
Abstract
In many research studies, the handling and procedures performed can generate stress and require the use of anesthesia or sedation. In this sense, plant-based extracts named essential oils have emerged as potentially effective and environmentally safe anesthetic/sedative options. Here, we sought to evaluate the essential oil of Varronia curassavica (VCEO) as an anesthetic/sedative ingredient. Here, the anesthetic and sedative capabilities of the VCEO using adult zebrafish (Danio rerio). The anesthetic induction was performed under four different concentrations (100, 200, 400, and 600 mg L−1), followed by evaluation of recovery time; furthermore, a 3 h long exposure was performed for sedative testing, where fish were exposed for 3 h at three different concentrations (50, 80, and 100 mg L−1) under a superpopulation condition, such as those faced during transport. To assess stress, we collected whole-body cortisol from each individual after each exposure. The concentrations of 400 and 600 mg L−1 induced anesthesia (S4) but promoted excessive recovery times; on the contrary, they reduced the cortisol levels of the fish compared with the positive and negative controls. In the sedative testing (3 h long exposure), the animals exposed to 50 mg L−1 remained in S2 stage from the first 10 min, and those exposed to 80 mg L−1 after 20 min. The 100 mg L−1 induced stage S3a. There were no significant changes in cortisol levels observed at these concentrations. Limitations include measurement of whole-body cortisol at a single endpoint and the consistently slower induction and recovery with VCEO compared with a reference anesthetic MS-222, a property that constrains its practical use and warrants further dose–response, time-course, and recovery-optimization studies. Therefore, VCEO appears to be a promising anesthetic and sedative alternative, effective and plant-based, without increasing the physiological stress of the animals.
Keywords
Introduction
The use of zebrafish (Danio rerio) as a model organism, due to its genetic similarity to humans 1 is growing in research on embryonic development, 2 mental, metabolic, pathological, 3 neurological, 4 and behavioral 5 disorders. The pharmaceutical field, in particular, uses this animal model primarily in research focused on toxicology and the development of new drugs.6–9 In all these studies, the handling and procedures performed can generate stress and require the use of anesthesia or sedation to reduce stress and prevent animal suffering. 10 Its subsequent effects can compromise the lives of these organisms and impair the accuracy of the research. 11
It is known that these anesthetic/sedative tools, synthetic or plant-based, have in some cases demonstrated an effect on inducing physiological stress responses in organisms, resulting in increased cortisol levels.12,13 This occurs through the recruitment of the hypothalamic–pituitary–interrenal (HPI) axis, 14 resulting from the disruption of homeostasis by stress, promoting the release of corticotropin-releasing hormone (CRH) by the hypothalamus. In the pituitary gland, CRH stimulates the release of adrenocorticotropic hormone, which in turn stimulates the adrenal cells to release cortisol.14,15 With this, it is important to find anesthetic/sedative options that do not cause stress to the animals, especially based on the evaluation of biochemical parameters such as the cortisol levels. 16
Over time, it has been noticed that phytotherapeutic alternatives have been considered in aquaculture13,17 for research or production purposes.16,18,19 Essential oils (EO) are examples of phytotherapeutic alternatives to sedatives/anesthetics. These can be allies in medical treatments, acting as antibiotics, antioxidants, and even as auxiliary medications in treatment protocols used in human medicine. 20 In ornamental or food fish farming, EOs can assist in nutrition, reproduction, and disease treatment.21,22 The chemical compositions and low environmental impact (at appropriate concentrations) due to the biodegradability of EO 23 indicate promising plant-based options with potential anesthetic and sedative effects on ichthyofauna.18,19,24 In light of this, EOs can be considered a good and effective methodological alternative in the field of anesthesiology.
A plant of interest in the field of phytotherapeutics is the plant species popularly known as “erva-baleeira” Varronia curassavica Jacq. (Cordiaceae), also known by the synonym Cordia verbenacea DC. (Boraginaceae). This plant has a shrub-like growth habit and is native, occurring throughout much of Brazil, mainly in the Atlantic Forest and Pampa biomes. 25 The leaves have glandular trichomes that secrete the EO, a product of the plants’ secondary metabolism, with a wide variety of chemical compounds such as monoterpenes and sesquiterpenes.19,26,27 In Brazil, the tradition of using this species in folk medicine has been cultivated for generations in the treatment of bruises and inflammations. 28 In fact, V. curassavica is on the list of medicinal plants indicated by the Agência Nacional de Vigilância Sanitária (ANVISA) in the Formulary of Phytotherapeutics for the treatment of localized inflammatory processes, 29 thus showing itself as an important therapeutic alternative.
Furthermore, different pharmacological actions have been described for the plant, such as antifungal, 30 antinociceptive, 31 antibacterial, 32 anti-inflammatory, and antiallergic.33,34 In the area of fish production, the essential oil of V. curassavica (VCEO) is also known to have sedative effects in a dose-dependent manner without causing visible adverse effects on the behavior of animals of the species Oreochromis niloticus and Rhamdia quelen.18,19
Therefore, this study had as its primary objective to evaluate the anesthetic and sedative potential of the VCEO in adult zebrafish. A secondary objective was to test the oil’s tranquilizing effect during prolonged exposure, simulating practical situations such as transport and routine handling, by exposing animals to subanesthetic/sedative concentrations for 3 h and assessing stress reduction. To this end, acute anesthesia and short sedative trials were used to characterize induction and recovery, while the 3-hour-long exposure experiments were specifically designed to evaluate changes in stress levels via whole-body cortisol.
