
Abstract
Hematopoietic stem cell (HSC) transplantation is an established therapy for malignant and nonmalignant hematologic disorders; however, clinical application remains constrained by limited graft availability and challenges in maintaining stemness during ex vivo manipulation, as well as transplant-related complications. Accordingly, alternative strategies to generate hematopoietic-competent cells from accessible stem cell sources are being actively explored. This study investigated whether OCT3/4 overexpression enhances the hematopoietic trans-differentiation potential of dental tissue-derived mesenchymal stem cells (DMSCs) and whether the resulting HSC-like cells exert therapeutic effects in a cyclophosphamide-induced myelosuppressed mouse model following intra-femoral delivery. OCT3/4 was introduced into DMSCs using the Neon transfection system, and OCT3/4-overexpressing DMSCs (DMSCsOCT3/4+) were subsequently exposed to hematopoietic cytokines to induce an HSC-like phenotype. Cytokine-treated DMSCsOCT3/4+ exhibited a rounded morphology, increased expression of HSC-associated surface markers (CD34 and CD45), and upregulated hematopoietic transcription factors, including GATA2, C/EBPα, RUNX1, and SCL. The derived HSC-like cells (D-HSCs) were transplanted into the femoral bone marrow cavity of myelosuppressed mice, and therapeutic outcomes were assessed by complete blood counts and histological analyses. D-HSC transplantation was associated with recovery of bone marrow cellularity and partial restoration of spleen and thymus cellularity and size, accompanied by improvement in body weight and peripheral blood parameters compared with myelosuppressed controls. Collectively, these findings indicate that OCT3/4-enhanced DMSCs can be directed toward an HSC-like state under hematopoietic cues and that the resulting cells may support hematopoietic and immune recovery in myelosuppressed hosts, supporting their potential as an alternative, autologous cell source for hematopoietic regeneration.
Keywords
Introduction
Hematopoiesis is the process by which hematopoietic stem cells (HSCs) self-renew and differentiate into all blood cell lineages throughout life. 1 Although HSCs are valuable for treating hematologic and immune-related disorders, their clinical application remains limited by low cell yield, loss of stemness, and difficulty maintaining a pure HSC population during ex vivo expansion.2–5 While alternative stem cell sources have been explored to overcome these limitations,6,7 the teratoma-forming potential of pluripotent stem cells remains a major barrier to clinical application. 8 Therefore, nontumorigenic and clinically compatible stem cell sources for HSC generation are needed.
Mesenchymal stem cells (MSCs) are nonhematopoietic stem cells and multipotent adult stem cells of mesodermal origin that can be isolated from various tissues, including bone marrow, adipose tissue, and dental tissue.9,10 MSCs exhibit self-renewal, multilineage differentiation potential, immunomodulatory activity, and low tumorigenic risk, making them attractive for regenerative therapy.11,12 Various diseases, such as myeloid leukemia, anemia, and congenital neutropenia, have been treated with MSCs.13–15 Among MSCs derived from various tissues, dental tissue-derived MSCs (DMSCs) have attained the advantage of being easily obtained noninvasively from wisdom teeth extracted after dental treatment. 10 Therefore, DMSCs are considered among the most promising candidates for stem cell–based therapies.
Octamer-binding transcription factor 3/4 (OCT3/4) is a critical regulator of pluripotency that maintains the undifferentiated state and controls their self-renewal and differentiation capacity.16,17 Recent studies have highlighted the role of OCT3/4 not only in maintaining pluripotency but also in promoting hematopoietic lineage reprogramming.18,19 Notably, it was demonstrated that fibroblasts transduced with OCT3/4 and hematopoietic factors directly converted into multilineage hematopoietic progenitors without passing through a pluripotent state. 18 These findings suggest that OCT3/4, in the presence of hematopoietic cytokines, can facilitate the generation of HSC-like phenotypes from non-hematopoietic cells.
In the present study, OCT3/4-overexpressing DMSCs were induced toward hematopoietic differentiation using hematopoietic cytokines. The resulting cells were evaluated for HSC-like phenotypes in vitro and for therapeutic potential after transplantation into an alkylating agent-induced myelosuppressed mouse model.
Materials and Methods
Chemicals and media
All chemicals were purchased from Sigma (St. Louis, MO, USA) and media from Gibco (Gibco Life Technologies, Gaithersburg, MD, USA), unless otherwise specified.
Isolation of mesenchymal stem cells in dental tissue
Dental tissues were obtained with informed consent under approval from the Ethics Committee of Gyeongsang National University Hospital (GNUH-IRB-2018-11-002-001). DMSCs were isolated from the dental follicle tissue as previously described. 20 Briefly, tissue was chopped into 1 mm3 explants, digested in DPBS containing collagenase type I (1 mg/mL) at 37°C for 40 min, mechanically dissociated, and cultured in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine serum (FBS), 100 µg/mL streptomycin, and 100 IU/mL penicillin. The cells were cultured in an incubator at 37°C in a humidified atmosphere containing 5% CO2. At 70%–80% confluence, cells were passaged using 0.25% trypsin–ethylene-diamine-tetra-acetic acid (Trypsin-EDTA) used at passage 3.
