
Abstract
Beyond enabling survival in extreme environments, tardigrades open new avenues for innovation across scientific fields. This review examines the physiological and molecular mechanisms underlying tardigrade tolerance, namely cryptobiosis, damage-suppressor proteins, intrinsically disordered proteins, trehalose accumulation, and robust DNA repair pathways. Furthermore, it highlights their potential applications in cryopreservation, radioprotection, astrobiology, biotechnology, and medicine. In cryobiology, tardigrade-derived molecules offer strategies to improve the viability of cells, tissues, and organs during freezing and thawing. Their natural radioprotective mechanisms may inform the development of novel cancer therapies and advanced shielding materials. Insights into tardigrade survival in the vacuum of space provide valuable models for life-support systems and planetary protection in long-duration missions. Moreover, engineering tardigrade proteins into microbial or human cells holds promise for enhanced stress tolerance in industrial bioprocessing and therapeutic contexts. By unravelling these unique survival strategies, researchers can leverage tardigrades as a blueprint for designing next-generation solutions to pressing challenges in human health, food security, and space exploration.
Introduction
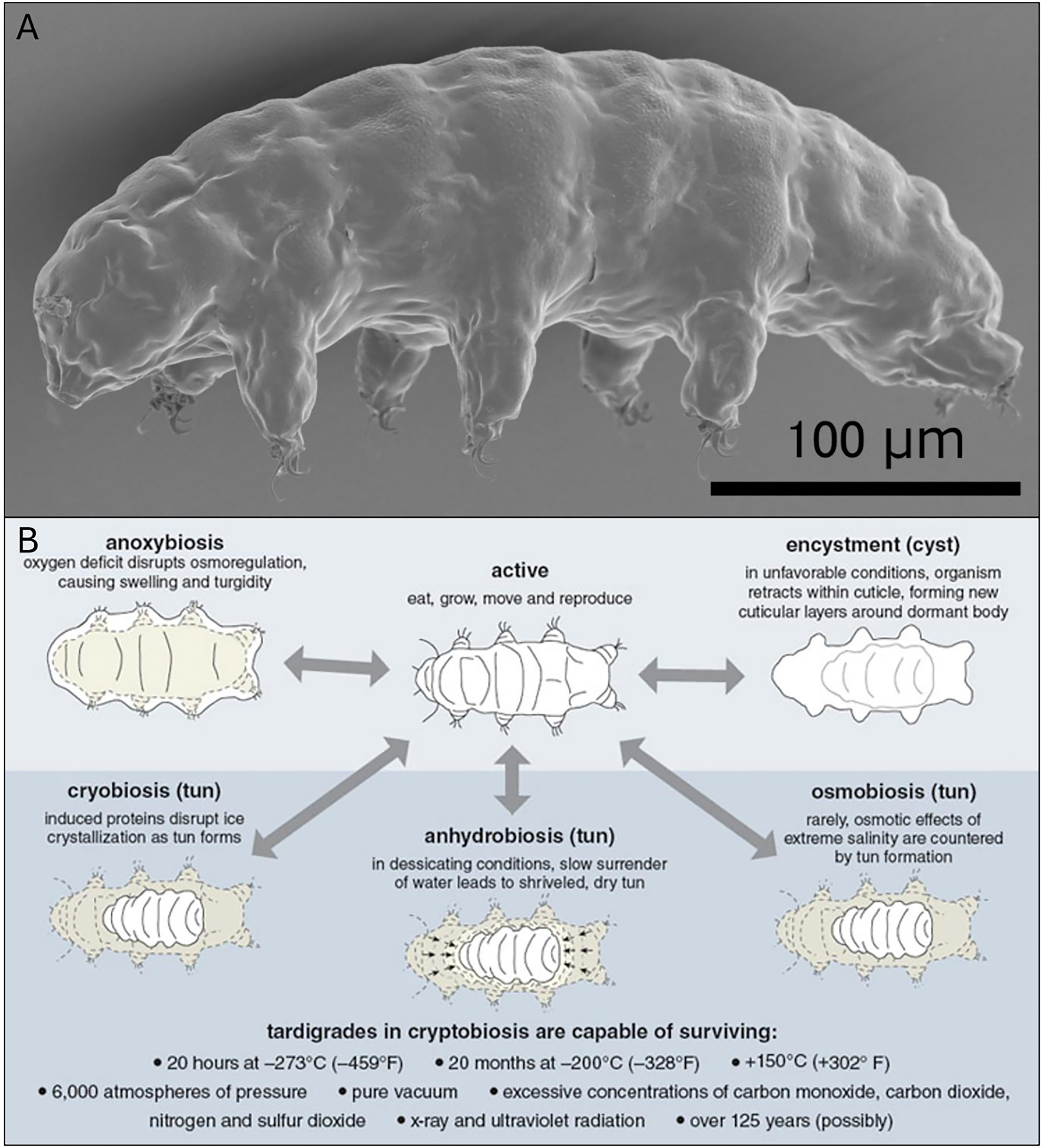
Tardigrades, often referred to as water bears, have attracted considerable attention due to their extraordinary ability to survive and adapt to extreme environmental stresses (Fig. 1a).3–5 Tardigrades have a broad ecological distribution and are considered cosmopolitan, inhabiting all over the world, including soil, sand, freshwater, marine sediments, and hot springs. 1 Most feed on plant fluids using a needle-like mouth, though some are predators or detritivores. They exhibit specialized adaptations that enable them to survive fluctuations in humidity and extreme temperature conditions.1,6 Among their most remarkable traits is the ability to endure environmental stressors such as desiccation, freezing, and radiation, that would be fatal to most other organisms. During anhydrobiosis, tardigrades can withstand extreme conditions, including temperatures ranging from −272°C (−457°F) to over 150°C (302°F) for up to 15 minutes, intense ionizing radiation, pressures reaching 600 MPa, and even the vacuum of space.7–13 This remarkable characteristic is owing to their unique adaptation called cryptobiosis or anabiosis, a reversible dormant phase in which metabolic activity nearly ceases, a trait found in some extremophiles. Furthermore, during this phase, tardigrades produce protective molecules known as tardigrade-specific intrinsically disordered proteins (TDPs), which help stabilize their cellular structures and protect them from damage. 14 Trehalose, a naturally occurring disaccharide composed of two D-glucose units, 15 is another important biomolecule that aids tardigrade survival. However, only a few tardigrades accumulate trehalose during desiccation; this is not universal across the phylum; many species rely predominantly on TDPs (e.g., cytoplasmic abundant heat-soluble [CAHS], secretory abundant heat-soluble [SAHS], mitochondrial abundant heat-soluble [MAHS]) rather than trehalose as their primary anhydrobiotic protectants. Trehalose stabilizes their biological processes, enabling them to withstand freezing temperatures and desiccation. 16 Similarly, brine shrimp embryos accumulate trehalose up to 15% of their dry weight before entering hibernation, which is essential for their survival. 17 Another important survival strategy of tardigrades is their efficient DNA repair mechanisms during radiation exposure. Studies show that specific proteins facilitate repair and recovery from radiation-induced DNA breaks, which are typically lethal in other species.1,18 Damage suppressor (Dsup), a unique protein in tardigrades, has been shown to stabilize cellular structures and protect against oxidative stress and radiation, thereby significantly enhancing their survival. These remarkable features possess unique evolutionary adaptations and make them ideal model organisms for studying how cells respond to stress and damage. 14