Material and Methods
Ethical note
This study was approved by the Animal Use Ethics Committee of the University of Passo Fundo under protocol number 007/2025.
Study strategy
Initially, we exposed eight zebrafish individuals to a concentration of 100 mg L−1 of VCEO to determine the concentrations for further anesthesia induction (S2 stage) and recovery tests, and 3 h long sedative testing. For the anesthetic induction and recovery test, eight fish were exposed to each of the concentrations (100, 200, 400, and 600 mg L−1) of V. curassavica essential oil for 30 min or until they reached the stage of anesthesia. The 3 h long sedative testing method was carried out at concentrations of 50, 80, and 100 mg L−1 with eight fish for each of the three concentrations, in triplicate, 24 fish by concentration. As behavioral observations, signs of adverse effects, and mortality over the course of 3 h (0, 10, 30, 50 min, 1, 2, and 3 h) were observed for 5 min each time period. Subsequently, fish from both experiments were euthanized with spinal cord sectioning after being desensitized in ice, frozen, and stored for later cortisol analysis. In the anesthetic induction and recovery test, we included a positive control consisting of 12 zebrafish of the same strain and origin treated with the reference anesthetic MS-222 (tricaine methanesulfonate) at a concentration of 150 mg L−1. See the schematic representation of our experimental design in Figure 1.
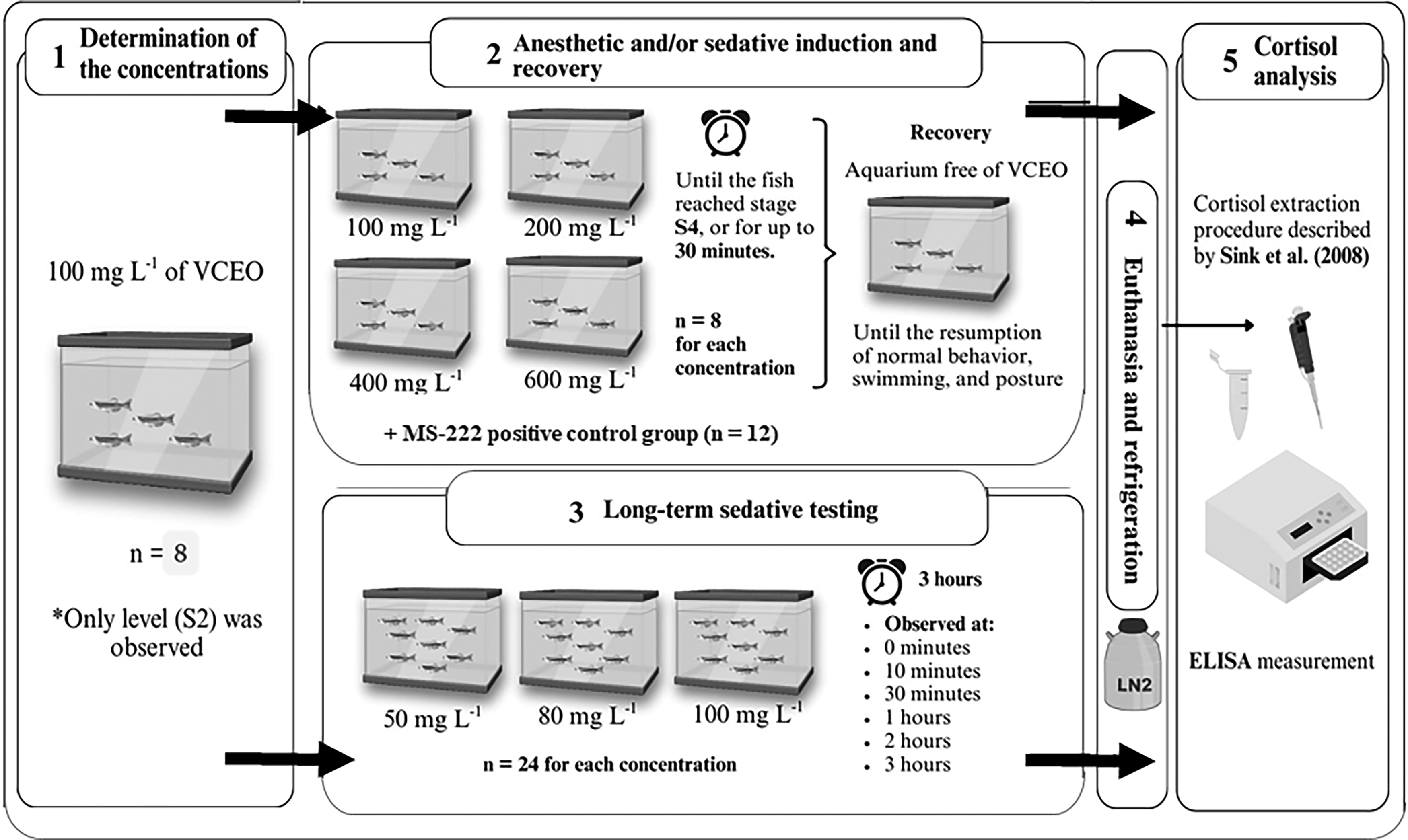
Schematic representation of the experimental design. Made with Biolcons and Biorender®.