Analysis of flow cytometry
DMSCs (1 × 105) were fixed in a 4% paraformaldehyde for 60 min at room temperature, washed by DPBS. The cells reacted with FITC-conjugated mouse antihuman CD34 (1:200; BD Biosciences, NJ, USA), FITC mouse antihuman CD45 (1:200; BD Biosciences), FITC mouse antihuman CD44 (1:200; BD Biosciences), FITC rat antihuman CD90 (1:200; BD Biosciences), and APC-conjugated mouse antihuman CD73 (1:200; BD Biosciences), APC mouse anti-human CD105 (1:200; BD Biosciences) for 60 min at room temperature. FITC mouse IgG1 (1:200; BD Biosciences) and APC mouse IgG1 (1:200; BD Biosciences) were used as isotype controls. After the reaction, the cells were washed with FACS buffer and analyzed using Flow-cytometry (BD FACS verse; Becton Dickson, NJ, USA). Data were analyzed by FlowJo v10 software.
Mesenchymal lineage differentiation
DMSCs were differentiated into osteoblasts, adipocytes, and chondroblasts for 21 days, as previously described. 20 Osteoblast differentiation was induced in DMEM containing 10% FBS, 50 μg/mL L-ascorbic acid 2-phosphate, 10 nM dexamethasone and 10 mM β -glycerophosphate. The osteoblasts differentiation was confirmed by Alizarin Red S. Adipogenic differentiation was induced in DMEM containing 10% FBS, 1 μM dexamethasone, 10 μM insulin, 100 μM indomethacin, and 500 μM isobutyl methylxanthine. Adipogenic differentiation was confirmed by Oil Red O. Chondrogenesis was induced in STEMPRO chondrogenesis media (STEMPRO chondrogenesis differentiation kit, Invitrogen, CA, US). Chondrocyte-differentiated cells were stained with 1% Alcian blue solution.
OCT3/4 plasmid transfection system
OCT3/4 gene overexpression was induced using Neon™ Transfection System 100 μL Kit (Invitrogen, MA, USA). DMSCs were seeded 1–2 days before transfection, resuspended in Buffer R at 1 × 107 cells/mL, and mixed with plasmid DNA (4 μg/106 cells). Electroporation was performed by setting neon transfection conditions with a single pulse at 1.7 KV/cm for 20 msec. DMSCs transfected with OCT3/4 (DMSCs OCT3/4+ ) were immediately replated and cultured for subsequent experiments.
Differentiation into HSC-like cells
DMSCs OCT3/4+ were differentiated into HSC-like cells (D-HSCs) as previously reported, with minor modifications.18,21 Briefly, cells were cultured on Matrigel-coated plates in DMEM/F-12 medium containing 10% knock out serum replacement, 1% L-glutamin, 1% non-essential amino acid, 0.1% β-mercaptoethanol supplemented with 150 ng/mL recombinant human stem cell factor (SCF, Peprotech, NJ, USA), 150 ng/mL recombinant human FMS-like tyrosine kinase 3 ligand (Flt3L, Peprotech), 20 ng/mL basic fibroblast growth factor (bFGF, R&D Systems, MN, USA), 30 ng/mL recombinant human insulin-like growth factor (IGF-II, Peprotech), 20 ng/mL granulocyte colony stimulating factor (GCSF, Peprotech), 50 ng/mL bone morphogenetic protein 4 (BMP4, R&D Systems) 20 ng/mL interleukin 3 (IL-3, Peprotech), and 20 ng/mL interleukin 6 (IL-6, Peprotech) for 20 days. CD45-positive DMSCs OCT3/4+ (CD45+ DMSCs OCT3/4+ ) were sorted by labeling CD45 microbead antibodies (Miltenyi Biotec, Germany). Sorted CD45+ DMSCs OCT3/4+ were seeded in low attachment cell culture plates and cultured in a hematopoietic medium containing 80% knockout DMEM, 20% FBS, 1% nonessential amino acid, 1% L-glutamin, and 0.1% β-mercaptoethanol supplemented with SCF, FLT3L, G-CSF, BMP-4, IL-3, and IL-6. The medium was replaced every 3 days, and morphological changes were monitored by a light microscope.
Isolation of CD45 positive DMSCsOCT 3 /4+
CD45+ DMSCs OCT3/4+ were isolated with the MACS magnetic separation kit (Miltenyi biotec) according to the manufacturer’s instructions. Briefly, harvested cells were labeled with the MicroBeads conjugated to monoclonal anti-human CD45 antibodies for 20 min at 4°C. After being washed with MACS buffer, the labeled cell mixture was then passed through a separation column. CD45 + cells were obtained by washing with MACS buffer on the separated column.
Real-time quantitative polymerase chain reaction
Total RNA was extracted using the easy-spinTM Total RNA Extraction Kit (iNtRON Biotechnology, Seongnam, Republic of Korea), and cDNA was synthesized from 500 ng RNA using HiSenScriptTM RH (-) RT PreMix Kits (iNtRON Biotechnology, Seongnam, Republic of Korea). Real-time quantitative polymerase chain reaction (RT-qPCR) was performed using a Rotor-Gene Q cycler (Qiagen, Hilden, Germany) and Quantinova SYBR® Green PCR Master Mix (Qiagen), each of which contained 50 ng of cDNA and 0.7 μM forward and reverse primers. RT-qPCR was run in a Rotor-Gene Q RT-qPCR machine (Qiagen) following predenaturation, denaturation, and a combined annealing/extension step. Cycling conditions were 95°C for 2 min, followed by 40 cycles of 95°C for 10 s and 60°C for 6 s. Melting curve analysis was performed from 60°C to 95°C. All experiments were performed in triplicate and used Tyrosine 3-Monooxygenase/Tryptophan 5-Monooxygenase Activation Protein Zeta (YWHAZ) as an internal control. The relative levels of gene expression were calculated by using the 2−ΔΔCT method. The primers used in the present study are listed in Table 1.