Beyond enabling survival in extreme environments, their unique adaptations offer critical insights relevant to biotechnology and astrobiology. Current research highlights their distinctive position at the intersection of ecology, evolutionary biology, and applied science, underscoring their significance as model organisms for understanding extreme survival. This review delves into their exceptional properties, specialized dormancy mechanisms such as cryptobiosis, and molecular adaptations, including protective proteins and DNA repair pathways. Furthermore, it examines how these remarkable survival strategies are driving innovation in diverse fields, including biotechnology, medicine, space exploration, and environmental monitoring, offering promising avenues for future scientific and technological advancements.
State of Dormancy
With over 1200 species discovered so far, tardigrades inhabit a wide range of ecosystems, from freshwater habitats to extreme environments such as mountain tops, deep-sea trenches, and tropical forests, and are even found in Antarctica and Arctic soil.19–22 Although many species are terrestrial or freshwater dwellers, most depend on a thin film of water surrounding their bodies to facilitate gas exchange and prevent desiccation. Tardigrades enter anhydrobiosis, a specific type of cryptobiosis, in response to harsh conditions, progressing through dehydration, a desiccated “tun” stage, and rehydration. Several experiments on tardigrade anhydrobiosis have shown that survival varies with species, the duration of the tun stage, and environmental conditions. Milnesium inceptum and Echiniscus testudo show high survivability up to ca. 20 years, while Hypsibius exemplaris displays low rates even after short tun stages. Studies indicate geographic variation, body size, and energy reserves influence recovery. Some factors affecting anhydrobiosis survival in tardigrades have been studied, while others remain poorly understood. Although age is often cited as influential, one study found no significant effect. 23 Survival is known to decrease with longer tun durations and repeated anhydrobiosis cycles, likely due to the accumulation of cellular damage. However, the combined effects of episode frequency and duration, as well as the roles of sex and group presence, remain unexplored. To address these gaps, a study on Paramacrobiotus experimentalis evaluated the influence of sex, age, group presence, and both the number and duration of tun episodes, including their interactions.24,25 Furthermore, tardigrades survive extreme stress through dormancy, enduring desiccation, freezing, oxygen deprivation, high salinity, and even space conditions by drastically reducing metabolic activity. 26 They achieve this via cryptobiosis, a reversible metabolic shutdown triggered by chemical signaling, and diapause, which involves encystment, cyclomorphosis, and the production of resting eggs. 27 While cryptobiosis and diapause are distinct, both are critical survival strategies that enable tardigrades to withstand harsh environments.
Diapause in tardigrades involves significant morphological and physiological changes in response to environmental stressors, allowing them to suspend development and metabolic activity as a survival strategy. 28 This programmed developmental arrest is associated with processes such as encystment, cyclomorphosis, and the production of resting eggs, all of which enhance their ability to withstand extreme conditions (Fig. 1b). Encystment is common among freshwater species and involves forming protective cysts by shedding the old cuticle and producing multiple new ones, thereby reducing metabolism and increasing resistance to stressors such as desiccation and temperature fluctuations.26,29 For instance, the eutardigrade, Amphibolus volubilis produces two types of cysts: type 1 in cold seasons and type 2 in warm seasons, indicating the importance of temperature in cyst formation. Moreover, cyclomorphosis refers to seasonal, cyclic changes in morphology and physiology tied to molting, enabling marine tardigrades to survive challenges such as freezing by entering freeze-tolerant stages.26,29 In addition, tardigrades produce resting eggs during diapause that can endure harsh conditions and only hatch in response to specific environmental cues, such as rehydration after desiccation, ensuring species survival even in highly unpredictable environments.