Animals
One hundred and 40 adult zebrafish (Danio rerio) of both sexes were used (1:1 male/female), obtained from the Fish Physiology Laboratory of the Veterinary Medicine Faculty at the University of Passo Fundo. Prior to the tests, the fish were kept in 3.7 L glass tanks (five fish per tank), with an artificial photoperiod (14 h light: 10 h dark) and constant aeration, supplied with filtered water and monitored for ideal levels of temperature (27 ± 2°C), oxygen (6.0 ± 0.4 mg L−1), pH (7.0 ± 0.25), and ammonia (concentration <0.5 mg L−1) in a bioassay room. They were fed twice a day with commercial Basic Alcon feed (45% raw protein).
Plant and phytochemicals
The 100% pure EO of V. curassavica (labeled as C. verbenacea) was commercially acquired from the company Herbia® (Joinville, Brazil). The technical specifications highlight its liquid appearance, yellow color, characteristic strong resinous aroma, density at 20°C of 0.870, and refractive index at 20°C of 1.489. The chemical composition of the commercial EO used in this study was provided by the manufacturer and is presented in Table 1. These values were validated and cited in our previous study by Fortes et al. 18 and were consistent with the manufacturer’s analysis.
Chemical Composition (%) of the Essential Oil of Varronia curassavica

VCEO, essential oil of Varronia curassavica.
Determination of the concentrations
The concentrations were determined from an initial experiment where eight zebrafish individuals were exposed to a concentration of 100 mg L−1 of VCEO, aiming to understand whether this concentration would exhibit anesthetic or sedative activity. With this concentration, only central depression at the sedation level (S2) was observed. Therefore, in order to achieve activity at the anesthesia level (S4), three other higher concentrations were tested: 200, 400, and 600 mg L−1. For the 3 h long sedative testing experiment, concentrations of 100 mg L−1 and two lower concentrations were used: 80 and 50 mg L−1.
Anesthetic and/or sedative induction and recovery
For the anesthetic/sedative testing, eight unsexed adult fish (8 months) were placed in 3.7 L tanks where they were exposed to different concentrations of VCEO up to 30 min (n = 8). VCEO was first diluted in ethanol (95%) at a 1:10 ratio and tested at four different concentrations: 100, 200, 400, or 600 mg L−1 of EO. After the exposure, each individual was stimulated using a glass rod and assessed by the observation of the sedative stages as described by Schoettger and Julin: 36 sedation (S2), where there is a reduction in response to visual stimuli, as well as a reduction in locomotor and opercular activities; partial loss of balance (S3a), in which the fish lose balance in the water current and have reduced swimming ability; total loss of balance (S3b), where there is a loss of locomotion ability, reduced opercular function, and responses only to pressure on the caudal peduncle; and anesthesia (S4) or loss of reflexes, with responses to external stimuli ceasing. Medullary collapse with cessation of opercular activity (S5) indicates mortality.
The fish remained in the tank with VCEO until they reached stage S4 (loss of reflexes and reaction to environmental stimuli), or for up to 30 min. Afterward, they were relocated to an aquarium free of EO for recovery analysis, where they were assessed for resumption of normal behavior, swimming, and posture, as observed prior to the test. Then, they were weighed, measured, frozen in liquid N2, and stored at −80°C. 37
Twelve fish of the same strain and origin were used as a positive control and exposed to buffered MS-222 (150 mg L−1). Each fish was transferred individually to a 3.7 L tank containing the MS-222 solution, which was maintained at the same temperature, dissolved oxygen, and pH conditions as the VCEO induction/recovery experiment. Induction and recovery were assessed using the same criteria, and the time to reach each criterion was recorded. After full recovery, fish were returned to the holding system and observed for 24 h for any delayed effects. Induction and recovery times for MS-222 were compared with those obtained for VCEO.
Three hours long sedative testing
To evaluate the sedative and stress-reducing effects of the VCEO, eight fish, in triplicate, were exposed in a 3.7 L aquarium to simulate a high-density environment similar to what occurs during the transport of farming fish. Each tank was exposed to one of the three determined concentrations of VCEO (50, 80, and 100 mg L−1) for 3 h, in triplicate.18,19 Observations were made at 0, 10, and 30 min, and 1, 2, and 3 h to assess responses to stimuli, using a glass rod, and to monitor for any adverse effects and/or mortality. The control used was ethanol at a concentration of 1000 µL L−1. Subsequently, the animals were euthanized with ice-cold water followed by a spinal cord section and frozen as described previously in the anesthetic induction and recovery test.
