
Abstract
Identification of natural compounds that delay aging and prevent age-related neurodegeneration is a key goal in gerontology. Fucoxanthin, a marine-derived xanthophyll, exhibits potent antioxidant properties, yet its effects on organismal aging and specific molecular mechanisms remain underexplored. Here, we investigated the pro-longevity and neuroprotective effects of fucoxanthin using Caenorhabditis elegans. Fucoxanthin supplementation significantly extended the mean lifespan of wild-type nematodes by 12.1% and improved health span, as evidenced by delayed age-related motility decline and enhanced resistance to oxidative stress. Notably, this lifespan extension occurred without compromising reproductive fitness. Genetic analysis revealed that the beneficial effects of fucoxanthin require the FOXO transcription factor DAF-16 and the autophagy-essential gene bec-1. Furthermore, fucoxanthin treatment increased autophagic flux and upregulated the expression of SKN-1/Nrf2-dependent detoxification genes, hsp-16.2 and gst-4. In nematode models of Alzheimer’s and Parkinson’s disease, fucoxanthin significantly ameliorated Aβ-induced paralysis and protected against dopaminergic neurodegeneration and α-synuclein accumulation in a DAF-16-dependent manner. Collectively, our findings demonstrate that fucoxanthin acts as a multitarget geroprotector that promotes healthy aging through the coordinated activation of DAF-16 and autophagy, suggesting its potential as a therapeutic intervention for age-related decline.
Introduction
Aging is a universal and inevitable biological process characterized by a progressive decline in physiological integrity, leading to impaired function and increased vulnerability to death. Understanding the molecular mechanisms underlying aging is a major focus of contemporary biomedical research. However, the precise molecular mechanisms of aging remain underexplored. The free radical theory of aging underscores reactive oxygen species (ROS)-mediated macromolecular damage as a pivotal driver of aging phenotypes. 1 ROS are produced as by-products of metabolic processes, mainly the mitochondrial electron transport chain reactions, and heighten vulnerability to age-related diseases such as Alzheimer’s disease (AD) and Parkinson’s disease (PD). 2 To defend against harmful ROS-induced damage, cells have evolved antioxidant defense mechanisms. Enzymatic defense mechanisms employ antioxidant enzymes, such as superoxide dismutase, catalase (CAT), and glutathione peroxidase (GPx). However, previous studies involving genetic manipulations of antioxidant enzymes in various model organisms suggest that antioxidant enzymes are not universal, primary causal determinants of aging. 3
Nonenzymatic defense is mediated by antioxidant molecules, such as vitamin E, vitamin C, and glutathione. The identification of natural antioxidants delaying aging and reducing the development of age-related diseases is one of the most important research in gerontology. Resveratrol, a plant-derived polyphenol rich in grapes and berries, increased the lifespan of experimental organisms, including yeast, Caenorhabditis elegans, and Drosophila melanogaster.4–6 SIR-2.1 was activated by resveratrol, and the lifespan-extending effect of resveratrol disappeared in a sir-2.1 null mutant, suggesting that SIR-2.1 mediated the increased lifespan induced by resveratrol. 7 Resveratrol also significantly augmented telomerase activity in vitro. 8 Curcumin is a major polyphenol found in Curcuma longa and is known to have strong antioxidant and anti-inflammatory activities. Supplementation with curcumin increased the lifespan of C. elegans and D. melanogaster. 9 It increased resistance to oxidative and heat stress and decreased cellular oxidative damage. 9 Spermidine, a natural polyamine found in plant- and animal-originated foods, has antitumor, antiobesity, and cardiovascular protective activities. 10 The longevity phenotype was observed in a model organism with spermidine treatment, which was mediated by activated autophagy. 11 Dietary supplementation with the plant flavonoids, fisetin and butein, increased lifespan and reduced the risk of age-related diseases, including AD and PD.12,13 Xanthoxyline, a plant-derived phenolic phytochemical, induced longevity and prevented neuronal damage. 14
Fucoxanthin is a marine-derived xanthophyll carotenoid abundant in edible brown seaweeds such as Undaria pinnatifida and Sargassum spp. It exhibits a broad spectrum of bioactivities, including antioxidant, anti-inflammatory, anti-obesity, and neuroprotective effects. 15 The antioxidant activity of fucoxanthin is thought to be derived from its chemical structure, including an allenic bond, an epoxide group, and a hydroxyl group. 15 Fucoxanthin activated Nrf2, a transcription factor regulating oxidative stress response, and increased the activities of antioxidant enzymes, such as CAT and GPx. 16 Accumulation of amyloid beta (Aβ), one of the pathological hallmarks of AD, was reduced by fucoxanthin through hydrophobic interactions with Aβ. 17 Supplementation with fucoxanthin decreased cognitive impairment observed in scopolamine-induced mice via inhibition of acetylcholinesterase. 18 Molecular docking and an in silico study revealed that fucoxanthin is a potential dopamine agonist binding to the active site of the dopamine receptor. 19 The levels of α-synuclein, a molecular marker of PD, and inflammatory factors, one of risk factors of PD, were reduced by fucoxanthin. 20 A previous study investigating the effects of two carotenoids, β-carotene and fucoxanthin, on lifespan in D. melanogaster and C. elegans revealed that β-carotene increased lifespan only in D. melanogaster, while fucoxanthin extended the lifespan of both experimental animals. 21 Transcriptional profiling using RNA sequencing suggested that the antiaging effect of fucoxanthin was mediated by mTOR signaling, autophagy, apoptosis, immune response, and DNA damage response pathways in D. melanogaster. 22
In this study, we sought to investigate the effects of fucoxanthin on stress response, aging, and age-related neurodegenerative diseases in C. elegans. Among the diverse animal models available, the nematode C. elegans has emerged as a powerful system for investigating the genetics and molecular biology of aging due to its short lifespan, defined cell lineage, and high genetic tractability. By employing both physiological assays and fluorescence-based reporters, we tested the hypothesis that fucoxanthin confers pro-longevity and neuroprotective effects, potentially via the activation of autophagy and DAF-16, a FOXO transcription factor. By bridging phenotypic observations with mechanistic insights, this study clarifies how fucoxanthin affects conserved longevity pathways and mitigates neurodegenerative phenotypes.
