
Abstract
Cannabis use during pregnancy is increasing. In rodent models of delta-9 tetrahydrocannabinol (Δ9-THC) exposure during pregnancy, placental pathology, including compromised labyrinth development, is reported. Cannabinoid receptor 1 (CB1/Cnr1) is the primary mediator of Δ9-THC action, with its expression reportedly limited to the placental junctional zone in the rodent. Given a Δ9-THC-induced labyrinth-specific pathology, we predicted more diverse expression. This study aimed to elucidate the spatiotemporal expression of CB1/Cnr1 in the rodent and assess whether it mediates Δ9-THC-induced alterations in trophoblast differentiation. Results revealed CB1 expression in all maternal blood-facing trophoblast cells. Furthermore, Δ9-THC exposure (at levels matching those reported in maternal serum) had a more significant effect on the expression of markers associated with differentiating trophoblast cells than on proliferating trophoblast stem (TS) cells. Δ9-THC impacted mouse (m) TS cell differentiation in a CB1-dependent manner, reducing the expression of syncytiotrophoblast (SynT) markers, driving differentiation along the junctional zone/trophoblast giant cell pathway. mTS cells without Cnr1/CB1 (mTS Cnr1 KO) did not express markers of SynT cells or the differentiated junctional zone cell types. However, at a higher than physiologically relevant concentration, Δ9-THC (15 μM) induced Gcm1 (SynT) expression in mTS Cnr1 KO cells. This study reveals a mechanism by which Δ9-THC may impact placental growth.
Keywords
Introduction
Stem cells are vital to the maintenance and regeneration of tissue because they both self-renew and differentiate into specialized cell types, making them an invaluable in vitro model of differentiation. 1 In the placenta, trophoblast stem (TS) cells give rise to the highly specialized differentiated trophoblast cells. 2 While TS cells were first isolated from the mouse placenta, 2 they have subsequently been isolated from the human placenta as well as the rat, bovine, and non-human primates.3–5 The study of mTS cells has significantly informed much of what is known about trophoblast proliferation and differentiation. While cannabinoids have been shown to affect function in differentiated human trophoblast cells,6–8 little is known about whether cannabinoid exposure impacts the TS cell and its differentiation potential.
Placental health stands as a crucial determinant in fetal development, serving as the primary interface between maternal influences and the developing fetus. 9 Cannabis use in pregnancy is increasing, and knowledge about how it impacts placenta development is limited.10,11 Notably, cannabinoids readily cross the placental barrier.12–14 Thus, it is possible that the impact of cannabinoid exposure on the fetus is due to a direct effect on the fetus or exerted indirectly via the placenta. 11 Delta-9 tetrahydrocannabinol (Δ9-THC), the most abundant cannabinoid in cannabis, 15 is an exogenous analog of the endocannabinoid anandamide 16 and is responsible for many of the pharmacological actions of cannabis, including its antiemetic activities. 17 Using a preclinical rodent model, we previously reported that maternal Δ9-THC exposure during pregnancy led to compromised vascular branching in the placental labyrinth layer and growth-restricted offspring. 18
Trophoblast cells, unique to the placenta, are crucial for placenta development and establish a barrier between the maternal and fetal blood. 19 In cultured human trophoblast cells, Δ9-THC interferes with turnover and differentiation. 20 However, the mechanism of action has not been elucidated. Well established, however, is that the development and function of the placenta depends on a tightly regulated endocannabinoid system (ECS), orchestrating the proliferation, differentiation, invasion, and apoptosis of trophoblast cells.21,22 The ECS consists of endocannabinoids, endo/cannabinoid receptors, and associated proteins regulating the synthesis, transport, and degradation of endocannabinoids.21,23 Endocannabinoids (anandamide [AEA] and 2-arachidonoylglycerol) are endogenous lipids that bind and activate endocannabinoid receptors (cannabinoid receptor 1 [CB1/Cnr1] and cannabinoid receptor 2 [CB2/Cnr2]). 23 Like endocannabinoids, exogenous cannabinoids also bind CB1 and CB2 receptors, 24 with Δ9-THC acting as a partial agonist at both.11,17,25 Interestingly, the mouse (m) TS cells that give rise to the differentiated trophoblast cells express CB1 but not CB2. 26 Mouse knockout (KO) models and human trophoblast studies have underscored the pivotal role of CB1 signaling in trophoblast invasion, migration, and differentiation, apoptosis, mitochondrial function, and placental layer development (reviewed in 11 ).
Sustaining a stem cell pool and enabling their long-term self-renewal are critical for the maintenance and regeneration potential of a tissue.27,28 Thus, understanding whether Δ9-THC exposure affects the self-renewal capacity and/or the maintenance of TS cells is important to understanding how exposure can impact placental development. In the mouse placenta, CB1 expression has only been reported in the junctional zone. 26 Due to the Δ9-THC exposure-induced changes within the placental labyrinth, 18 we questioned whether (1) CB1 expression may be more extensive than previously reported or (2) Δ9-THC may alter the differentiation of mTS cells to labyrinth trophoblast subtypes via its action at CB1. Because high levels of Δ9-THC can cause CB1 desensitization in some tissues, 29 this study also aimed to assess the effect of physiologically and higher than physiologically relevant Δ9-THC exposure on undifferentiated mTS cells and their differentiation potential using wild-type (WT) and previously-derived Cnr1-KOmTS cells, 26 and further to characterize CB1 expression in the human, rat, and mouse placenta.
Materials and Methods
Placenta preparation for histological assessment of CB1 expression
Animals caged under standard conditions (12-h light/12-h dark cycle) had ad libitum access to water and food. Mouse work was conducted per the University of California, San Diego (UCSD) guidelines for animal models in research, with user protocols and experiments reviewed and approved by the Institutional Animal Care and Use Committee. Rat work was conducted per the Canadian Council on Animal Care guidelines with approval from the Western University Animal Care Committee; 3–6 placentae (1 placenta/dam) were stained to ensure that expression patterns were similar across samples.
Mouse placentae
Adult female and adult male C57BL/6 mice (12 weeks; Envigo) were mated and inspected the following morning for evidence of a copulatory plug, which was considered embryonic day (E) 0.5. Pregnant dams were euthanized by cervical dislocation at E10.5 and E18.5. Placentae were dissected, fixed in fresh 4% paraformaldehyde (pH 6.9), and either processed for embedding in paraffin18,30–33 or Optimal Cutting Temperature compound.34,35
Rat placentae
Pregnant female Wistar rats (Charles River, QC, Canada), shipped at E3.0, were euthanized at E19.5 using an overdose of pentobarbital (100 mg/kg i.p.), followed by decapitation. Placentae were dissected, fixed, and processed for paraffin embedding as described above.
Human placentae
Human term placental samples were sourced from the repository of placental tissues within the Biorepository at the UCSD Center for Perinatal Discovery and generously gifted from the Parast Lab (UCSD). All participating patients provided informed consent for the acquisition of tissue samples that were then deidentified. Tissue samples were processed employing established protocols within the UCSD Department of Pathology for fixation of tissue in neutral-buffered formalin, followed by processing and paraffin embedding. Sections from five normal placentae were assessed to ensure that staining patterns in villi were similar across samples.
Immunohistochemistry (IHC)
IHC analysis (5 μm paraffin sections), per the manufacturer’s protocol (ImmPRESSHorse Anti-Rabbit IgG Kit; Vector Laboratories), included antigen retrieval (Citra Buffer; BioGenex) in a 2100-Retriever (Electron Microscopy Science). CB1 antibody (Abcam, ab23703; 1:50) was visualized using Dako DAB with hematoxylin counterstain. Omission of the primary antibody served as the negative control.
In situ hybridization (ISH) by RNAscope®
ISH/RNAScope (10 μm cryosections), per the manufacturer’s protocol (RNAScope® Multiplex Fluorescent Reagent Kit v2 assay; Advanced Cell Diagnostics), used target or control probes: negative control (kit supplied), positive control (kit supplied), Cnr1 (Mm-Cnr1-C4), Prl8a8 (Mm-Prl8a8-C2). The probe signal was developed using Opal Dyes, diluted at 1:750 (Opal 690/Cnr1, Opal570/Prl8a8a), counterstained with DAPI, and mounted. Slides were scanned (M7000; Life Technologies), capturing regional images of the placenta. Comparative analysis was performed with IHC images to confirm that CB1 and Cnr1 expression matched.
mTS cell culture
WT mTS (mTSWT) cells (previously derived in our laboratory) 32 and mTS cells with the CB1 gene, Cnr1, knocked out (mTS Cnr1 KO; previously derived and generously gifted by the Sun laboratory) 26 were used for all experiments and maintained and differentiated as previously described.30–32,35 The rationale for using mouse TS cells was that existing mTS Cnr1 KO cells 36 were available and the Cnr1−/−placentae characterized, 26 allowing for results to be interpreted relative to in vivo findings. mTS cells were cultured in proliferating conditions (2 days) or in differentiating conditions (2, 4, and 6 days).30,31 As cell culture has less biological variability than tissue or animal studies, 37 a minimum of three independent biological replicates was selected for the experimental design. Experiments were seeded at 2.5 × 103 cells/cm2 and repeated to generate biological replicates (n = 3).
