
Abstract
Russian bryozoans and other invertebrate biocenosis material collected in the Arctic Ocean over a period of more than 250 years, and the corresponding literature citing them, are deposited in the Zoological Institute of the Russian Academy of Sciences, Saint Petersburg, Russia. This paper provides a general description and historical quantitative information about the Historical Bryozoan Collection kept in this Russian institute. Access to species-level data is available via the data portal (www.zin.ru). The quantitative distribution of benthos, specifically bryozoans, in the Russian Arctic seas remained nearly unexplored until recent times. The first systematic quantitative studies were conducted by the Zoological Institute in the Laptev Sea, in its southeastern part, on the New Siberian Shoal in 1973; the Chaun Bay of the East Siberian Sea in 1986; and near Wrangel Island and Cape Schmidt of the Chukchi Sea in 1976.
Keywords
Introduction
The history of Arctic research dates back to 1733, with the Great Northern Expedition, on board Expedition and Ob. From then to recent times there have been numerous scientific expeditions (see Table 1 with the most important expeditions). The Zoological Institute of the Russian Academy of Sciences (ZIN RAS) mainly holds specimens collected from 1878 onward, including systematic quantitative studies in the Laptev, East Siberian, and Chukchi seas. The Institution’s collection of extant invertebrates includes bryozoans and the fauna of associated biocenoses.
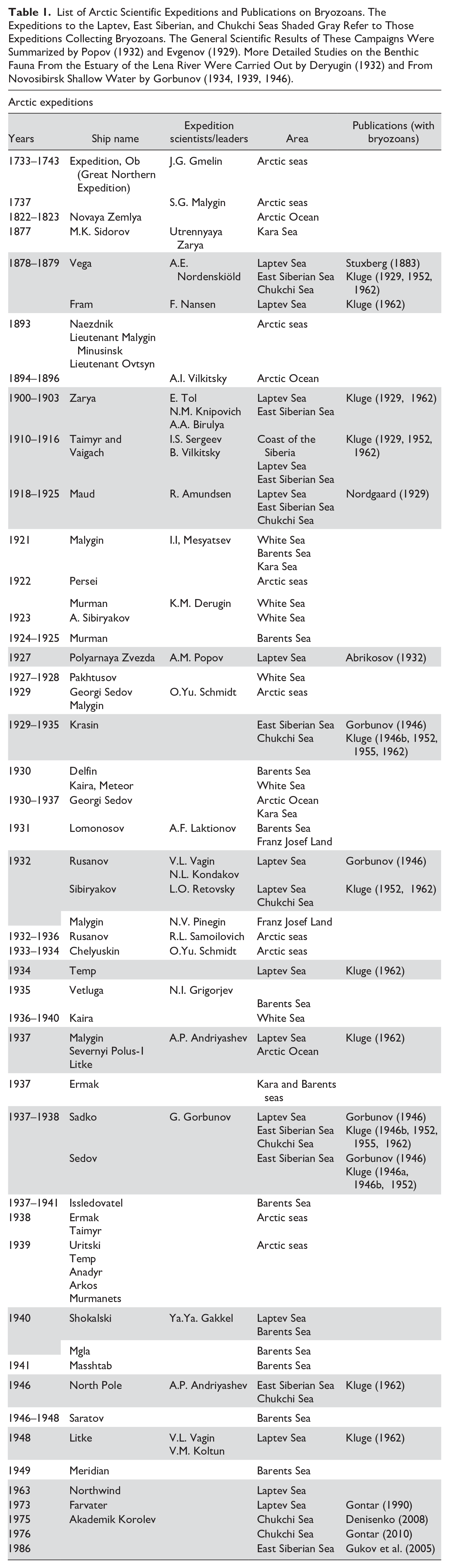
List of Arctic Scientific Expeditions and Publications on Bryozoans. The Expeditions to the Laptev, East Siberian, and Chukchi Seas Shaded Gray Refer to Those Expeditions Collecting Bryozoans. The General Scientific Results of These Campaigns Were Summarized by Popov (1932) and Evgenov (1929). More Detailed Studies on the Benthic Fauna From the Estuary of the Lena River Were Carried Out by Deryugin (1932) and From Novosibirsk Shallow Water by Gorbunov (1934, 1939, 1946).
Bryozoans play an important role in most marine shelf benthic communities and their biocenoses. They are reliable indicators of environmental conditions and their change in time and space. They have a wide distribution across the waters of the world’s ocean and high species diversity. The benthic fauna of the Arctic Ocean and its regional composition reflect its origin and history. Although this fauna is highly autochthonous, several biogeographic subdivisions and Arctic subregions can be recognized. The subregion boundaries are determined by the topography of the seabed and the system of constant oceanic currents of the Northern Hemisphere.
There are numerous works dealing with the Arctic Ocean Bryozoan fauna (Gostilovskaya 1978; Kluge 1962, 2009; Nordgaard 1906, 1912, 1918, 1923, 1929; Norman 1906; Osburn 1913, 1932, 1936; Packard 1867; Smitt 1879; Vigelius 1881–1882, 1884; Waters 1900, 1904 and recent papers by Gontar and Denisenko [Denisenko 2008, 2011; Gontar 1990, 1994, 2010; Gontar and Denisenko 1989]). Data on the bryozoan fauna of the Siberian seas were very scarce until 1962, with 105 species listed in seven works (Abrikosov 1932; Kluge 1929; Levinsen 1886; Nordgaard 1912, 1929; Osburn 1923; Smitt 1879) that were followed by a monograph (Kluge 1962) that it is key to the study of Arctic bryozoans. Currently, the bryozoan fauna consists of more than 240 species.
