
Abstract
Muse cells are endogenous pluripotent-like stem cells identified as stage-specific embryonic antigen-3 (SSEA-3)-positive subpopulations in the bone marrow, peripheral blood, and connective tissues of various organs. Clinical trials conducted by intravenous injection of donor-Muse cells, without the use of immunosuppressive drugs, have demonstrated safety and efficacy across multiple diseases. Since the epitope recognized by the anti-SSEA-3 antibody is a glycolipid, rather than a protein produced by a genetic code, the antibody may detect Muse cells across different species. Muse cells possess unique properties, including the ability to survive under stressful conditions, spontaneously turn into different cell types from all three primary layers of the body, and repair tissues in living organisms. They have been isolated from several mammalian species. However, their presence and characteristics in companion animals, such as canine and feline, remain unexplored, despite the growing demand for treatments that regenerate tissues in veterinary medicine. Adipose-derived stem cells (ADSCs) were established from adipose tissue taken during routine veterinary procedures. SSEA-3-positive cells were isolated using fluorescence-activated cell sorting. SSEA-3-positive cells were found in both canine (0.93 ± 0.16%) and feline (2.9 ± 0.15%) ADSCs, similar to human rates. Gene expression analysis revealed that SSEA-3-positive cells exhibited significantly higher levels of the pluripotency markers Oct3/4 and NANOG compared with SSEA-3-negative ADSCs. In suspension culture, SSEA-3-positive cells formed ES cell-like M-clusters. These cells could differentiate into endodermal (SOX17, AFP), mesodermal (GATA2, DESMIN, SMA), and ectodermal (NESTIN, NF) marker-positive cells, as measured by quantitative polymerase chain reaction and immunocytochemistry. These results show that canine and feline ADSCs contain SSEA-3-positive cells. These cells express pluripotency markers and can differentiate into endodermal, mesodermal, and ectodermal lineages. Their properties match those of Muse cells in humans and other mammals. This study offers basic evidence for isolating Muse cells from pets and demonstrates their potential for use in veterinary regenerative therapies.
Introduction
Multilineage-differentiating stress-enduring (Muse) cells, stress-tolerant endogenous pluripotent-like stem cells, are found in the bone marrow (BM), organ connective tissues, including those of the umbilical cord (UC) and amnion, and peripheral blood (∼0.01%∼0.2% of the mononuclear fraction) as pluripotent surface marker stage-specific embryonic antigen (SSEA)-3-positive.1–8 They are also found in cultured mesenchymal stem cells (MSCs) and fibroblasts as a subpopulation, corresponding to 1% to several percent of the total population. Different from SSEA-3 negative non-Muse MSCs, Muse cells express pluripotency markers, including Oct3/4, Nanog, Sox2, survive in the suspension culture and form Muse cell-derived clusters which are similar to embryonic stem (ES) cell-derived embryoid body, and are able to differentiate into endodermal-, mesodermal-, and ectodermal-lineage cells either spontaneously on the gelatin-coated dish or by cytokine-induced differentiation control.1,9,10
Preclinical studies have demonstrated that Muse cells outperform MSCs or non-Muse MSCs after intravenous injection (i.v.) in various models of organ damage. These include chronic kidney disease, 11 spinal cord injury, 12 stroke,13,14 liver damage, 15 myocardial infarction, 16 lung fibrosis, 17 aortic aneurysm, 18 peripheral arterial disease, 19 amyotrophic lateral sclerosis, 20 diabetic skin ulcer, 8 and osteochondral damage. 21 This is because Muse cells have unique abilities. They can differentiate into many cell types in vivo and home to damaged sites by sensing tissue damage signals, such as sphingosine-1-phosphate. Unlike other cells, they are not trapped in the lung after i.v. injection. Muse cells evade immune responses and stay in the target tissue for extended periods. They also provide lasting effects, such as reducing inflammation and apoptosis, protecting blood vessels, and preventing fibrosis. Non-Muse MSCs have limited differentiation capacity (mainly into osteocytes, chondrocytes, and adipocytes), are often trapped in the lungs, and provide only temporary bystander effects. 16