Cortisol analysis
After euthanasia, fish carcasses were stored at −80°C until processing to preserve hormone integrity. The whole-body of fish was weighed and individually macerated in 30 volumes of phosphate-buffered saline pH 7.4 (PBS). The cortisol extraction procedure was performed according to the methodology described by Sink et al. 38 Briefly, one zebrafish (weighing more than 0.5 g) was euthanized, minced, and homogenized in a test tube for 30 s using 3 mL of PBS (pH 7.3). After that, 3 mL of ethyl ether was added to a different test tube containing a 1 mL aliquot. After homogenizing the mixture for 30 s, liquid nitrogen (N2) was added. After three repetitions of this procedure, the tube was immersed in a water bath at 37°C for 12 h in order to fully evaporate the ethyl ether. The lipid extract was then resuspended in 200 μL of PBS and kept at −20°C for cortisol immunoassay. Cortisol levels were quantified using an enzyme-linked immunosorbent assay kit (ELISA; EIAgen CORTISOL test, Bio Chem Immuno Systems). Cortisol analysis control was performed using ethanol 15 at a concentration of 6000 μL L−1 for the anesthetic and/or sedative induction and recovery test, and at 1000 μL L−1 for the 3 h long sedative test.
Statistics
The data were presented as mean ± SD or as median ± interquartile range and analyzed using either one-way Analysis of Variance (ANOVA) followed by Tukey’s multiple comparisons test or the Kruskal–Wallis test followed by Dunn’s test, depending on data normality as assessed by the Bartlett and Brown–Forsythe tests. All statistical analyses were conducted with GraphPad Prism version 8.0.0 for Windows (GraphPad Software, San Diego, California, USA). Statistical significance was considered at p < 0.05.