The extant Arctic fauna can be grouped into three types (Guryanova 1970), which belong to (1) the autochthonous Arctic marine waters; (2) Atlantic Ocean; and (3) Pacific Ocean. Most of Arctic exploration-related collections are deposited in ZIN RAS under the name of the studied region. Most of these collections can be found in the online Systematic Catalog using the subject heading Arctic regions, or the expedition name and specific geographic area. In addition to the general Arctic historical collections, there are three quantitative collections recorded from the Arctic. Access to the species-level data is available via the ZIN RAS data portal, selecting the list of species of free-living invertebrates of Eurasian Arctic seas and adjacent deep waters (https://www.zin.ru/projects/CNAMF/arctlstq.asp), reflecting how the collections are organized, mainly by sampling geography. The bryozoan species dataset is also provided as Supplemental Material.
In this article a general description of the ZIN RAS Arctic Bryozoa Collection and historical collections, which are housed in the Laboratory of Marine Research of the same institution, will be presented with their biocenosis data displayed in maps and charts.
The first systematic quantitative studies were conducted in the southeastern part of the Laptev Sea (New Siberian Shoal), and northeastward of Kotelny and Bennett Islands, between both islands, from July to September 1973 (Table 1). A few years later the Chukchi Sea, near Wrangel Island and Cape Schmidt, was studied in 1976. Finally, Chaun Bay on the East Siberian Sea was explored in 1986 (Figure 1). These expeditions formed the three main Arctic collections that are described below. The upper sublittoral zones were examined by diving to 35 to 40 m depth, using the pyramidal system of benthic algae and invertebrate quantitative inventory (Golikov et al. 1974).
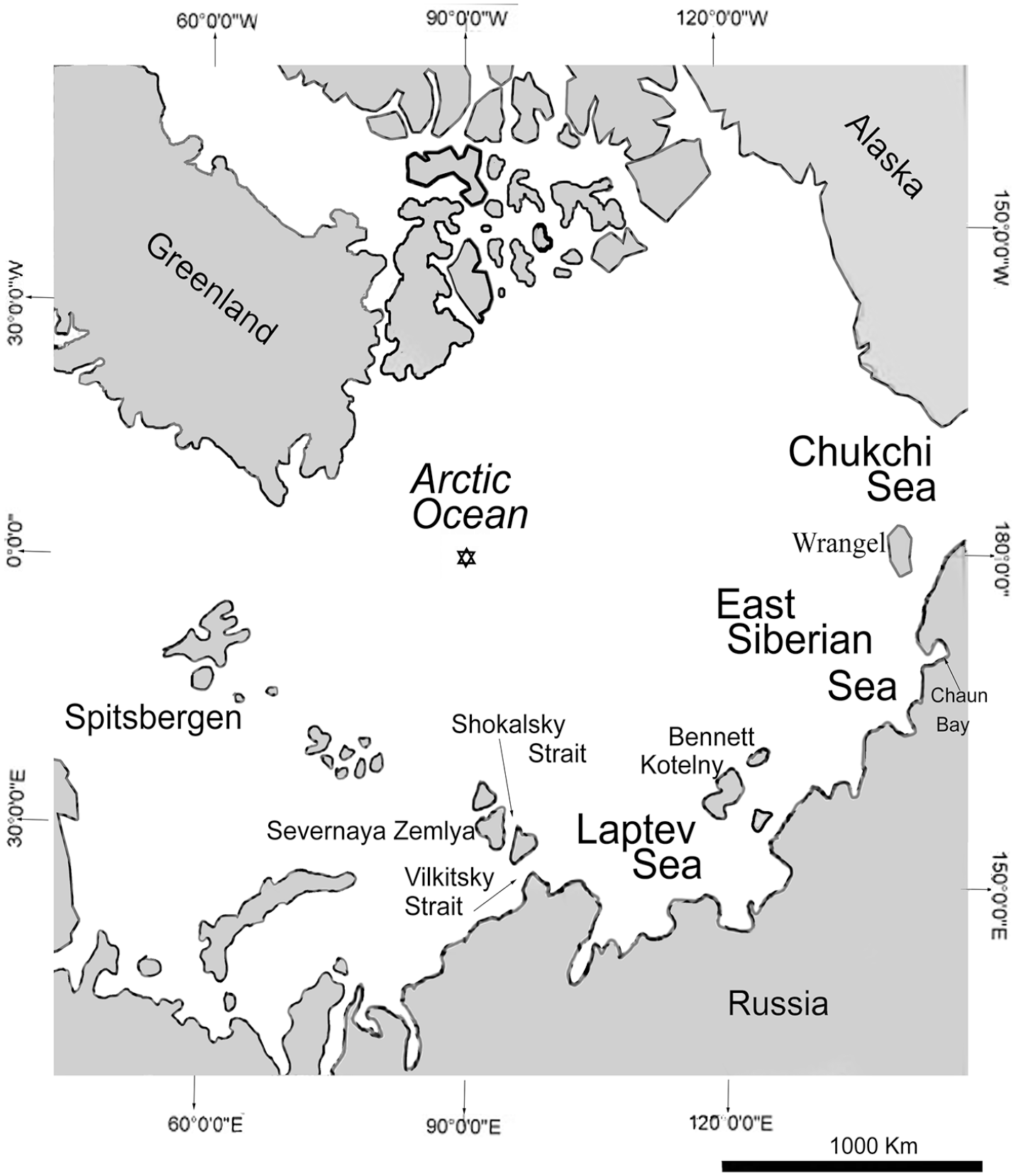
Location of the three Russian systematic quantitative studies, in the Laptev, East Siberian, and Chukchi seas.