Clinical studies support these preclinical findings. Muse cells from HLA-mismatched donors have been given intravenously without immunosuppression in trials for ischemic stroke, 22 spinal cord injury, 23 neonatal hypoxic–ischemic encephalopathy, 24 acute myocardial infarction, 25 epidermolysis bullosa, 26 and amyotrophic lateral sclerosis. 27 All studies showed safety and therapeutic benefits for 1–1.5 years with no signs of immune rejection. In one double-blind, placebo-controlled trial for subacute ischemic stroke, about 70% of patients treated with donor Muse cell i.v. improved greatly. They went from being dependent or bedridden to regaining the ability to travel, eat, use public toilets, and sign documents on their own. These improvements persisted for 52 weeks. 22
SSEA-3, also called globoside-5, is a glycosphingolipid. It is composed of five carbohydrate units attached to a sphingolipid. 28 Since SSEA-3 is not a protein, it is not encoded by a gene. This lets it remain consistent across species. Researchers have isolated Muse cells using SSEA-3 from many mammals, including mice,29,30 rats, 31 rabbits, 16 swine,32,33 sheep,34,35 goats, 36 and horses. 37 These cells express pluripotency factors, differentiate into three germ layers, and show better therapeutic effects than MSCs in models such as rabbit myocardial infarction 16 and swine liver injury.32,33 Therefore, Muse cells with effects similar to those in humans can be isolated from many mammals using SSEA-3.
Canines and felines are among the most common pets worldwide, and demand for cell therapy is increasing. This study examined whether cells similar to human Muse cells can be isolated from their adipose tissue as SSEA-3-positive cells.
Materials and Methods
Animals
Canine and feline tissue was provided by the “Obstetric Services For Dogs” in Fukaya, Japan. Veterinarians performed all procedures with the owner’s consent for sample use in this study. Samples were treated according to Tohoku University’s animal care regulations. The Animal Care and Experimentation Committee of Tohoku University Graduate School of Medicine approved the experiments (permission No. 2019MdA-265-03).
Isolation and expansion of canine and feline adipose-derived mesenchymal stem cells
Adipose tissue was collected from the uterine broad ligament and subcutaneous tissue during contraceptive surgery for three canines and felines. The adipose tissue was minced thoroughly using sterile scissors in phosphate-buffered saline (PBS) at room temperature, and then enzymatically digested using 2 mg/mL collagenase/Dispase (Roche, Mannheim, Germany) in Dulbecco’s modified Eagle’s medium (DMEM, Invitrogen, Carlsbad, CA, USA) in a 37°C shaker for 3 h. The mixture was checked every 30 min until the tissue was digested, at which point no visible tissue pieces were observed. The mixture was slowly passed through a 100 µm filter into a 50 mL tube to remove tissue debris. The filtrate was further centrifuged at 300g for 10 min at 15°C, and the supernatant was discarded. The remaining pellet was seeded onto 100 mm dishes at density of 3.5 × 104 cells/cm2 in growth media consisting of DMEM supplemented with 10% fetal bovine serum (FBS; HyClone, Logan, UT, USA), 1% Kanamycin sulfate (ThermoFisher Scientific, Waltham, MA, USA), and 1% antibiotic antimycotic solution (MilliporeSigma, St Louis, MO, USA). Dishes were incubated at 37°C in a humidified incubator containing 5% CO2 for 2 days before changing media to 10% FBS/DMEM. At 80%–90% confluence, the cells were detached with a trypsin–EDTA solution (ThermoFisher Scientific) and subcultured repeatedly.
Collection of SSEA-3-positive cells from canine- and feline-ADSCs
Cells from passages five through seven were used for the isolation of SSEA-3-positive cells. SSEA-3 positive and SSEA-3 negative cells (Muse and non-Muse cells, respectively) were isolated by fluorescence-activated cell sorting (FACS; BD FACS Aria™II cell sorter, BD Biosciences [BD], San Jose, CA, USA), as described previously. 1 To isolate SSEA-3 positive cells, canine-ADSCs, and feline-ADSCs were incubated with mouse anti-SSEA-3 antibody (1:100, YKR-04, AlbaMedicine, Sendai, Japan) and stained with secondary antibody to fluorescein isothiocyanate (FITC)-conjugated anti-mouse IgM (1:100, Jackson ImmunoResearch Laboratories, West Grove, PA, USA).