Results
Anesthetic and/or sedative induction and recovery
Individuals exposed to 100 mg L−1 VCEO reached only the sedation stage (S2). Fish exposed to 400 and 600 mg L−1 VCEO reached S2 in times similar to those observed with MS-222 (Fig. 2a), whereas individuals exposed to 200 mg L−1 attained stage S3b (incomplete anesthesia). Fish exposed to 600 mg L−1 VCEO reached S3b in times comparable to MS-222 (Fig. 2c). Both 400 and 600 mg L−1 produced full anesthetic activity at the S4 level (Fig. 2d); however, animals in these VCEO groups reached S4 more slowly than those exposed to MS-222.
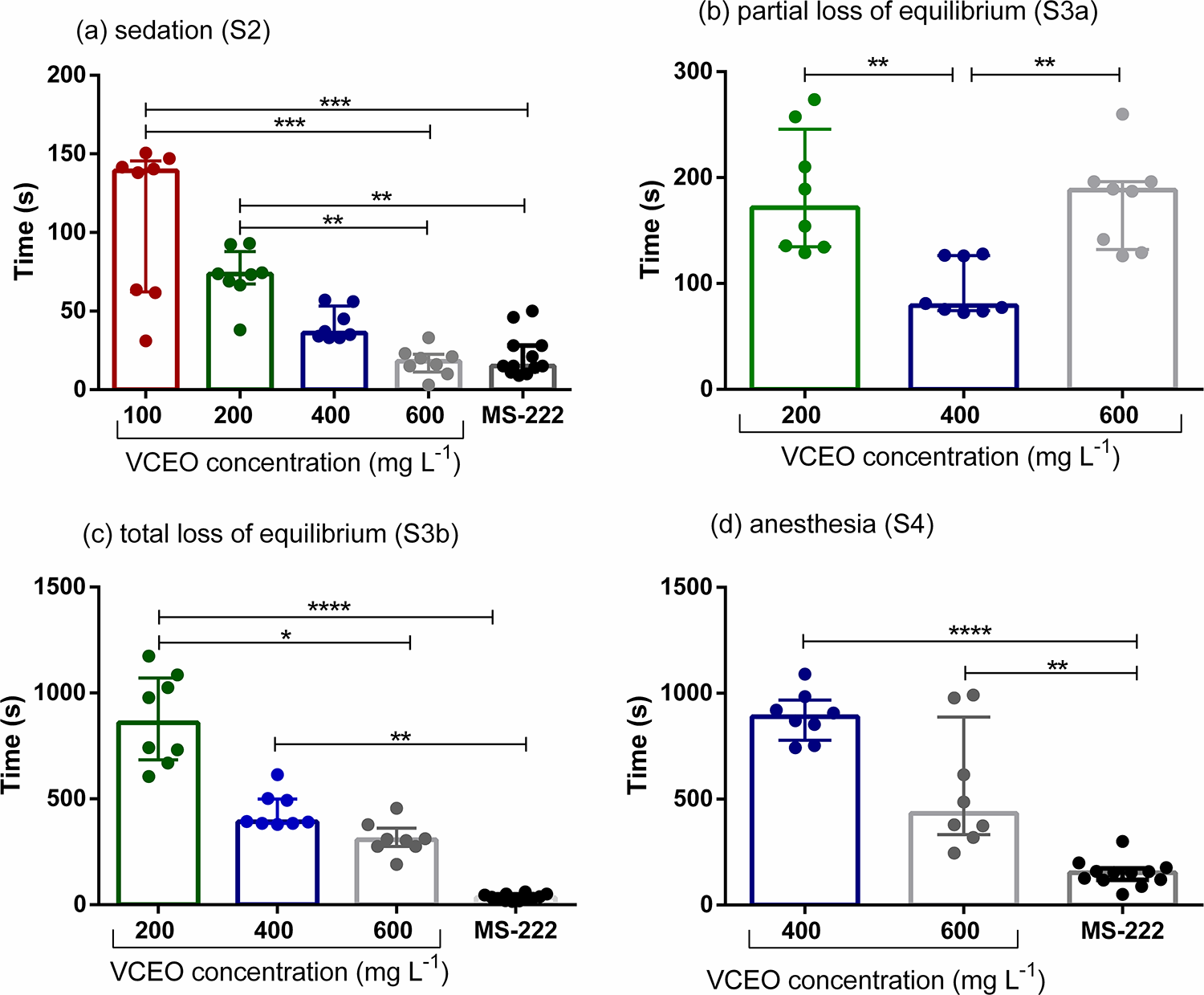
Time at which zebrafish (Danio rerio) individuals reached each stage under the different concentrations of VCEO and to MS-222 (Stages S2, S3b, and S4), namely
Regarding recovery, animals exposed to 400 and 600 mg L−1 VCEO required similar times to regain normal behavior and activity; however, both groups recovered significantly more slowly than fish exposed to MS-222 (Fig. 3). Recovery times for the 100 and 200 mg L−1 VCEO groups were excluded from this analysis because those fish did not reach the S4 stage.
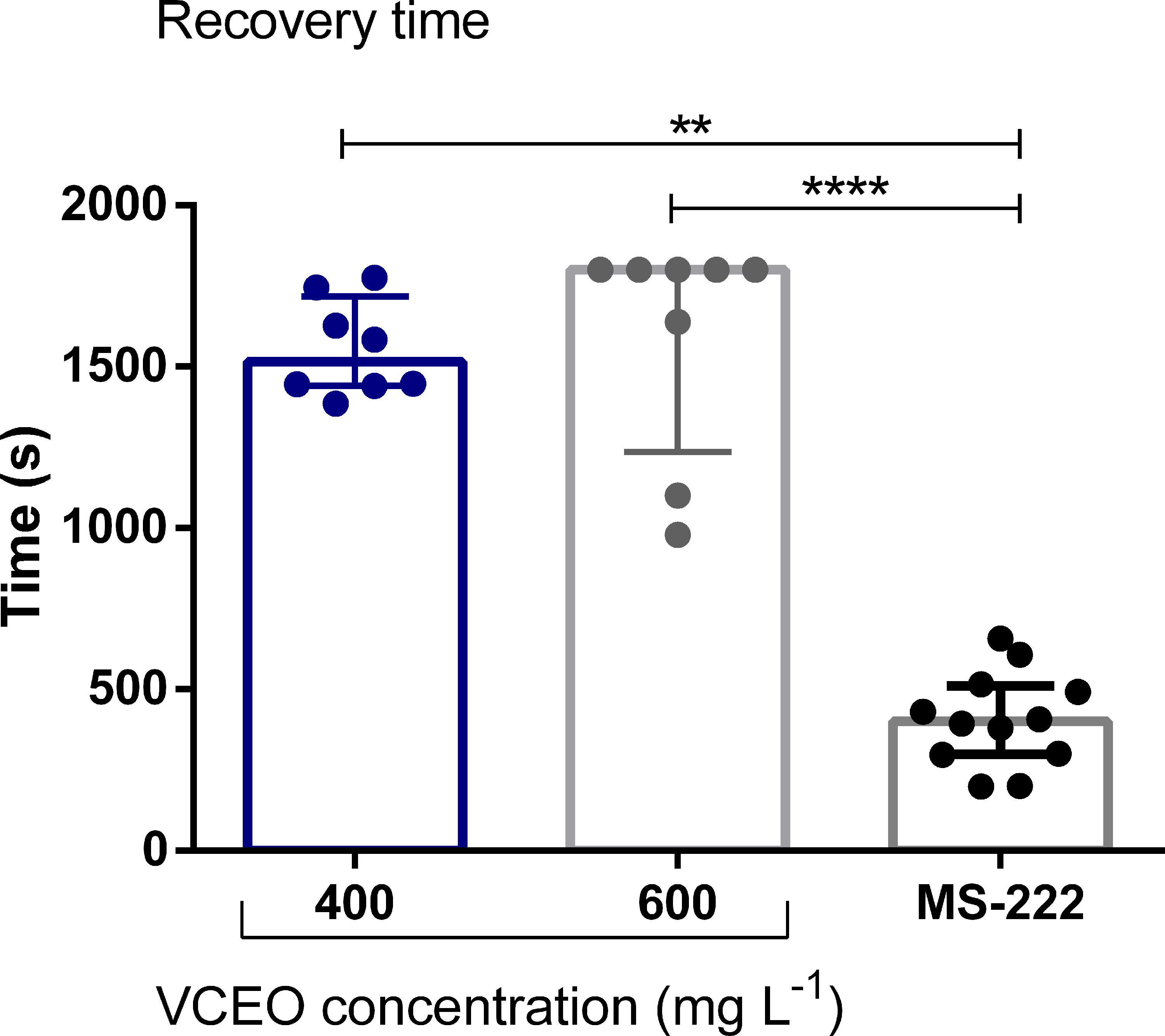
Recovery time of zebrafish (Danio rerio) individuals exposed to anesthetic concentrations of VCEO (400 and 600 mg L−1) and to MS-222. Data expressed as median ± interquartile range and analyzed by Kruskal–Wallis test followed by Dunn’s (**p < 0.01 and ****p < 0.0001).