The Laptev Sea Collection
Only short descriptions of historical studies are given on the Siberian seas until the first ZIN RAS quantitative diving expedition in 1973. The research into of the Laptev Sea, one of the most inaccessible seas in Russia, has been developed intermittently for more than 120 years (Table 1). The first information about the qualitative composition of the fauna and, to a lesser extent, the flora of the Laptev Sea was obtained after an expedition on board Vega (1878–1879) under the guidance of the Swedish researcher A.E. Nordenskiöld and the famous drift of the ship Fram led by F. Nansen. Bryozoans were recorded at nineteen stations located along the seacoast during the last two years of this expedition.
Significant materials from the Laptev Sea were also collected by the Russian Polar Expedition led by E. Tol on the schooner Zarya (1900–1903). Some of these materials were processed by N.M. Knipovich and A.A. Birulya. Further collection sampling from the Laptev Sea was carried out by the Hydrographic Expedition on Taimyr and Vaigach (1914–1916) under the leadership of B. Vilkitsky and an expedition led by R. Amundsen on board Maud (1918–1925). The next step in the study of the fauna of this sea was made thanks to A.M. Popov’s sampling in the southeastern part of the sea and at the mouth of the Lena River during the Yakut expedition of the Academy of Sciences of Russia on board Polyarnaya Zvezda (1927). More material was accessioned by ZIN RAS coming from the icebreaker Rusanov with a more detailed survey of the western part of the sea in the areas of the Vilkitsky and Shokalsky straits in 1932, with V.L. Vagin and N.L. Kondakov. In the same year, the icebreaker Sibiryakov provided more samples near Severnaya Zemlya and the southern part of the Laptev Sea compiled by L.O. Retovsky. Much smaller samples were also collected in the southern part of the sea on the vessel Temp in 1934. A few years later there was a significant addition to the Laptev Sea benthic fauna made by G. Gorbunov on board Sadko (1937–1938), A.P. Andriyasheva on Malygin (1937), V.L. Vagin and V.M. Koltun on Litke (1948), and the American expedition on Northwind (1963).
Benthic fauna was mainly collected using the quantitative diving technique in the southeastern part of the Laptev Sea, on the New Siberian Shoal and northeastward of Kotelny and Bennett islands from July to September 1973. The work was done on board the schooner Farvater and the lugger Zenith from the Tiksi Hydrobase. Quantitative samples were also collected using a Petersen seabed grab sampler, with a grip area of 0.025 sq m, and a Sigsby trawl in the coastal continental waters, where there is high turbidity—brown water—with less than 0.3 m transparency. The lack of visibility reduces diving efficiency. On hard substrates with zero visibility, samples were collected blind, using only a seabed grab sampling an area of 0.05sq m. The quantitative diving technique was used in sufficiently clear water, though visibility remained limited near the seabed. At the same time, measurements of homogeneous biotope areas were made. Large and uncommon organisms were collected from three differentiated areas of 5 sq m each along a measuring halyard or from frames of 1 sq m in area. Furthermore, three samples with a surface of 0.1 sq m were collected from each biotope region using a frame or a diver’s seabed grab. Some bibliographical references suggest the presence of biocenosis and decreased salinity levels of the area, evidencing river discharge, with salmon migrating to this area to feed.
Stuxberg (1883) recorded bryozoans at nine stations of the Laptev Sea, stations numbers 66–67, 70, 74–76, 79, 83–84. Station 66 had bryozoans associated with worm tubes. Station 67 is significant for its high bryozoan species diversity, the most abundant being Infundibulipora lucernaria (recorded as Defrancia lucernaria). Stations 74 and 79 had mainly Alcyonidium mamillatum, which was recorded at the Khatanga River southeast estuary. Stuxberg (1883) described an Alcyonidium-formation with silt or coarse sand at 4 to 6 feet depth and seabed of Station 74. Stations 83 and 84 had freshwater bryozoans encrusted on wood. Station 84 had an undetermined species of Alcyonidium.
In 1929 Kluge published a paper on the bryozoans collected by the Nordenskiöld expedition on board Vega, the Russian Polar Expedition on Zarya and the Hydrographic Expedition on Taimyr and Vaigach and listed eighty-five bryozoan taxa (species and varieties with outdated taxonomic names) belonging to three bryozoan orders (Cyclostomatida, Ctenostomatida, Chelostomatida). The same year Nordgaard (1929) published his research on the samples collected by the Norwegian Polar Expedition on board Maud and cited three bryozoan species for the Laptev Sea at Station 29 at 23 m depth: (1) Eucratea loricata, which was attached to Portlandia arctica and Serripes groenlandicus valves (Bivalvia); (2) Serratiflustra serrulata (see Kluge 2009); and (3) Rhamphostomella bilaminata on the burrowing cardiid S. groenlandicus.