Cells were treated with a vehicle control [VEH, 0.1% (v/v) ethanol] or Δ9-THC [0.75, 1.5, or 15 μM, in a final concentration of 0.1% (v/v) ethanol; Cayman Chemicals CAS 192-08-03; ≥95%]. The Δ9-THC concentrations were selected to (1) be within the physiological range of measured plasma concentrations reported in human38–42 and rat 43 studies (60–200 ng/mL; ∼0.7 μM at the high end) and (2) represent concentrations within the range used in trophoblast studies.20,44 0.75 μM was considered the physiologically relevant concentration.38–43 Both 1.5 and 15 μM treatments were included because (1) trophoblast cells can tolerate higher concentrations of Δ9-THC without a significant impact on viability 20 and (2) high dose and repeated Δ9-THC exposure has been reported to cause CB1 desensitization. 29 Therefore, including higher concentrations in the experimental design allowed for the assessment of whether all concentrations produced the same effect, or whether concentrations greater than those found in maternal serum have an incremental effect or no effect at all. This is of importance as not all exposure in pregnancy is a result of the same delivery method, Δ9-THC concentration, or frequency of use. As mTS Cnr1 KO cells did not adhere to the plates as well as the mTSWT cells, culture experiments were replicated using gelatin-coated plastic (0.1% gelatin in water, Stem Cell Technologies).
RNA, cDNA, and quantitative reverse transcription polymerase chain reaction
Total RNA was isolated (Aurum micro spin columns, Bio-Rad) per the manufacturer’s protocol from each biological sample, and reverse transcribed (iScript RT Kit, Bio-Rad) with quantitative reverse transcription polymerase chain reaction (qRT-PCR) performed (in technical triplicate) with iTaq Universal SYBR Green Supermix (Bio-Rad) in a QuantStudio5 thermocycler (ThermoFisher). Commercially available primers (Bio-Rad, USA; meeting MIQE guidelines,45,46 see 47 for efficiency and validation criteria; primer efficiency determined and provided by Bio-Rad to fall within 90%–110%) included markers of undifferentiated TS cells,32,35,48,49 Eomes (qMmuCID0009039)30,31; Sca-1 (Ly6a; qMmuCED0003761)30–32 ; Esrrb (qMmuCED0039638) 30,32; Elf5 (qMmuCID000610); Fabp5 (qMmuCED0004811); labyrinth progenitor marker, 50 Epcam; (qMmuCID0017186)30–32 ; differentiated labyrinth trophoblast subtypes,51–53 syncytiotrophoblast layer II SynT-II, Gcm1 (qMmuCID0023712)30–32 ; syncytiotrophoblast layer I (SynT-I), Syna (qMmuCED0003216);30,32 sinusoidal trophoblast giant cell (S-TGC), Ctsq (qMmuCID0022572)31,32; junctional zone progenitor 54 Ascl2 (qMmuCED0001620)30,31; differentiated junctional zone trophoblast subtypes, 55 Tpbpa (qMmuCID0007168)30–32 ; spongiotrophoblast (Sp-T), 55 Prl8a8 (qMmuCID0008489); 31 glycogen trophoblast (GlyT), 56 Adlh1a3 (qMmuCED0045366) 30 ; spongiotrophoblast (Sp-T), trophoblast giant cells53,55 (P-TGC) Prl2c2 (qMmuCID0061688)30–32 ; and canonical 11 Cnr1 (qMmuCED0037834), Cnr2 (qMmuCED0038254), and non-canonical cannabinoid receptors 11 Gpr55 (qMmuCID0007644), Trpv1 (qMmuCID0005370), Pparg (qMmuCID0018821). All primer sets used were tested for one distinct melt peak, matching the details provided by the manufacturer. Data analysis utilized the comparative cycle threshold (ΔΔCt) method with Ppia (qMmuCED0041303) and Ywhaz (qMmuCED0027504) used as reference genes.30–32 Reference genes were selected for use in mTS cells following a previous analysis using a reference gene panel (Reference Genes M96, 10029130; Bio-Rad, USA), identifying Ppia and Ywhaz as stably expressed in both proliferating and differentiating TS cell cultures. The relative quantity of the target genes was calculated using the formula 2−ΔΔCT (QuantStudio 5 software; Life Technologies).
Statistical analysis
Analysis used Prism 9 software, with results from proliferating experiments analyzed by one-way ANOVA and differentiating experiments analyzed by 2-way ANOVA to assess (1) how Δ9-THC affected gene expression and (2) how days in culture affected gene expression. Where significant interaction was identified, post-hoc analysis was performed using Tukey’s multiple comparisons with a single pooled variance with a multiplicity-adjusted P value for each comparison. Significance was identified as a P value or adjusted P value ≤ 0.05, with data expressed as mean ± SEM.
Results
CB1/Cnr1 localized to trophoblast cells in direct contact with maternal blood in mouse, rat, and human placentae
Spatiotemporal CB1/Cnr1 assessment in the mouse confirmed CB1 expression in the Gly-T and the Sp-T of the junctional zone and identified expression throughout the labyrinth layer, in the S-TGCs (identified based on their contact with maternal blood and the size of their nuclei31,53,57) and in the interhaemal space (Fig. 1A–C). In the rat placenta, like that of the mouse, positive CB1 staining was identified in the junctional zone and labyrinth layer, with the CB1-positive TGCs lining maternal blood spaces (Fig. 2A). CB1 staining was confirmed in human term placentae, with CB1-positive villous SynT cells (Fig. 2B), demonstrating that maternal blood-facing trophoblast in all three species express CB1.
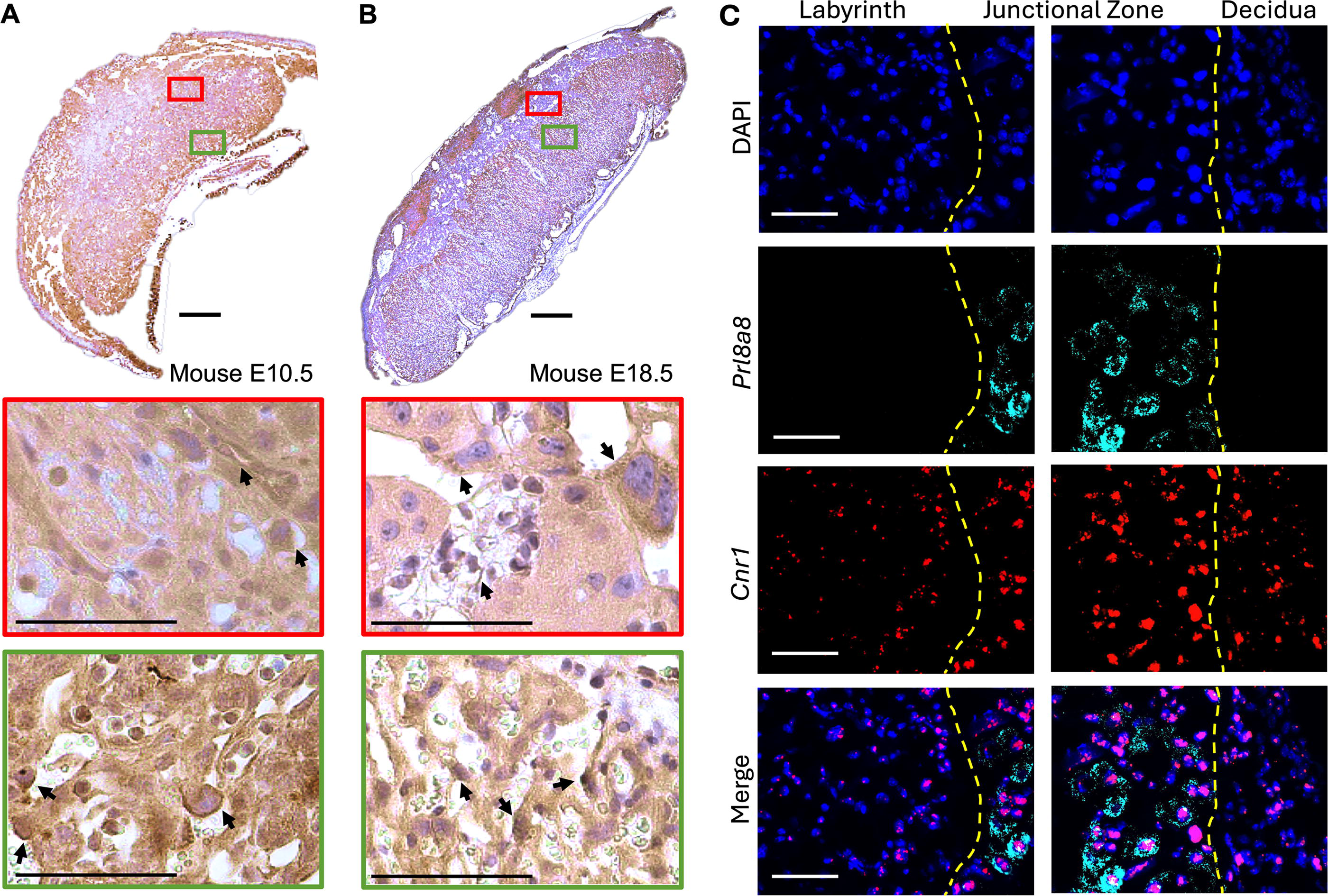
Spatiotemporal CB1 and Cnr1 staining in mouse placenta.
