
Abstract
NIMA-related kinase 4 (NEK4) is a serine/threonine kinase implicated in microtubule stabilization, cilia function, and DNA damage response (DDR), with emerging roles in cancer progression through context-dependent effects on proliferation, epithelial-to-mesenchymal transition (EMT), and metastasis. Despite its significance, site-specific phosphorylation dynamics of NEK4 remain underexplored. Here, we conducted a comprehensive computational phosphoproteomic analysis by curating Class-1 phosphosites from over 3800 public datasets, identifying NEK4 phosphosites, including four predominant sites (S563, S661, S461, S639) outside the kinase domain that exhibit high detection frequencies and differential regulation. Coregulation analysis revealed phosphosites in other proteins (PsOPs) that coordinate with these NEK4 sites, linking them to DDR pathways (e.g., via interactions with DNA-PK complex components), EMT signaling, microtubule organization, and mitochondrial function. Network mapping integrated predicted upstream kinases (e.g., CDK13, RPS6KA1/3), downstream substrates (e.g., MKI67, INCENP), and binary interactors (e.g., TMPO, RRP1B), highlighting NEK4’s integration into cancer-associated networks involving cell cycle regulation, apoptosis, and autophagy. Functional enrichment underscored NEK4’s potential in modulating genotoxic stress responses and tumorigenic reprogramming. These findings provide a phospho-centric framework for NEK4 signaling, positioning it as a therapeutic target in DDR-defective and EMT-driven cancers, and lay the groundwork for experimental validation of its site-specific roles.
Introduction
NEK4 is a serine/threonine kinase belonging to the NIMA-related kinase (NEK) family, which spans NEK1 to NEK11 within the larger protein kinase superfamily (Li et al., 2025). Originally identified by Cance and colleagues in human breast cells as serine/threonine-protein kinase 2 (STK2) (Cance et al., 1993), NEK4 shares its ortholog with NIMA (NEVER IN MITOSIS GENE A) from Aspergillus nidulans, the founding member of the family (Basei et al., 2015). Comprising 841 amino acids, it is encoded on chromosome 3p21.1, transcribed into a ∼4 kb mRNA (Basei et al., 2015), and has a molecular weight of 94,597 Da. The protein localizes primarily to the cilium and cytoplasm (Coene et al., 2011).
NEK4 exists in two isoforms: isoform 1 (NEK4.1) and isoform 2 (NEK4.2). These isoforms contribute to microtubule stabilization, ciliary function, and the DNA damage response (DDR) (Basei et al., 2015). NEK4.1, distinguished by an insertion in its regulatory domain, overlaps with NEK4.2 in many interactions and processes but also exhibits unique roles, such as DNA repair, cilia regulation, translation control, and cell adhesion, that are novel within the NEK family (Basei et al., 2015). In contrast, NEK4.2 is primarily involved in mRNA processing, apoptosis, and transcription (Basei et al., 2015). Early studies on the murine homolog, initially termed STK2 (Chen et al., 1999; Hayashi et al., 1999), led to its reclassification as NEK4.
Like other NEK kinases, NEK4’s N-terminal catalytic domain is enriched in serine/threonine residues. All 11 human NEK proteins share a conserved His–Arg–Asp motif in this domain, characteristic of kinases activated by phosphorylation, including a regulatory serine or threonine in the activation loop (Fry et al., 2012). As one of the largest human NEKs, NEK4 includes an N-terminal catalytic domain and a C-terminal regulatory domain that regulates microtubules, primary cilia assembly, and DDR (Basei et al., 2015). Similar to NEK1 and NEK8, NEK4 supports primary cilia stabilization (Meirelles et al., 2014). In Arabidopsis thaliana, NEK5 interacts with NEK4 and NEK6 to regulate epidermal cell expansion and microtubule organization, potentially via β-tubulin phosphorylation (Motose et al., 2011). Additionally, NEK1, NEK4, NEK6, NEK8, NEK10, and NEK11 coordinate DDR and the G2/M cell cycle checkpoint (Meirelles et al., 2014).
Research by Nguyen and colleagues has shown that NEK4 interacts with the DNA-PK complex (including DNA-PKcs, Ku70, and Ku80), facilitating DNA-PKcs recruitment to DNA damage sites. This promotes efficient nonhomologous end-joining (NHEJ) repair, H2AX activation, and p53 signaling in response to double-strand breaks (DSBs; Nguyen et al., 2012; Pavan et al., 2021). Without NEK4, the Ku70/Ku80 complex dissociates prematurely from DSB sites, impairing DNA-PKcs binding, reducing H2AX phosphorylation, and weakening DNA repair and cell cycle arrest (Nguyen et al., 2012; Pavan et al., 2021).
Beyond DDR, NEK4 influences mitochondrial function: its overexpression boosts respiration, elevates mitochondrial membrane potential, and protects against mtDNA damage, while depletion impairs respiration, reduces mtDNA integrity, and causes mitochondrial elongation, possibly by modulating DRP1 (Basei et al., 2022). NEK4 inhibition also sensitizes cancer cells to TRAIL-induced apoptosis by downregulating survivin (Park et al., 2016). In cancer, NEK4’s role is context-dependent; it suppresses proliferation in triple-negative breast cancer but promotes epithelial-to-mesenchymal transition (EMT), invasion, and metastasis in lung cancer. For instance, suppressing NEK4 in A549 lung adenocarcinoma cells reduces migration and invasion (Ding et al., 2018).
Phosphoproteomic studies have advanced our understanding of kinase signaling and therapeutic targeting, yet phosphosite-specific data on NEK4 across biological and pathological contexts remain limited. To bridge this gap, we employed a computational phosphoproteomic approach to analyze NEK4 phosphorylation sites from public datasets, akin to large-scale kinase profiling. This revealed co-regulation patterns between NEK4 and its interacting protein-phosphosites in various processes. Notably, NEK4 phosphorylation potentially links key pathways such as DDR, EMT, and microtubule organization. Integrative network mapping highlights its role in DSB repair. With growing evidence tying NEK4 overexpression to EMT and cancer progression, deeper phosphoproteomic analysis could uncover its molecular contributions to tumorigenesis and validate it as a promising therapeutic target.