Materials and Methods
Worm strains and culture conditions
All C. elegans strains were obtained from the Caenorhabditis Genetics Center (Minneapolis, MN). The N2 strain was used as the wild-type control. Long-lived mutants age-1 (hx546) and eat-2 (ad465) were used to investigate mechanisms underlying lifespan extension. Autophagy activation was assessed using BC12921 (sIs10729 [rCes T12G3.1::GFP, pCeh361]). The strains CL4176 (dvIs27 [myo-3/Aβ1-42/let UTR, rol-6]) and CL2331 (dvIs37 [myo-3p::GFP::Aβ3-42 + rol-6]) express amyloid-β (Aβ) in muscle tissue. BZ555 (egIs1 [dat-1p::GFP]) expresses GFP in dopaminergic neurons, while NL5901 (pkIs2386 [unc-54p::α-synuclein::YFP + unc-119(+)]) expresses fluorescent α-synuclein in muscle. Worms were maintained on nematode growth medium (NGM) plates (25 mM NaCl, 2.5 mg/mL peptone, 50 mM KPO4 buffer, 5 µg/mL cholesterol, 1 mM CaCl2, 1 mM MgSO4, 1.7% agar) at 20°C. Escherichia coli OP50 was used as the food source. Fucoxanthin (Sigma-Aldrich, St. Louiis, MO, USA, Cat. No. F6932, purity ≥95% by HPLC) was dissolved in 99.9% ethanol to prepare a 10 mM stock solution stored at −20°C protected from light and diluted in NGM to the indicated final concentrations. The final ethanol concentration was kept constant at 0.1% (v/v) on all plates, including the untreated (vehicle) control, to exclude solvent-related artifacts.
In vitro radical-scavenging activity
ABTS [2,2’-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid)] solution was made by mixing equal volumes of 7.4 mM ABTS and 2.45 mM potassium persulfate and incubating the mixture at 20°C for 24 hours in the dark. Then, ABTS solution was diluted with 10% ethanol to an absorbance of 0.7 ± 0.03 at 734 nm. Fucoxanthin (20 μL) was mixed with ABTS solution (180 μL) in a 96-well plate. After incubation for 5 minutes, absorbance was measured at 734 nm. Ascorbic acid (100 µM) was used as a positive control.
Survival after environmental stresses
Three-day-old worms were randomly selected and exposed to various concentrations of fucoxanthin for 24 hours (n = 30). Subsequently, worms were transferred to individual wells of a 96-well plate containing 2 mM H2O2 in cholesterol-free S-basal medium (5.85 g NaCl, 1 g K2HPO4, 6 g KH2PO4 per liter of distilled water). After 8 hours of incubation under oxidative stress, worm survival was recorded. For heat and ultraviolet (UV) stress, synchronized 3-day-old worms (n = 60) were subjected to 8 hours of 35°C heat shock or 1 minute of UV exposure (20 J/cm2/min), respectively. Survival was monitored daily under a microscope until all worms had died.
Lifespan assay
At 3 days posthatching, 60 worms were transferred to NGM plates containing 5-fluoro-2’-deoxyuridine (FUdR) (12.5 mg/L) to prevent progeny production. Live and dead worms were counted daily, and worms lost, damaged, or internally hatched were excluded.
Fertility assay
Twelve adult worms were randomly selected at 48 hours posthatching. Each was placed on a separate NGM plate and allowed to lay eggs at 20°C. After 24 hours, the adults were transferred to new plates. Eggs were incubated at 20°C for 48 hours, and the number of progeny was counted. This procedure was repeated daily throughout the reproductive period.
Motility assay
Locomotor activity was evaluated at 5, 10, 15, 20, and 25 days posthatching (n = 100). Activity levels were categorized as phase 1: spontaneous movement, phase 2: movement only upon mechanical stimulation, and phase 3: restricted head movement upon stimulation. Additionally, the number of body bends per minute in the M9 buffer was recorded for 15 worms.
RNAi
RNAi clones targeting daf-16, skn-1, and bec-1 were obtained from the Ahringer RNAi library. 23 The empty L4440 vector served as the negative control (EV). HT115(DE3) bacteria carrying each construct were grown overnight in LB with 50 µg/mL ampicillin, seeded onto NGM plates supplemented with 1 mM IPTG, and induced at room temperature for 24 hours prior to transferring worms.
Autophagic activity
Synchronized 3-day-old BC12921 worms (n = 25) expressing fluorescent sequestosome-1 (SQST-1) were treated with fucoxanthin at 20°C for 48 hours. Worms were immobilized on 2% agarose pads with 1 M sodium azide and observed under a fluorescence microscope. Images were analyzed quantitatively using ImageJ software.
Quantitative real-time PCR
Total RNAs were extracted from approximately 300 9-day-old worms using a Trizol reagent (Thermo Fisher Scientific, Waltham, MA, USA) and reverse-transcribed to complementary DNA. Quantitative real-time PCR was performed using a ReverTra Ace qPCR RT Master Mix (TOYOBO, Osaka, Japan), 2× SyGreen Mix Hi-ROX (qPCRBIO, London, UK), and a StepOne Plus Real-Time PCR System (Applied Biosystems, Waltham, MA, USA). Primer pairs for each gene are shown in Supplementary Table S1. The ama-1 gene was used for normalization and the relative expression of each gene was calculated using the 2–ΔΔCt method.
Genetic model of AD
Eggs from CL4176 worms were incubated at 15°C for 24 hours. Sixty worms were then transferred to 25°C for 24 hours to induce Aβ expression. Paralysis was assessed at specific time intervals until all worms were paralyzed. CL2331 worms were grown at 15°C to reach the late L4 stage. Then, worms were incubated in a 25°C incubator for 48 hours. Worms were immobilized on 2% agarose pads with 1 M sodium azide and observed under a fluorescence microscope. Images were analyzed quantitatively using ImageJ software (Washington, D.C., USA).