Lists of Primers Used in RT-qPCR Analysis

RT-qPCR, real-time quantitative polymerase chain reaction.
Ethics approval and animals
All animal experiments were approved by the Animal Ethics Committee of Gyeongsang National University (Permit No: GNU-210201-M0007). Female C57BL/6 mice (7–8 weeks old, 20∼22 g) were purchased from Central Lab Animal Inc. (Seoul, Republic of Korea). The mice were maintained under standard laboratory conditions (12 h light/dark cycle, 30%–40% humidity and 25 ± 2°C).
Induction of myelosuppression mouse model and D-HSCs transplantation
Mice were randomly assigned to Control, MSM, and D-HSCs transplantation (HSCT) groups (n = 5/group). Myelosuppression was induced by intraperitoneal busulfan (BU; 20 mg/kg/day) for 4 days followed by cyclophosphamide (CY; 120 mg/kg/day) for 2 days with minor modifications. 22 After 1 day from MSM induction, the mice were anesthetized with Avertin (2, 2, 2- Tribromoethanol, Sigma, Saint Louis, MO, USA) and injected with D-HSCs (1 × 106 cells/100 μL PBS) into the intra-femoral space. All mice measured their body weights every day for 20 days. Blood, spleen, thymus, and femur samples were collected at sacrifice for subsequent analyses.
Analysis of the cell population on D-HSC transplantation
At 14 days after transplantation, mice were sacrificed. Cells were obtained from spleen and thymus by mechanical dissociation. Bone marrow cells were obtained via flushing the femur. Spleen, thymus, and bone marrow cells were stained with fixable viability dye (eBioscienceTM, San Diego, CA, USA) antibody. Stained cells were analyzed using flow cytometry and raw data were analyzed using FlowJo v10 software.
Histological assessment of D-HSCs transplantation