Materials and Methods
Assembly of global phosphoproteomic datasets of NEK4
The assembly and analysis of the global phosphoproteomic datasets of NEK4 employed the previously established methodology developed in our laboratory (George et al., 2026; Kumar et al., 2026; Sheela et al., 2025). Briefly, to assemble the phosphosites of NEK4, we screened the human cellular global phosphoproteome datasets available in PubMed using the search term “phosphoproteomics” OR “phosphoproteome” NOT “Plant” NOT “Review” to identify those containing Class-1 NEK4 phosphosites (localization probability ≥75%; A-score >13). The datasets were categorized into qualitative profile datasets, which analyzed test and control samples separately, and quantitative differential datasets, which compared test samples under specified biological or experimental settings to their matched controls. To further refine our findings, datasets were classified based on the type of phosphosite enrichment, either serine/threonine/tyrosine (S/T/Y) residues. In differentially expressed datasets, phosphosites with a fold change of ≥1.3 were considered as upregulated, and those with a fold change of ≤ 0.76 were considered as downregulated. To ensure significant and statistically reliable results across various biological conditions, we applied a significance threshold of p < 0.05. To avoid discrepancies in protein names, we mapped individual proteins to their respective HUGO Gene Nomenclature Committee (HGNC) gene symbols (Seal et al., 2023). Using our in-house mapping tool, phosphosites in each protein were mapped to corresponding UniProt accession numbers (downloaded May 13, 2023) (UniProt Consortium, 2023).
Determining the predominant phosphosites in NEK4
To determine the predominant sites in NEK4, we evaluated qualitative datasets collected from human cells and enriched for S/T/Y phosphoproteomes. Phosphosites were ranked based on their frequency of detection in these datasets. The most frequently observed phosphosites in NEK4 were then selected based on quantitative data to explore the coregulated phosphosites in other proteins (PsOPs).
Identification and analysis of coregulated PsOPs with NEK4 predominant phosphosites
To assess the role of NEK4 phosphosites, we extracted PsOPs that were coregulated across several quantitative datasets, using the method described earlier (George et al., 2026; Kumar et al., 2026; Sheela et al., 2025). Because of the significant variety in experimental designs and biological systems, re-analyzing the raw data was impractical. To ensure data reliability, we only included the phosphosites with the localization probability ≥75% or A-score >13, resulting in significant site-specific identification. PsOPs were grouped based on their expression changes relative to the NEK4 phosphosite to interpret coregulation patterns. The pair was labeled as UU when PsOPs and the NEK4 site were upregulated. If the PsOPs were downregulated while NEK4 was upregulated, it was assigned to the UD group. Likewise, when NEK4 phosphosites were downregulated along with co-upregulated PsOPs, they were categorized as DU, and co-downregulated PsOPs as DD. To enhance clarity and reduce variability, we further organized the PsOPs into two broad categories: Positively coregulated phosphosites (UUDD) indicated PsOPs that are either upregulated or downregulated simultaneously with NEK4 phosphosites, while negatively coregulated phosphosites (UDDU) demonstrated PsOPs that are upregulated when NEK4 phosphosites are downregulated, or vice versa.
To assess coregulatory patterns between NEK4-predominant phosphosites and their associated PsOPs across the quantitative profile datasets, we ran a one-sided Fisher’s exact test (FET) using a contingency table (George et al., 2026; Kumar et al., 2026; Priyanka et al., 2024; Sheela et al., 2025). PsOPs with a p-value of <0.05 in UUDD or UDDU categories were considered statistically significant and included for further studies. This approach is similar to a previously described method for detecting co-occurring phosphosites in qualitative data from PhosphoSitePlus (Hornbeck et al., 2012). Differential FET scores for each NEK4 phosphosite PsOP pair were calculated using a total number of differential datasets. The high-confidence identification cutoff was 10% of the total differential datasets relating to a single NEK4 phosphosite. To avoid potential bias from overrepresentation of multitemporal datasets and datasets derived from similar experimental stimuli, we applied a cutoff of PMID (Pubmed ID) confidence (≥3) and an experimental code confidence (experiments carried out with the same stimuli, ≥3). The PsOPs that obey these four criteria were considered as the high-confidence PsOPs and were used to analyze the biological processes, binary interactors, upstream kinases, and downstream substrates of NEK4. In addition, the other kinases associated with predominant sites of NEK4 are assembled from KinHub (Manning et al., 2002).
Co-occurrence analysis of NEK4 phosphosites
A co-occurrence analysis was performed to examine the relationships between phosphorylation sites in NEK4, with a special focus on the codifferential regulation patterns of phosphosite pairs within NEK4. Differential datasets containing multiple NEK4 phosphosites observed under the same experimental conditions were selected. For each phosphosite pair, we determined the frequencies of all four regulation combinations: UU, UD, DD, and DU. Positive coregulation was calculated using the ratio ∑(nUU + nDD)/∑(nUD + nDU), while negative co-regulation was calculated using ∑(nUD + nDU)/∑(nUU + nDD).
Identification of potential phosphocentric regulatory network of predominant NEK4 phosphosites
To explore the regulatory network associated with NEK4 phosphosites, we investigated both experimentally validated and computationally predicted upstream kinases and downstream substrates. Proteins showing either positive or negative coregulation with specific NEK4 phosphosites were selected for further analysis. Information on known kinases and substrates of NEK4 was assembled from curated phosphoproteomic databases, including PhosphoSitePlus (downloaded May 2023) (Hornbeck et al., 2012), Phospho.ELM (downloaded May 2023) (Dinkel et al., 2011), and RegPhos 2.0 (downloaded May 2023) (Huang et al., 2014). To broaden this dataset, we incorporated predictions from tools such as NetworKIN (Linding et al., 2008), Automatic Kinase-specific Interactions Detection (AKID) (downloaded June 2023) (Johnson et al., 2019), and the in vitro Kinase-to-Phosphosite Database (Mari et al., 2022). Additionally, all of the substrates and kinases classified by Johnson et al. (2023) based on synthetic peptide screening to evaluate substrate specificity within the kinome and using a 90-percentile cut-off were extracted for further analysis (Gopalakrishnan et al., 2025).
Identification of protein and phosphosite-specific binary interactors of NEK4
To construct a comprehensive protein interaction network of the binary interactors, we integrated experimentally validated protein–protein interactions, including kinase and substrate data, from major databases such as the Human Protein Reference Database (Goel et al., 2012), Biomolecular Interaction Network Database (Bader et al., 2003), BioGRID (Oughtred et al., 2021), ConsensusPathDB (version 35) (downloaded May 2023), CORUM (downloaded March 2023) (Steinkamp et al., 2025), and RegPhos 2.0 (downloaded May 2023; Huang et al., 2014).
Data visualization-related tools
For data visualization, we employed the R/Bioconductor package trackViewer (10.18129/B9.bioc.trackViewer) to generate lollipop. https: Cytoscape (version 3.10) (Shannon et al., 2003) was used to map the relationship of PsOPs within the substrate, interactor, and kinase networks. RAWGraphs 2.0 (10.1145/3125571.3125585) was applied for clustering and hierarchical visualization, while Matplotlib and Pandas were employed for quantitative analysis of phosphosite distribution patterns.