Triploblastic differentiation of SSEA-3-positive cells in vitro
Culture plates were coated with poly-HEMA (2-hydroxyethyl methacrylate) (MilliporeSigma), which prevented cells from adhering to the bottom of the 96-well culture plates (ThermoFisher Scientific), and were air-dried overnight on a clean bench. The collected SSEA-3-positive cells were seeded at a density of 1 × 10^4 cells per well in a coated 96-well plate to form clusters. After 3 days, the clusters were replated onto gelatin-coated 4-well plates and expanded for an additional 7 days.
Immunocytochemistry of cells expanded from clusters
Cells were fixed with 4% paraformaldehyde (PFA; Millipore) in PBS for 1 h at 4°C and were subjected to immunocytochemistry as described previously [1]. In brief, samples were incubated overnight at 4°C with antibodies against α-fetoprotein (AFP, 1:500, ab75705, Abcam), neurofilament (NF, 1:500, ab1987, Abcam), and smooth muscle actin (SMA, 1:500, MS-113, Thermo). All antibodies were diluted in PBS/0.1% BSA solution. After washing with PBS three times, cells were incubated with secondary antibodies, either of Alexa488-conjugated anti-mouse IgG or anti-rabbit IgG (1:500; Jackson ImmunoResearch) for 2 h at room temperature (RT). Nuclear staining was performed using 4′,6-diamidino-2-phenylindole (DAPI; 1:500; Sigma-Aldrich). The cells were then inspected using a fluorescence microscope (BZ-X710, KEYENCE, Osaka, Japan).
Quantitative reverse transcriptase-polymerase chain reaction
Total RNA was collected using the NucleoSpin®RNA XS (Macherey–Nagel, Duren, Germany), and cDNA was synthesized using Oligo(dT) 20 primer (ThermoFisher Scientific) and SuperScript®IV reverse transcriptase (ThermoFisher Scientific). DNA was amplified with the Applied Biosystems 7500 Fast real-time PCR system according to the manufacturer’s instructions. Data were processed using the ΔΔCT method. Canine and feline pluripotent markers were evaluated using a Fast SYBR Green expression assay (ThermoFisher Scientific) with the following primers:
Canine Oct3/4-F: 5′-CTGAAGCAGAAGAGGATCAC-3′, Canine Oct3/4-R: 5′-GCCGCAGCTTACACATATTC-3′; Canine SOX2-F: 5′- AACCCCAAGATGCACAAСTС-3′, Canine SOX2-R: 5′-CGGGGCCGGTA TITATAATC-3′; Canine NANOG-F: 5′-CTAGGGACCCTTСTССAATG-3′, Canine NANOG-R: 5′-CTTCTGTTTCTTGCCCTGG-3′; Canine ACTB-F: 5′-CAGCAAATGTGGATCAGC-3′, Canine ACTB-R: 5′-ССААTСTCA TCTCGGTTT-3′.
Feline Oct3/4-F: 5′-CCAAACAACCATCTGCCGTT-3′, Feline Oct3/4-R: 5′-TGTCTCTGCTTTGCATATCTCCTG-3′; Feline SOX2-F: 5′-ACCAGCTCGCAGACCTACAT-3′, Feline SOX2-R: 5′-TGGAGTGGGAGGAAGAGGTA-3′; Feline NANOG-F: 5′-TCCAGTGCCTGAGATTGATGG-3′, Feline NANOG-R: 5′-AGGAAGGAAGAGGAGAGACGGT-3′; Feline ACTB-F: 5′-AGCCTTCCTTCCTGGGCAT-3′, Feline ACTB-R: 5′-AGGGCGGTGATCTCCTTCT-3′.