Genetic model of PD
BZ555 worms were exposed to 50 mM 6-hydroxydopamine (6-OHDA) with 10 mM ascorbic acid at 20°C for 1 hour, with gentle mixing every 10 minutes. After washing with M9 buffer, worms were transferred to NGM plates containing OP50 and 12.5 mg/L FUdR and incubated at 20°C for 72 hours. Degeneration of dopaminergic neurons was visualized using fluorescence microscopy (470/22 nm excitation, 525/50 nm emission). For NL5901 worms, 3-day-old adults were bleached, and their eggs were cultured in M9 buffer at 20°C for 24 hours. Hatched larvae were then transferred to NGM and incubated for 5 days at 20°C. α-Synuclein fluorescence was measured (500/24 nm excitation, 542/27 nm emission), and intensity in the head region was quantified using ImageJ.
Fluorescence imaging and ImageJ quantification
All fluorescence images within a given experiment were acquired with identical microscope settings (fixed exposure time, gain, and lamp intensity). Raw 16-bit images were imported into ImageJ (NIH, v1.54). Background subtraction was performed with the rolling-ball algorithm (radius = 50 pixels). Regions of interest (ROIs) were drawn manually around the head region (for Aβ and α-synuclein) or the pharyngeal–anterior intestinal region (for SQST-1::GFP) or around individual dopaminergic neurons, deliberately excluding the intestinal lumen to avoid lipofuscin autofluorescence. Mean gray values from ≥20 worms per condition were averaged and normalized to the untreated control set to 100%.
Statistical analysis
ABTS radical-scavenging data were analyzed by one-way analysis of variance followed by Tukey’s post hoc test (n = 3 independent experiments, each in triplicate). Survival data (lifespan, oxidative stress, paralysis) were analyzed by the log-rank test. 24 Quantitative PCR, fluorescence intensity, motility, and progeny data were compared using Student’s two-tailed t-test. A value of p <0.05 was considered statistically significant.
Results
Fucoxanthin reduced free radicals and enhanced resistance to oxidative stress
The radical-scavenging activity of fucoxanthin was evaluated using the ABTS assay. Fucoxanthin demonstrated potent radical-scavenging activity (Fig. 1A). At 100 μM, fucoxanthin significantly reduced free radical levels (41.4 ± 2.75%, p < 0.001), compared to the untreated control (100.8 ± 0.59%, mean ± standard error), and showed greater efficacy than the same concentration of ascorbic acid, a widely used antioxidant (72.7 ± 0.60%, p < 0.001). Lower concentrations of fucoxanthin (10 μM) also significantly reduced free radicals to 83.0 ± 0.71 (p = 0.003), while 1 μM of fucoxanthin failed to show a significant effect (95.3 ± 0.42, p = 0.175). In addition, fucoxanthin supplementation significantly enhanced resistance to oxidative stress (Fig. 1B). Survival after oxidative stress increased from 33.3 ± 3.33% in the untreated control to 55.6 ± 1.11% in worms pretreated with 10 μM fucoxanthin (p = 0.003). However, fucoxanthin had no effect on resistance to other environmental stressors, such as heat shock and UV irradiation (Fig. 1C, D). This stressor-selective profile, efficacy against H2O2 but not heat or UV, suggests that fucoxanthin acts predominantly through redox-buffering rather than via general proteotoxic or DNA-damage stress–response programs.
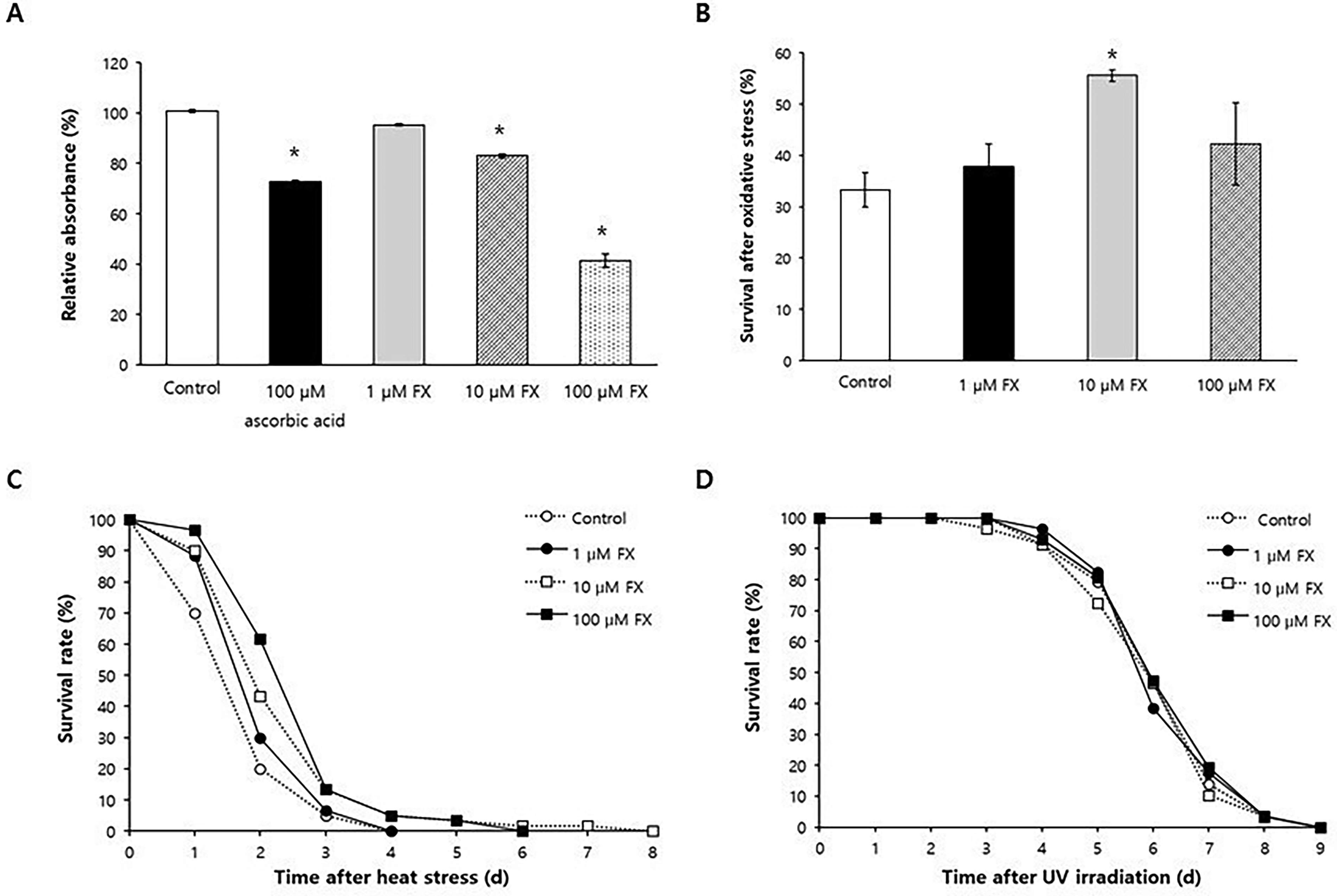
Effect of fucoxanthin on response to environmental stressors.