Results
Class I phosphosites and predominant phosphosites of NEK4 in human global phosphoproteome datasets
By systematically screening, integrating and curating phosphoproteomics data from >3800 PubMed-indexed studies, we compiled cellular phosphoproteomics datasets covering a broad spectrum of experimental conditions. Out of the compiled datasets, 492 were classified as qualitative profiles and 95 as quantitative differential datasets, each providing data on Class-1 NEK4 phosphosites. Across these cellular datasets, a total of 31 NEK4 phosphosites were detected, of which 12 showed quantitative measurements indicating differential expression compared to control samples. The complete list of phosphosites from both the differential and profile datasets is provided in Supplementary Tables S1 and S2. The datasets revealed considerable variation in how often individual NEK4 phosphosites were detected, with some sites occurring much more frequently than others. Such variation likely arises from several factors, including reduced protease cleavage near phosphorylated residues (Solari et al., 2015), differences in peptide ionization and fragmentation efficiencies that favor peptides with optimal length and charge (Yu and Veenstra, 2021), and rapid dephosphorylation by phosphatases, which can reduce site stability.
To determine the predominant phosphosites in NEK4, all the Class-1 phosphosites detected in NEK4 were ranked based on the frequency of detection in both profiling and differential datasets. Among the 12 phosphosites that showed differential expression, four sites, including S563, S661, S461, and S639, were detected at higher frequencies, appearing in 356, 229, 141, and 164 profiling datasets, and showed differential regulation in 42, 26, 24, and 21 datasets, respectively. As mass spectrometry relies on quantifying tryptic peptides, we further checked whether these phosphosites were present within the same peptide sequence to ensure that the interpretation was not redundant. Based on this analysis, all four predominant sites are on distinct peptides and are thus designated as predominant phosphosites of NEK4, reflecting consistent detection across both profiling and differential datasets. The phosphosite distribution across the profiling and the differential datasets is shown in Figure 1. Notably, all four predominant phosphosites of NEK4 are located outside the kinase domain, classifying NEK4 as a phosphosite outside kinase domain (PoKD) kinase, consistent with the report by Gopalakrishnan et al. (2025).
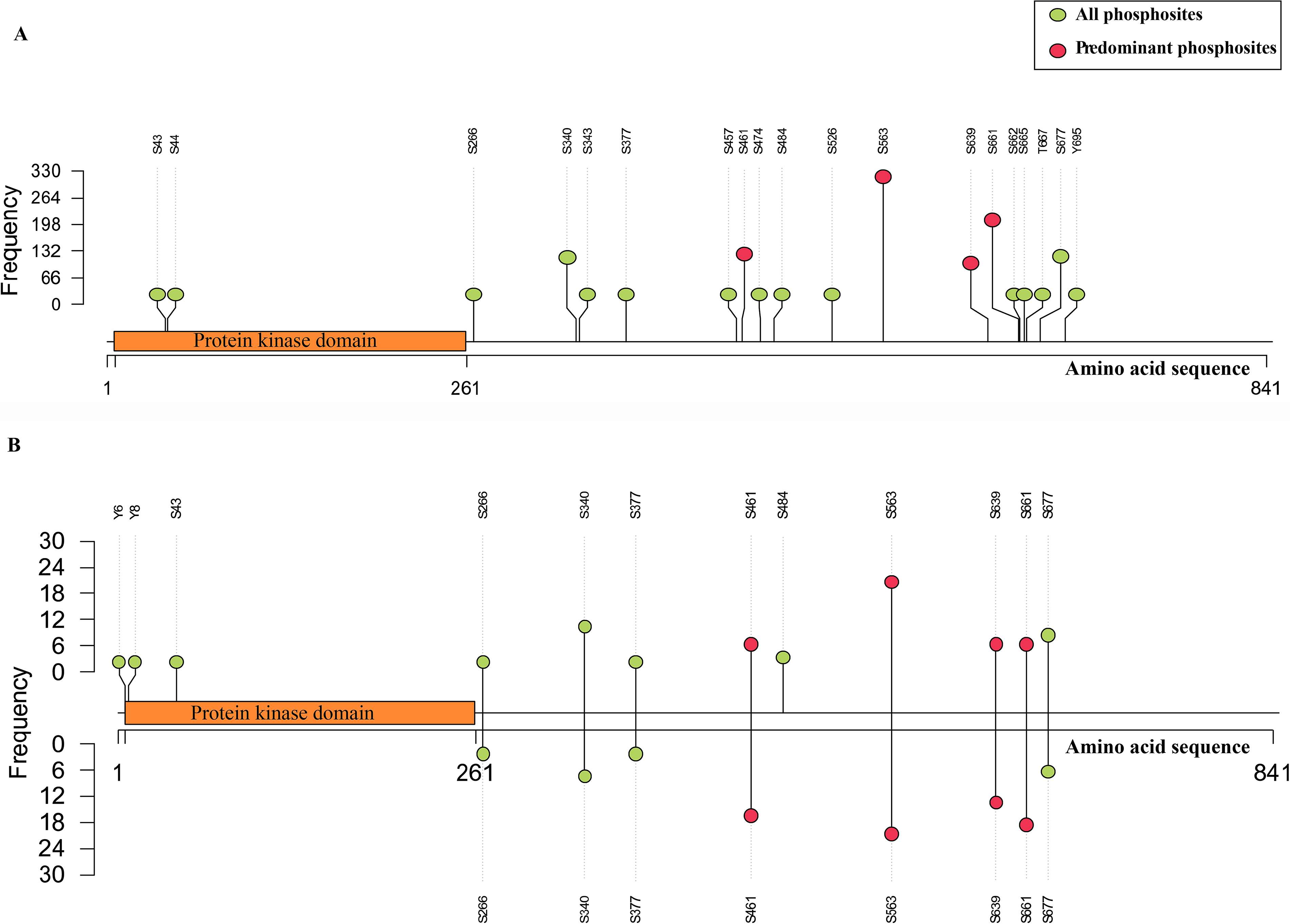
Lollipop plot illustrating frequency-based NEK4 phosphosites. The length of the amino acid sequence is represented by the X-axis, while the dataset frequency is represented by the Y-axis.