CRYPTOBIOSIS
Cryptobiosis represents an extreme survival mechanism in which tardigrades reduce their metabolism to undetectable levels to endure harsh environmental stressors. Depending on the specific stressor, cryptobiosis can occur in four distinct forms. 5 Anhydrobiosis occurs in response to desiccation, during which tardigrades retract their legs, expel internal water, and enter a tun state, forming a barrel-shaped structure that reduces surface area. In this phase, they synthesize trehalose and TDPs (tardigrade-specific TDPs) to stabilize cellular structures during dehydration. 30 Cryobiosis allows tardigrades to survive subzero temperatures by preventing intracellular ice formation, a process aided by the accumulation of cryoprotectants such as glycerol, which help preserve cellular integrity. 31 Anoxybiosis occurs under oxygen-deprived conditions; unlike other forms, the tun state is not formed. Instead, tardigrades absorb water to maintain osmotic balance and become immobile, surviving until oxygen becomes available again.26,32 Osmobiosis is a response to high salinity, in which tardigrades adjust their internal osmotic balance by producing osmo-protectants, such as trehalose and TDPs, to sustain cellular function in hyperosmotic environments. 30 Moreover, a study by Hvidepil and N. Møbjerg argues that additional forms of cryptobiosis (such as chemobiosis) may exist beyond the traditionally cited four types. Chemobiosis refers to the condition induced by exposure to toxic chemical stressors, as demonstrated by E. sigismundi entering the tun state upon exposure to 2,4-Dinitrophenol (DNP). This finding shows that chemical toxicity alone can trigger latent-life pathways, expanding the known conditions that induce cryptobiosis. 33
While both diapause and cryptobiosis are dormancy strategies, they differ in purpose and onset. Diapause is a preprogrammed developmental arrest triggered by environmental cues, whereas cryptobiosis is an immediate physiological response to extreme conditions such as desiccation or freezing. Depending on the species and environment, these strategies may occur independently or simultaneously.26,29
MOLECULAR AND GENETIC PERSPECTIVE
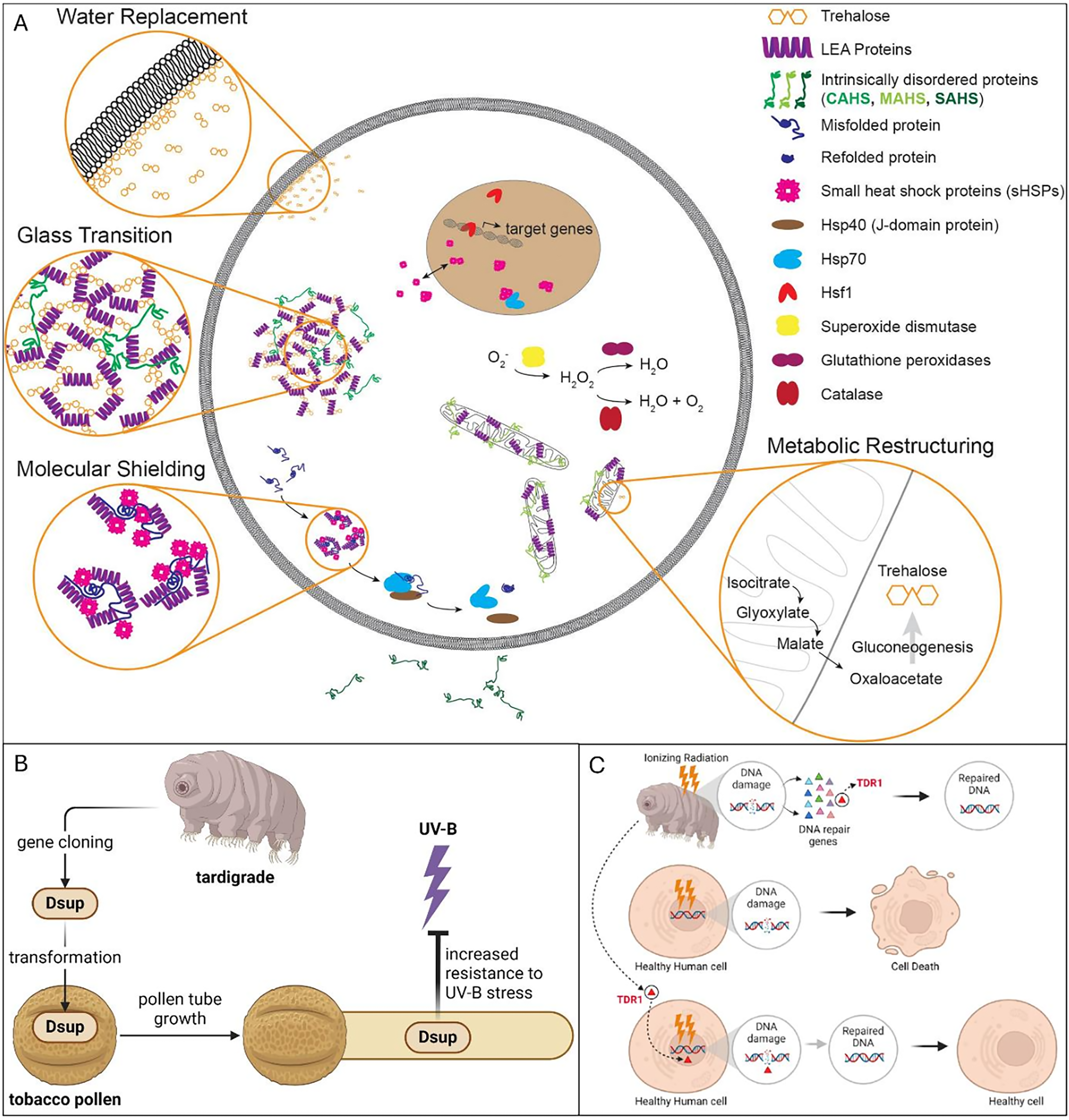
The dormancy mechanisms of tardigrades are supported by specific molecular and genetic factors that enable their remarkable survival strategies. Recent advances in ribosomal RNA sequencing and genomic analysis have provided more profound insights into the genetic basis of these mechanisms. These studies reveal that genes associated with stress responses are upregulated during dormancy, facilitating the transition into cryptobiotic states. 26 Two key proteins, Dsup and tardigrade DNA damage response 1 (TDR1), which are unique to tardigrades, play critical roles in protecting cells under extreme stress. 34 Dsup, on the other hand, binds to chromatin, shielding DNA from hydroxyl radicals and reactive oxygen species (ROS), thus reducing strand breaks. Additionally, its interaction with nucleosomal DNA facilitates the recruitment of DNA repair factors, thereby enhancing genomic stability.35,36 In contrast, TDR1 supports DNA repair by aiding the repair of double-stranded breaks 37 (Fig. 2a). In addition, tardigrade disordered proteins (TDPs), including CAHS, SAHS, and MAHS, are essential for maintaining cellular integrity during desiccation and for the formation of the tun state in anhydrobiosis. These proteins prevent protein aggregation and preserve cellular function even under conditions of extreme dehydration.14,30
Tardigrades unique physiological adaptations and molecular mechanisms not only underscore their evolutionary success but also offer exciting potential for advancements in biotechnology, space exploration, and medical research. Unraveling the mechanisms behind their dormancy and resistance to extreme environments may pave the way for innovative products and bioinspired applications.