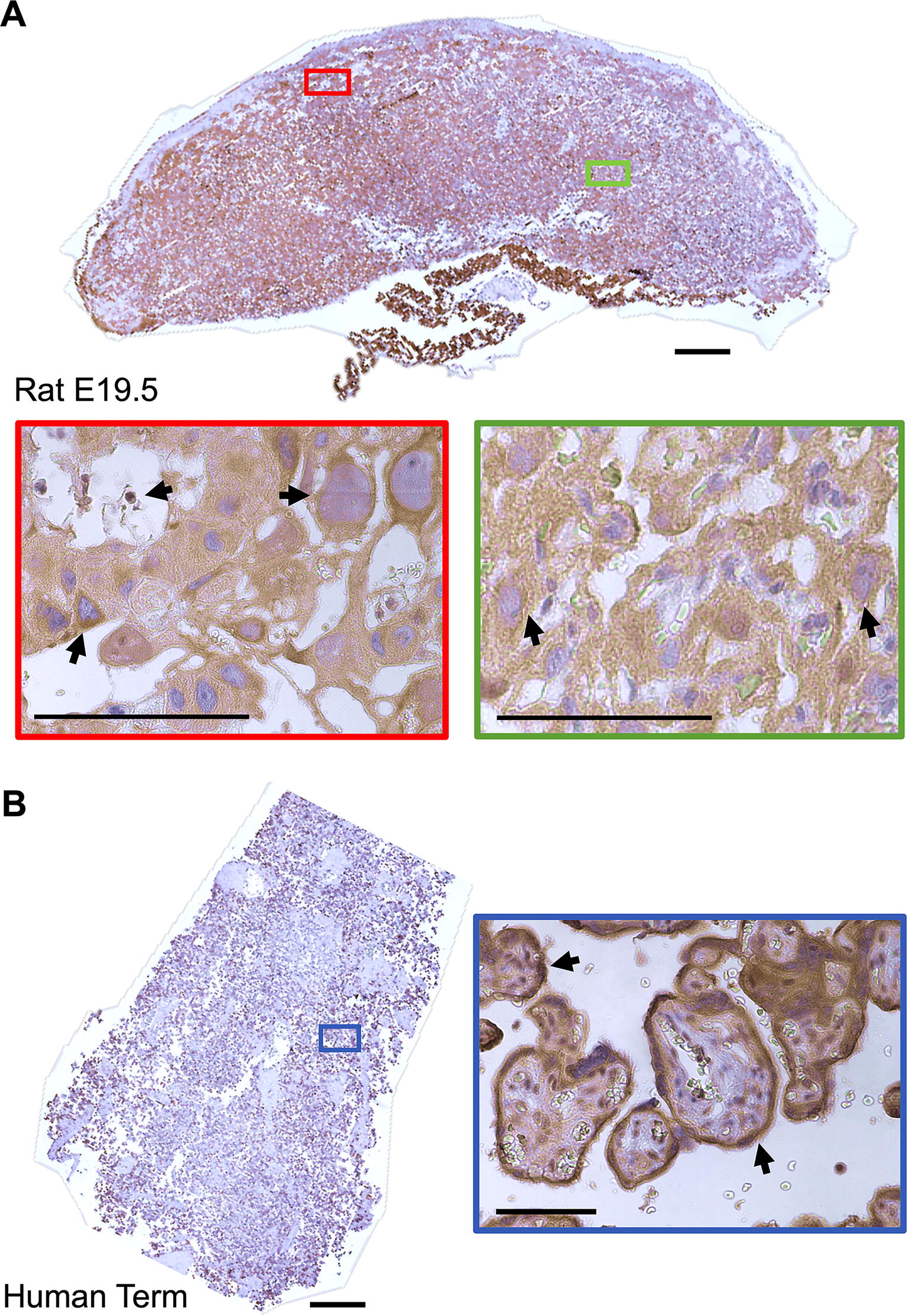
Spatiotemporal CB1 staining in rat and human placentae.
In the absence of Cnr1, mTS cells express Cnr2
Given that Δ9-THC can also signal through non-canonical receptors (Pparg, Trpv1 and Gpr55), their expression was assessed in undifferentiated mTSWT and mTS Cnr1 KO cells. As previously reported, mTSWT cells expressed canonical receptor Cnr1 but not Cnr2 (Fig. 3A). 26 Cnr2 was detected in the mTS Cnr1 KO cells (Fig. 3A), suggesting that its expression may be compensatory for the absence of Cnr1. Both mTSWT and mTS Cnr1 KO cells expressed the non-canonical receptor Pparg but not Trpv1 or Gpr55 (Fig. 4A).
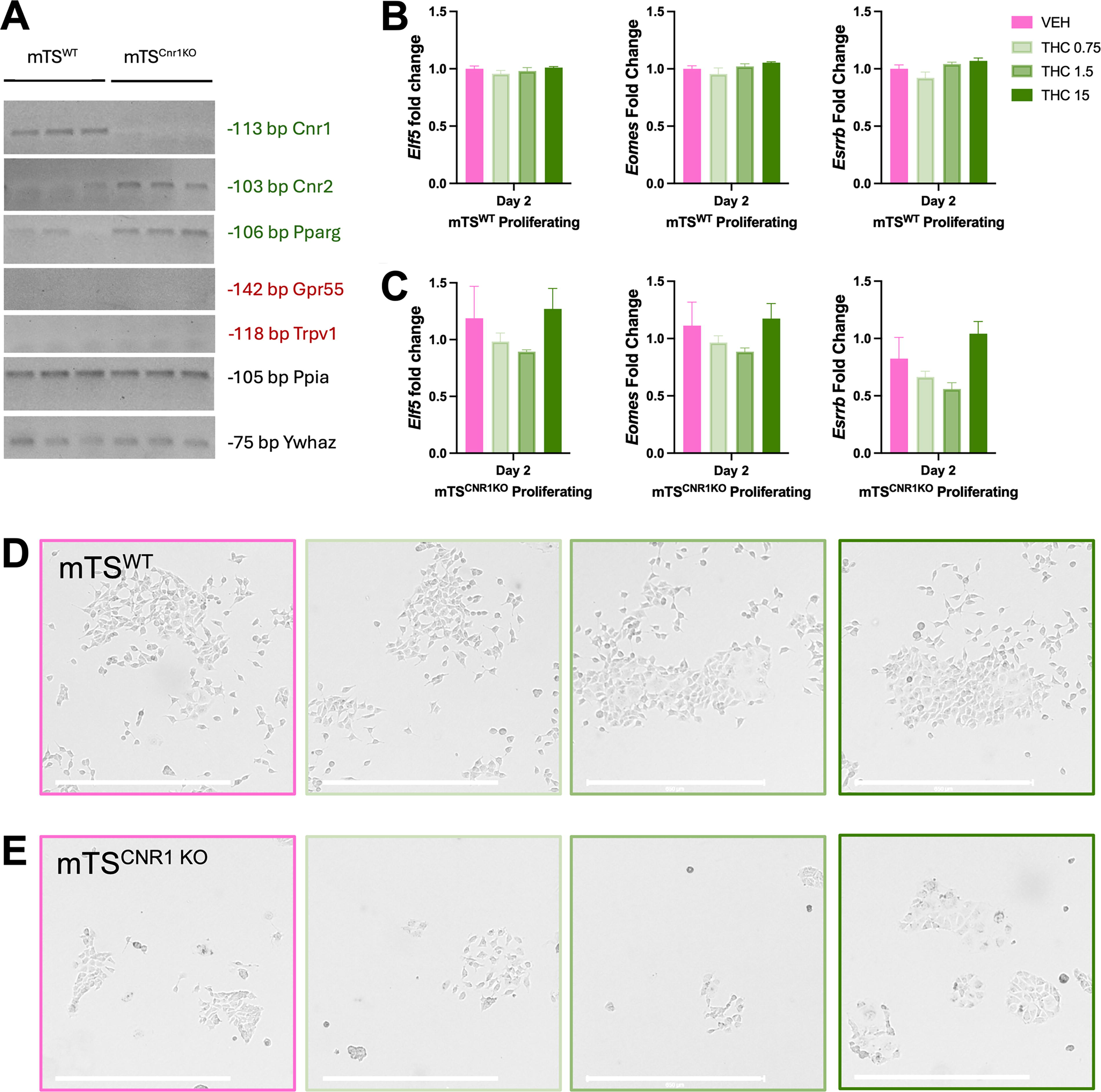
Δ9-THC treatment of mTSWT and mTS
Cnr1
KO in proliferating conditions.
qRT-PCR analysis of markers of undifferentiated TS cells, labyrinth progenitors, and syncytiotrophoblast.