Differentially coregulated protein PsOPs with predominant sites of NEK4
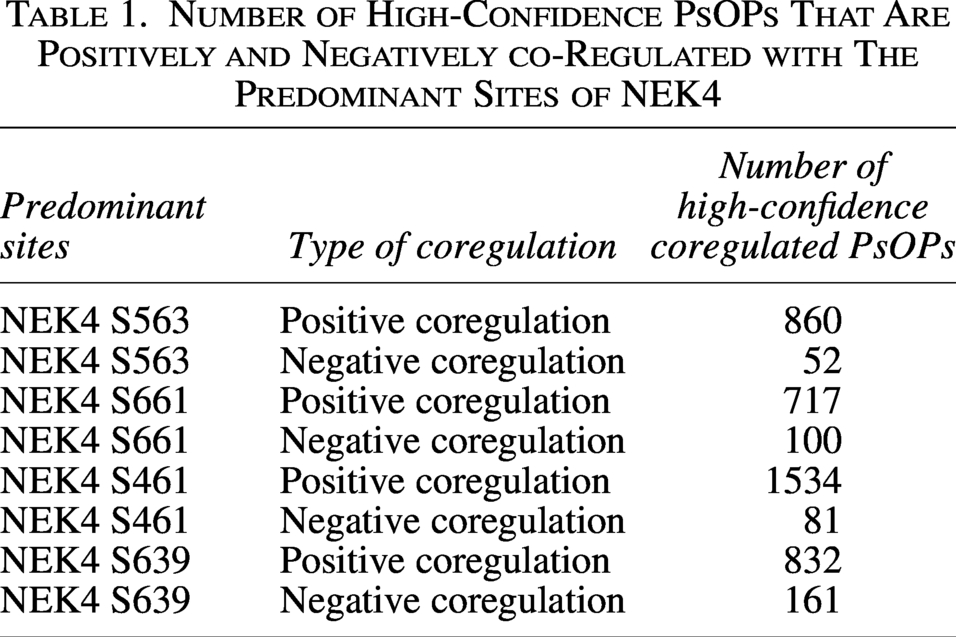
To explore the cellular phospho-signaling networks associated with the predominant sites of NEK4, we conducted a co-regulation analysis. This analysis mapped alterations in phosphosites of other proteins (PsOPs) that showed coordinated regulation with the primary NEK4 phosphosites across multiple phosphoproteomics datasets. The observed coregulation patterns showed substantial variability in both their frequency and direction of association. To ensure reliable results and limit bias from individual datasets, stringent selection criteria were implemented, as outlined in the Materials and Methods section—Determining the predominant phosphosites in NEK4. The number of high-confidence PsOPs showing positive and negative coregulation with the NEK4 predominant sites is summarized in Table 1 (Supplementary Tables S3, S4, S5, S6, S7, S8, S9, and S10).
Number of High-Confidence PsOPs That Are Positively and Negatively co-Regulated with The Predominant Sites of NEK4
Some of the positively co-regulated PsOPs with NEK4 (S563) were Eukaryotic Translation Initiation Factor 4 Gamma 2 (EIF4G2) (T508) and Establishment Of Sister Chromatid Cohesion N-Acetyltransferase 2 (ESCO2) (S75), with positive co-regulation frequencies of 26 and 24, respectively, and the top negatively co-regulated PsOPs with NEK4 (S563) were Mitogen-activated protein kinase 3 (MAPK3) (T202), Zinc transporter 6 (SLC30A6) (S382) and BSD domain-containing protein 1 (BSDC1) (S387), with negative co-regulation frequencies of 17 and 16 for both, respectively. Similarly, the top positively co-regulated PsOPs with NEK4 (S661) were ATP-citrate synthase (ACLY) (S455) and Zinc finger CCCH domain-containing protein 4 (ZC3H4) (S1275), with positive co-regulation frequencies of 18 for both, respectively, and the top negatively co-regulated PsOPs with NEK4 (S661) were Spastin (SPAST) (T303) and Zinc finger C3HC-type protein 1 (ZC3HC1) (S335), with negative co-regulation frequencies of 11 for both, respectively. Furthermore, the top positively co-regulated PsOPs with NEK4 (S461) were Ribosomal RNA processing protein 1 homolog B (RRP1B) (S706) and Telomere-associated protein RIF1 (RIF1) (S2196), with positive co-regulation frequencies of 18 for both, respectively, and the top negatively co-regulated PsOPs with NEK4 (S461) were B-cell lymphoma 3 protein (BCL3) (S374) and Eukaryotic translation initiation factor 4B (EIF4B) (S406), with negative co-regulation frequencies of 11 for both, respectively. Additionally, the top positively co-regulated PsOPs with NEK4 (S639) were CTD small phosphatase-like protein 2 (CTDSPL2) (S28) and Serine/threonine-protein phosphatase 2 A 56 kDa regulatory subunit delta isoform (PPP2R5D) (S573), with positive co-regulation frequencies of 17 and 15, respectively and the top negatively co-regulated PsOPs with NEK4 (S639) were Serine/threonine-protein kinase A-Raf (ARAF) (S257) and Proliferation marker protein Ki-67 (MKI67) (S827), with negative co-regulation frequencies of 11 and 10. The top 25 positively and negatively co-regulated PsOPs with predominant sites of NEK4 are shown in Supplementary Figure S1. However, for most of these co-regulated phosphosites, their functional significance at the phosphosite level remains unclear. Further, our analysis identified one PsOP that showed positive co-regulation with the predominant sites of NEK4. Notably, BAG family molecular chaperone regulator 3 (BAG3) (T406) showed positive co-regulation with all of these sites. These co-regulation pattern suggests the common functional significance of NEK4 predominant sites.
Co-occurrence of phosphorylation sites in NEK4
According to Li et al. (2017), co-occurring phosphorylation events within the proteins tend to exhibit functional interdependence (Li et al., 2017). To determine interdependent, mutually exclusive or coordinated signaling relationships between NEK4 phosphosites, we examined their co-occurrence across cellular differential phosphoproteomics datasets. We categorised those that were co-upregulated or co-downregulated as positive co-occurrence, whereas those with opposite expression patterns (one up-regulated and the other down-regulated) were classified as negative co-occurrence. Additionally, the positive and negative co-occurrence scores of the phosphosite pairs were calculated across the differential datasets to assess concordance, as shown in Supplementary Figure S2. Our analysis showed that the phosphosite pair S461-S563 exhibited positive co-occurrence in 10 datasets and negative co-occurrence in two datasets. A similar trend was observed in the S340-S563 pair, which showed positive co-occurrence in seven datasets and negative co-occurrence in one dataset. These positive co-occurrence patterns indicate potential functional relationships between the phosphosite pairs, suggesting coordinated signaling at these sites may support similar molecular functions of NEK4. However, experimental validation is still needed to confirm the biological relevance of these coordinated signaling patterns. The co-occurrence frequencies for all NEK4 phosphosites are listed in the Supplementary Table S15.
