption of this barrier is associated with various dermatological conditions, including AD, psoriasis, contact dermatitis, ichthyosis, seborrheic dermatitis, acne, and rosacea.37–39 The significance of accurate methods for assessing skin barrier function cannot be overstated as they are essential for understanding the pathophysiology of skin diseases, guiding therapeutic strategies, and developing effective skincare products.
In clinical research, TEWL serves as the primary indicator of skin barrier function. TEWL measures the evaporation of water from the dermis through the epidermis, reflecting the integrity of the barrier. An increase in TEWL signifies a compromised barrier that allows more water to evaporate from the skin surface.40–42 Although human testing offers the most reliable data, preclinical evaluation methods are essential because of the environmental sensitivity of tests, ethical concerns, and high costs involved.
In preclinical research, reconstructed human epidermis (RhE) models and excised human skin are frequently used to assess the permeability of chemical substances through the skin barrier. 43 Previous studies have highlighted the importance of accurate and reliable in vitro models for predicting the permeability of substances when applied to human skin. For example, Gabbanini et al. 44 conducted an in vitro evaluation of essential oils from cosmetic formulations using RhE; Netzlaff et al. 45 assessed the permeability of the Episkin model in comparison to various human skin preparations. Additionally, animal models are commonly used in preclinical research to evaluate barrier function by observing pathological changes in the skin. Despite this, the research throughput on mammalian barrier function remains low; therefore, there is a need for efficient screening models. Zebrafish have long been recognized as reliable vertebrate models for skin physiology research. However, to the best of our knowledge, a fully mature model for studying zebrafish skin barrier function is yet to be established. Therefore, this study aimed to develop an in vivo zebrafish embryo model to evaluate skin barrier function, offering a novel approach that complements existing preclinical methods.
Lactic acid is a known irritant that can damage human skin structure, disrupting the skin barrier. For instance, Hasan et al. 46 conducted a transcriptional analysis of the RhE following the topical application of lactic acid, revealing that lactic acid induced the expression of damage-associated genes and provided valuable insights into the molecular mechanisms underlying the response of the skin to lactic acid. Hsiao et al. 47 treated human keratinocytes (HaCaT) with lactic acid and performed western blotting, which revealed that lactic acid induces oxidative damage and apoptosis in HaCaT by increasing the expression of fas, baxa, and casp3a. In addition, Ding et al. 48 conducted a human lactic acid sting test (LAST) that revealed significant differences in the sensitivity scales of stinging, itching, and tightness in the positive group. The LAST scores were negatively correlated with stratum corneum hydration and positively correlated with TEWL. These findings are important for understanding how lactic acid can compromise skin barrier function.
In this study, to establish a moderate level of skin barrier damage in zebrafish, we first explored and optimized lactic acid conditions. To enable the rapid and effective visualization of barrier function, we used crystal violet, a water-soluble fluorescent dye that cannot permeate through the healthy skin of zebrafish embryos, as a tracer to assess barrier damage. The results indicated that the red fluorescence of crystal violet could mark the location and extent of damage to the skin barrier caused by lactic acid, whereas the positive control, echinacea, significantly reduced this fluorescent signal. This method facilitates rapid and straightforward visualization of barrier damage. A quantitative analysis of the barrier function can be conducted by statistically analyzing the intensity of the fluorescent signal.
Lactic acid induces skin damage, which manifests as abnormal peeling, inflammation, and apoptosis of skin cells, ultimately affecting the skin barrier integrity. To validate the pathological alterations in our model, we analyzed skin structure and apoptosis in zebrafish embryos following lactic acid exposure. Histopathological sections revealed significant damage to the embryonic epidermis, with a marked reduction in the epidermal thickness surrounding the lesions and the presence of overt inflammatory cells. Although skin damage was apparent, Masson’s and Victoria blue staining yielded no distinct positive signals, which may be due to insufficient accumulation of dermal collagen and elastic fibers in the zebrafish embryos at 48 hpf. Apoptosis was significantly induced by lactic acid treatment, as evidenced by the increased number of positive cells following AO in situ and PI staining in flow cytometry. Additionally, a significant upregulation of pro-apoptotic genes, including fas, casp3a, and baxa, was noted. Therefore, the data suggest that exposure to lactic acid under the conditions of this model leads to compromised epidermal integrity in zebrafish embryos, accompanied by upregulation of cellular apoptosis, culminating in diminished skin barrier function. Although lactic acid promoted the apoptosis of zebrafish skin cells, it failed to represent the death of skin-specific cells. Furthermore, lactic acid induced considerable biological variability in skin barrier function damage, which had certain limitations.