Potential Applications of Tardigrades
Tardigrades' exceptional survival abilities, including cryptobiosis and protective proteins such as TDPs and Dsup, have sparked interest across multiple scientific fields. These adaptations are driving innovations in cryopreservation, radiation shielding, and space research.
TARDIGRADE AS A MODEL IN ASTROBIOLOGY
Scientists first considered tardigrades for space research in 1964 because of their extreme radiation resistance.21,40 Among species, Milnesium tardigradum is noted for its extremotolerance and was included in the 2007 Foton-M3 mission along with Paramacrobiotus richtersi and Richtersius coronifer. 21 This mission was the first to demonstrate that tardigrades can survive direct exposure to the Extravehicular Activity. Subsequent missions, such as the TARDIKISS (Tardigrades in Space) and Tardigrade Resistance to Space Effects (TARSE) projects, were launched to investigate tardigrade biology in space further in Low Earth Orbit (LEO), focusing on survival mechanisms, life cycle progression, enzymatic activity, and DNA integrity under both hydrated and desiccated conditions. 41 The findings indicated that microgravity and space radiation had minimal effects on the survival and fertility of R. varieornatus. The desiccated specimens of Richtersius coronifer and M. tardigradum were aboard the TARDIS mission, where they were first exposed to the combined effects of a space vacuum (10–6 Pa), cosmic radiation (100 mGy), and two different UV doses for 10 days. Tardigrades, shielded from solar radiation, were largely unaffected by cosmic radiation and vacuum, while those exposed to UV light showed high mortality, with none surviving the full UV spectrum. Remarkably, 12% of M. tardigradum endured combined exposure to vacuum, cosmic rays, and UVA+B, representing the first documented case of survival under true space conditions.
In contrast, space vacuum had no significant effect on egg-laying rates in either R. coronifer or M. tardigradum. Eggs produced by vacuum-exposed tardigrades hatched successfully at rates comparable to those of control groups in both species.42,43 However, UV radiation markedly reduced survival and egg production, though it did not affect egg hatchability or the fertility of subsequent generations. 44 This study provides the first evidence of an animal surviving both space vacuum and exposure to cosmic radiation, demonstrating that tardigrade cells, including developing eggs, can resist extreme desiccation at the expense of potential DNA damage. 42
On the other hand, the Rotifers, Tardigrades, and Radiation experiment examined how limno-terrestrial tardigrades respond to cosmic radiation, revealing a notable degree of resistance 45 Similarly, the TARDIKISS (Tardigrades in Space) project, conducted during the 2011 Endeavor mission, explored tardigrade DNA repair mechanisms under space-induced stress, further confirming their robustness in both hydrated and desiccated states. Horikawa et al. (2013) demonstrated that Ramazzottius varieornatus employs a dual survival strategy against UVC, involving both DNA protection and repair. In its desiccated anhydrobiotic state, it prevents the formation of UVC-induced thymine dimers, whereas in the hydrated state it efficiently repairs them via light-dependent mechanisms involving the photolyase gene phrA. Compared with hydrated specimens, anhydrobiotic tardigrades accumulate far fewer DNA lesions, resulting in much higher survival and reproductive ability. This explains their extraordinary UV radiation tolerance relative to less-resistant species such as Hypsibius dujardini, which lacks the phrA gene and adequate DNA repair capacity. 46 The unintentional release of tardigrades during the 2019 Beresheet lunar lander crash has raised concerns about potential biological contamination of the Moon. 47 Scientists acknowledged the possibility that some tardigrades may have survived the impact, inadvertently introducing Earth-based organisms to the lunar environment. This incident highlighted the lack of stringent planetary protection protocols for private space missions and sparked global discussions on the need for clearer regulations to prevent forward contamination. As space exploration becomes increasingly commercialized, the event underscored the importance of establishing international guidelines to safeguard celestial bodies from unintended biological transfer.
ENHANCING OIL RECOVERY
Conventional oil recovery methods, including primary recovery using natural reservoir pressure and secondary recovery through water or gas injection, typically recover only 35–65% of the original oil, leaving up to 55% of the oil trapped in the reservoir. 48 To address this limitation, enhanced oil recovery (EOR) techniques have been developed and are generally categorized into chemical, thermal, and gas injection methods. Chemical EOR utilizes polymers, surfactants, and alkaline agents to reduce interfacial tension and improve oil mobility; thermal EOR, including steam injection and in-situ combustion, lowers oil viscosity to enhance flow. Gas injection methods employ CO2, nitrogen, or natural gas to increase reservoir pressure and reduce oil viscosity. Despite their effectiveness, these methods face challenges such as high operational costs, reservoir heterogeneity, and the need for secure CO2 sources. Recent innovations, such as smart polymers, nanotechnology, and microbial EOR, aim to overcome these challenges and improve recovery efficiency while reducing environmental impact. 49 In this context, a study by Nmegbu et al. explored the potential of tardigrades, in conjunction with nanoparticles such as aluminum oxide and iron oxide, for EOR. 50 Five test formulations using crude oil from the Otakikpo marginal oil field were evaluated. These formulations include crude oil alone (Sample A), crude oil with tardigrades (Sample B), crude oil with aluminum oxide (Sample C), crude oil with iron oxide (Sample D), and a combination of tardigrades and both nanoparticles (Sample E). Sample B (tardigrades) yielded 15 L of oil, outperforming Sample A (7.8 L), Sample C (9 L), and Sample D (11 L). Sample E demonstrated the highest recovery, yielding 19 L within 10 minutes, indicating a synergistic effect between tardigrades and nanoparticles. These results suggest that tardigrades can modify crude oil viscosity and density, enhancing flow and recovery efficiency, and their resistance to extreme conditions positions them as promising, cost-effective candidates for application in challenging EOR environments.
Although current studies remain limited and largely conceptual, tardigrades' unique biological properties may have a role in bio-assisted EOR, suggesting potential for ex situ applications. For instance, tardigrade-derived enzymes or biomolecules could be explored for their stability and functionality during storage or processing stages outside the reservoir environment. Further investigation is needed to elucidate the molecular mechanisms by which tardigrade systems might interact with hydrocarbons. As no direct in-reservoir mechanistic data currently exist, this represents a novel conceptual frontier rather than an established oil recovery technology.