Co-regulated protein phosphosites in currently known binary interactors of NEK4
As phosphorylation influences protein-protein interactions, we curated a set of proteins known to interact with NEK4 from databases such as BioGrid and IntAct (Davis, 2011). Phosphosites in 376 NEK4-interacting proteins were identified to be co-regulated with predominant sites of NEK4. Among these, 211 phosphosites exhibited positive co-regulation, whereas 98 showed negative co-regulation, and the list of binary interactors is given in Supplementary Table S11. Figure 2 depicts the phosphosites of these interactors that demonstrate strong co-regulation with NEK4 predominant sites. Notably, phosphorylation at S306 of Lamina-associated polypeptide 2, isoforms beta/gamma (TMPO), S1041 of Retinoblastoma-like protein 1 (RBL1), and T227 of Splicing factor 3B subunit 1 (SF3B1) showed the highest frequency in 21 datasets, and they are positively co-regulated with S563 of NEK4. Interestingly, multiple phosphosites in TMPO (T74, S351, S424, S67, S66) showed positive co-regulation with the predominant sites of NEK4. Specifically, Phosphorylation at T74 was co-regulated with S563, while phosphorylation at S351, S424, S67, and S66 were co-regulated with S661 of NEK4. Additionally, phosphorylation at S706 of Ribosomal RNA processing protein 1 homolog B (RRP1B) was positively co-regulated with S461 and S639 of NEK4 in 18 and 8 datasets, respectively. Phosphorylation at S513 of RRP1B was positively co-regulated with S461 and S639 of NEK4 in 17 and 9 datasets. Furthermore, phosphorylation at S245 of RRPIB was positively co-regulated with S461, S661 and S639 of NEK4 in 9 datasets and 8 datasets for both S661 and S639. Phosphorylation at T1589 of Mediator of DNA damage checkpoint protein 1 (MDC1) was also positively co-regulated with S563 and S461 of NEK4 in 9 and 7 datasets, respectively. Despite their consistent co-regulation with NEK4, the precise molecular roles of these phosphosites remain unclear. Therefore, further investigations will be necessary to define the functional relevance of these signaling patterns.
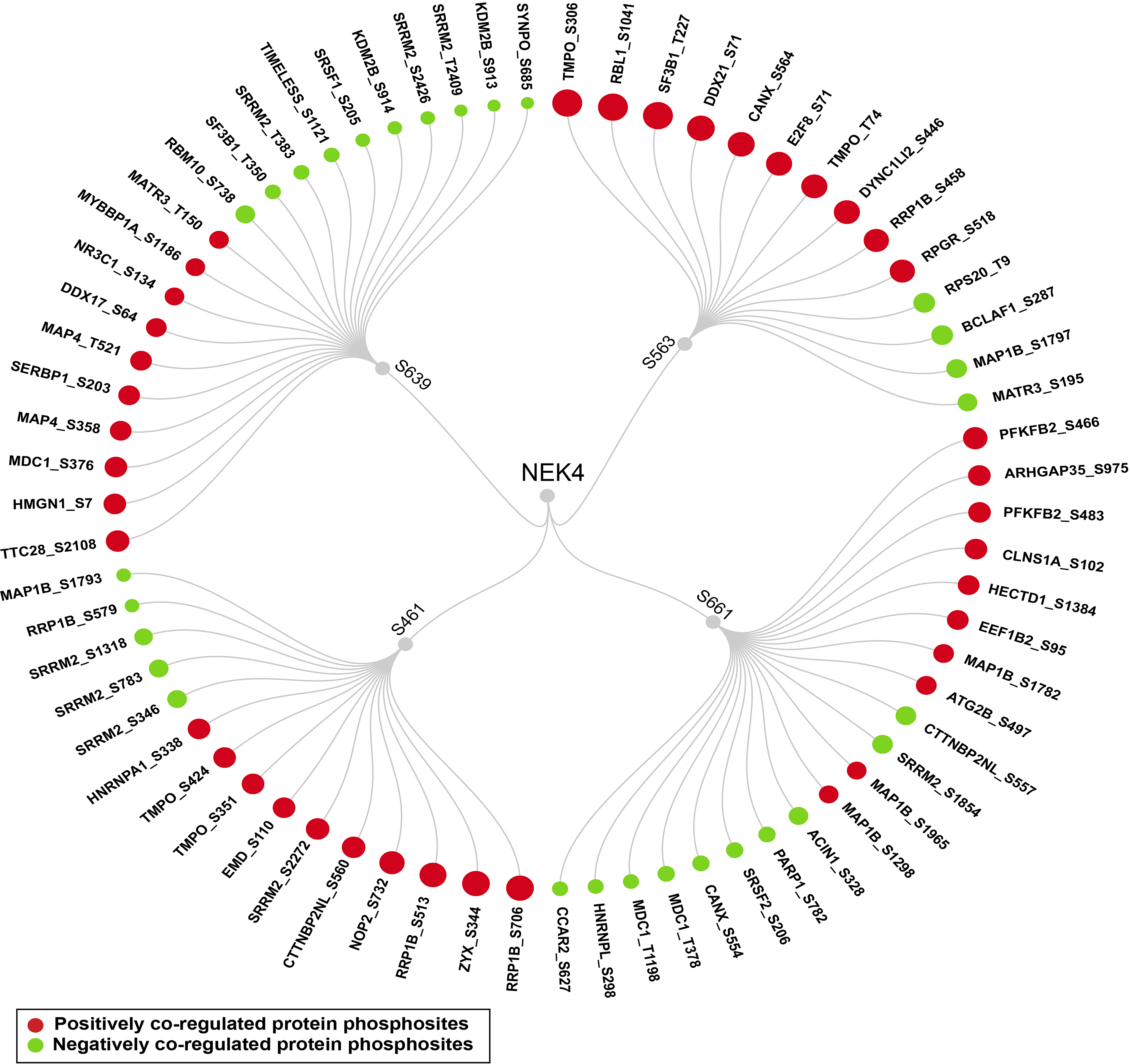
Binary interactors of NEK4. Positively and negatively coregulated binary interactors of NEK4 predominant sites.