Later, Abrikosov (1932) described ten cheilostome species collected at seven stations of the Yakut expedition carried out by the Hydrobiological Working Group in 1927. Some bryozoan species, mainly on polychaete tubes, were found on silty ground with high salinity (more than 28‰).
The first quantitative collection of the ZIN RAS expedition in 1973 (Gontar 1990) found twelve bryozoan taxa (species and subspecies), the subspecies new to the fauna of this region. For instance, Alcyonidium gelatinosum was found with the Balanus crenatus+Suberites domuncula+Eucratea loricata biocenosis at 12 to 18 m depth, with the highest colony density (1.16 ± 0.45 colony spec/sq m). The highest biomass (10 ± 3.53 g/sq m) was reached by the cited bryozoan species in the Musculus glacialis+Suberites domuncula biocenosis at 12 m depth. The cheilostome species Eucratea loricata was found in the New Siberian Shoal in the Saduria entomon biocenosis at a depth of 7−8 m with the highest colony density of 70 ± 24.7 colony spec/sq m and the highest biomass of 44 ± 15.5 g/sq m. On the other hand, Alcyonidium disciforme was recorded in association with Portlandia arctica and Rhizomolgula globularis at 4.8 m depth with the highest colony density of 40.2 ± 1 colony spec/sq m and the highest biomass of 14.5 ± 4.72 g/sq m in the nuculoid Ennucula tenuis (previously Leionucula bellotii) biocenosis (Figure 2).
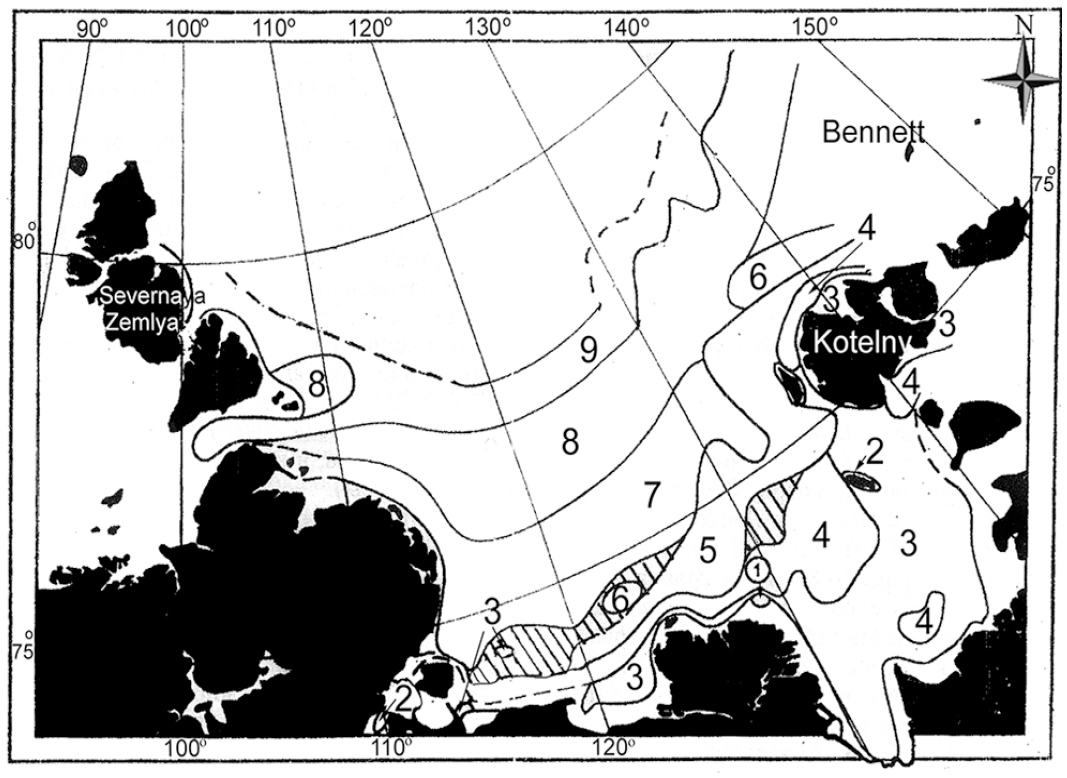
Biocenoses of the Laptev Sea shelf seabed (>10 m depth). 1. Alcyonidium disciforme. 2. Suberites domuncula. 3. Portlandia arctica. 4. Astarte borealis+A. montagui+Portlandia arctica. 5. (Leionucula) Ennucula tenuis. 6. Ocnus glacialis. 7. Astarte borealis (+Astarte montagui+Maldane sarsi). 8. Ophiocten sericeum. 9. Ophiopleura borealis+ Ophiocten sericeum+Ophiacantha bidentata. Areas with insufficient data are in black (after Petryashev et al. 2004).