Δ9-THC did not alter colony formation or expression of TS cell markers in undifferentiated TS cells
The maintenance of undifferentiated mTS cells requires growth factors, enabling their self-renewal. 2 As various physiological and toxicological factors disrupt this capacity and can lead to stem cell survival with diminished growth or compensatory and prioritized differentiation, 58 the effect of Δ9-THC exposure on mTS cell self-renewal, colony formation, and the expression of mTS cell markers was evaluated. In both mTS cell populations, Δ9-THC (all concentrations) did not change their expression of mTS cell markers (Fig. 3B and C), their capacity for self-renewal, colony formation, or cell morphology (Fig. 3D and E). As previously reported, mTS Cnr1 KO cells did not self-renew at the same rate as mTSWT cells, 26 resulting in smaller, less confluent mTS Cnr1 KO colonies (Fig. 3D and E).
Absence of CNR1 alters cell morphology and adherence
Growth factor removal induces mTS cell differentiation along two lineages:59,60 (1) junctional zone and trophoblast giant cells and (2) syncytiotrophoblast cells (SynTI and II). 61 Differentiating mTSWT and mTS Cnr1 KO cells were exposed to a VEH or one of three concentrations of Δ9-THC. Cultures were morphologically assessed (Supplementary Fig. S1), with the expression of the genes associated with differentiated trophoblast assessed by qRT-PCR (Figs. 4 and 5).
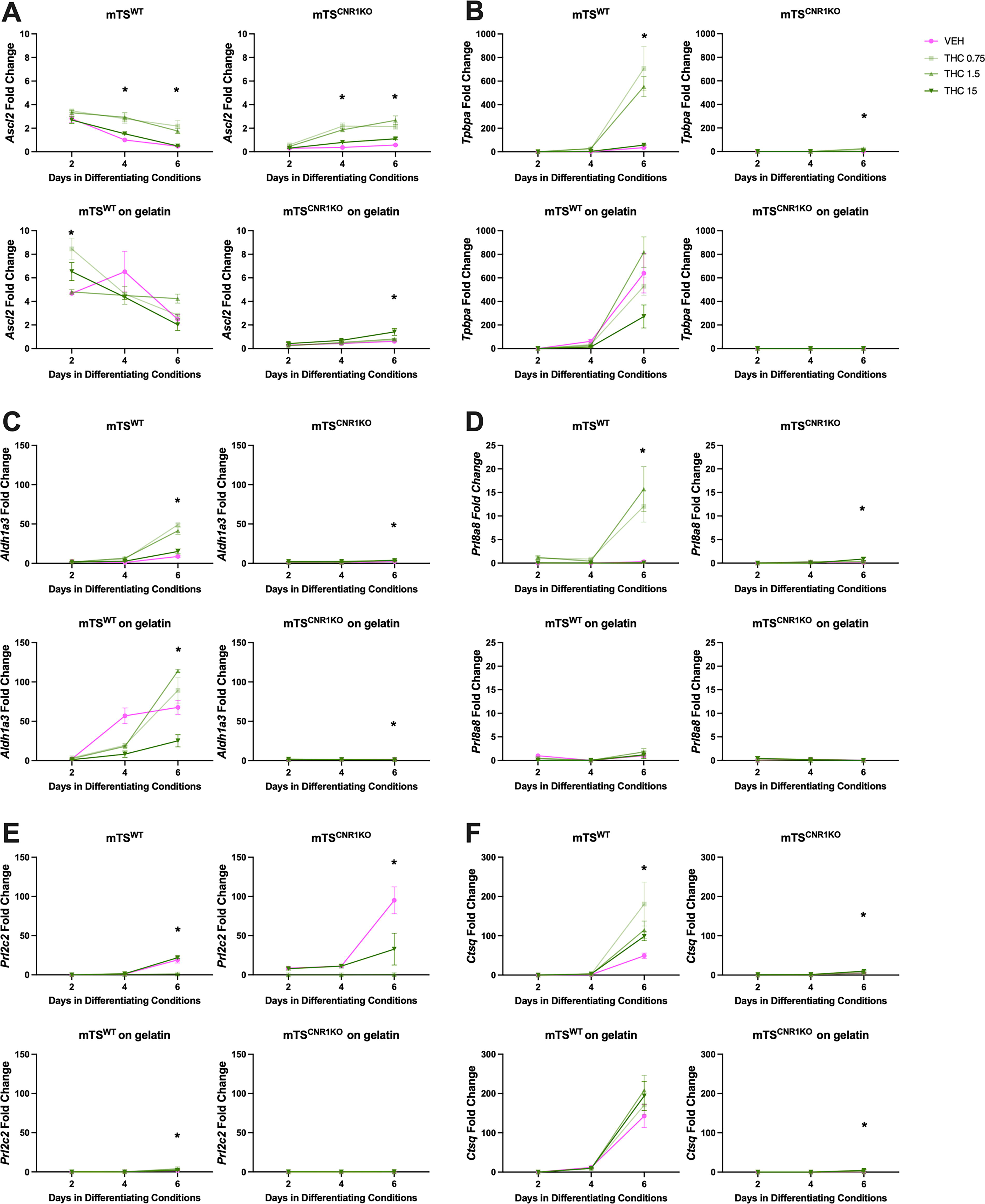
qRT-PCR analysis of markers of the junctional zone and trophoblast giant cell markers.
mTSWT vs mTSCnr1KO cells
Differentiating mTSWT cells were more confluent over 6 days than differentiating mTS Cnr1 KO cells (Supplementary Fig. S1A and B), with the differentiating VEH mTS Cnr1 KO cells less firmly adhered and a less differentiated cell morphology (small colonies and cuboidal in appearance; Supplementary Fig. S1B). Interestingly, differentiating mTS Cnr1 KO cells treated with Δ9-THC were less susceptible to adherence issues. The experiment was repeated on gelatin-coated plates to ensure differences in differentiation were not due to poor adherence on tissue culture plastic (Supplementary Fig. S1C and D). On gelatin-coated plastic, day 6 differentiated mTSWT cells were more confluent than those on plastic (Compare Supplementary Fig. S1A with C), while differentiating mTS Cnr1 KO cells had improved adherence but not confluence (Compare Supplementary Fig. S1B with D). Cell morphology of both cell types appeared flatter and more spread out (Supplementary Fig. S1A–D) on gelatin-coated plastic (day 6).
Δ9-THC vs VEH treatment
VEH-treated differentiating mTSWT cells on plastic or gelatin-coated plastic appeared more confluent (days 4 and 6) when compared with the Δ9-THC-treated groups (Supplementary Fig. S1A and C), though morphology was not obviously altered. Δ9-THC treatments did not alter the morphology or confluence of differentiating mTS Cnr1 KO cells on plastic or gelatin-coated plastic on days 2 and 4 (Supplementary Fig. S1B and D), but on day 6, confluence on plastic was increased (all Δ9-THC concentrations; Supplementary Fig. S1B). Given that the cells did not grow at equal rates, the effect of Δ9-THC on the expression of differentiation markers was interpreted independently for mTSWT and mTS Cnr1 KO cells.