Spleen and thymus tissues were fixed in 4% paraformaldehyde for overnight. The fixed tissue samples were embedded in paraffin and sectioned at 5 μm thickness. Then, the slides were stained with hematoxylin and eosin (H&E). The slides observed using a light microscope (Nikon Eclipse 80i) with Photo Imaging System (Canon 600 D).
Complete blood cell count analysis
Blood collection via direct cardiac puncture. The samples were transferred into anticoagulant EDTA tube for complete blood count (CBC) analysis. CBC was conducted using a hematology analyzer (ProCyte Dx Hematology Analyzer; IDEXX, Cumberland County, ME, USA).
Statistical analysis
Data were presented as mean ± standard error of the mean (SEM). The statistical differences were estimated by student’s t-test and one-way analysis of variance (ANOVA) followed by Tukey’s post-hoc in GraphPad Prism 8. The statistical difference at P < 0.05 was considered significant.
Results
Isolation and characterization of dental tissue-derived mesenchymal stem cells
DMSCs were isolated from abandoned dental tissue after wisdom teeth extraction. At passage 3, the cells showed adherent growth and spindle-like shape morphology (Fig. 1A). Flow-cytometry showed that DMSCs expressed MSC-specific markers including CD44, CD73, CD90, and CD105 but not hematopoietic markers CD34 and CD45 (Fig. 1B). DMSCs were successfully differentiated into mesenchymal lineages including osteoblasts, adipocytes and chondroblasts as confirmed by Alizarin Red S, Oil Red O, and Alcian Blue staining, respectively (Fig. 1C).
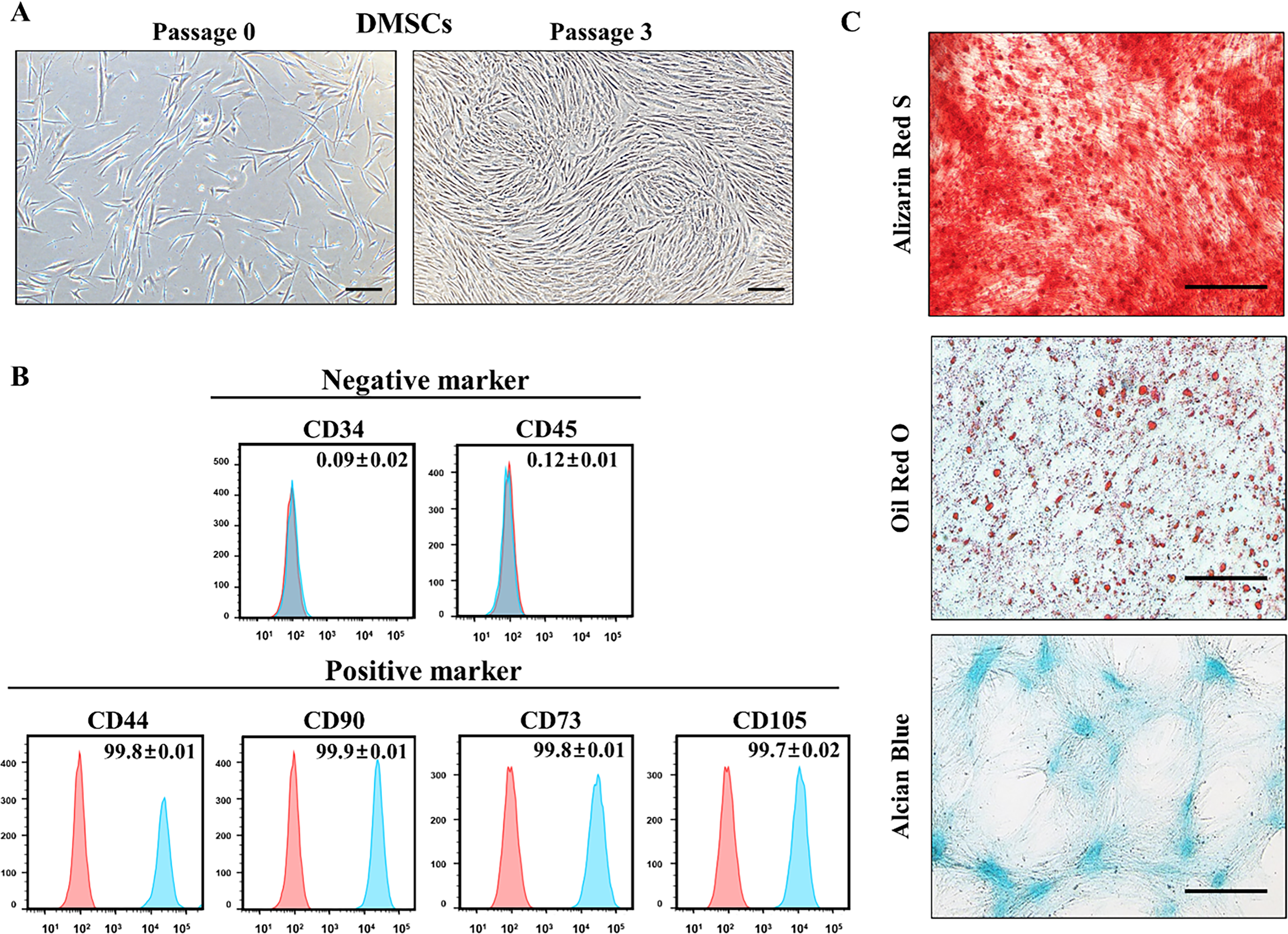
Characterization of DMSCs.
In vitro differentiation of DMSCsOCT3/4+ into HSC-like cells