The East Siberian Sea Collection
The first collections of bryozoans from the East Siberian Sea (Table 1, Figure 1) were made by A. Nordenskiöld on board Vega along the coast of Siberia in 1878 to 1879. Taxonomic work on these collections was published by Stuxberg (1883), who only recognized two bryozoan taxa (Alcyonidium sp. and Carbasea carbasea? as Flustra carbasea?). Later, Nordgaard (1929) published a list of eighteen bryozoan species after studying the material of the Norwegian expedition on Maud in the northern part of the Siberian Sea during the drift in 1918 to 1925. A publication by Nordgaard, published in the same year, with the bryozoan fauna identified from several expeditions, such as the Swedish one on Vega (1878–1879) and the Russian ones on Zarya (1900–1903), and Taimyr and Vaigach (1913–1915), was published by Kluge (1929).
Research on benthic fauna, including bryozoans, from the East Siberian Sea was carried out thanks to the support of the Russian polar expeditions. In the period between the two World Wars of the twentieth century, zoological works were taken on the icebreaker Krasin in the eastern part of the sea (1935) and on the icebreaker Sadko (1937–1938), which drifted for two years, which Sedov (1937–1938) also did in the northwestern part of the sea and in New Siberia shoals. However, the results of these expeditions were published almost a decade later, after the Second World War (Gorbunov 1946; Kluge 1946a, 1946b, 1952). Kluge (1946b, 1955) described seventeen new taxa (including taxonomic names for species and subspecies that have not been updated) from the East Siberian Sea with material of Sadko. In the case of Gorbunov (1946), he published a list of eighty-nine bryozoan species in a general article of the zoobenthos of the Arctic region.
In the first post-wars years, under the leadership of A.P. Andrijashev, an expedition was carried out to the North Pole (1946). These samples were partly studied by G.A. Kluge and the results were included in his monograph “Bryozoans of the Northern Seas of the USSR” (Kluge 1962), which also included all previously published information about bryozoans of the East Siberian Sea.
The first systematic quantitative studies conducted in the East Siberian Sea region were carried out in July–August 1986 by ZIN RAS. A total of twenty-one bryozoan taxa (species and subspecies, the taxonomic names of which have not been updated) were found in Chaun Bay (Figures 1 and 3). Only five species, one subspecies and one form have already been recorded from this region, the rest of the taxa being new records for the East Siberian Sea.
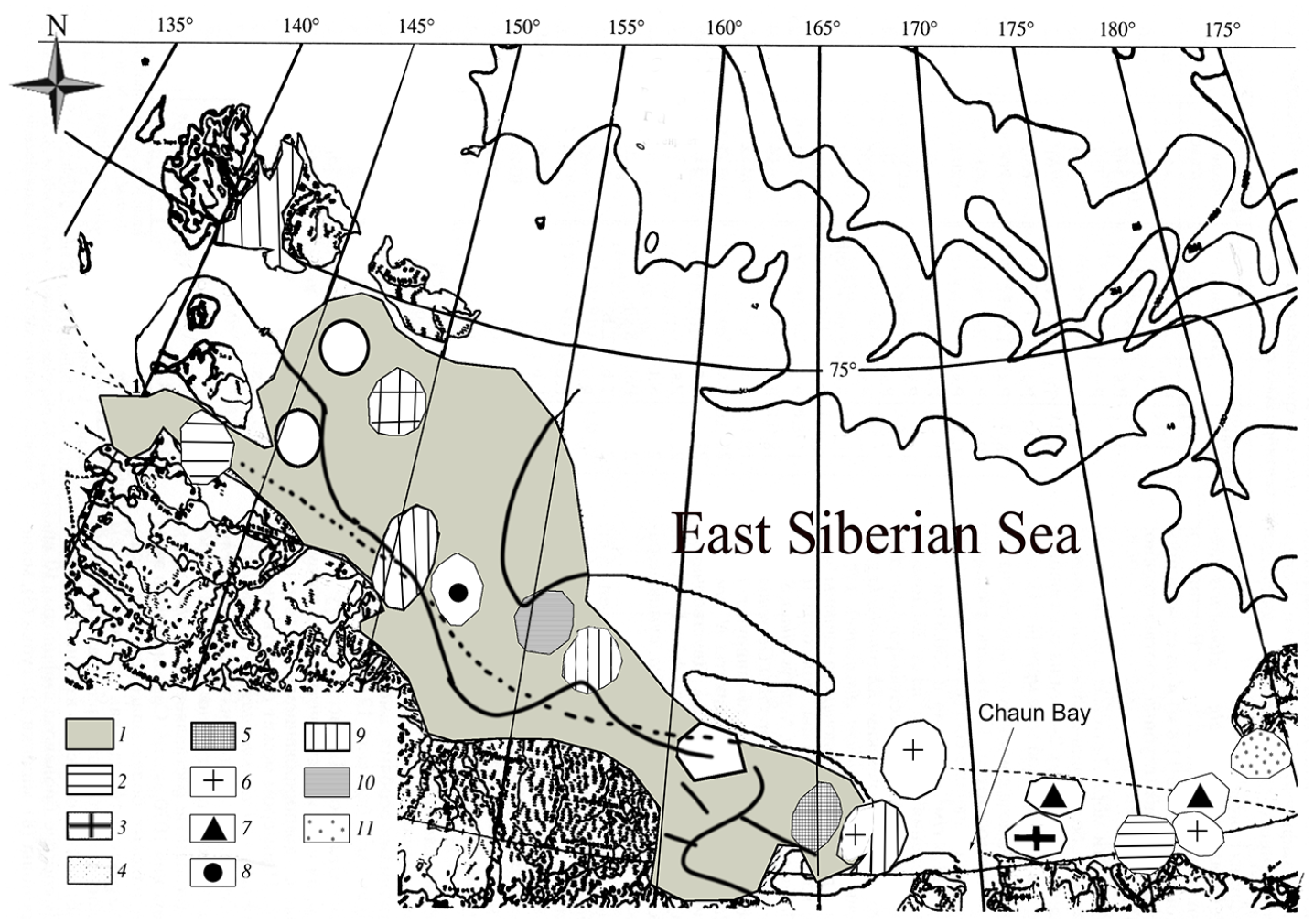
Distribution of seabed biocenosis within sublittoral ground (8–50 m) of the East Siberian Sea. 1. Portlandia siliqua. 2. Rhizolmolgula globularis. 3. Portlandia siliqua +Polychaeta. 4. Astarte borealis+A. montagui+ Portlandia siliqua. 5. Portlandia siliqua+ Ennucula tenuis. 6. Astarte borealis+ Ennucula tenuis. 7. Ennucula tenuis. 8. Polychaeta+Saduria sibirica. 9. Nephtys longosetosa. 10. Portlandia siliqua+ Saduria sibirica+Saduria sabini. 11. Ocnus glacialis (after Gukov et al. 2005).