TARDIGRADE AS BIO-INDICATORS
Environmental pollution poses a serious threat to ecosystem health, negatively impacting biodiversity, soil quality, and water resources. Industrialization, agricultural runoff, and urbanization have led to elevated concentrations of heavy metals, pesticides, and other chemical pollutants, resulting in ecosystem degradation and loss of species diversity. Practical assessment of these changes requires bioindicator organisms whose presence, absence, or physiological condition reflects environmental quality. 51 Tardigrades, despite their reputation for resistance, exhibit notable sensitivity to specific pollutants while showing exceptional tolerance to environmental stressors. A long-term warming experiment demonstrated that elevated temperatures did not significantly affect their abundance, richness, or diversity, unlike many other soil invertebrates. Their thermotolerance, cryptobiosis, and the insulating properties of soil litter likely contribute to this stability. 52 Notably, many tardigrades have been identified as sensitive to heavy metals (e.g., copper and zinc) and to nutrient pollution (e.g., nitrates and phosphates), making them suitable for monitoring industrial and agricultural contaminants.53–56 In a study conducted at a wastewater treatment facility in Poznań, Poland, Thulinius ruffoi showed high abundance (up to 8300 individuals/L) in the first of four nutrient-rich, well-oxygenated activated sludge, where chemical oxygen demand (COD) ranged from approximately 783 to 1760 mg/L and biochemical oxygen demand (BOD) ranged from about 370 to 850 mg/L, with suspended solids at their highest levels. Abundance decreased to around 423 individuals/L in the fourth tank, coinciding with significant reductions in nutrient levels: chemical COD dropped to 27–453 mg/L, and BOD decreased to 2–110 mg/L. Seasonal peaks occurred in summer and early autumn, indicating a preference for warmer conditions. Despite the presence of pollutants, including heavy metals, T. ruffoi demonstrated resilience, making it a promising bioindicator of changes in nutrient load in wastewater treatment systems. 56 Furthermore, 20 tardigrade species were identified from moss samples collected across Hungary, with species and specimen numbers varying widely. Using European relative limit values for heavy metals, samples with all metals below limits averaged five species and 142 specimens, while increasing the number of metals above limits progressively reduced biodiversity and abundance, down to 1 species and one specimen when seven metals exceeded limits, demonstrating clear pollution impacts on tardigrades. 57 However, the limitations of tardigrades as a bioindicator were examined through acute toxicity testing with ammonia. Where T. ruffoi exhibits acute toxicity to ammonia, with no surviving individuals after 24-hour exposure at ≥ 125 mg/L total ammonia nitrogen, the LC50 values are 52 mg/L for total ammonia nitrogen and 0.65 mg/L for unionized ammonia. Despite this sensitivity, T. ruffoi tolerates ammonia levels above typical regulatory limits for wastewater, limiting its suitability as an ammonia bioindicator. The study highlights the need for standardized ammonia testing methodologies and further research on the effects of ammonia on tardigrade life-history traits for environmental monitoring applications. 58
Compared with conventional bioindicators such as chironomid larvae and diatoms, tardigrades offer unique advantages due to their ability to survive in extreme or degraded habitats where other organisms cannot. 59 However, their small size and often specialized ecological niches may limit their applicability in broad-spectrum monitoring, positioning them instead as valuable tools for targeted assessment of specific environmental stressors.
IN RADIATION PROTECTION
Radiation exposure can cause severe DNA damage and oxidative stress, posing significant risks in contexts such as cancer radiotherapy, space exploration, and nuclear accidents. As discussed earlier, tardigrades have evolved unique molecular mechanisms that enable them to tolerate extreme stressors, including high doses of radiation. The resistance is attributed mainly to protective proteins such as Dsup and TDR1, which shield DNA from radiation-induced breaks and oxidative damage.14,60 Several studies on Hypsibius exemplaris reveal striking ionizing radiation resistance driven by rapid, robust DNA repair involving massive upregulation of key repair genes and the tardigrade-specific gene TDR1, which stabilizes DNA. These rapid and massive upregulation of DNA repair pathway genes, particularly BER and NHEJ, with at least one gene, XRCC5, are shown to be functionally required for radiation tolerance.34,61 A similar experiment comparing transcriptomic and proteomic analyses identified TDR1 in H. exemplaris as strongly induced by radiation, which enhances DNA damage resistance, including when expressed in human cells. 34 Multi-omics analysis of H. henanensis reveals three key radiotolerance mechanisms: a horizontally transferred DODA1 (DOPA dioxygenase 1) gene encoding antioxidant betalains, a tardigrade-specific disordered protein, TRID1, promoting DNA repair via phase separation, and mitochondrial proteins BCS1 (ubiquinol–cytochrome c reductase (bc1) synthesis)/NDUFB8 (NADH dehydrogenase (ubiquinone) 1 beta subcomplex subunit 8) enhancing NAD+ regeneration.