In transfected cells, expression of OCT3/4 by neon transfection was significantly higher than DMSCs by expression of mRNA level (Fig. 2A). After hematopoietic induction, DMSCs OCT3/4+ exhibited morphological changes, which altered from spindle-like structure to round shape (Fig. 2B). Differentiated cells were positively expressed hematopoietic markers CD34 and CD45 (Fig. 2C). To evaluate lineage-specific differentiation, the expression of mesodermal genes such as GATA2 and hematopoietic specific genes including C/EBPα, RUNX1, and SCL mRNA level were analyzed by RT-qPCR in DMSCs OCT3/4+ and D-HSCs sorted after differentiation. Expression of GATA2, C/EBPα, RUNX1, and SCL in D-HSCs was significantly increased compared with DMSCs OCT3/4+ (Fig. 2D).
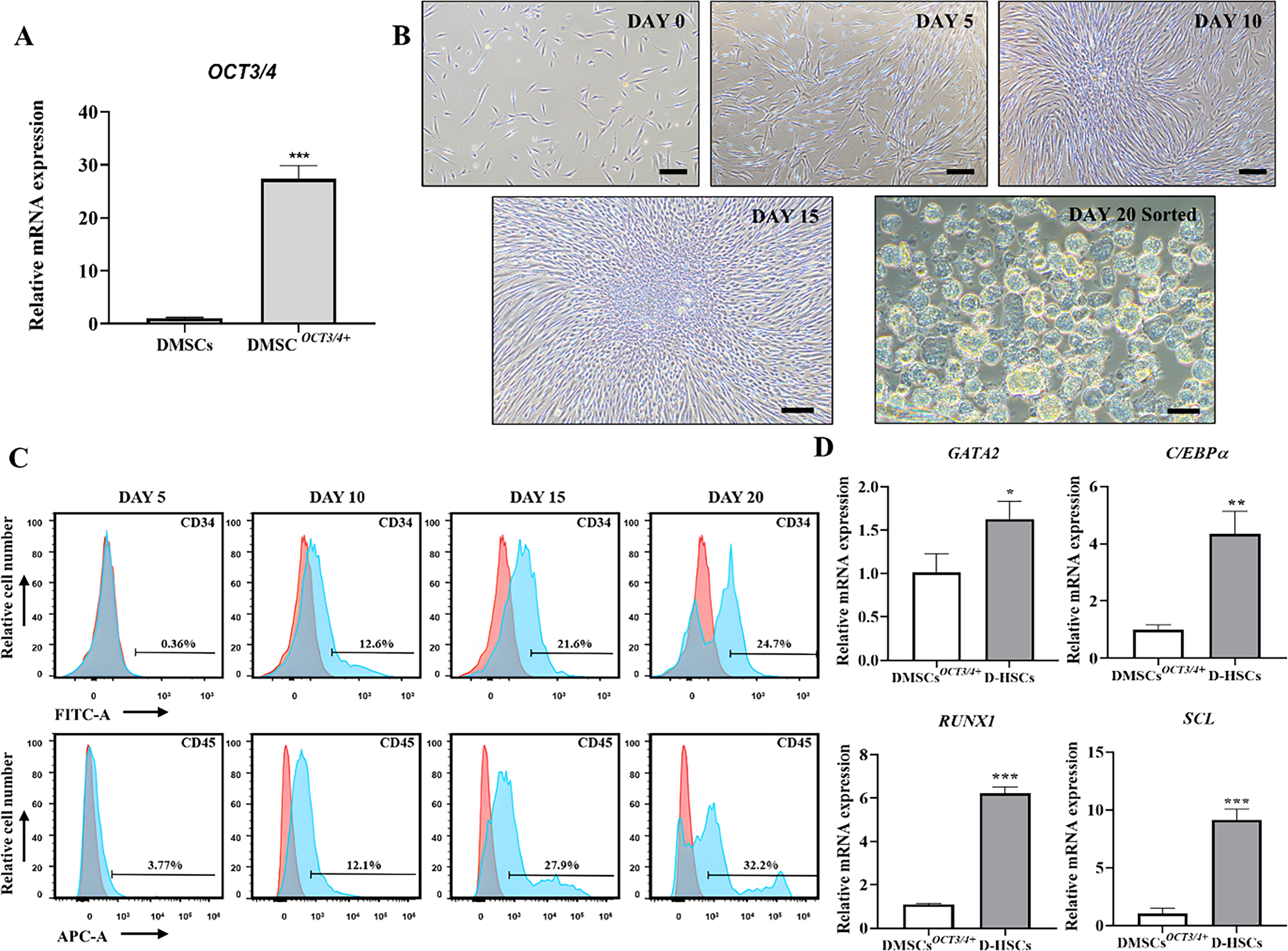
Differentiation into HSC-like cells derived from DMSCs
OCT3/4+
.
Restoration of body weight and hematopoiesis associated organ by D-HSCs transplantation
Following MSM induction, the HSCT group was performed D-HSC transplantation into the femur (Fig. 3A). Body weight in MSM group was significantly lower than Control group, which was confirmed as the MSM model was induced. After D-HSCs transplantation, compared with MSM group, the body weight in HSCT group was significantly increased (Fig. 3B). Spleen weight was significantly decreased in MSM group compared to Control group and showed significant recovery in HSCT group. The thymus weight was also significantly reduced in MSM group, while it was significantly recovered in the HSCT group. Similarly, size of spleen was significantly smaller in MSM group than in the Control group, whereas HSCT group was significantly larger than MSM group. Thymus size also significantly decreased in MSM group and showed recovery trend in the HSCT group (Fig. 3C).
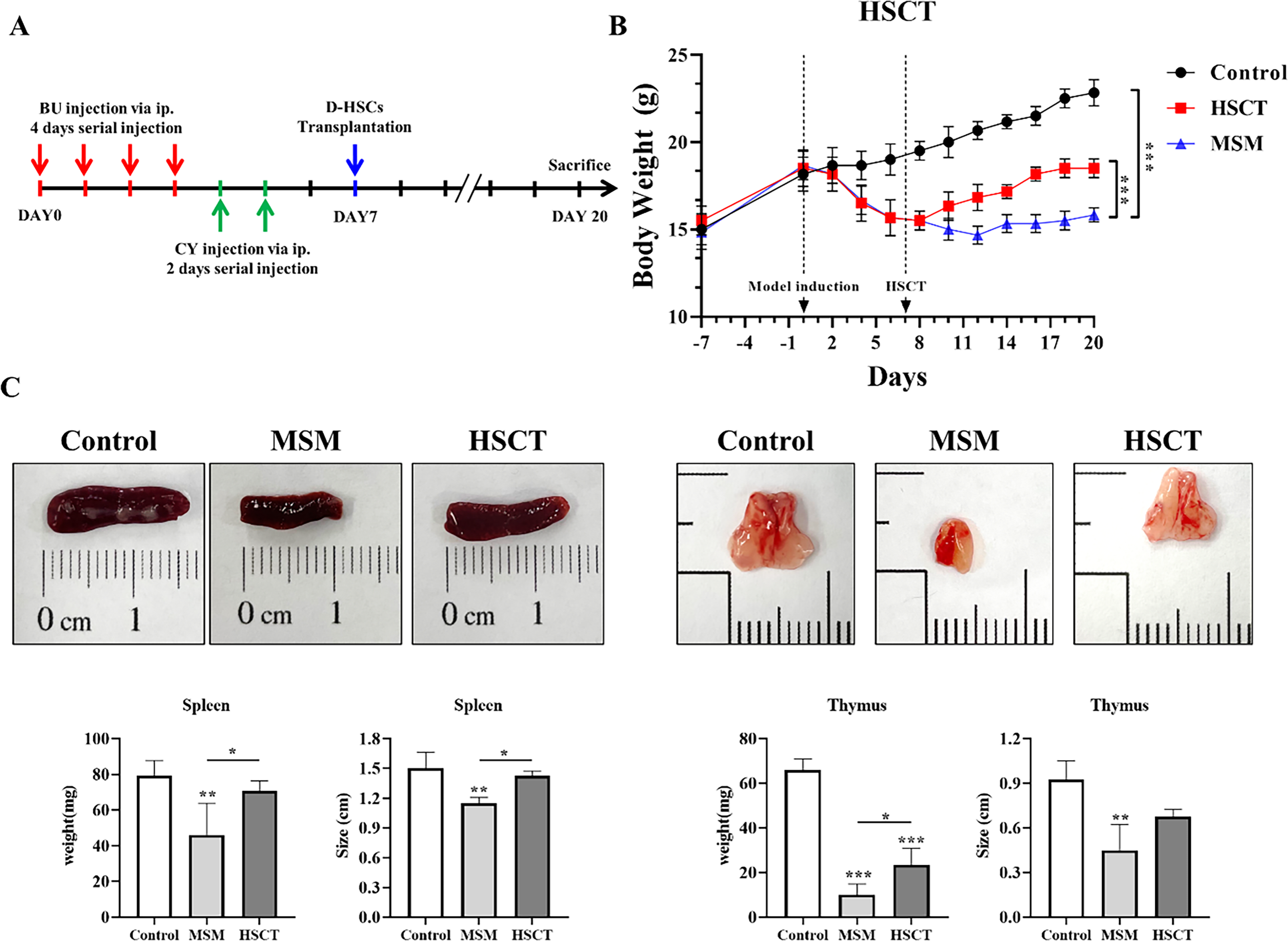
Recovery of body weight and hematopoietic tissue by D-HSC transplantation.