The analysis of bryozoan vertical distribution in Chaun Bay shows the maximum number of species (15) and the maximum total biomass at 5 to 10 m depth. In the case of the Rhizolmolgula globularis biocenosis, this was found on a sandy seabed near Cape Billings in the Long Strait at a depth of 11.8 m and in the Dmitry Laptev Strait at a depth of 10.2 and 13.0 m (Gukov et al. 2005) (Figure 2). The dominant species was Alcyonidium disciforme. In the southern part of the Laptev Sea, this biocenosis is usually distributed at depths of 2 to 19 m on sandy silt and sand in the freshened part of the sea.
The Ennucula tenuis biocenosis within the New Siberia Polynya on sandy silt and silty ground at depths from 22.5 to 24.5 m had the richest species composition (Figure 4). The following bryozoan species are present: Eucratea loricata (frequency of occurrence 20 col/sq m, biomass 1.2 g/sq m), Alcyonidium disciforme (frequency of occurrence 40 col/sq m, biomass 0.7 g/sq m), Escharella ventricosa (frequency of occurrence 10 col/sq m, biomass 0.08 g/sq m), Leieschara subgracilis (as Myriapora subgracilis) (with 20 col/sq m, 0.03 g/sq m). In the East Siberian Sea, this biocenosis was found at several stations on silty ground and silt with stones, at depths of 27.5 to 49.0 m, and five bryozoan species were found in its composition. On the traverse of Cape Blossom (Wrangel Island), Eucratea loricata and Alcyonidium gelatinosum were found in the Ocnus glacialis biocenosis.

Benthic Ennucula tenuis biocenosis. (A) General view. (B) Species present in the biocenosis. 1. Ampelisca sp. 2. Gammarus setosus. 3. Mertensia ovum. 4. Tubularia indivisa. 5. Diastylis glabra. 6. Amauropsis islandica. 7. Eucratea loricata. 8. Myriotrochus rinkii. 9. Suberites domuncula. 10. Onisimus sibirica. 11. Saduria entomon. 12. Rhizomolgula globularis. 13. Semisuberites cribrosa. 14. Mysis oculata. 15. Saduria sabini. 16. Balanus balanus. 17. Ennucula tenuis. 18. Astarte montagui. 19. Portlandia siliqua. 20. Cryptonatica affinis. 21. Priapulus caudatus. 22. Astarte borealis. 23. Scoloplos armiger. 24. Nereis zonata. 25. Astarte crenata (after Gukov 1999).
Naturally, there is an increase in benthos biomass from the shallow waters of large river estuaries to deeper seabed with an increase in salinity and a decrease in temperature.
The Chukchi Sea Collection
Chukchi Sea bryozoans were studied by numerous Russian and international researchers, some of the classic publications are those of Stuxberg (1883), Osburn (1923, 1950, 1952, 1953, 1955), Nordgaard (1929), Powell (1968), Kluge (1952, 1962), Gontar and Denisenko (1989), Gontar (2001, 2004, 2015a, 2015b, 2016), and Denisenko and Kuklinski (2008).
The first effort to collect bryozoans of the Chukchi Sea region (Table 1, Figure 1) was undertaken by the Swedish expedition on Vega (1878–1879). The Siberian coast of the Chukchi Sea was studied between 1875 and 1878 and the first publication citing bryozoans was written by Stuxberg (1883) with Alcyonidium mamillatum (from the Russian marine stations 96 and 97). The bryozoan samples from the Vega expedition were delivered to ZIN RAS where they were subsequently identified by G.A. Kluge. In 1929, Nordgaard published a bryozoan list including those bryozoans collected by the Norwegian Polar expeditions on board the vessel Maud (1918–1925). Unfortunately, he only recorded three bryozoan species from the northern part of the Chukchi Sea: Eucratea loricata, Serratiflustra serrulata, and Myriozoella crustacea.