Co-regulated upstream kinases and kinase phosphosites associated with predominant NEK4 phosphorylation
Currently, no experimentally validated upstream kinases have been identified that phosphorylate the predominant sites of NEK4. However, to uncover the potential upstream kinases that may phosphorylate these NEK4 phosphosites, we employed in silico prediction tools and incorporated peptide-based kinase-substrate screening data from Johnson et al. (2023) and Yaron-Barir et al. (2024). To evaluate the reliability of these predictions, we performed co-regulation analysis between the phosphorylation levels of the predicted upstream kinases and the corresponding NEK4 phosphosites, using this relationship as a confidence measure for potential coordinated signaling. Phosphosites from 33 predicted upstream kinases showed co-regulation with the predominant phosphosites of NEK4; among them, 31 exhibited positive co-regulation and two exhibited negative co-regulation. Our co-regulation analysis identified that phosphorylation at T1246, T1058 and S1054 of Cyclin-dependent kinase 13 (CDK13) was positively co-regulated with S563 of NEK4. Phosphorylation at S419 and T507 of MAP/microtubule affinity-regulating kinase 3 (MARK3) was positively co-regulated with S661 of NEK4. Phosphorylation at S380, T359 and S369 of Ribosomal protein S6 kinase alpha-1 (RPS6KA1) was positively co-regulated with S661 and S639 of NEK4. Phosphorylation at S227 and S415 of Ribosomal protein S6 kinase alpha-3 (RPS6KA3) was positively co-regulated with S661 and S639 of NEK4. Phosphorylation at S309, T507 and S506 of Protein kinase C delta type (PRKCD) was positively co-regulated with S661 and S639 of NEK4, respectively. Phosphorylation at S205 and S208 of Serine/threonine-protein kinase D1 (PRKD1) was positively co-regulated with S639 of NEK4, respectively. Interestingly, the phosphorylation sites S227 of RPS6KA3, S241 of PDPK1, and T507 of PRKCD have been reported to induce the enzymatic activity of these kinases and contribute to carcinogenesis (Shimura et al., 2012; Viswanathan et al., 2022; Yamada et al., 2021). The observed co-regulation patterns, particularly those involving activating kinase sites, suggest that NEK4 may interact with its predicted upstream kinases in a more complex and coordinated manner than initially expected. The list of positively and negatively co-regulated predicted upstream kinases, as reported by Johnson et al. (2023) and htp, is detailed in Supplementary Table S12 and illustrated in Figure 3.
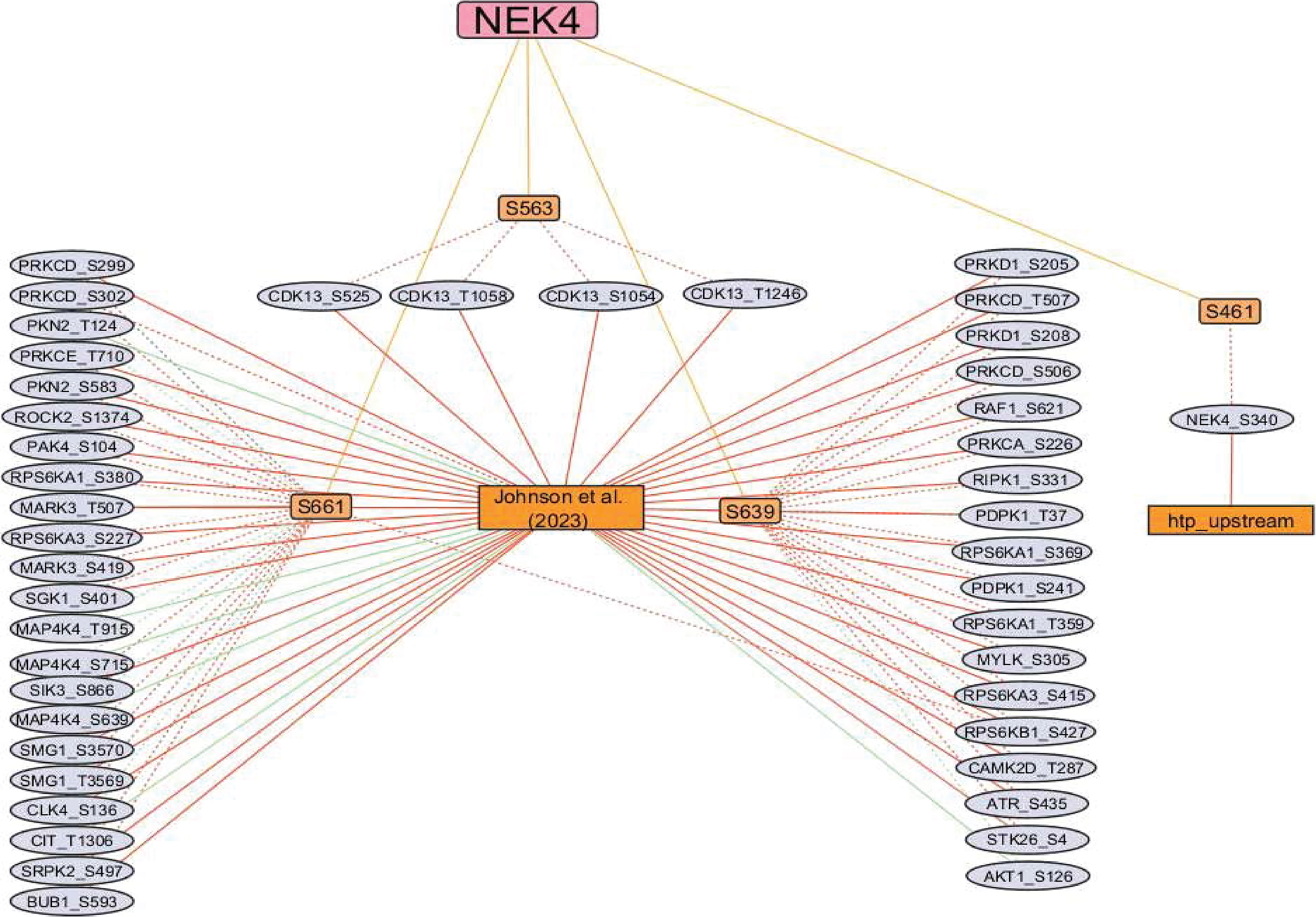
Upstream kinase coregulation network of NEK4. Red edges indicate positive co-regulation, whereas green indicates negative coregulation. Orange edges represent the predominant sites of NEK4.
Given the central role of kinases in phosphorylation-based regulation and their reliance on phosphorylation for functional modulation within complex signaling networks, we analyzed kinase phosphosites that displayed consistent co-regulation with the predominant NEK4 phosphosites. Phosphosites from a total of 253 kinases were identified among the high-confidence co-regulations with the NEK4 predominant phosphosites. Among them are phosphosites that enhance kinase activity, notably Serine/threonine-protein kinase N1 (PKN1) (S562) and 3-phosphoinositide-dependent protein kinase 1 (PDPK1) (S241), which were positively co-regulated with S563 of NEK4 with frequencies of 16 and 12 datasets, respectively. Additionally, ATP-dependent 6-phosphofructokinase, platelet type (PFKP) (S386), PDPK1 (S241), 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3 (PFKFB3) (S461), and Protein kinase C delta type (PRKCD) (T507) were positively co-regulated with S639 of NEK4 with frequencies of 11, 8, 8 and 7 datasets. Furthermore, Ribosomal protein S6 kinase alpha-3 (RPS6KA3) (S227) and Mitogen-activated protein kinase 14 (MAPK14) (Y182) were positively co-regulated with S661 of NEK4, with frequencies of 9 and 8 datasets. Serine/threonine-protein kinase PAK 4 (PAK4) (S474) and Ribosomal protein S6 kinase beta-1 (RPS6KB1) (T444) were positively co-regulated with S461 of NEK4 with frequencies 8 and 6 datasets. These phosphorylation events have been reported to increase the kinase activity of their respective kinases and contribute to carcinogenesis (Deng et al., 2024; Lim et al., 2022; Park et al., 2018; Shimura et al., 2012; Thirusangu et al., 2022; Viswanathan et al., 2022; Yamada et al., 2021; Zeng et al., 2020). Detailed information on these kinases and their co-regulated phosphosites is provided in Supplementary Table S14.