Live/dead cell population of MSM model and transplantation with D-HSCs
At 14 days after D-HSCs transplantation, cells were isolated from spleen, thymus, bone marrow of each experimental group. To assess cell viability, the cells were analyzed by flow-cytometry using a fixable viability (Fig. 4A). Compared with Control group, the proportion of viable cells was significantly reduced in the MSM group in spleen, thymus, and bone marrow. In contrast, the HSCT group showed significantly higher viability than the MSM group in the spleen, thymus, and bone marrow (Fig. 4B). However, cell viability in the HSCT group remained significantly lower than Control group across all tissue, indicating partial recovery.
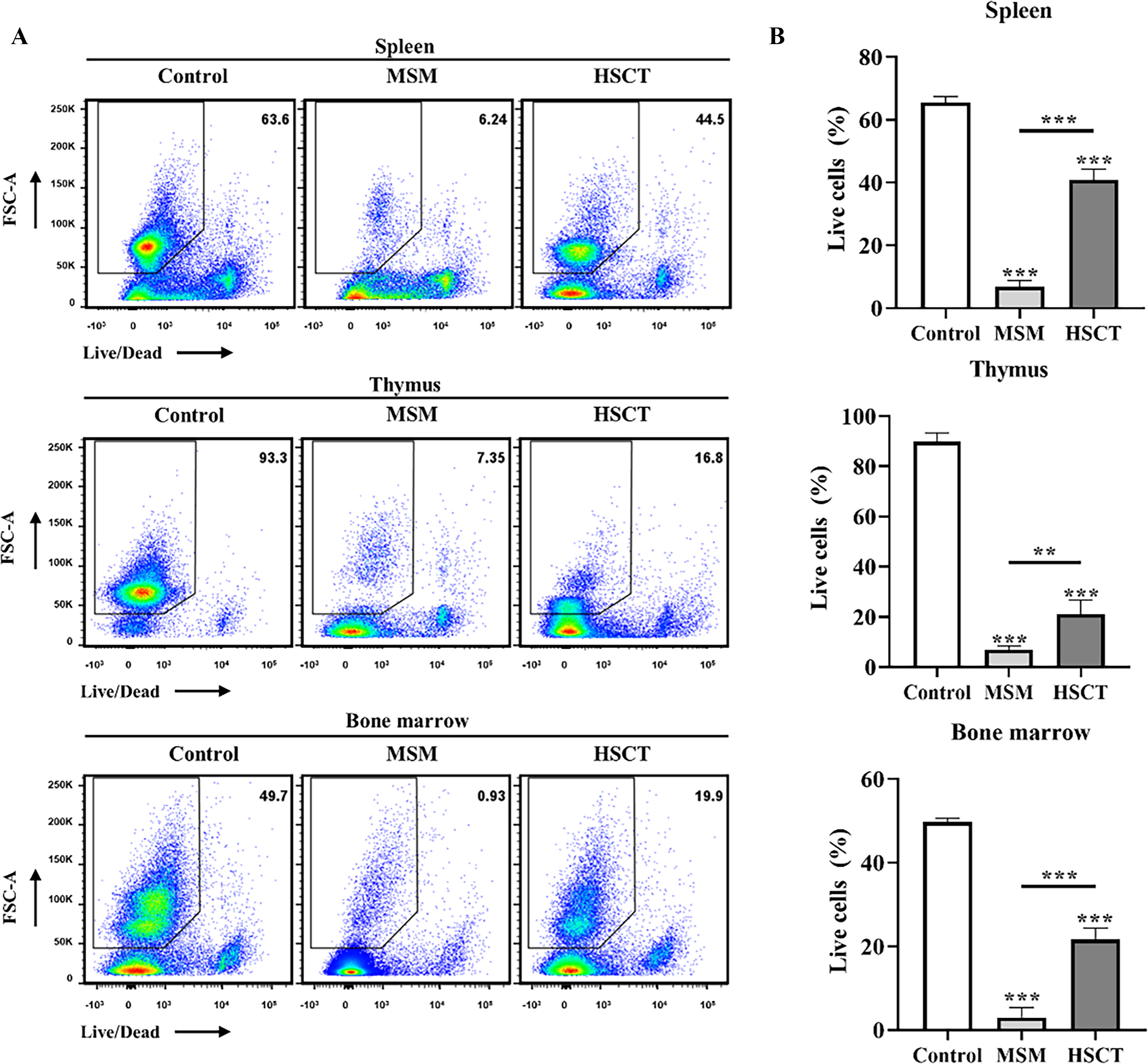
Therapeutic effect of D-HSCs on live/dead cell populations.