Fucoxanthin extended lifespan without reducing fertility
Based on the increased survival under oxidative stress, we examined the antiaging effect of fucoxanthin at 10 μM. Lifespan was significantly extended in fucoxanthin-treated worms, with a 12.1% increase in mean lifespan (23.5 days for the untreated control vs. 26.4 days for the fucoxanthin-treated group, p = 0.006) (Fig. 2A). Maximum lifespan also increased slightly (34 vs. 36 days, respectively). These findings were consistently observed with independent experiments (Supplementary Table S2). To assess potential trade-offs, we measured fertility, which is often reduced in lifespan-extending interventions.12,13,25 The average number of progeny per worm was not significantly different between the untreated control and the fucoxanthin-treated group (223.6 for the untreated control vs. 238.0 for the fucoxanthin-treated group, p = 0.291), and the daily progeny distribution during the reproductive period remained unaffected by fucoxanthin (Fig. 2B).
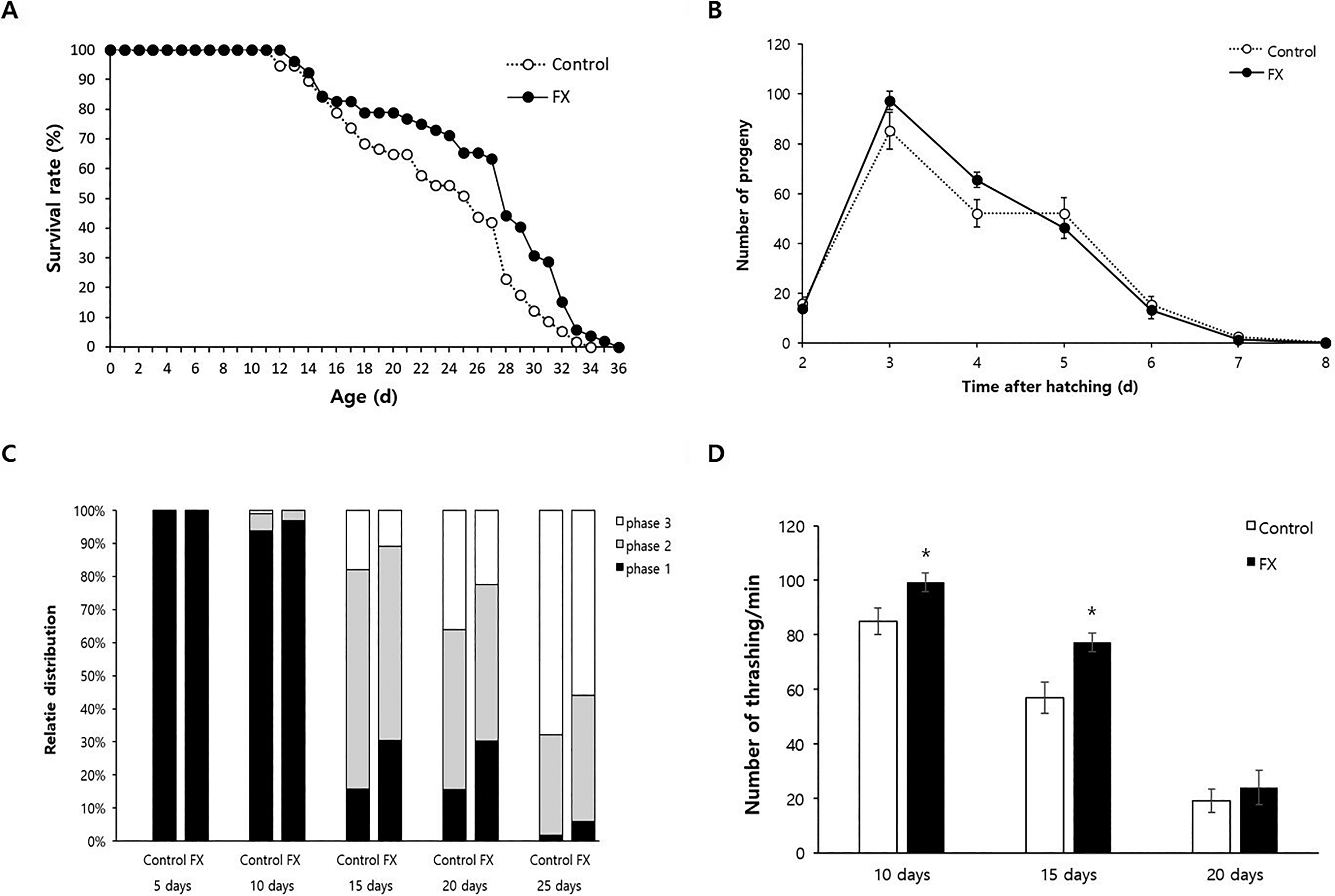
Effect of fucoxanthin on lifespan, fertility, and motility.
Age-related decline in motility was delayed by fucoxanthin
To assess the effect of fucoxanthin on age-related motor decline, we monitored the motility of individual worms over time. As expected, worm motility declined with age, with a decrease in spontaneous movement (phase 1) and an increase in head-only movement (phase 3) (Fig. 2C). Fucoxanthin significantly mitigated this decline. For 15-day-old worms, the proportion of phase 1 animals increased from 15.7% in the untreated control to 30.4% in fucoxanthin-treated worms, while phase 3 animals decreased from 18.0% to 10.9%. Similar improvements were observed for 20-day-old worms (Fig. 2C). Quantitative assessment using thrashing assays further confirmed this effect: in 10-day-old worms, thrashing rates were significantly higher in fucoxanthin-treated animals (99.3 vs. 84.9, p = 0.020), as well as in 15-day-old worms (77.2 vs. 58.9, p = 0.014) (Fig. 2D).