Together, these pathways support efficient free-radical scavenging, PARP1 (poly(adenosine diphosphate–ribose) polymerase 1)-mediated DNA repair, and cellular recovery under extreme radiation. 62
Similarly, Dsup has attracted attention for its potential biomedical applications. A recent study demonstrated that ionizable lipid nanoparticles (LNPs) encapsulating Dsup mRNA can protect oral and rectal tissues from radiation-induced DNA damage. Administered to healthy tissues surrounding tumors before radiation therapy, these LNPs effectively reduced DNA damage by enabling local cellular expression of antioxidant enzymes that scavenge ROS. These findings suggest that Dsup mRNA delivery via LNPs may serve as a novel therapeutic strategy to reduce the side effects in cancer patients undergoing radiation treatment. 63
Beyond cancer radiotherapy, the unique biomolecular adaptations of tardigrades are inspiring a range of biomedical innovations. The Dsup protein, initially identified in R. varieornatus, has been shown to reduce DNA strand breaks and cellular damage caused by ionizing radiation when expressed in human cells, yeast, and mammalian models.64,65 This has fueled interest in Dsup for applications in gene therapy, radioprotective drug development, and tissue engineering. For instance, Dsup-expressing stem cells could be employed to enhance the survival of grafted tissues in high-radiation environments, such as during cancer treatments or organ transplantation procedures that involve preconditioning radiation. 66 In addition, synthetic biology platforms are exploring the integration of Dsup into cellular systems for long-term storage, preservation, and biobanking, where radiation and oxidative damage are significant concerns.67,68 On the other hand, the protective effects of the tardigrade Dsup protein on tobacco pollen under UV-B stress were studied. Transgenic pollen expressing Dsup showed enhanced antioxidant levels, improved pollen tube growth, restored callose plug formation, and increased stress-related proteins (tubulin, HSP70). These findings suggest that Dsup expression can reduce UV-B-induced damage (Fig. 2b). 38
Other tardigrade-derived molecules, such as TDPs and late embryogenesis abundant (LEA) proteins, show promise for biostabilizing pharmaceuticals, such as vaccines and biologics, under extreme conditions. TDPs, including CAHS, SAHS, and MAHS families, are intrinsically disordered and can form non-crystalline, glass-like matrices that preserve cellular and molecular integrity during desiccation, potentially enabling room-temperature stable formulations that bypass the cold chain. 69 Experimental studies have characterized CAHS proteins that gel upon drying, forming strong, reversible matrices capable of protecting client proteins. 70 In addition to their role in biostabilization, recent reviews highlight the therapeutic potential of tardigrade-derived proteins such as Dsup, CAHS, and LEA in attenuating oxidative stress, reducing DNA damage, and enhancing radiotolerance in transfected human cells and various model organisms. The radioprotective role of Dsup has been validated in several model organisms, including transgenic Caenorhabditis elegans, Drosophila melanogaster, Escherichia coli, and mammalian cells. In these systems, Dsup has been shown to reduce DNA strand breaks, oxidative stress, and radiation-induced mortality, while also functioning as a non-specific transcriptional repressor that modulates gene expression.60,65,71,72 Notably, human cells engineered to express Dsup exhibit 40–50% less DNA damage following X-ray exposure, underscoring their potential for astronaut protection against space radiation. Their successful expression in transgenic plants, animals, and human cells has consistently demonstrated improved survival rates and reduced DNA damage under radiation exposure, highlighting their potential in fields ranging from space medicine to radiotherapy support and synthetic biology.
APPLICATIONS IN THE FOOD INDUSTRY
Today, trehalose has attracted interest in many food industrial processes due to its protective role in molecular stabilization. 73 Research on trehalose in tardigrades dates back to 1974, when studies first demonstrated that tardigrades and nematodes undergoing anhydrobiosis synthesize trehalose to survive extreme desiccation. 74 These findings highlighted trehalose' critical role in protecting cellular structures during dehydration, laying the groundwork for understanding its broader function in stress tolerance and biological preservation. Trehalose acts as a protective sugar by forming a glassy matrix around proteins and membranes, stabilizing them and preventing denaturation and aggregation during freezing, drying, and other extreme conditions (Fig. 2c). This unique property has led to its application as a natural preservative in food products, where it helps retain texture, flavor, and aroma, while reducing moisture loss and preventing ice crystal formation during freezing. Trehalose is generally recognized as a safe compound widely used for protein and membrane stabilization, and hence successfully employed in preserving liposomes, which are used to encapsulate and deliver drugs, nutrients, and bioactive compounds, demonstrating its potential for stabilizing sensitive food and pharmaceutical ingredients.73,75
TDPs offer promising potential in enhancing the stability of probiotics, bioactive compounds, and other sensitive food ingredients by protecting them from degradation during processing and storage. TDPs are experimental, species-specific biomolecules that physically bind to and protect DNA, providing unique mechanisms of radiation resistance currently under investigation. These proteins, particularly CAHS, form noncrystalline, amorphous solids upon drying, creating a protective vitrified matrix around biological molecules. This matrix prevents protein aggregation and denaturation during dehydration and supports proper protein refolding upon rehydration.14,76–78 By enhancing resistance to drying and thermal stress, TDPs help maintain the functional efficacy of sensitive components and extend shelf life. Specifically, CAHS contribute to desiccation tolerance by interacting with residual water molecules, thus stabilizing biomolecules during drying. In the food industry, CAHS-based technologies have the potential to both improve crop drought resistance and extend the shelf life of dry foods minimizing dehydration-related damage. 79 While research on trehalose and its role in liposome and protein preservation has laid an important foundation, tardigrade-specific proteins such as CAHS and Dsup are still in the early stages of application in food systems. Recent developments include a patent application proposing the use of CAHS proteins to stabilize probiotic formulations, though clinical and industrial validation remains forthcoming. 77
IN BIOMEDICAL ADVANCES
Tardigrades exceptional resistance has inspired significant medical advancements, particularly in drug stabilization, cryopreservation, and organ preservation. Their TDPs, CAHS proteins, and trehalose are being explored to improve the shelf life of pharmaceuticals, enhance the viability of cryopreserved tissues and cells, and extend the preservation window for transplant organs under suboptimal conditions.