Histological analysis of spleen and thymus after transplantation with D-HSCs
H&E staining showed normal splenic white pulp architecture and dense lymphocyte populations in the Control group. In contrast, the MSM group showed disrupted white pulp integrity and marked lymphocyte depletion. These changes were partially restored in the HSCT group (Fig. 5A). In the thymus, the Control group showed a clear capsule and distinct cortical and medullary regions, whereas the MSM group exhibited cortical thinning, loss of corticomedullary distinction, and reduced cellularity. The HSCT group showed partial recovery of thymic architecture and lymphocyte density (Fig. 5B).

Histological analysis of the spleen and thymus after D-HSCs transplantation.
Complete blood count of MSM model and transplantation with D-HSCs
CBC analysis showed that RBC count, hematocrit, and hemoglobin were significantly reduced in the MSM group compared with the Control group. Hematocrit and hemoglobin were significantly restored in the HSCT group compared with the MSM group, whereas RBC count did not differ significantly between the MSM and HSCT groups. Reticulocyte counts were significantly higher in the HSCT group than in both the Control and MSM groups (Fig. 6A).
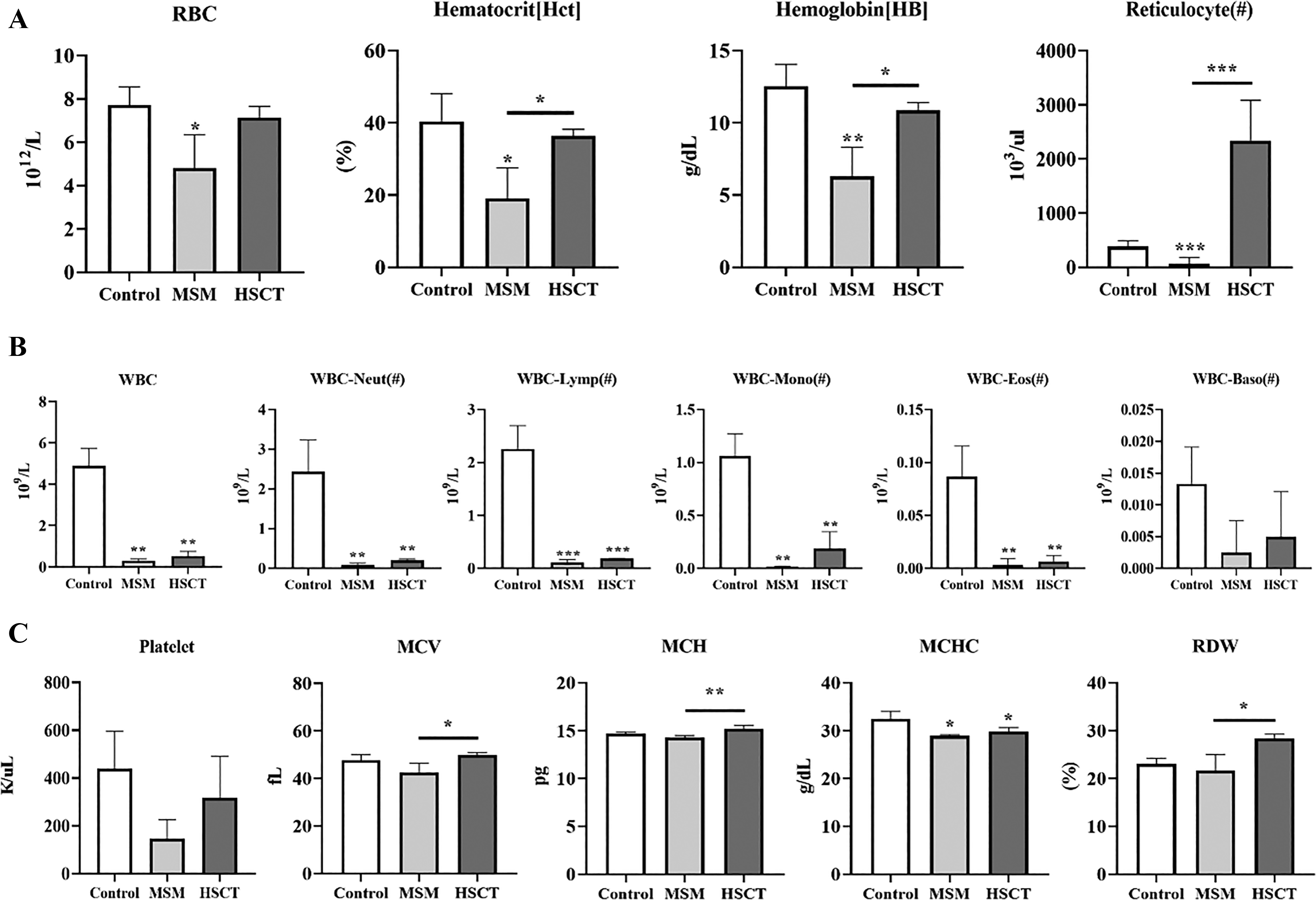
Complete blood count analysis on effect of D-HSC transplantation.