Lifespan extension by fucoxanthin required daf-16 and bec-1
To elucidate the mechanisms of fucoxanthin-induced longevity, we examined its effects in long-lived mutants. In the age-1 mutant, which has reduced insulin/IGF-1-like signaling, fucoxanthin did not further extend lifespan (p = 0.337) (Fig. 3A). Similarly, no additional effect was observed in the eat-2 mutant, a genetic model of dietary restriction (DR) (p = 0.225). The results of replicative experiments were shown in Supplementary Material (Supplementary Table S3). These findings suggest that fucoxanthin’s lifespan-extending mechanism overlaps with pathways involved in insulin/IGF-1-like signaling and DR. We further tested whether key regulators in these pathways were required for the fucoxanthin-induced longevity. RNAi-mediated knockdown of daf-16, a FOXO transcription factor in insulin/IGF-1-like signaling, completely abolished the lifespan-extending effect of fucoxanthin: mean lifespans were 14.4 days without fucoxanthin and 14.1 days with fucoxanthin (p = 0.622) (Fig. 3B). Likewise, inhibition of bec-1, a key autophagy gene, also eliminated the longevity phenotype conferred by fucoxanthin. Mean lifespan of the untreated worms (19.3 days) was not significantly different from that of fucoxanthin-treated worms (19.1 days, p = 0.979). Results of repeated experiments were shown in Supplementary Table S4.
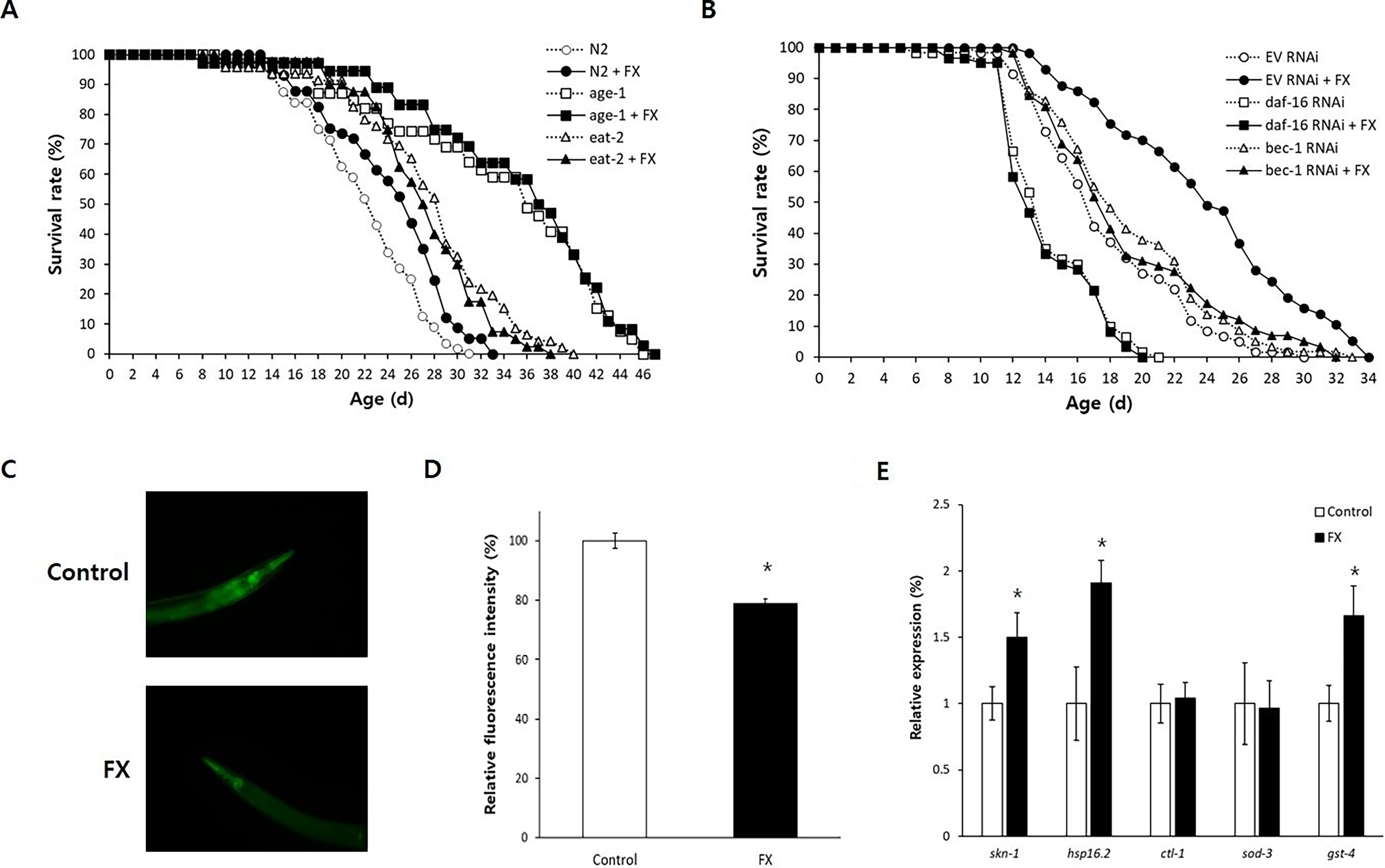
Genetic analysis of fucoxanthin-induced longevity.