Inferred downstream substrates coregulated with the predominant phosphosites of NEK4
Although NEK4 functions as a serine/threonine kinase, its downstream phosphorylation signaling is still poorly understood. Currently, no experimentally validated downstream substrates are known to phosphorylate the predominant sites of NEK4. Using an approach similar to upstream kinase prediction, we identified multiple putative NEK4 substrates computationally. Their potential importance was assessed by analyzing how their phosphosites coregulated with the NEK4 predominant phosphosites. Yet, the biological functions of these sites have not been defined and remain part of the largely unexplored dark phosphoproteome.
Additionally, 56 PsOPs were predicted as NEK4 substrates by Johnson et al. (2023), 125 PsOPs were predicted as NEK4 substrates by NetworkIN/AKID, and one PsOP was predicted as a high-throughput (htp) substrate of NEK4 predominant sites. Exhibiting consistent coregulation with NEK4 predominant sites. The distribution of predicted substrate phosphosites showing positive or negative coregulation is presented in Table 2. Among these, phosphorylation at S538 of proliferation marker protein Ki- (MKI67) exhibited positive coregulation with NEK4 (S661, S461 and S639), which was predicted as a downstream substrate by Johnson et al. (2023) and NetworKIN/AKID in 8,12, and 6 datasets. Similarly, phosphorylation at S1207 of MKI67 exhibited positive co-regulation with NEK4 (S661, S461 and S639), was predicted as a downstream substrate by Johnson et al. (2023) and NetworKIN/AKID in 6, 9, and 6 datasets. Phosphorylation at T219 and S197 of inner centromere protein (INCENP), predicted by NetworkIN/AKID as a potential downstream substrate of NEK4, showed positive co-regulation with NEK4 S563 and S461, but negative coregulation with the S661 site. Specifically, T219 exhibited positive coregulation in 15 and 5.5 datasets and negative coregulation in 10 datasets, while S197 exhibited positive coregulation in 12 and 5 datasets and negative coregulation in 7 datasets, respectively. These consistent co-regulation patterns between predominant NEK4 phosphosites and predicted substrate sites strengthen the confidence in these candidates as potential NEK4 substrates and establish the basis for future experimental validation. The list of positively and negatively co-regulated downstream substrates is detailed in the Supplementary Table S13 and illustrated in Figure 4.
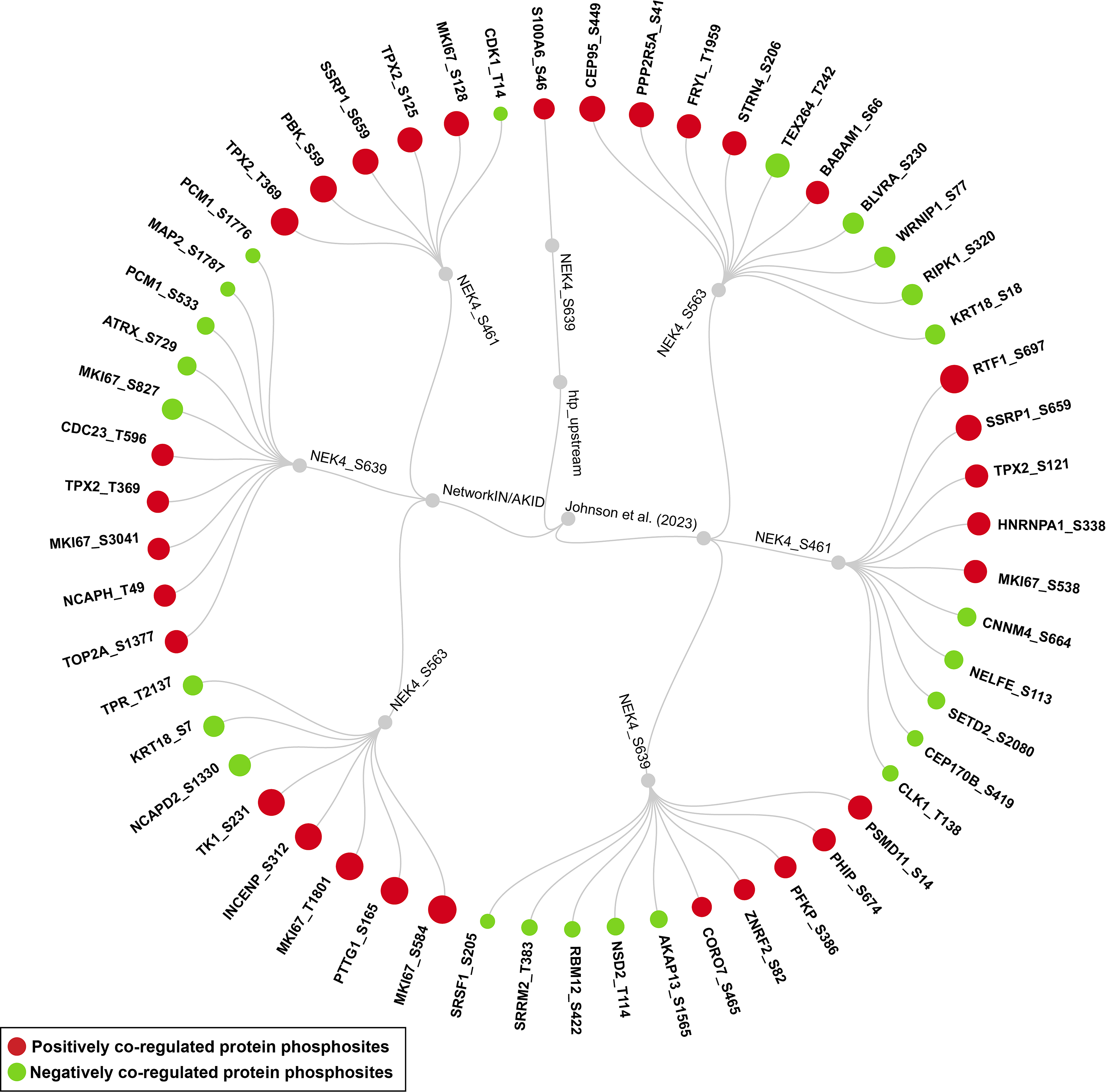
Downstream substrates associated with NEK4. Circular dendrogram showing positively and negatively co-regulated downstream substrates of NEK4 predominant sites.