In biopharmaceuticals
The cold chain requirement remains a critical barrier in the global distribution of vaccines and biologics. Most biologics, including monoclonal antibodies, recombinant proteins, and traditional vaccines, must be stored and transported at 2–8°C to maintain their structural integrity and therapeutic efficacy. Even brief temperature excursions can result in irreversible protein denaturation, loss of antigenicity, and reduced potency, rendering the product ineffective. This stringent requirement poses serious logistical challenges, particularly in low-resource settings where electricity supply is unreliable, refrigeration infrastructure is limited, and transportation networks are underdeveloped. In such regions, maintaining the cold chain is not only technically challenging but also economically burdensome, with estimates suggesting that cold chain logistics can account for up to 80% of the total cost of vaccination programs in some developing countries.80,81
Trehalose plays a vital role in the development of thermostable vaccine formulations by protecting biological molecules from denaturation during desiccation and thermal stress. Recent studies have shown that trehalose can effectively stabilize vesicular stomatitis virus vectors during spray drying, maintaining their immunogenicity even after extended storage at 37°C, whereas liquid controls lost their immune-stimulating capabilities.82,83 In addition to trehalose, CAHS have demonstrated strong potential to enhance the stability of vaccines and biologics. CAHS proteins form protective glass-like matrices during drying, enabling room-temperature storage without compromising bioactivity.14,77,78 Besides vaccines, TDPs have been shown to preserve bacteria, yeast, and other biologics, suggesting future applications in temperature-stable drug manufacturing and even in cosmeceuticals, such as anti-ageing formulations. 14
Tardigrade-inspired stabilization mechanisms have shown promising potential in preserving biologics beyond vaccines. For example, Packebush et al. (2023) demonstrated that human blood clotting factor VIII could be successfully stabilized during drying and rehydration through cytoplasmic abundant heat soluble protein(CAHS D), thereby demonstrating the desiccation tolerance of Hypsibius exemplaris. 84 Hibshman et al. used an expression-cloning approach in E.coli as a model cargo to identify tardigrade proteins that enhance desiccation survival. By expressing cDNA libraries from two tardigrade species and subjecting bacteria to desiccation-rehydration cycles, they selected survivors to reveal potent protectants. Among the strongest identified were mitochondrial single-stranded DNA-binding proteins, whose DNA-binding activity was necessary and sufficient for protection, thereby improving bacterial survival by over 100-fold. 85 These findings highlight the broader biomedical relevance of tardigrade-derived molecules such as trehalose and TDPs, which are known to prevent protein denaturation and aggregation during dehydration and promote correct protein refolding upon rehydration. This has significant implications for the development of thermostable vaccines and biologics that retain efficacy without refrigeration, alleviating cold-chain challenges in global healthcare delivery. 84 Notably, no licensed biological currently uses tardigrade proteins for storage stabilization, indicating this is a novel area for future biotechnological development.
Platelet preservation
Platelets stored under conventional conditions undergo a range of damaging changes known as the platelet storage lesion, including morphological degradation, impaired adhesion and aggregation capabilities, and altered membrane markers. 86 Oxidative stress is a primary contributor, with platelet functionality declining significantly after 6 days and becoming severely compromised by day 8. Cryopreservation using 5–6% dimethyl sulfoxide (DMSO) at −80°C can extend storage for nearly 2 years but introduces complications such as altered platelet phenotype, heightened procoagulant activity, reduced aggregation response, and adverse patient reactions such as seizures, cardiac hypertension, and hypersensitivity. 87 Trehalose offers a promising alternative by inhibiting phagocytosis and preventing apoptosis, thus improving refrigerated platelet stability.17,88 Studies comparing in vivo and in vitro trehalose have demonstrated that trehalose-treated platelets significantly accelerate wound healing, with one study showing that trehalose-treated platelets closed wounds 6 days faster than untreated controls. Lyophilized platelet preparations with trehalose also enhanced wound closure, granulation tissue formation, neovascularization, and cell proliferation in diabetic mice, supporting their potential for managing chronic and nonhealing wounds. 89
MAHS proteins derived from tardigrades have shown promising potential for cellular preservation. Recent studies have demonstrated that MAHS expression in human adipose-derived stem cells significantly enhances cell viability under stress. Specifically, MAHS increased cell survival by up to 61% during a 72-hour incubation under nutrient-deprived conditions and improved survival by 14% following DMSO exposure for the same duration. 90 These findings underscore the utility of MAHS in improving cell resilience under preservation-related stressors. In the context of platelet storage, tardigrade-intrinsic proteins like MAHS could help maintain membrane integrity, reduce oxidative stress, and preserve platelet functionality during extended storage. However, these claims emphasize conceptually promising aspects and the requirement for controlled experimental validation to assess aggregation, clot retraction, viability, and sterility. By mimicking tardigrade’s ability to tolerate desiccation and freezing, these proteins may lead to advanced cryopreservation techniques with reduced dependence on toxic cryoprotectants such as DMSO.
In organ transplantation
Current organ preservation techniques are constrained by limited storage durations, particularly for organs such as the heart and lungs, which remain viable for only 4–6 hours post extraction. Even kidneys, which can be stored for up to 24 hours, often suffer from ischemia-reperfusion injury (IRI), a major contributor to poor transplant outcomes due to oxidative stress and inflammation following reoxygenation.91,92 Tardigrade-derived biomolecules have shown promising proof-of-concept potential in specific organ models. The Dsup protein, known for reducing DNA damage by up to 40% in human embryonic kidney cells exposed to X-rays, may help mitigate IRI-related oxidative stress.14,43 This protective effect suggests that Dsup could mitigate oxidative stress-induced DNA damage during ischemia-reperfusion in organ transplantation. 43 Additionally, CAHS proteins stabilize enzymes under stress, with their protective effect linked to charge density, suggesting potential for bioengineering optimization.14,93 Trehalose, a sugar associated with tardigrade anhydrobiosis, has already inspired advances in organ preservation. For instance, the ET-Kyoto solution has shown superior results in lung transplantation compared to the Euro-Collins solution. 81 Trehalose has also demonstrated therapeutic potential in mammalian eye care 94 and extended rat heart preservation up to 96 hours when used with perfluorocarbon immersion and perfusion. 95
While these findings provide strong proof-of-concept for tardigrade biomolecules enhancing organ preservation in kidney, lung, and heart models, they primarily come from small-animal or cellular studies. Significant challenges remain, such as obtaining large-animal validation and ensuring compatibility with perfusion-based clinical preservation systems. Additionally, a comprehensive evaluation of cost-effectiveness and scalability is needed to facilitate translation into clinical practice. Furthermore, regulatory approval will require extensive safety and efficacy trials, adding considerable time and expense before these approaches can be widely implemented clinically.