Fucoxanthin activated autophagy and SKN-1-dependent gene expression
Given the requirement of bec-1 for fucoxanthin-induced lifespan extension, we investigated whether fucoxanthin enhances autophagic activity. In vivo autophagy was assessed via levels of sequestosome-1 (SQST-1), a substrate of autophagy. Relative protein levels of SQST-1 were significantly decreased in worms treated with fucoxanthin (79.0 ± 1.62) compared to the untreated control (100.0 ± 2.55, p < 0.001), indicating enhanced autophagy in fucoxanthin-treated worms (Fig. 3C, D). Previous studies suggest that the antiaging effect of fucoxanthin is mediated by Nrf2, a stress-responsive transcription factor, in mice. 26 We analyzed the expression of skn-1, a worm homolog of Nrf2, and its downstream targets. The expression of skn-1 was significantly induced by fucoxanthin treatment (1.5-fold, p = 0.049). Among downstream targets of SKN-1, hsp-16.2 and gst-4 were 1.9- and 1.7-fold upregulated by supplementation with fucoxanthin, respectively (p < 0.05). However, the expression of ctl-1 and sod-3 was not affected by fucoxanthin (Fig. 3E).
Aβ-induced toxicity was reduced by fucoxanthin
To examine the effects of fucoxanthin on age-related diseases, we utilized a C. elegans model expressing human Aβ in muscle. Fucoxanthin significantly delayed Aβ-induced paralysis (p < 0.001) (Fig. 4A). This protective effect of fucoxanthin on Aβ-induced paralysis completely disappeared when daf-16 or skn-1 was knocked down. In control worms, the time to 50% paralysis increased from 7.6 to 8.4 hours with fucoxanthin (p = 0.006), but this effect was lost under daf-16: the times to 50% paralysis were 7.4 and 7.3 hours in the untreated control and fucoxanthin-treated group, respectively (p = 0.655) (Fig. 4C). The time to 50% paralysis was not affected by fucoxanthin when the expression of skn-1 was inhibited (8.1 vs. 7.7 hours, p = 0.267). Furthermore, fucoxanthin reduced Aβ accumulation as shown in Figure 4C. The relative fluorescence intensity associated with Aβ accumulation was decreased to 61.8 ± 4.48% with fucoxanthin treatment, compared to the untreated control (100.0 ± 9.90%, p = 0.004) (Fig. 4D). Similar results were obtained with independent replicate experiments (Supplementary Table S5).
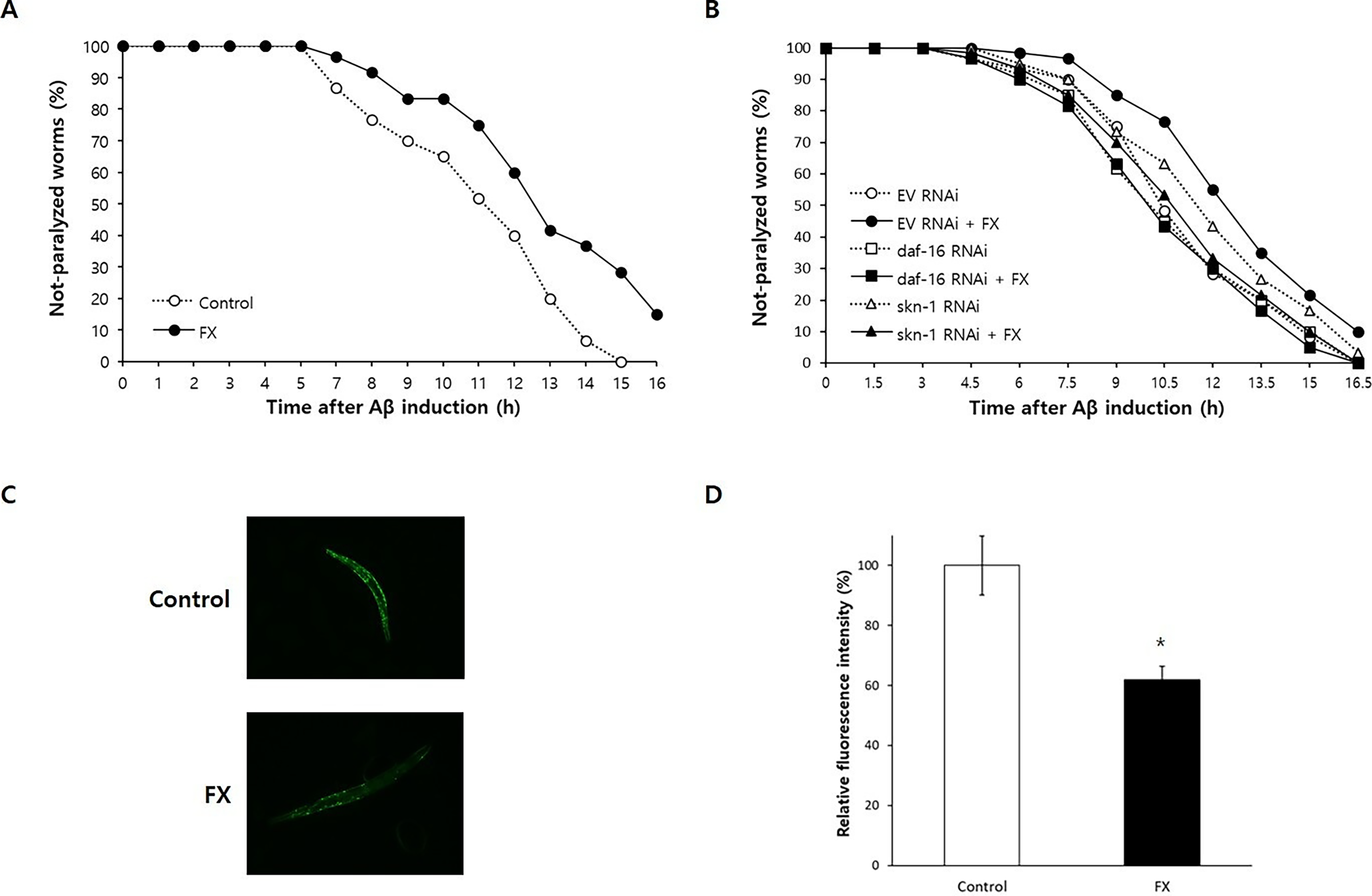
Positive effects of fucoxanthin on the risk of AD.