Total WBC count was significantly higher in the Control group than in both the MSM and HSCT groups. Neutrophils, lymphocytes, monocytes, eosinophils, and basophils were reduced in both the MSM and HSCT groups compared with the Control group, although the HSCT group showed a trend toward recovery across all WBC subtypes relative to the MSM group (Fig. 6B).
Platelet count was decreased in the MSM group and restored in the HSCT group. MCV and MCH were significantly increased in the HSCT group compared with the MSM group, respectively. MCHC was significantly higher in the Control group than in both the MSM and HSCT groups. RDW also significantly improved after D-HSC transplantation compared with that in the MSM group (Fig. 6C).
Discussion
HSCs sustain lifelong hematopoiesis through self-renewal and multilineage differentiation. Although HSCT is an established therapy for hematologic disorders, its broader application remains limited by immune incompatibility, graft-versus-host disease, and graft failure. 22 Therefore, alternative hematopoietic cell sources that support hematopoietic regeneration are still needed. Because of their plasticity, MSCs have been investigated as candidate cell sources for hematopoietic regeneration. OCT4-driven programs have been implicated in lineage remodeling and hematopoietic/erythroid conversion in nonhematopoietic cell.18,21 In this study, it was evaluated whether OCT3/4 overexpression in DMSCs enhances hematopoietic trans-differentiation, and whether the resulting HSC-like cells confer therapeutic effect in MSM mouse model.
Human DMSCs displayed typical MSC morphology, MSC markers expression, and mesodermal differentiation capacity, consistent with prior study. 23 Dental MSCs have also been broadly characterized as clinically accessible sources of MSCs with regenerative and immunomodulatory potential. 10 Although MSCs are classically defined by mesenchymal lineage differentiation, trans-differentiation toward non-mesodermal fates has been reported under specific microenvironmental conditions and instructive signaling.24,25 Similarly, pluripotent stem cells can be directed toward HSC-like states by hematopoietic cytokines and growth factors, supporting the concept that an epigenetically primed stem/progenitor state may improve hematopoietic induction efficiency.26,27
After OCT3/4 overexpression and cytokine induction, DMSCs acquired a rounded morphology, expressed CD34 and CD45, and upregulated C/EBPα, RUNX1, and SCL, indicating hematopoietic trans-differentiation. These findings align with reports that OCT4-related programs can contribute to lineage conversion in specific contexts, rather than acting solely as a factor for pluripotency maintenance.16–18 C/EBPα is a key regulator of myeloid commitment and hematopoietic fate decisions, while RUNX1 and SCL cooperate within transcriptional regulatory networks required for definitive hematopoietic specification and emergence.28,29 These findings suggest that OCT3/4 overexpression increases cellular permissiveness for hematopoietic gene activation, which is further reinforced by hematopoietic cytokine signaling. However, because marker expression and transcriptional changes alone do not establish bona fide long-term HSC identity, further studies should include functional assays such as long-term multilineage reconstitution and secondary transplantation.
In vivo, intrafemoral transplantation of D-HSCs into MSM mice promoted recovery of bone marrow cellularity and partial restoration of spleen and thymus size and architecture, together with improved body weight. Previous studies have demonstrated that transplanted HSCs can reconstitute all major hematopoietic lineages including lymphoid and myeloid cells and promote the recovery of immune organs and hematopoietic cell populations following myelosuppressive injury.30,31 These results suggest not only enhanced cell survival but also functional restoration of hematopoietic activity. In addition, histological analysis revealed restoration of the structural integrity of the spleen and thymus following transplantation. Consistent with previous studies, restoration of the splenic structure and recovery of the thymus suggest that hematopoietic reconstitution and cytokine-mediated pathways contribute to immune organ regeneration following injury.30,31 Nevertheless, additional immune-functional analyses will be required to confirm restoration of immune competence.
Peripheral blood findings also indicated hematopoietic recovery after transplantation. In MSM mice, RBC and reticulocyte reductions were consistent with impaired erythropoiesis, while transplantation was associated with partial normalization of these parameters. Leukocyte subsets also showed a trend toward recovery after transplantation, although some changes did not reach statistical significance within the evaluated timeframe. Because hematopoietic reconstitution after transplantation follows defined kinetics and may require weeks to stabilize across lineages, modest early recovery can be biologically plausible and time-dependent.32–34 Platelet counts and erythrocyte indices, including MCV, MCH, MCHC, and RDW, also showed partial improvement. Recovery of these indices has been associated with red cell regeneration and erythropoietic activity. 35 Together, these results suggest that D-HSC transplantation contributes to recovery of both marrow and peripheral hematologic parameters after myelosuppressive injury.
In conclusion, OCT3/4-overexpressing DMSCs could be directed toward an HSC-like state, and the resulting D-HSCs were associated with improved hematopoietic and immune-related endpoints in a myelosuppressed mouse model. This strategy may provide a practical approach for generating hematopoietic progenitor-like cells without full pluripotent reprogramming. However, long-term stability, multilineage reconstitution capacity, genomic safety, and the mechanisms linking OCT3/4 priming to hematopoietic fate acquisition require further investigation.
Data Availability Statement
All primary data are included in the article Figures. All original data are available upon reasonable request from the corresponding author.
Footnotes
Author Disclosure Statement
No potential conflict of interest was reported by the author(s).
Funding Information
This study was supported by a grant from the National Research Foundation (NRF) and was funded by the Government of the Republic of Korea (grant no.