Three hours long sedative testing
Animals evaluated after a high-density group exposure to 50, 80, and 100 mg L−1 (Fig. 4) of VCEO demonstrated that up to the first 10 min, only the 50 mg L−1 concentration did not reach the sedation stage (S2), with all fish showing normal response to stimuli, while the other concentrations induced sedative activity. At 80 mg L−1, the animals reached sedation within the first 10 min. From 20 min onward, 50 and 80 mg L−1 of VCEO maintained a sedative effect at the S2 level on zebrafish individuals, while those exposed to the 100 mg L−1 concentration reached stage S3a in the second and third hours. No mortality or adverse effects were observed during the 3 h of exposure.
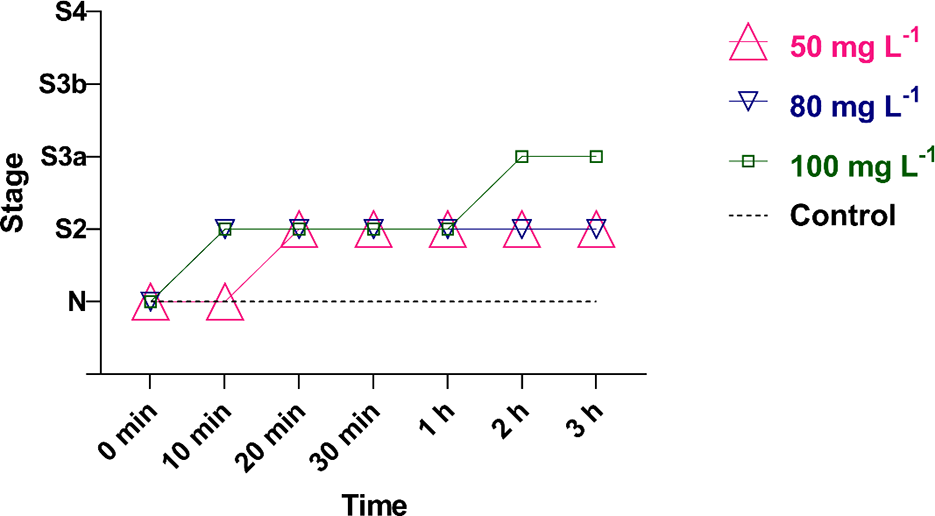
Sedation/anesthesia stages observed in zebrafish (Danio rerio) exposed to the essential oil of Varronia curassavica. N—normal behavior, S2—sedation, S3a—partial loss of equilibrium, S3b—total loss of equilibrium, and S4—anesthesia.
Cortisol analysis
The results obtained from the whole-body cortisol analysis of zebrafish (Danio rerio) are presented in Figure 5. The whole-body cortisol levels of zebrafish exposed to VCEO did not show statistically significant differences among the lower concentrations of 100 and 200 mg L−1 in the anesthetic induction test, nor among all three concentrations used in the long-term sedative testing test (50, 80, and 100 mg L−1), as compared with the control group. In contrast, fish exposed to the higher concentrations used for anesthetic induction (600 mg L−1) exhibited a significant reduction in cortisol levels when compared with the control group.