Fucoxanthin prevented dopaminergic neurodegeneration
Using PD models, we assessed the effect of fucoxanthin on dopaminergic neuron integrity. As shown in Figure 5A, 6-OHDA specifically degenerated dopaminergic neurons, and those neuronal damages were reversed by supplementation with fucoxanthin. 6-OHDA reduced neuronal fluorescence to 58.0 ± 4.02%, compared to the untreated control (100.0 ± 4.53%, p < 0.001). However, simultaneous treatment with fucoxanthin restored it to 100.5 ± 8.70%, p < 0.001 compared with worms treated with 6-OHDA alone (Fig. 5B). This neuroprotective effect was abolished by daf-16 RNAi, where neuronal fluorescence remained low despite fucoxanthin treatment (Fig. 5A). Decreased fluorescence intensity by 6-OHDA (62.4 ± 6.99%, p < 0.001 compared with the untreated control, 100.0 ± 6.12%) was not restored by supplementation with fucoxanthin (59.2 ± 8.01%, p = 0.760 compared with worms treated with 6-OHDA only) when the expression of daf-16 was blocked (Fig. 5B). Additionally, fucoxanthin reduced α-synuclein accumulation, a pathological hallmark of PD (Fig. 5C). Compared to the untreated control (100 ± 10.65%), worms treated with fucoxanthin showed a decreased fluorescence intensity (70.3 ± 4.56, p = 0.015) (Fig. 5D).
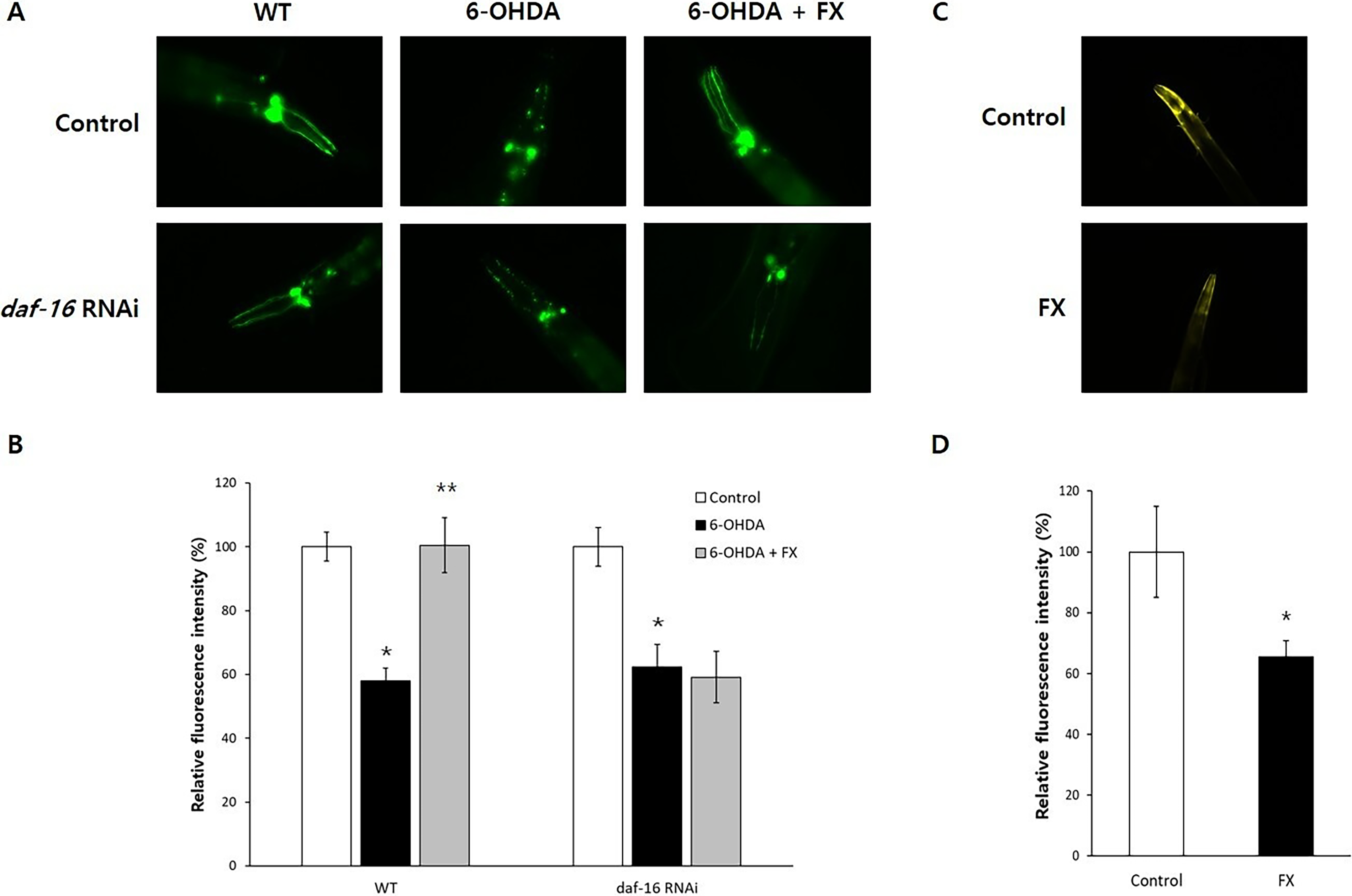
Effect of fucoxanthin on neuronal damage in PD models.