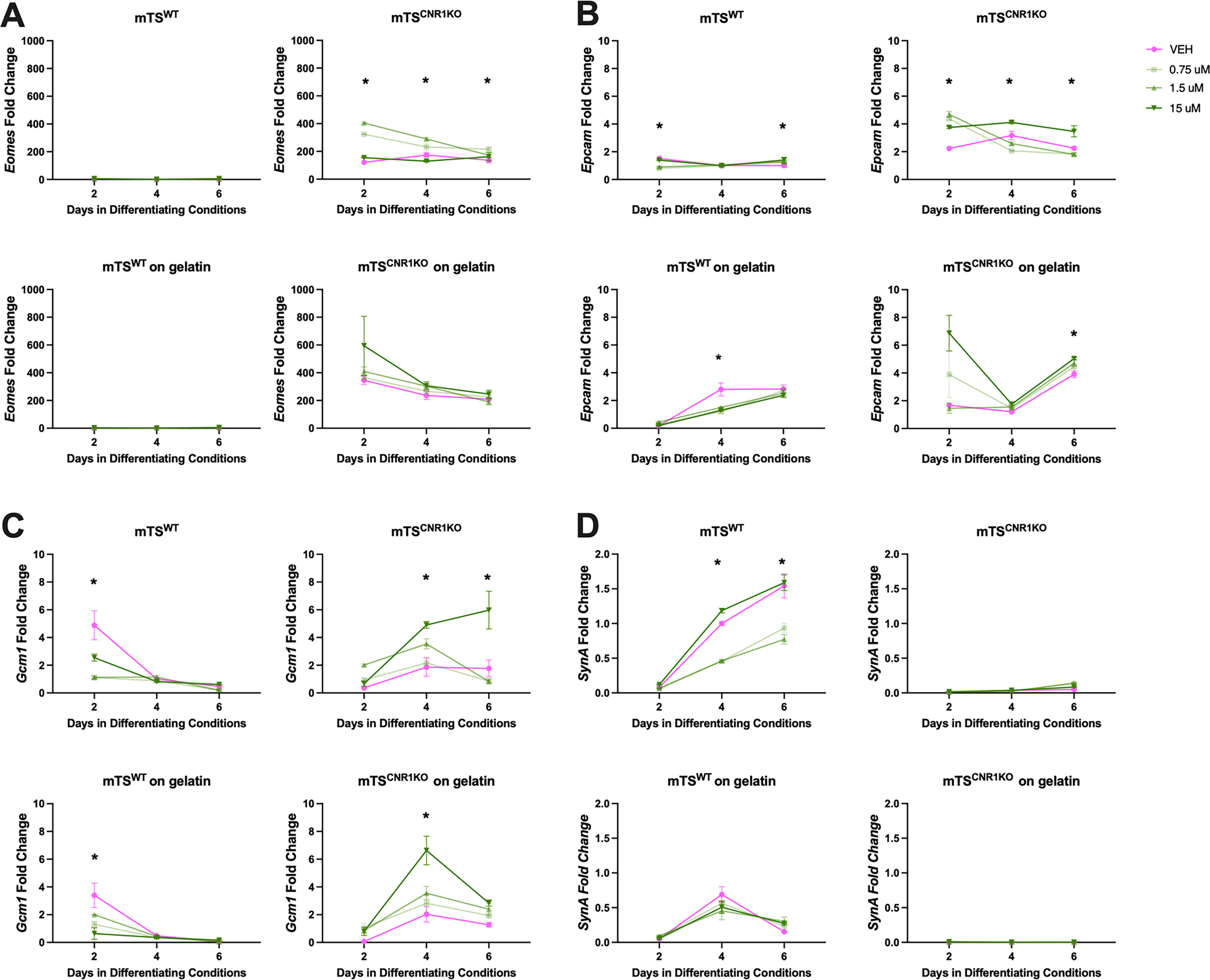
Differentiating VEH- and Δ9-THC-treated mTSWT cells expressed virtually no Eomes on plastic or gelatin-coated plastic, indicating their differentiation 35 (Fig. 4A). On plastic, differentiating VEH-treated mTSWT cells expressed markers of differentiated trophoblast in patterns as previously published,30,32,35 with labyrinth progenitor Epcam and SynT-II Gcm1 peaked on day 2, SynT-I SynA elevated on days 4 and 6 (Fig. 4B–D), junctional zone progenitor Ascl2 on day 2, Gly-T Aldh1a3, 62 TGC Prl2c2, S-TGC Ctsq and junctional zone Tpbpa on day 6 (Fig. 5A–C, E, and F) and levels of Sp-T Prl8a8 low with no observable peak of expression (Fig. 5D). On gelatin-coated plastic, differentiating VEH-treated mTSWT cells exhibited a similar expression of SynT-II Gcm1, Sp-T Prl8a8 and S-TGC Ctsq as those on plastic (Figs. 4C and 5D,F). Notable changes in the VEH group included elevated labyrinth progenitor Epcam expression on days 4 and 6, SynT-I SynA peaked on day 4, but rather than being maintained, its expression dropped off (Fig. 4B and D). Junctional zone Tpbpa and TGC Prl2c2 both similarly peaked on day 6, but Tpbpa expression was 12 times higher on gelatin, while Prl2c2 was reduced (10x) when compared with plastic (Fig. 5B and E, respectively). Junctional zone progenitor Ascl2 expression was elevated (6x) over plastic, while GlyT Aldh1a3 expression was elevated (25x) over plastic (Fig. 5A and C, respectively).
On plastic, Δ9-THC appeared to inhibit SynT differentiation pathways, with all concentrations reducing peak expression of SynT-II marker Gcm1 (Fig. 4C) and the two lower concentrations (0.75 and 1.5 µM) reducing peak SynT-I marker SynA and TGC marker Prl2c2. Conversely, Δ9-THC promoted junctional zone pathways with increased junctional zone Ascl2 and Tpbpa, GlyT Aldh1a3 and SpT Prl8a8 (Figs. 4 and 5). S-TGC marker Ctsq was only elevated by the lowest concentration of Δ9-THC (Fig. 5F). On gelatin-coated plastic, the two lower Δ9-THC concentrations reduced expression of SynT-II marker Gcm1 (Fig. 4C) and TGC Prl2c2 (Fig. 5E) and increased junctional zone progenitor Ascl2 (day 2; Fig. 5A) and GlyT Aldh1a3 (day 6; Fig. 5C). No Δ9-THC concentration affected the expression of SynT-I SynA, junctional zone Tpbpa, SpT Prl8a8 or S-TGC Ctsq on gelatin (Figs. 4D and 5B, D, and F).
15µM Δ9-THC treatment increased the expression of genes associated with labyrinth progenitor and SynT-II differentiation in mTSCnr1KO cells
Differentiating VEH-treated mTS Cnr1 KO cells on plastic and gelatin-coated plastic had high expression levels of Eomes, labyrinth progenitor and SynT-II markers Epcam and Gcm1 (Fig. 4A–C) and junctional zone progenitor marker Ascl2 (Fig. 5A). They had very low levels/no expression of most markers of SynT-I cells (Fig. 4D) or junctional zone populations (Fig. 5B–D). Together, these findings supported the morphological observations that mTS Cnr1 KO cells were less differentiated after 6 days. On plastic, VEH-treated mTS Cnr1 KO cells expressed TGC marker Prl2c2 (day 6), but not on gelatin-coated plastic. Culturing differentiating VEH-treated mTS Cnr1 KO cells on gelatin-coated plastic did not improve the expression of markers of terminal differentiation (Figs. 4C and 5B–D and F) but promoted the expression of labyrinth progenitor marker Epcam (day 6, compared with plastic). Increased Epcam coincided with reduced TS cell marker Eomes and TGC marker Prl2c2 (Figs. 4A,B and 5E), suggesting that the improved adherence on gelatin promoted more differentiation.
Despite the morphological differences between the mTSWT and the mTS Cnr1 KO cells, the effect of Δ9-THC treatment on mTS Cnr1 KO differentiation was assessed, considering that the undifferentiated cells expressed Cnr2 rather than Cnr1. On plastic, mTS Cnr1 KO cells remained largely undifferentiated when treated with low-dose Δ9-THC (0.75 and 1.5 µM). Specifically, expression of stem Eomes (all days), progenitor Epcam (day 2) and Ascl2 (Day 4 and 6) were elevated (Figs. 4A, B and 5A), with very low levels or no expression of markers of terminally differentiated trophoblast. Conversely, high-concentration Δ9-THC (15 µM) most significantly affected mTS Cnr1 KO cells, with evidence of differentiation towards the SynT-II pathway, increased expression of labyrinth progenitor marker Epcam and SynT-II marker Gcm1 (Fig. 4B,C). Furthermore, while expression remained low, the same concentration increased expression of Sp-T marker Prl8a8 and the S-TGC marker Ctsq (Fig. 5D and F). All Δ9-THC concentrations reduced the expression of TGC marker Prl2c2 (Fig. 5E). Culturing the mTS Cnr1 KO cells on gelatin-coated plastic and treating with Δ9-THC (15 µM) yielded similar results to those on plastic, with increased labyrinth progenitor marker Epcam and SynT-II marker Gcm1 (Fig. 4B and C), although the timing of expression was different, with Epcam elevated on day 6 and Gcm1 on day 4 (Fig. 4B and C). Similarly, while expression levels were low, Δ9-THC (15 µM) elevated expression of junctional zone progenitor marker Ascl2 and S-TGC marker Ctsq (Fig. 5A and F). Lower doses of Δ9-THC had a limited effect on differentiation: Δ9-THC (1.5 µM) did not significantly alter the expression of any markers of differentiation, and Δ9-THC (0.75 µM) only altered Aldh1a3 expression, which, while low, was elevated (Fig. 5C).