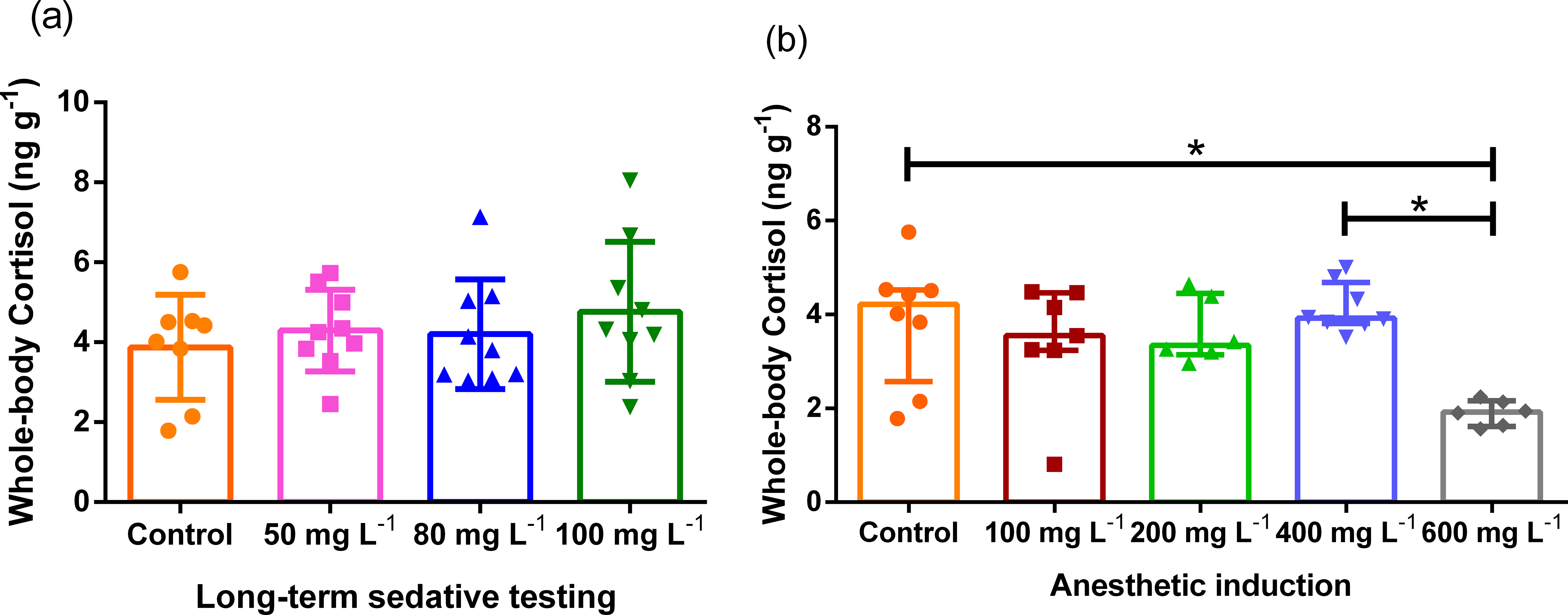
Mean cortisol levels in zebrafish (Danio rerio) exposed to different VCEO treatments and concentrations, where
Discussion
Here, we show that VCEO at concentrations of 400 and 600 mg L−1 exhibited anesthetic effects on zebrafish, reaching stage S4. However, fish exposed to these concentrations recovered from anesthesia between 24 and over 30 min, which is considered an excessively long recovery time compared with the recovery time of zebrafish anesthetized with MS-222 (2 min and 30 s),39,40 one of the most commonly used synthetic anesthetics in fish. In addition, other synthetic anesthetics, such as isoflurane 41 and propofol, 42 administered in aqueous solutions, are characterized by extremely fast induction (1–2 min) and consistent recovery, while etomidate 43 is valued for its minimal cardiovascular depression despite slightly longer recovery times. In addition, eugenol, 44 derived from clove oil, shows similar induction and recovery times, though with greater variability.
The induction and recovery time observed for MS-222 are consistent with values reported in the literature, supporting its use as a reference anesthetic in our study; although induction and recovery with MS-222 were significantly shorter than those observed for VCEO at 400 and 600 mg L−1, these longer induction and recovery times may be managed according to the specific procedure being performed under anesthesia, particularly if a longer recovery period is required or if the animal is to be euthanized for the collection of biological materials.
Regarding the 3 h long exposure sedative testing, the aim was to simulate a stressful environment by exposing a large number of individuals to VCEO in the same tank with a limited volume of water, similar to conditions encountered during transport. The 50 mg L−1 concentration resulted in continuous sedation for 3 h, while 80 mg L−1 produced the same effect from the first 20 min onward, indicating the potential use of VCEO as a sedative under stressful conditions. No adverse effects were observed during the entire period of prolonged exposure or anesthetic induction, highlighting the positive and promising impact of VCEO in promoting animal welfare and reducing stress-related effects that could interfere with experimental protocols.
The anesthetic potential of VCEO identified in the present study is likely directly related to α-pinene, a monoterpene that has already demonstrated central nervous system (CNS) inhibitory activity through modulation of GABAa receptors at benzodiazepine binding sites,45,46 with its effect being blocked when administered together with GABAa receptor antagonists. 30 Furthermore, activation of Gamma-Aminobutyric Acid a (GABA) receptors not only promotes anesthetic and sedative effects but also produces anxiolytic outcomes due to its inhibitory nature within the CNS, 47 recognizing that GABAergic receptor agonists are among the most widely used drugs for treating anxiety disorders. 48 While several major constituents of the oil have been associated with sedative or neuromodulatory effects in other systems, the present study did not test isolated compounds or fractions. Therefore, any attribution of the observed behavioral effects to specific constituents is tentative. Future work will include fractionation and testing of major components to determine their individual and synergistic contributions.