Oncofertility
Mature oocytes pose unique cryopreservation challenges due to their large size, high intracellular water content, and low surface area-to-volume ratio, all of which increase the risk of intracellular ice crystal formation during freezing and thawing. The metaphase II (MII) spindle apparatus is particularly vulnerable to cryoinjury, often resulting in chromosomal misalignment and increased aneuploidy. Although vitrification techniques employing high concentrations of cryoprotective agents (CPAs) such as DMSO, ethylene glycol, and propanediol successfully prevent ice formation, they are associated with cytotoxicity, osmotic stress, and potential epigenetic modifications. Despite advancements that yield post-thaw survival rates of 80–90%, oocyte cryopreservation still suffers from reduced fertilization efficiency and impaired developmental competence. 96 Trehalose-based cryopreservation has become a foundational approach in oncofertility research, mostly tested in animal models such as mouse oocytes. Experimental studies using mouse MII oocytes have demonstrated that both intracellular and extracellular trehalose significantly improve cryosurvival, with extracellular concentrations up to 0.5 M maintaining high viability even after exposure to liquid nitrogen temperatures (−196°C). Notably, combining trehalose with reduced concentrations of permeating CPAs, such as 0.5 M DMSO, further improved fertilization and embryonic development, yielding outcomes comparable to those of fresh oocytes. These protocols have successfully resulted in the birth of healthy offspring, underscoring trehalose’s potential in human fertility preservation.81,97 In addition to trehalose, tardigrade-derived intrinsically disordered proteins (IDPs), such as CAHS, SAHS, and MAHS, may provide molecular protection for cryopreserved oocytes. However, these proteins are still in experimental and investigational stages. CAHS proteins are hypothesized to stabilize intracellular structures such as mitochondria, the endoplasmic reticulum, and cytoskeletal components during freezing, thereby preserving essential functions required for embryogenesis. SAHS proteins may reinforce extracellular matrices, including the zona pellucida, thereby reducing the risk of zona hardening, a phenomenon that impairs fertilization following cryopreservation. Furthermore, MAHS proteins could mitigate mitochondrial damage by preserving membrane integrity and reducing ROS generation, both of which are critical to maintaining oocyte quality post-thaw.14,69 These studies demonstrate trehalose’s protective effects by stabilizing cellular membranes and reducing cryoinjury during freeze-thaw cycles. In contrast, tardigrade protein-based approaches, including proteins such as Dsup and CAHS, remain investigational and are not yet established in clinical or preclinical oncofertility applications. Furthermore, it is important to emphasize that protein and gene delivery vectors based on tardigrade proteins (e.g., Dsup) require careful regulatory evaluation due to potential safety and ethical concerns before any clinical use can be considered.
Collectively, tardigrade-inspired molecules hold significant promise in advancing oocyte cryopreservation technologies. By enhancing cytoplasmic and mitochondrial stability, reducing oxidative stress, and preserving extracellular architecture, these biomolecules may substantially improve post-thaw survival, fertilization capacity, and developmental competence, offering new avenues for fertility preservation in clinical and research settings.
Challenges Associated with Developing Applications
Although tardigrades provide compelling models for extreme stress tolerance, translating their biology into practical applications presents several scientific challenges. Many protective mechanisms, such as CAHS/SAHS or Dsup, exhibit strong context dependence, with protective effects varying across organisms, cell types, expression levels, and stress modalities.2,35,77 Moreover, increasing evidence suggests that tardigrade resilience arises from the coordinated action of DNA repair pathways, chromatin remodeling, metabolic regulation, and stress-responsive transcriptional programs, rather than from single molecules acting in isolation.43,62 This systems-level complexity complicates attempts to engineer tardigrade-inspired solutions using simplified or reductionist approaches.
Significant challenges also exist at the translational and regulatory levels. Large-scale production of tardigrade proteins, particularly TDPs, poses difficulties related to stability, consistency, and functional validation across environments. For biomedical, agricultural, or industrial applications, additional hurdles include potential immunogenicity, delivery and dosing strategies, off-target effects, and compliance with regulatory frameworks governing genetically modified organisms or biologically derived therapeutics.14,80,83,86,91,93 As a result, most proposed applications of tardigrade biology remain at an early proof-of-concept stage. Addressing these challenges will require a deeper mechanistic understanding, standardized evaluation frameworks, and cautious assessment of feasibility to ensure realistic translation of biological insights into applied technology.
Conclusion
Tardigrades are micro-metazoans that exhibit extraordinary resistance through cryptobiosis and molecular adaptations. Molecules such as trehalose, Dsup, and TDPs are not only key to tardigrade survival but also hold transformative potential across diverse biotechnological domains. Trehalose enables cold-chain-independent stabilization of vaccines, platelets, and oocytes. Dsup provides radioprotection by shielding DNA, with implications in cancer therapy, organ transplantation, and space exploration. TDPs confer desiccation tolerance, enhancing applications in cryopreservation, food preservation, and beyond. CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) breakthroughs in R. varieornatus have advanced our understanding of these mechanisms, supporting their use in synthetic biology, bioindicator systems, and stress-resistant bioengineering. However, challenges such as production scalability, immune safety, and regulatory hurdles must be addressed. The key challenge is the scalable, cost-effective production of tardigrade-derived proteins. Delivery mechanisms must overcome immune system interactions, and extensive safety and efficacy testing is needed for regulatory approval. Environmental considerations, especially in large-scale applications, must meet the criteria of ecological safety, minimal biodiversity disruption, and lifecycle sustainability. This includes ensuring that tardigrade-derived molecules or engineered systems do not negatively impact local ecosystems, maintaining biosafety in microbial production platforms, and adhering to regulatory frameworks for environmental release. Sustainable design should emphasize biodegradability, low energy inputs, and compatibility with circular bioeconomy principles to align with global environmental goals.
Authors’ Contributions
N.P. and A.S.: Conceptualization, literature review, drafting, and revision of the article. B.M., A.C., G.M., M.G., M.A., M.C., S.K., I.S., A.S., and G.S.: Data collection, analysis, visualization, and formatting. A.K.: Critical review and editorial support. G.D.M. and A.K.S.: Supervision, technical support; administration, project completions, review and approval.
Footnotes
Acknowledgments
The authors are grateful to Amity University Uttar Pradesh, India, for research support.
Author Disclosure Statement
All authors declare that they have no financial or personal relationships that could have appeared to influence the work reported in this article. No interests to disclose.
Funding Information
No funding was received for this article.
Availability of Data and Material
The data and supporting information are available within the article.