As trophoblast cells differentiate towards the junctional zone and TGC lineage, Cnr1 expression was reduced, and Pparg and Cnr2 expression were increased
With the expression of canonical and non-canonical receptors in undifferentiated TS cells defined and the spatiotemporal CB1 staining revealed, we investigated the expression of other cannabinoid receptors (Cnr2, Pparg, Trpv1, and Gpr55) in differentiating trophoblast. The results showed an inverse relationship between Pparg and Cnr1 (compare Fig. 6A with B) in differentiating mTSWT cells. Relative to the mTSWT, the mTS Cnr1 KO cells had very low levels of Pparg (Fig. 6B). Both populations had increased expression of Cnr2 as days in differentiating conditions increased (Fig. 6B). Neither population expressed measurable levels of Trpv1 or Gpr55 (Fig. 6C). Interestingly, as the mTSWT cells differentiated towards the junctional zone and TGC lineages (day 6), Cnr1 expression was reduced, and Pparg and Cnr2 expression were increased (Compare Fig. 5 with Fig. 6B).
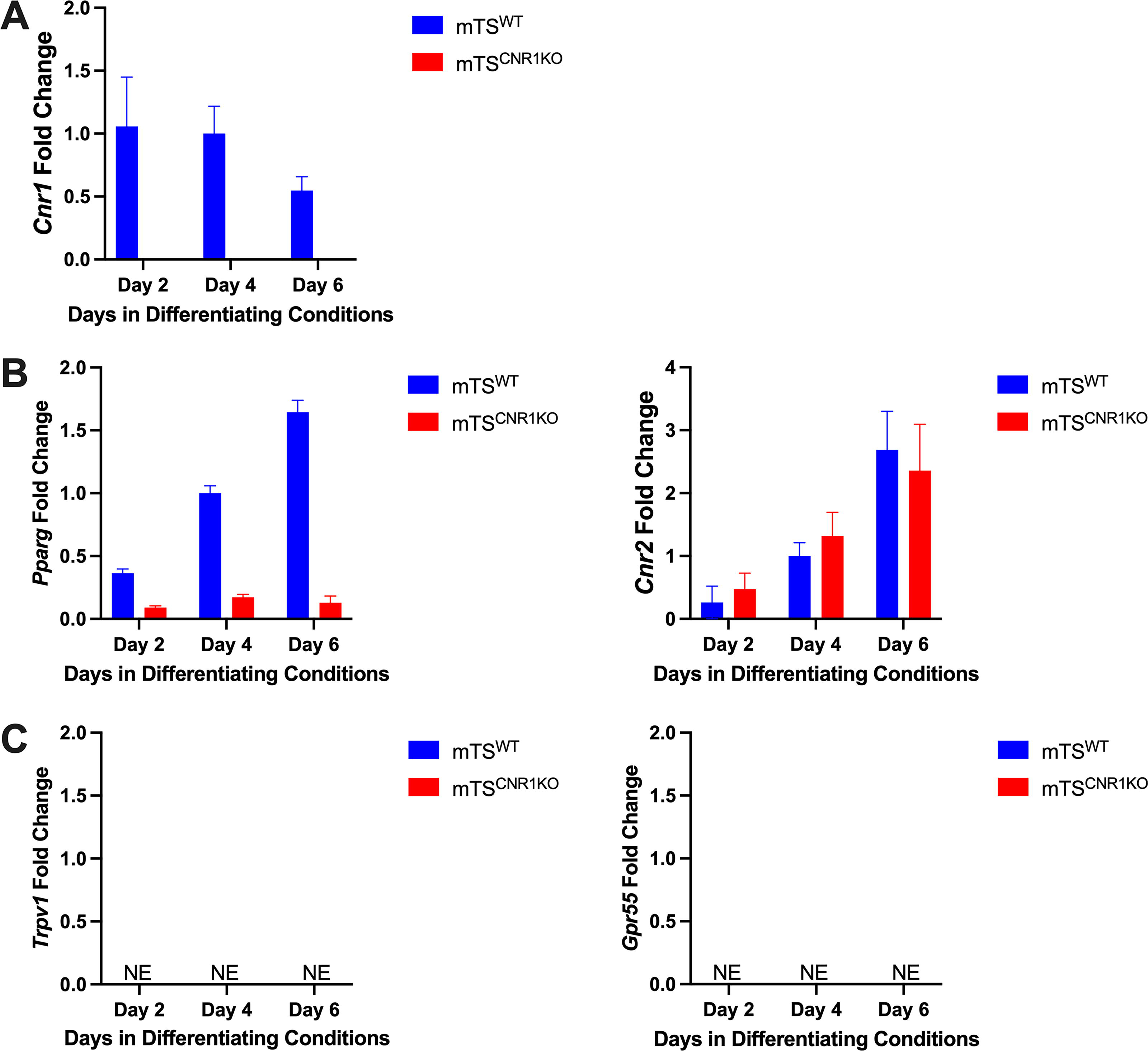
qRT-PCR analysis of cannabinoid receptors.
Discussion
It is well established that the ECS is critical to trophoblast differentiation and healthy placental development,26,36,63–68 with evidence that exogenous cannabinoid exposure during pregnancy impacts placental development and fetal growth.18,33,69–71 Stem cells are undifferentiated cells with the capacity to self-renew (creating more stem cells) and to differentiate into the specialized cells that enable organ function. 72 While the dynamics of how human (h)TS cells contribute to the human placenta throughout gestation has not been established, we have previously shown using an in vivo lineage tracing model that Eomes-expressing mTS cells contribute to the trophoblast cells of both layers, 35 demonstrating that in vitro differentiation recapitulates the lineage differentiation occurring in vivo. While Δ9-THC has been shown in vitro to impact trophoblast function,7,20,73 its effect on TS cells has not been reported. With cannabinoid exposure during pregnancy on the rise,74–76 it was imperative to elucidate which, if any, cell populations in the placenta are vulnerable to physiologically relevant concentrations of cannabinoids. Thus, to gain a better understanding of the placental response to cannabinoid exposure, we undertook an analysis of mTS cell Δ9-THC exposure, including a concentration of Δ9-THC (0.75 µM) in a range with that reported in maternal serum,38–43 a concentration 2x that found in maternal serum and 20× the concentration (as previously used for many of the functional trophoblast studies20,44) as elevated and repeated Δ9-THC exposure can contribute to receptor desensitization. 29 To easily visualize results, a summary figure representing the mTS cell findings is presented in Figure 7.