EOs comprise a wide array of chemical compounds, and consequently, various pharmacological effects may be observed. 49 In this context, the experiments conducted also revealed a potential stress-mitigating effect associated with higher concentrations of the EO (400 and 600 mg L−1), as evidenced by reductions in cortisol levels in exposed zebrafish. This effect is likely linked to the modulation of GABAa receptors previously mentioned, as well as to other constituents within VCEO, such as β-caryophyllene—a cannabinoid sesquiterpene that, in rodents, has exhibited anxiolytic effects analogous to benzodiazepine drugs. 50 This outcome appears to be directly associated with the agonist potential of β-caryophyllene on endocannabinoid receptors, eliciting anxiolytic and antidepressant effects through modulation of CB2 receptors present in the CNS, 5 including in limbic areas involved in the regulation of anxiety-type and stress responses. 51
Furthermore, analysis of cortisol results indicated that the group exposed to a higher concentration during anesthetic induction showed lower whole-body cortisol concentration, suggesting that this dose can mitigate the elevation of cortisol promoted by the exposure methodology verified in control and lower concentrations.
We acknowledge that the 3-h time sampling can in principle lead to attenuation of the HPI axis via negative feedback and thus lower cortisol levels; however, in our study, the cortisol decrease was observed only at the 600 mg L−1 VCEO treatment and not at the lower VCEO concentrations, all sampled at the same time points (Fig. 5b). If the reduction were driven primarily by time-dependent axis attenuation, a similar decrease would be expected across all VCEO treatments sampled under identical temporal conditions. The restriction of the effect to the highest concentration therefore weakens the negative-feedback explanation and instead supports a dose-dependent effect of VCEO (e.g., greater suppression of HPI activity or a stronger sedative action at 600 mg L−1). We therefore interpret the cortisol result as more likely reflecting a pharmacological effect at the highest dose, while noting the limitation of our single time-point design and recommending targeted time-course experiments to fully disentangle temporal attenuation from dose-dependent mechanism.
Considering these findings, the importance of analyzing the chemical composition of VCEO becomes evident. As a plant-derived material, VCEO undergoes changes in its compounds and their respective concentrations due to genetic, 26 geographic, 27 and seasonal factors, 52 as well as the life stage of the specimens, 53 which may consequently alter both the efficacy and potency of its pharmacological effects.
In summary, this study introduces a novel pharmacological alternative in the field of fish anesthesiology, derived from a plant species native to Brazil’s rich biodiversity, which encompasses approximately 45,000 species. 25 Many of these species remain underexplored in terms of their medicinal potential. Research of this nature not only expands the possibilities for applying plant diversity in healthcare but also enhances our understanding of plant–animal interactions in natural environments.54 As VCEO is extracted from a plant species, it is organic, biodegradable, and easily accessible, positioning it as an effective and low-cost therapeutic option.
We acknowledge important limitations. First, whole-body cortisol was measured at a single end point only, which precludes characterization of the temporal dynamics of the HPI response and therefore requires caution when interpreting the observed cortisol decrease; targeted time-course sampling would be necessary to distinguish true dose-dependent suppression from time-dependent axis attenuation or negative feedback. Notably, the cortisol reduction was observed only at 600 mg L−1 and not at 400 mg L−1, which weakens a simple time-dependent/negative-feedback explanation and instead suggests a concentration-dependent effect of VCEO. With respect to the oil itself, although VCEO exhibited sedative and anesthetic activity, induction, and recovery were consistently slower than with the field standard (MS-222), a property that imposes practical constraints on its use (e.g., for procedures requiring rapid induction or brief recovery) and warrants further optimization. Future studies should therefore include multiple sampling times, more detailed dose–response experiments, recovery-optimization protocols, and longer term safety and mechanistic investigations before VCEO can be recommended as a routine alternative to established anesthetics.
In conclusion, VCEO exhibited clear sedative and anesthetic effects in zebrafish: 50 and 80 mg L−1 reliably produced prolonged sedation (S2) under stressful conditions, while 600 mg L−1 induced full anesthesia (S4) and was associated with a significant reduction in whole-body cortisol. Importantly, no stress-related behaviors or mortality were observed under our experimental conditions, suggesting an acceptable acute safety profile. Taken together, these results indicate that VCEO has pharmacological potential as a plant-derived sedative and anesthetic for zebrafish, but further work, including detailed dose–response studies, time-course cortisol sampling, recovery-optimization protocols, mechanistic investigations, and longer term safety assessments, is required before VCEO can be recommended as a routine alternative to established anesthetics.
Authors’ Contributions
A.J.R.: Conceptualization; methodology, writing and editing, and data analysis. L.K.B.: Supervision, review, writing, and analysis. C.H.M.F.: Methodology, materials, and data analysis. A.P.G.F.: Methodology. E.B.: Methodology. A.C.S.: Methodology. J.A.D.L.: Methodology. W.A.T.: Writing and review. L.J.G.B.: Supervision, project administration, writing, review, methodology, and data analysis.
Data Availability
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Footnotes
Acknowledgments
The authors wish to thank the University of Passo Fundo, the Fish Physiology Laboratory at the University of Passo Fundo, and the Federal University of Santa Maria for their support with laboratory facilities and materials essential for the development of this research.
Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by grants from the National Council for Scientific and Technological Development (CNPq, process no.