There were more scientific expeditions which studied the Chukchi Sea (Table 1) such as those on Chelyuskin and Krasin (Figure 5). The former was a scientific research vessel that sank in the Chukchi Sea in February 1934 while attempting to sail the Northern Sea Route from Murmansk to the Bering Strait and on to Vladivostok. Krasin was the first icebreaker to reach the northern shores of Novaya Zemlya. One of the researchers involved in Arctic fauna exploration was Pavel Vladimirovich Ushakov (Figure 5) who was on board Krasin and observed of the different faunas between the Chukchi and Bering seas. He worked for ZIN RAS for forty years, from 1945.

On the left, the Chelyuskin and Krasin ships. Image taken in 1933, from the World Digital Library (Library of Congress Control Number 2018683523). On the right Pavel Vladimirovich Ushakov (1903–1992) who 1935, on board Krasin. Image courtesy of ZIN RAS.
The most detailed and comprehensive bryozoan study in the Chukchi Sea, and in the Arctic in general, was carried out by the Russian bryozoologist G.A. Kluge. Kluge (1952) published a list of 123 bryozoan taxa (species and subspecies) from the Chukchi Sea and the Bering Strait, of which fifteen were new bryozoan species. Six species were previously cited only from the Bering Strait. Kluge’s research was based on all the specimens collected in the previous Arctic expeditions up to his time (Vega, 1878–1879; Taimyr and Vaigach, 1910–1914; Sibiryakov, 1902; Dalnevostochnik, 1932; Red Army, 1933; Krasin, 1935; Okhotsk, 1938), and Kluge and Gekker’s private collections made near Wrangel Island in 1938.
On the other hand, Osburn (1923) published a list of sixteen species of bryozoans from two marine stations at the northwest coast of Alaska; and Nordgaard (1929) wrote about three bryozoan species from two stations at north of Wrangel Island (Figure 1). In 1975 benthic samples were collected along the Siberian Coast by the expedition conducted on Akademik Korolev.
The Bryozoa fauna in Chukchi Sea is formed under Arctic conditions, influenced by the Pacific fauna via the Bering Strait. In the first quantitative collections of the ZIN RAS expedition in July-August 1976 near Wrangel Island and Cape Schmidt, twenty-seven bryozoan species and subspecies were found. Four species were new to the Chukchi Sea fauna. The highest bryozoan diversity near Wrangel Island was recorded at relatively shallow depths, from 3 to 6 m. This is probably due to the development of seabed plants at these depths. Along the coast of the Chukotka Peninsula, a cold Chukchi Current emerges in summer, and it is constant during autumn and winter, carrying freshened waters of the East Siberian Sea from the southwest to the Bering Strait. These freshened waters are cold in summer and relatively warm in winter (temperature is about −1.6°C). According to the zoologist L.I. Moskalev (see Gontar 2010), amongst the most important forms in the seabed biocenosis of the external Chukchi Sea shelf, more than a third are sestonophagous and there are at least three bryozoan species. The map of distribution of food groups and benthic biocenosis (Figure 6) shows that bryozoans play an important role in at least two of the food groups.
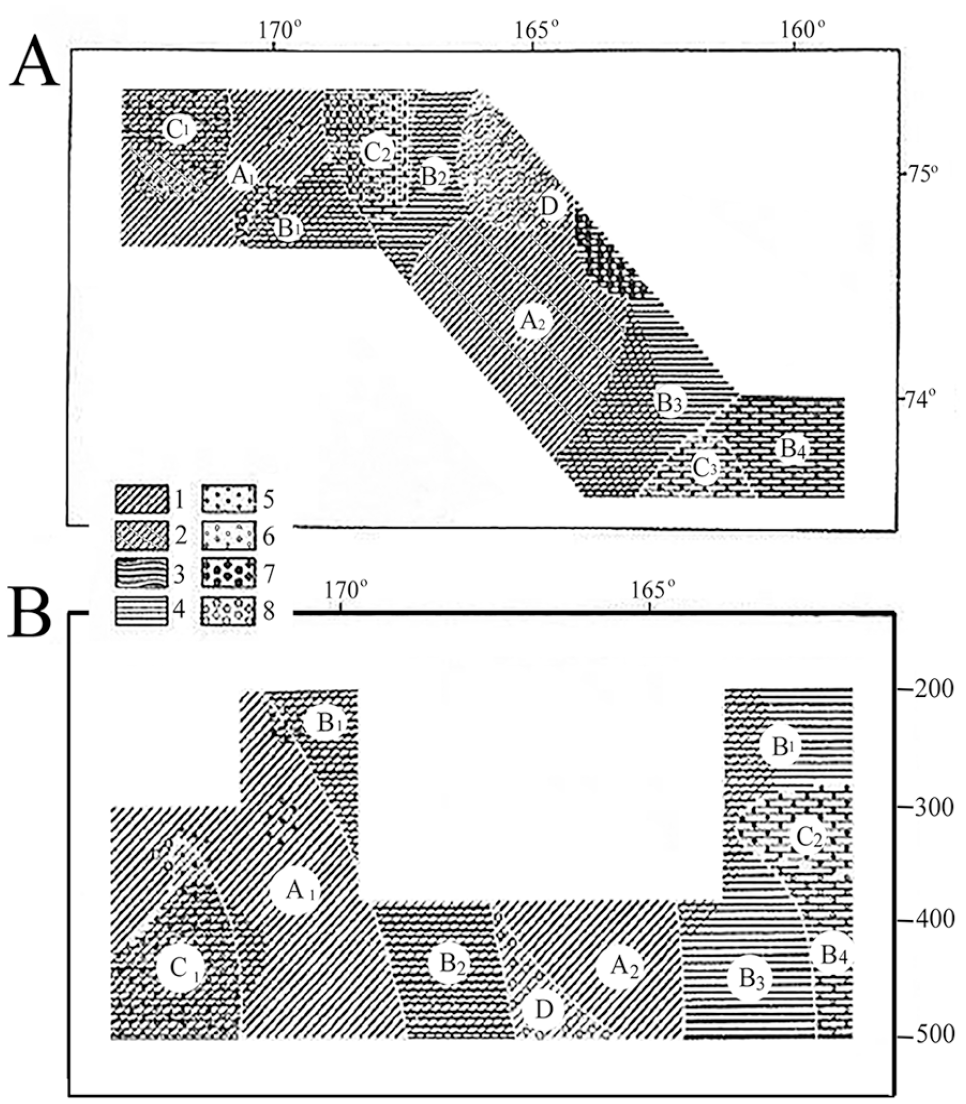
(A) Distribution of food groups. (B) Benthos biocenosis in the northern Chukchi Sea shelf. Food groupings (in percent of the total weight of the trawl catch). Sestonophagous: 1. at >50 percent; 2. <50 percent. Detritivores: 3. at >50 percent; 4. <50 percent. Predators/carnivores: 5. at >50 percent; 6. <50 percent. Organisms with an indeterminate type of nutrition: 7. at >50 percent; 8. <50 percent. Biocenosis: A1-Umbellula-Heliometra-Bryozoa-Jasmineira-Gersemia; A2-Gersemia; B1-Molpadia-Ctenodiscus; B2-Astarte; B3-Ctenodiscus-Pontaster; B4-Kolga-Portalesia; C1-Anthosactis-Bryozoa; C2-Anthosactis-Pontaster; C3-Anthosactis-Ctenodiscus-Pontaster; D-Ophioscolex (after Gontar 2010).