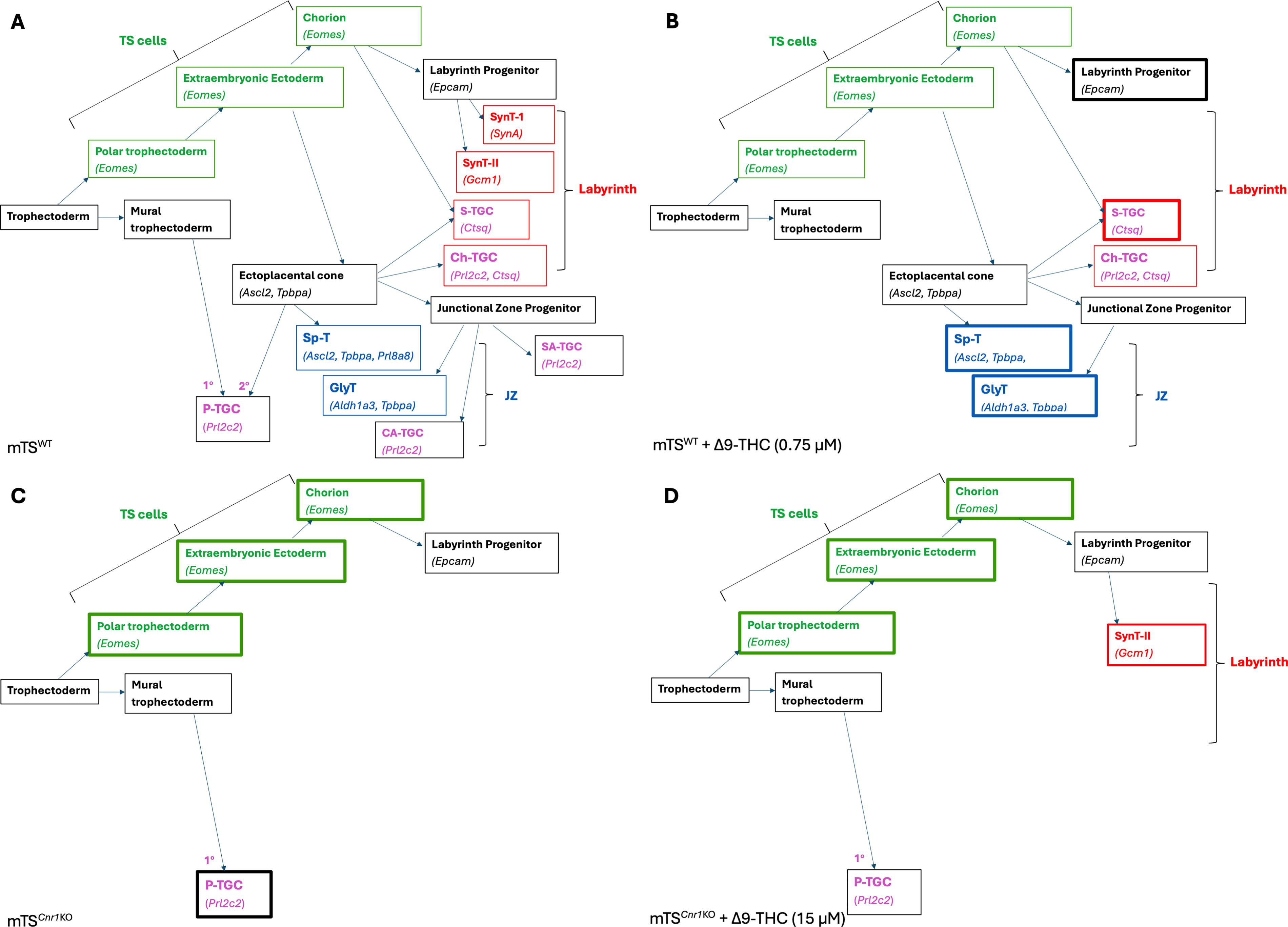
Overview of study findings. Differentiation potential of both WT and Cnr1KO mTS cells, treated with and without Δ9-THC.
While analysis of cannabinoid exposure is ideally studied in human pregnancies, pre-clinical animal studies allow for the study of isolated cannabinoids, reduce confounding factors, and mid-gestation placental analysis. In the human placenta, it has been shown that the endo/cannabinoid receptor CB1 is expressed in the trophoblast population in direct contact with maternal blood, enabling a direct effect of cannabinoids on this population. In the rodent placenta, however, its expression has only been reported in the junctional zone, 26 despite labyrinth-specific pathology in a rat in utero Δ9-THC-exposure model. 18 As such, updated spatiotemporal expression of CB1 in rodent placentae is critical for elucidating the signaling pathways that underlie the pathology. With the antibody in the original study no longer available, 26 this study aimed to update the spatiotemporal expression of CB1 in rodent placentae and found that it is expressed throughout the rodent placenta, with CB1/Cnr1-positive cells within the interhaemal space. Importantly, like the human placenta, the trophoblast cells in direct contact with maternal blood are CB1/Cnr1 positive.
Δ9-THC, like its analogous endogenous cannabinoid AEA, 16 is a partial CB1 agonist. 17 AEA acts on trophoblast cells, inhibiting proliferation and impairing syncytiotrophoblast-related protein synthesis and differentiation in BeWo64,66 and primary human trophoblast cells. 20 Studies using human cells show that Δ9-THC exposure similarly impacts differentiation to SynT. 8 Results from this study build on those findings, with Gcm1 expression reduced ∼fivefold in mTSWT cells at both the physiologically relevant (0.75 μM) and 1.5 μM Δ9-THC concentrations. Expression was reduced by half at 15 μM, suggesting that receptor desensitization may be occurring at the higher concentration. While this study used mTS cells, it is worth considering that GCM1 is a transcription factor crucial for proper placentation in mice but is also highly expressed in human SynT cells. 77 While mouse and human SynT cells are not identically located, in both, GCM1 is considered a master regulator of SynT formation. 77 Specifically, in mouse Gcm1-deficient placentae, chorionic trophoblast cells do not fuse to form SynT, 78 with similar results reported in hTS cell studies where GCM1 has been knocked down. 77 Thus, we believe our finding showing that Δ9-THC impairs Gcm1 expression in differentiating mTS cells is likely relevant to hTS cells. Supporting this possibility, THC exposure has been shown to reduce SynT fusion and associated GCM-1 expression in a BeWo SynT model at a 20 μM concentration. 8 Still, it is unknown whether there would have been a more significant effect in the lower concentration range reported in maternal serum.38–43 Furthermore, in the mouse, Gcm1 initiates chorionic branching morphogenesis, and its absence leads to a block in the branching of the chorioallantoic interface.78,79 As such, diminished GCM1 expression in placentae exposed to in utero Δ9-THC has the potential to impact the development of the placental vasculature significantly.
The core mechanism of SynT fusion involves the syncytin proteins in both human and mouse placenta, though the specific proteins are distinct. 80 Our results show that SynA, the gene encoding syncytin A, was (like Gcm-1) reduced by Δ9-THC exposure. It was reduced by half in both the physiologically relevant and the 2x concentration, but expression was not altered at the 20x concentration. While this current study did not examine the syncytialization/fusion process, our results align with a BeWo study demonstrating that Δ9-THC-inhibited fusion was associated with reduced expression of the gene encoding human Syncytin-1. 8 Together with findings from others, these results suggest that Δ9-THC directly affects the expression of genes associated with SynT differentiation and the fusion process in both species. Thus, SynT studies using human isolated trophoblast populations or cell lines may have a greater impact using concentrations close to those found in maternal serum.38–43 A further consideration of Δ9-THC exposure affecting GCM1 expression in the human placenta is that it has also been shown to regulate extravillous trophoblast (EVT) development and function using a hTS cell model. 77 Since Gcm-1 expression is limited to the labyrinth layer in mice, this aspect of trophoblast differentiation would need to be examined using human cells and models.
CB1 is a pivotal receptor within the ECS, and mouse KO models underscore its role in trophoblast differentiation and placental development,11,21,26 with reduced weight and trophoblast proliferation in CB1−/−placentae. 26 In this study, while vehicle-treated mTSWT cells expressed markers associated with SynT on day 2–4 of differentiation, vehicle-treated mTS Cnr1 KO did not. SynT markers were not assessed in the published CB1−/−placenta study. 26 Still, as resorptions were greatest at E12.5–14.5 26 (the window of branching angiogenesis), it is tempting to speculate that expression of SynT markers was similarly compromised in vivo. We showed that the undifferentiated mTSWT population expresses Cnr1 and Pparg, while the Cnr1KO population expresses Cnr2 and Pparg. As PPARγ activation promotes SynT formation 81 and Δ9-THC exposure in the mTSWT cells inhibited the expression of SynT-associated markers, we speculate that signaling Δ9-THC occurred via CB1. We further speculate that at 15 µM Δ9-THC, either receptor desensitization occurred, or that Δ9-THC activated both CB1 and SynT-promoting PPARγ. Interestingly, it was the highest concentration of Δ9-THC (15 µM) that induced expression of Gcm1 in the absence of Cnr1, with no expression of SynA at any concentration, including the vehicle control. This would suggest that while the mTS Cnr1 KO cells express Gcm1, they do not progress along the pathway to Syn-T differentiation to express syncytins. However, it begs the question of why it was the highest concentration that induced Gcm1 expression. mTS Cnr1 KO cells expressed Pparg and, unlike mTSWT cells, expressed Cnr2; thus, it is possible that the highest Δ9-THC concentration enabled CB2 signaling and/or activated PPARγ (activation increases Gcm1 expression 82 and SynT differentiation 81 ) to promote Gcm1 expression.