Canine and feline triploblastic differentiation markers were evaluated using the following primers:
Canine AFP-F: 5′-CTTTGGCTTGCCCACTATGGC-3′, Canine AFP-R: 5′-AACCGTTATGGCTCGGAAGGT-3′; Canine NESTIN-F: 5′-AGCCCTACTTCCCTCTCCTT-3′, Canine NESTIN-R: 5′-CTGAAGTGTGGGCGGGATGGGG-3′; Canine GATA-2-F: 5′-GCCACTGACCATGAAGAAGGAA-3′, Canine GATA-2-R: 5′-ACAGCTCCTCAAAGCACTCTGC-3′.
Feline SOX17-F: 5′-TGTTCGATTCTGGTTGCCCA-3′, Feline SOX17-R: 5′-AAACACACCCAAGGCGAAAG-3′; Feline NESTIN-F: 5′-AGTTTAGAGGCCAAAGGCGG-3′, Feline NESTIN-R: 5′-CTAAAGACCTCAAGGACCTCTGG-3′; Feline DESMIN-F: 5′-GGTACAAGTCAAAGGTGTCCGA-3′, Feline DESMIN-R: 5′-TTTCGCAGGTGTAGGACTGG-3′.
Statistics
The data were analyzed using one-way analysis of variance and an unpaired Student’s t-test to determine statistical significance, as performed in Microsoft Excel software. All data represent three independent experiments. The results are presented as the mean ± SE of the mean. A P value of less than 0.05 was considered significant.
Results
Isolation of canine- and feline-SSEA-3-positive cells from ADSCs
We established ADSCs from each of three canines and felines adipose tissues and isolated SSEA-3-positive cells using AlbaMedicine antibody. Representative FACS analysis revealed the presence of SSEA-3-positive cells in both canine (0.93 ± 0.16%) and feline (2.9 ± 0.15%) ADSCs (Fig. 1A). The SSEA-3 positivity in ADSCs from canine and feline was shown in Table 1.

Isolation and characterization of canine and feline SSEA-3-positive cells from ADSCs.
SSEA-3 Positivity Percentage in Canine and Feline Adipose-Derived Stem Cells

Adipose-derived stem cells (ADSCs) were obtained from the adipose tissues of three canines and three felines, and SSEA-3-positive cells were isolated using the AlbaMedicine antibody.
Pluripotency marker expression in canine- and feline-SSEA-3-positive cells
We analyzed the expression of pluripotency-associated genes in SSEA-3-positive cells and SSEA-3-negative ADSCs. In canine ADSCs, SSEA-3-positive cells exhibited significantly higher expression of Oct3/4 (P < 0.01) and Nanog (P < 0.05) compared with SSEA-3-negative ADSCs (Fig. 1B). Similarly, feline SSEA-3-positive cells showed significantly elevated expression of Oct3/4 (P < 0.01) and NANOG (P < 0.01) relative to SSEA-3-negative ADSCs (Fig. 1C). Although SOX2 expression tended to be higher in SSEA-3-positive cells from both species, there was no statistical significance (Fig. 1B,C).
In vitro triploblastic differentiation potency of canine- and feline-SSEA-3-positive cells
To evaluate the in vitro triploblastic differentiation capacity of SSEA-3-positive cells, canine and feline SSEA-3-positive cell clusters were generated in suspension culture (Fig. 2). After 3 days, the clusters were transferred onto gelatin-coated dishes and maintained under adherent conditions for an additional 7 days, during which proliferating cells expanded outward from each cluster. Cells derived from these canine and feline clusters were collected and subjected to qPCR analysis.

Morphology of canine and feline SSEA-3-positive cell clusters. Suspension culture of canine and feline SSEA-3-positive cells at day 3, showing the formation of M-clusters. Scale bars = 100 μm.
In canine cluster-derived cells, the expression of lineage markers for all three germ layers was detected, including AFP (endoderm; P < 0.01), NESTIN (ectoderm; P < 0.001), and GATA-2 (mesoderm; P < 0.001) (Fig. 3A). Immunocytochemistry further confirmed the presence of AFP-positive endodermal cells, SMA-positive mesodermal cells, and NF-positive ectodermal cells (Fig. 3B).

In vitro triploblastic differentiation of canine-SSEA-3 positive cells.