The study of this fauna has led to the conclusion that bryozoans are often dominant in faunal associations, which makes them important indicators based on their thermopathy, halinopathy, and living depth. On the other hand, they are also biogeographic markers in the regionalization of the Laptev shelf, East Siberian, and Chukchi seas.
A relatively recent comparative analysis of the research carried out in Laptev Sea and East Siberian Sea (Evseeva 2021) at the end of the twentieth century (1986, 1987, and 1993–1998) based on literature data (Gontar 1990, 1994, 2004, 2015a, 2015b, 2016) concludes that there has been a significant increase in the proportion of Boreal-Arctic species in the Arctic Circle due to a significant decrease of Arctic species (by almost a third), which probably reflects climate change resulting from global warming. An increase in abundance of uncalcified (soft-bodied) species of the bryozoan genus Alcyonidium (found in numerous biocenoses on soft grounds and in freshened water masses, these ones occupy large parts of the seas) has also been observed in the Laptev, East Siberian, and Chukchi seas compared to other regions. This confirms the hypothesis that under low salinity and low temperature conditions, it is difficult for invertebrates to extract calcium from water (Akhmetchina 2018), which inhibits the development of calcareous bryozoans, but is probably favorable for the development of soft-bodied species (like Alcyonidium). According to Evseeva (2021), the bryozoan branching bushy colonies are increasing in the Laptev Sea by more than half (58 percent, twenty-six species) and in the north-western region of the East Siberian Sea by three quarters (75 percent, twelve species) of the forty-eight bushy bryozoan species recorded. Bryozoans which raise above the substrate in colonies are developed mainly in biotopes with intense sedimentation and moderate hydrodynamics.
Conclusions
The journey of more than 120 years through the history of scientific expeditions in the Arctic Circle, and the study of the invertebrate fauna collected in these expeditions, confirm a significant increase in the proportion of eurybiont Boreal-Arctic species in the Arctic Circle during the few last decades. Bryozoans have demonstrated that they play one of the leading roles in the benthic fauna from Arctic high latitudes. The quantitative samples carried out in the Arctic, mainly in the Laptev Sea, have allowed us to define and identify spreading trends of Atlantic origin for species in the upper shelf regions. The spread of these species presumably depends on the hydrological regime which has a quantitative and qualitative effect on the extant biocenoses of the studied Arctic seas.
Supplemental Material
sj-csv-1-cjx-10.1177_15501906231159042 – Supplemental material for A Short Research Guide on Arctic Historical Bryozoan Collections and a Few Associated Biocoenosis at the Zoological Institute of the Russian Academy of Sciences
Supplemental material, sj-csv-1-cjx-10.1177_15501906231159042 for A Short Research Guide on Arctic Historical Bryozoan Collections and a Few Associated Biocoenosis at the Zoological Institute of the Russian Academy of Sciences by Valentina I. Gontar in Collections: A Journal for Museum and Archives Professionals
Footnotes
Acknowledgements
I am very grateful to Consuelo Sendino (NHM) for assisting with this article; the late Aleksey Alexandrovich Golikov, son of the former head of the Marine Research Laboratory; and head of the Marine Research Laboratory the late Alexander Nikolaevich Golikov, who provided the expedition datasets.
Declaration of Conflicting Interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was conducted on the State Topic no. 122031100274-7 for 2022-2024.
Supplemental Material
Supplemental material for this article is available online.
Author Biography
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.