Interestingly, unlike genes associated with SynT expression, the physiologically relevant (0.75 µM) and 2x Δ9-THC concentrations elevated the expression of genes associated with several junctional zone and giant cell subtypes in mTS cells. Differentiation of mTS cells upon the withdrawal of stemness factors (FGF4 and TGF-β family members) typically drives differentiation to a mixed lineage of trophoblast cells. 2 To drive specific differentiation towards junctional zone subtypes would require suppressing SynT lineage by suppressing SynT-promoting PPARγ and Wnt signaling.83–85 Thus, these results suggest that the action of these two Δ9-THC concentrations suppress the SynT lineage in favor of junctional zone and giant cells differentiation. Further supporting this possibility is that at the highest concentration, expression of Gcm-1 was only reduced by half, and correspondingly, the increase in junctional zone markers was not as notable. As the absence or suppression of PPARγ signaling would be required for this action, we speculate that these effects of 0.75 and 1.5 µM Δ9-THC concentrations are mediated via action at Cnr1 in mTS cells. Unlike the other junctional zone and giant cell markers, expression of Prl2c2 was reduced, suggesting that differentiation to all TGC subtypes except for S-TGC cells was inhibited. When taken together with the Gcm1 results, it is noteworthy that these lower Δ9-THC concentrations reduced the expression of genes associated with early differentiation, specifically Prl2c2, expressed in parietal (p)TGC cells (the first trophoblast to differentiate) 53 and Gcm1, expressed in the progenitors of the SynT-II in the chorion, prior to chorioallantoic attachment. 78 Together with evidence that animal models with early Δ9-THC exposure (before E5) have increased spontaneous fetal loss, 86 it is tempting to speculate that physiologically relevant Δ9-THC concentrations may significantly impact early gestation, despite limited studies investigating pregnancy loss and cannabis use in human pregnancy. These findings provide impetus for exploring windows of cannabinoid exposure, as morning sickness (an often-cited reason for cannabis exposure 76 ) frequently occurs early in pregnancy.
In differentiating WT mTS cell populations, the response to the Δ9-THC concentrations was not linear. Perhaps most significant is that a number of the effects in mTSWT were mediated at the lowest Δ9-THC concentration. Δ9-THC has been shown to produce biphasic, dose-dependent effects (reviewed in ref. 11 ). Whether this is due to endocannabinoid competition at the receptor, signaling through multiple receptor pathways or receptor desensitization 11 at higher concentrations was not explored. Briefly, the CB1 receptor, a G protein-coupled receptor, primarily couples with Gi/o proteins or recruits both β-arrestin1 and β-arrestin2. 87 It also couples (to a lesser degree) with Gs, Gq/11, and G12/13 proteins. 87 All coupling and recruitment lead to ERK1/2 phosphorylation, which is induced by independent downstream signaling events stemming from the G-protein pathways. 87 However, independent of ERK1/2 phosphorylation, β-arrestin1 and β-arrestin2 recruitment also lead to CB1 desensitization and downregulation (reviewed in ref. 87 ) and may explain the differences observed between concentrations in the mTSWT cells.
A limitation to this study was that, as in previous studies, differentiating mTS Cnr1 KO cells displayed slowed growth, dramatic morphological changes, and adherence issues when compared with their WT counterparts. 26 While their adherence was moderately improved by both gelatin and Δ9-THC exposure, the slowed growth and morphological differences were not. We propose that altered adherence may underlie the observed compromised proliferation and self-renewal. Specifically, cell division requires the duplication of cellular structure with polar segregation, followed by the physical separation of the parent cell. Evidence suggests that cell cycle, cell adhesion, and cytoarchitecture are significantly intertwined (reviewed in 88 ). Thus, the decreased cell adhesion and confluence could signify compromised adhesion signaling. Worthy of consideration is that CB1 activates focal adhesion kinase (FAK)89,90 in many cell populations via CB1/G12/13 protein coupling. 87 FAK regulates cytoskeletal organization, gene expression, and cell behavior, such as cell spreading and migration. 91 As such, in the absence of CB1, FAK activation may be reduced or inhibited, contributing to poor attachment and limited cell spread. Supporting this possibility is that when the mTS Cnr1 KO cells were cultured on gelatin (which activates FAK 92 ) adherence was ameliorated. Furthermore, FAK plays a role in regulating cell cycle progression.93–95 As MacPhee et al have demonstrated that FAK is a key mediator of trophoblast proliferation and differentiation during early pregnancy, 96 it is tempting to speculate that CB1 activation of FAK may be necessary for mTS cell adhesion and self-renewal. In contrast, Δ9-THC treatment improved mTS Cnr1 KO cell confluence and adhesion, suggesting that Δ9-THC, acting via a different receptor (perhaps CB2 or PPARγ), may directly or indirectly ameliorate aberrant cell adhesion. While correlating Cnr1KO and WT mTS cell differentiation results to interpret Δ9-THC signaling was not possible (due to the adherence differences), we believe that the Δ9-THC exposure of the mTS Cnr1 KO cells demonstrated that Δ9-THC can signal via other receptors in mTS cells, and that this signaling promotes adhesion. Further, with the continued expansion and focus on organoid studies, these results highlight the importance of adhesion in trophoblast differentiation and support a role for endocannabinoid signaling in TS cell interaction with the extracellular matrix.
In the first half of gestation, as the placenta expands, the trophoblast population is heterogeneous with both undifferentiated and differentiating/differentiated trophoblast cells.32,35,97 In the latter half of gestation, the TS cell population is significantly reduced, with large populations of differentiated trophoblast cells.32,97 Results from this study show that Δ9-THC exposure had little impact on the status and colony-forming potential of the undifferentiated mTS cells. Several stem cell populations report no effect of Δ9-THC on colony-forming potential, but report increased proliferation, even at nanomolar concentrations. 98 Interestingly, adipose and bone marrow-derived stem cells 99 had increased proliferation, while stem cell populations responsible for embryonic development responded to Δ9-THC differently: naïve pluripotent embryonic stem cells had increased proliferation, whereas epiblast-like cells in a primed pluripotent state 100 did not (regardless of Δ9-THC concentration). Fatty acid amide hydrolase (FAAH) controls the content and biological activity of N-arachidonoylethanolamide and other bioactive endocannabinoids. 101 Cnr1KO mTS cells have reduced cell growth, 26 and similarly, FAAH null TS cells present a similar, though less pronounced, reduction in cell growth, suggesting that TS cells may have endogenous endocannabinoid activity at CB1. Thus, it is possible that in proliferating mTS cells, Δ9-THC either does not alter the endocannabinoid activity, or it is simply CB1 activation (by either an endocannabinoid or Δ9-THC) and possible downstream FAK signaling that is important to mTS cell proliferation. The results from this study suggest that mTS cells in their undifferentiated state are less vulnerable to the effects of Δ9-THC. A factor to consider when translating these findings to an in vivo TS cell proliferation assessment model is that in addition to increased proliferation the TS cell population size can change due to compromised SynT differentiation, which is often associated with an accumulation of undifferentiated cells.78,102
This study is limited in that it did not assess the impact of Δ9-THC exposure on the functional properties of differentiated trophoblast, including transport of nutrients, metabolism or production of hormones and growth factors, among others. Further analysis is warranted, as functional changes, like compromised trophoblast differentiation, can also contribute to placental pathology. Furthermore, as Δ9-THC and cannabis exposure studies in animals reveal that exposure (i.p. or inhaled) compromises placental efficiency, with more grams of placental tissue per gram of fetal tissue,18,103 it underscores compromised differentiation and/or function as possible contributors to placental pathology and fetal development.
In summary, with the rate of cannabis use in pregnancy increasing and animal exposure studies supporting compromised placental development, we have demonstrated that Δ9-THC treatment impacts the differentiation of mTS cells at a dose matching that reported in maternal serum38–43 and 2x that concentration. These concentrations do not affect mTS cell status or colony-forming potential but reduce the expression of genes associated with SynT differentiation, while increasing the expression of markers associated with junctional zone differentiation in a CB1-dependent manner. These results make it tempting to speculate that Δ9-THC at the concentrations used in this study would have a limited effect on the self-renewing capacity of in vivo TS cells but a significant impact on their differentiation to SynT, potentially impacting branching angiogenesis. Thus, the results from this study suggest that Δ9-THC imparts a targeted disruption in placental development rather than a general toxicity to mTS cells. Additionally, our study demonstrates that CB1/Cnr1 expression in the rodent, like in the human, is localized in trophoblast cells in direct contact with maternal blood across gestation. Given the strong links between placental insufficiency, fetal growth restriction, and trophoblast differentiation, there is a great impetus to examine the effects of gestational Δ9-THC exposure on the placenta and its individual cell populations, with translation to human organoid models.
Footnotes
Acknowledgments
The authors wish to thank Dr. Xiaofei Sun for generously providing the mTS Cnr1 KO cell line, the members of the Natale Lab for participating in helpful discussions throughout this study, Hoejeong Kim for sectioning the slides used in RNAscope®, the Parast lab for the deidentified placenta samples, and the Department of Biomedical and Molecular Sciences at Queen’s University for generously sharing equipment.
Author Disclosure Statement
The authors declare no competing interests.
Funding Information
This work was supported by grants from the Canadian Institutes of Health Research (CIHR) to DRCN and DBH, the Canadian Foundation for Innovation to DRCN and the NIH/NICHD (1RO1HD100179-01) to DRCN.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.