Similarly, feline cluster-derived cells exhibited significant expression of SOX17 (endoderm; P < 0.01), NESTIN (ectoderm; P < 0.001), and DESMIN (mesoderm; P < 0.001) (Fig. 4A). Consistent with the gene expression data, immunocytochemistry demonstrated AFP-, SMA-, and NF-positive cells in the differentiated cell population (Fig. 4B).
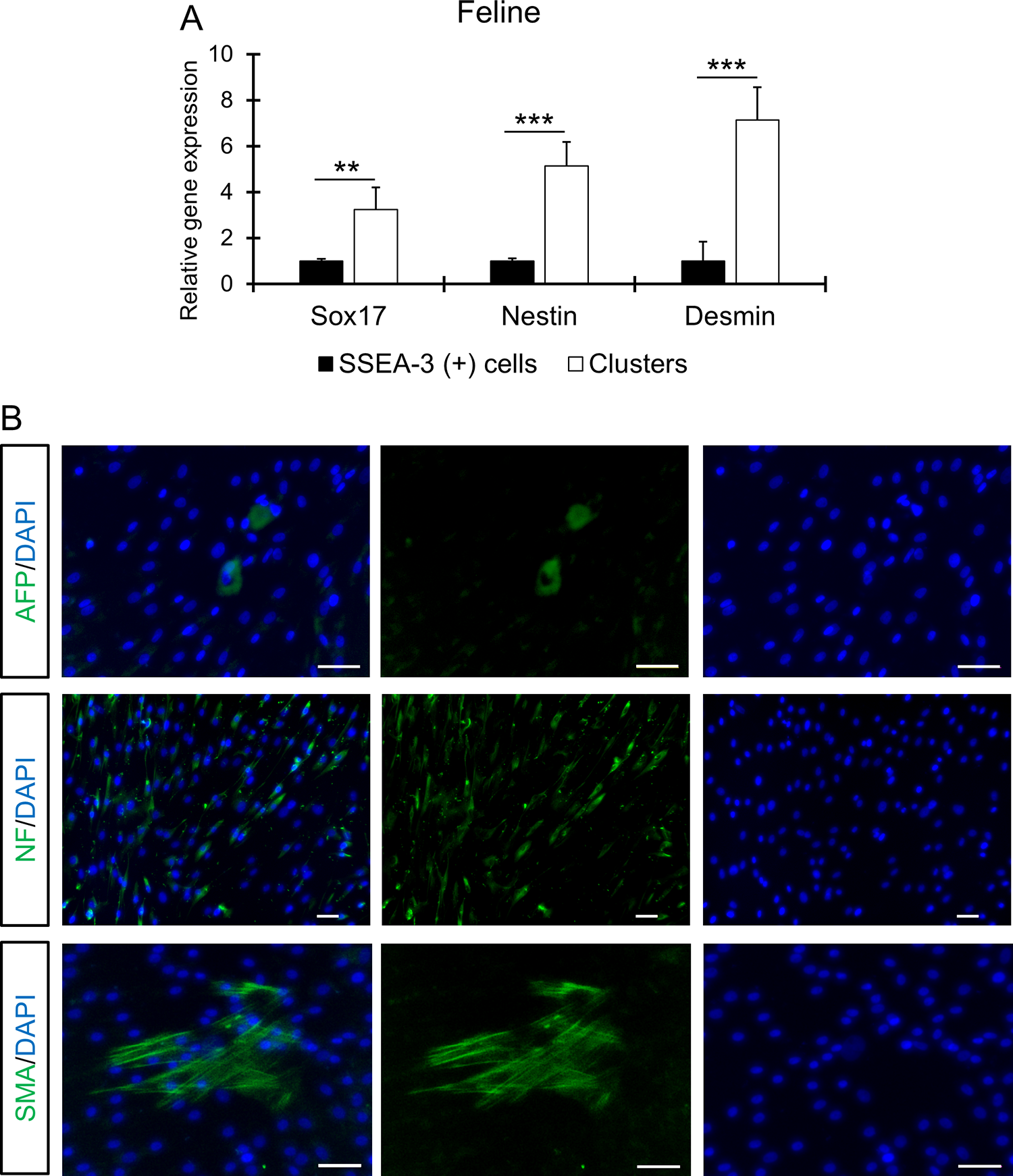
In vitro triploblastic differentiation of feline-SSEA-3 positive cells.