Ectoine, a raw material frequently used to enhance skin barrier function, has been extensively investigated. Research has indicated that ectoine can protect the application site from inflammatory responses induced by dryness, enhance the fluidity and functionality of the stratum corneum, and consequently decrease TEWL. 49 Given these findings, ectoine was used as a positive control agent in the present study. To determine the applicability of this model, various substances, including small-molecule compounds and botanical extracts, were collected and subjected to assays to improve skin barrier function, with glucose serving as a negative control. Glucose did not exhibit any significant effects across all tested concentrations, which was consistent with our expectations. Other substances have demonstrated the potential to enhance skin barrier function to varying extents, suggesting that their efficacy may be influenced by factors, such as concentration and chemical composition. However, a decline in the effectiveness was observed at excessively high concentrations. This reduction in effectiveness could be attributed to the presence of irritating impurities in the extracts or the adverse effects of high ionic concentrations. These findings highlight the critical importance of determining optimal exposure concentrations when applying this model to research.
Although the skin of zebrafish is similar to that of mammals, consisting of three layers, the epidermis, dermis, and subcutaneous layer, differences in skin structure and development between zebrafish and mammals have been observed. 50 Hair follicles and sebaceous glands are absent in zebrafish, which possess mucus-secreting cells and special aquatic structures, such as a lateral line containing enlarged hair cells for mechanosensation. Under normal circumstances, zebrafish epithelial cells are not keratinized but are only replaced upon death or injury, and the ability to proliferate is retained by all epidermal cells. 51 In the middle layer, most epithelial cells remained undifferentiated. The middle layer, which is composed of dispersed ionocytes, sensory, and mucus cells, forms a mucus layer on the surface of zebrafish. 52 The functions produced by these cells are similar to those of mammalian bronchial epithelial cells. 53 In addition to oxygen exchange, they can also provide many important functions such as physical protection, osmosis and ionic balance, and acid/ammonia excretion. 54 Due to the differences in the skin structure between zebrafish and humans, fish lack a cuticle; therefore, it is difficult to reflect the effect of cuticle improvement, which imposes some restrictions on the practical application of raw materials. In addition, the skin barrier can be damaged by chemicals and factors, such as ultraviolet radiation and physical damage, which can affect its normal function. The impact of chemical factors on skin barrier function is reflected in this model and the protective effects of compounds on chemically damaged skin barriers should be evaluated.
In conclusion, although the skin structure of zebrafish embryos is less complex than that of mammals, their suitability as whole organisms for high-throughput screening makes this in situ and in vivo evaluation method a valuable complement for preclinical skin barrier function research. This approach is particularly promising for its application in pathology research, development of therapeutic strategies, and assessment of pharmaceuticals and skincare products.
Authors’ Contributions
Q.T.: Conceptualization, data curation, methodology, original draft, and writing—review and editing, supervision, and project administration. M.H.: Methodology, formal analysis, investigation, validation, data curation, writing—original draft, visualization, and project administration. J.Z.: Formal analysis, investigation, and validation. Y.W.: Validation. X.C.: Writing—original draft. Y.L.: Data curation. E.Y.: Conceptualization, methodology, supervision, and project administration. H.Z.: Conceptualization, methodology, writing—review and editing, supervision, and project administration.
Footnotes
Acknowledgment
The authors thank Guangzhou Shanhai Biotechnology Co. Ltd. for collecting samples.
Data Availability Statement
Data supporting the findings of this study are available upon request from the corresponding authors. The data are not publicly available because of privacy and ethical restrictions.
Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This study did not receive any specific grants from funding agencies in the public, commercial, or nonprofit sectors.