Discussion
Fucoxanthin substantially reduced free radical levels and improved resistance to oxidative stress, while leaving heat- and UV-stress survival unchanged, indicating selectivity for redox rather than general proteotoxic challenge. The absence of protection against heat shock and UV irradiation is mechanistically informative. Carotenoids, including fucoxanthin, intercept singlet oxygen and peroxyl radicals through their conjugated polyene backbone but do not directly stabilize misfolded proteins or repair DNA lesions. 27 Accordingly, fucoxanthin does not appear to engage the cytosolic proteostasis program or the DNA-damage response. This stressor-selective profile distinguishes fucoxanthin from broad-spectrum geroprotectors such as rapamycin and supports a model in which redox buffering is the proximal trigger of the downstream DAF-16/SKN-1/autophagy axis. Building on this redox-selective action, fucoxanthin extended lifespan and preserved motility into late adulthood without compromising fertility, a profile that decouples somatic maintenance from the reproductive cost predicted by the disposable-soma theory and that mirrors recent observations for phlorizin and xanthoxyline.14,28
Elucidating the molecular architecture of fucoxanthin-induced longevity revealed an overlapping effect of fucoxanthin with reduced insulin/IGF-1-like signaling and DR, and a strict dependence on conserved signaling nodes. The pro-longevity effect of fucoxanthin is strictly dependent on the FOXO transcription factor DAF-16 and the autophagy-essential gene bec-1. Coupled with the direct observation of enhanced autophagic flux, our data support the “convergent mechanism” hypothesis proposed in landmark studies. Previous studies established autophagy as an essential, common downstream effector required for diverse longevity regimens, including DR and insulin/IGF-1 signaling inhibition. 29 Recent studies revealed that lifespan extension with antioxidants, including fisetin, butein, and xanthoxyline, was mediated by activated autophagy.12–14 Fucoxanthin appears to tap into this universal pathway, promoting the recycling of damaged cellular components to maintain homeostasis. Concomitantly, fucoxanthin upregulated the expression of the Nrf2 homolog skn-1 and its downstream phase II detoxification genes, hsp-16.2 and gst-4. Activation of Nrf2 signaling was also observed in suppression of osteoclastogenesis and Aβ-induced oxidative damage by fucoxanthin.26,30 The requirement of both daf-16 and bec-1 suggests that fucoxanthin-mediated longevity is not merely a result of direct radical scavenging but rather a coordinated transcriptional program that reinforces cellular quality control through the recycling of damaged macromolecules.
The lack of additive lifespan extension by fucoxanthin in the eat-2 (ad465) genetic DR model, combined with the strict requirement for bec-1, is consistent with fucoxanthin acting as a partial DR mimetic. DR-induced longevity in C. elegans is autophagy-dependent and is mediated in part by the PHA-4/FOXA and SKN-1 transcription factors.31–33 Our observations of skn-1 induction, increased autophagic flux, and bec-1-dependence collectively position fucoxanthin within this DR axis. Importantly, unlike DR, fucoxanthin did not reduce progeny number, indicating that it preferentially activates the somatic-maintenance arm of DR signaling without reproductive trade-offs.
Beyond somatic aging, fucoxanthin exhibited exceptional neuroprotective properties in models of protein conformational disorders. In an AD model, fucoxanthin significantly delayed the onset of paralysis and reduced amyloid-beta aggregation. This effect was contingent upon both daf-16 and skn-1, highlighting the necessity of these transcription factors in managing proteotoxic stress. These results are comparable to recent findings with 4,4′-methylenediphenol and epigallocatechin-3-gallate, which mitigate Aβ toxicity through similar signaling axes.34,35 However, the unique structural features of fucoxanthin, specifically its allenic bond and 5,6-monoepoxide group, may confer superior hydrophobic interactions with amyloid peptides, potentially inhibiting their assembly into toxic oligomers more effectively than planar polyphenols. In our PD models, fucoxanthin not only restored dopaminergic neuron viability following 6-OHDA exposure but also significantly suppressed α-synuclein accumulation. The complete loss of this neurorestorative effect under daf-16 RNAi further reinforces the critical link between the IIS pathway and neuronal survival. Similar neuroprotective effects have been reported for extracts from Spirulina platensis, which mitigate α-synuclein-induced neurotoxicity. 36 Supplementation with xanthoxyline also showed a neuroprotective effect in a PD model, which was dependent on DAF-16. 14 We initially prioritized daf-16 RNAi over skn-1 RNAi in the 6-OHDA model because the asymmetry of prior literature dictated different confidence levels for the two factors. In the AD paradigm, both DAF-16 and SKN-1 have been independently shown to mediate protection against Aβ proteotoxicity in C. elegans, thus justifying parallel testing.35,37 In contrast, DAF-16 dependence has been established across multiple C. elegans PD models, including 6-OHDA, MPP+, and α-synuclein paradigms, whereas evidence for SKN-1 involvement in dopaminergic neuroprotection has so far been restricted to the metal-toxicity context and to bacterial biofilm-mediated α-synuclein attenuation.14,38–41 To our knowledge, no published study has directly demonstrated a requirement of skn-1 in 6-OHDA-induced dopaminergic neurodegeneration. Accordingly, daf-16 represented the higher-confidence first target in our experimental design. A limitation of the present study is therefore that the requirement of skn-1 in fucoxanthin-mediated dopaminergic neuroprotection was not experimentally addressed. Future work using skn-1 RNAi in the BZ555 background will be required to determine whether SKN-1 acts in parallel with, or downstream of, DAF-16 in this paradigm.
Although our genetic data establish that DAF-16 and bec-1 are individually required for fucoxanthin-mediated longevity and that skn-1 is required in the Aβ-toxicity model, the precise hierarchy among these factors cannot be unambiguously resolved here. DAF-16 and SKN-1 are known to engage in extensive crosstalk in regulating stress-response and autophagy genes, and autophagy can act both downstream of FOXO/Nrf2 signaling and as a feedback regulator of these transcription factors.42–45 We therefore favor a model in which DAF-16 and SKN-1 operate in parallel to converge on bec-1-dependent autophagy as a common effector. Collectively, our study provides compelling evidence that fucoxanthin serves as a multitarget modulator of aging, leveraging the DAF-16/Autophagy/SKN-1 axis to promote longevity and neuroprotection. These findings position fucoxanthin as a promising, high-value marine nutraceutical for mitigating the global burden of age-related neurodegenerative diseases.
Conclusion
In conclusion, fucoxanthin significantly extends the lifespan and health span of C. elegans without reproductive trade-offs, mediated by the coordinated activation of DAF-16/FOXO signaling and autophagy. This mechanism upregulates SKN-1-dependent antioxidant defenses, fortifying the organism against oxidative stress. Furthermore, fucoxanthin exhibits robust neuroprotection in AD and PD models by mitigating proteotoxicity. These findings underscore fucoxanthin’s potential as a multitarget nutraceutical for promoting healthy aging and delaying neurodegenerative decline.
Authors’ Contributions
S.-K.P. designed the study. S.K. conducted the experiment. S.-K.P. and S.K. contributed to data analysis, wrote the first draft, reviewed the article, and approved the final version of the article as submitted.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.