Discussion
In this study, we isolated SSEA-3-positive cells from ADSCs established from canine subcutaneous adipose tissue and feline adipose tissue obtained from the uterine broad ligament using AlbaMedicine antibody. The percentage of positive cells was similar to that in human ADSCs, ranging from 1% to 3%. 7 These SSEA-3-positive cells expressed pluripotent stem cell markers, such as Oct3/4, NANOG, and SOX2, at higher levels than those in SSEA-3-negative ADSCs. Similar to Muse cells from other species, including humans, they formed clusters resembling embryoid bodies derived from ES cells in suspension culture.1,2 When these clusters were transferred to gelatin-coated dishes, they spontaneously differentiated into cells positive for endodermal (AFP, SOX17), mesodermal (GATA-2, DESMIN, SMA), and ectodermal (NESTIN, NF) markers, without the need for cytokine differentiation inductions, as confirmed by qPCR and immunocytochemistry. All of these characters were similar to those of Muse cells reported in human,1,2 mouse, 29 rabbit, 16 and swine. 32
Muse cells have been isolated from various tissues, including the BM, 1 fat,7,8 UC,5,6 amnion, 3 and blood. 4 However, in pet animals, fat is considered more practical than the BM, which requires anesthesia due to the painful nature of BM aspiration, or the UC and amnion, which are difficult to collect aseptically during natural birth and are eaten after birth. Fat can be easily and aspetically collected from the subcutaneous and uterine broad ligament during castration, neutering, or cesarean section, and can be collected and used both autologously and allogenically. Particularly, autologous Muse cells make it suitable for use in pedigree animals where donor cells should not be introduced into the body.
Once ADSCs have been established, they can be expanded and cultured to select SSEA-3-positive Muse cells. In rodent models of cerebral infarction, 14 liver disease, 38 and aortic dissection, 12 a Muse cell rate of 70% or more (using techniques such as MACS) yielded therapeutic effects significantly superior to MSCs and/or non-Muse MSCs. The main administration route is generally i.v. No differentiation induction, gene transfer, or surgery is required. This suggests that practical treatments tailored to market needs can be provided.
In particular, allogenic Muse cells were administered without immunosuppressants in a rabbit acute myocardial infarction16,25 and a swine liver damage model.32,33 Notably, in the former model, they survived as cardiomyocyte marker-positive cells in the postinfarct tissue even after 6 months without immunosuppressants, and cardiac function recovery did not decline. Based on this, it is possible that allogenic Muse cell therapy without immunosuppressants may also be possible in canines and felines.
In a previous canine study, SSEA-1-positive cells, not SSEA-3, were found to increase after stress treatment, such as long-term trypsin treatment. 39 SSEA-1 and SSEA-3 are different molecules: SSEA-3 belongs to the Globo series, while SSEA-1 belongs to the Lacto series is defined as a carbohydrate antigen expressed on the surface of cells during early embryogenesis and mouse pluripotent stem cells. Further investigation is needed to determine whether SSEA-1-positive cells are equivalent to the Muse cells observed in humans and other animals, and whether SSEA-3-positive cells can be detected using another antibody, such as the AlbaMedicine antibody.
In this study, SSEA-3-positive cells, similar to those found in humans, were detected in canines and felines. Further investigation into their therapeutic efficacy and safety is required. In particular, the possibility of allogenic Muse cell treatment, including the requirement for an HLA matching test and immunosuppressant treatments, remains to be examined. Meanwhile, because both canine and feline are genetically diverse and may have undergone extensive hybridization, the yield and differences in the properties of Muse cells will need to be investigated.
Footnotes
Acknowledgments
The article publishing charge for this article was raised through a crowdfunding campaign. The author would like to deeply thank all 42 crowdfunders. This work did not receive any grant from funding agencies.
Disclosure Statement
MuseCell Innovations PTE Ltd (MCI, Singapore) holds exclusive licensing and patents for MuseCells.
Funding Information
No funding was received for this article.