Number of Predicted Substrate Phosphosites Showing Expression Coregulation with Specific NEK4 Predominant Phosphosites

Phosphosite-specific protein enrichment identifies coregulation linked to predominant NEK4 phosphorylation sites
Although phosphorylation and other post-translational modifications (PTMs) play an important role in cellular signaling, phosphoproteomic and PTM datasets are commonly analyzed at the gene level due to the limited availability of site-resolved PTM data and analytical approaches (Krug et al., 2019). The PhosphositePlus database offers curated, site-resolved functional information on phosphosites extracted from experimental studies (Hornbeck et al., 2012). To assess the molecular significance of the consistent co-regulation patterns, we used PhosphositePlus to retrieve and map known functional annotations for phosphosites associated with the predominant sites of NEK4. The phospho-signaling networks consistently coregulated with the NEK4 predominant sites were primarily associated with cell cycle regulation, apoptosis, cell growth and autophagy. This suggests that phospho-signaling associated with NEK4 may modulate a wide range of cellular functions. A detailed overview of the site-specific functional annotations retrieved from PhosphositePlus is presented in the Supplementary Tables S3, S4, S5, S6, S7, S8, S9, and S10 and visualized in Figure 5.
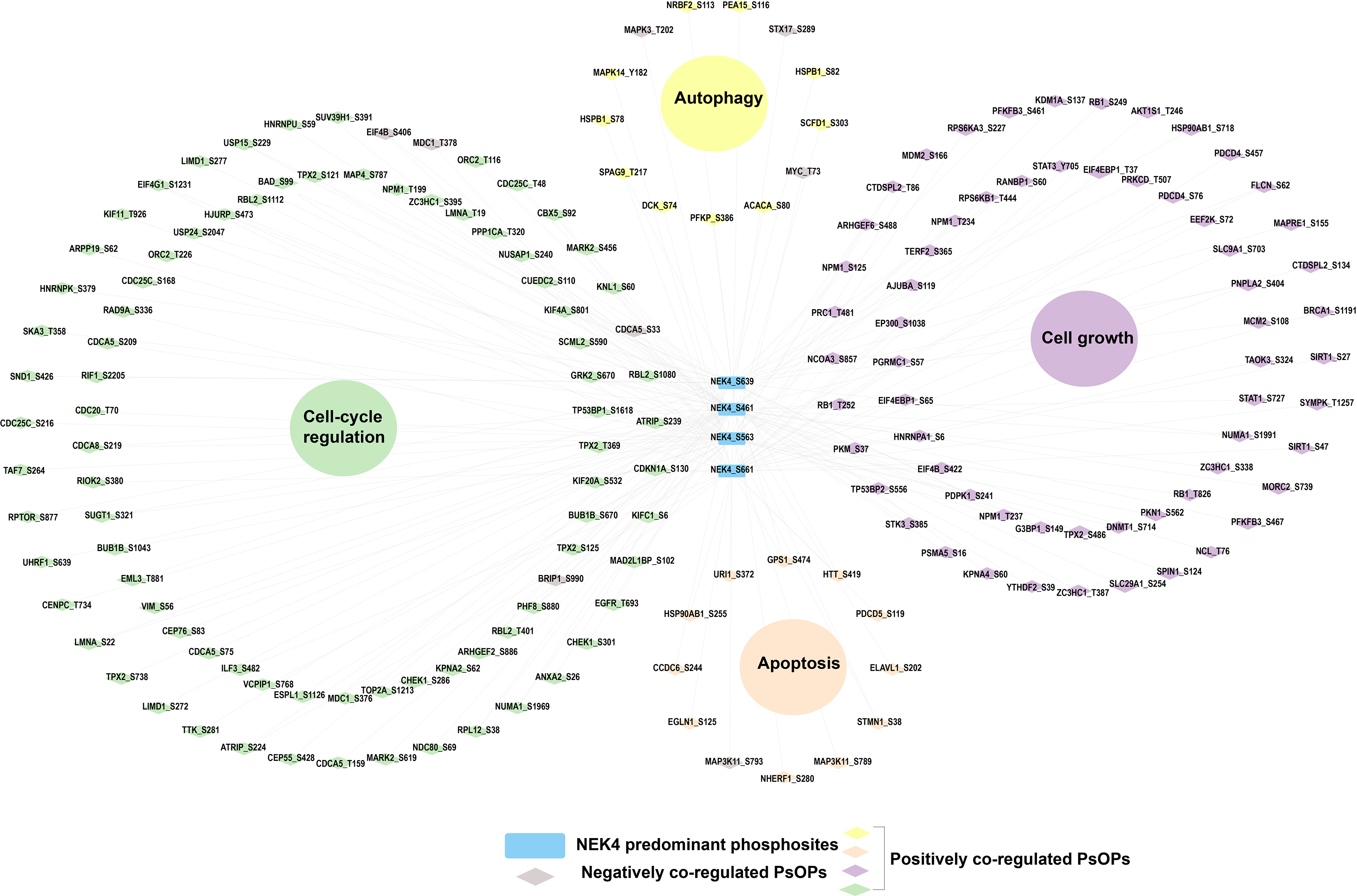
Functional enrichment of phosphoproteins coregulated with NEK4’s predominant sites. Network-based functional enrichment analysis illustrating biological processes associated with phosphoproteins that are positively and negatively coregulated with predominant NEK4 phosphosites. Enriched functional categories are shown as large colored circles, highlighting major biological processes including cell-cycle regulation, cell growth, autophagy, and apoptosis.
Discussion
Our phosphoproteomic analysis of NEK4 reveals a complex landscape of site-specific phosphorylation events that underscore its multifaceted roles in cellular signaling, particularly in DDR and cancer-related pathways. By integrating qualitative and quantitative datasets from over 3,800 studies, we identified 31 Class-1 phosphosites in NEK4, with four predominant sites, S563, S661, S461, and S639, exhibiting high detection frequencies and differential regulation across diverse biological contexts. These sites, located outside the kinase domain, align with NEK4’s classification as a PoKD kinase, consistent with prior kinome-wide classifications by Gopalakrishnan et al., 2025.
The co-regulation analysis of PsOPs with NEK4’s predominant sites provide compelling evidence for NEK4’s integration into broader phospho-signaling networks. Specific examples include the positive co-regulation of NEK4 S563 with EIF4G2 T508 and ESCO2 S75, NEK4 S661 with ACLY S455 and ZC3H4 S1275, NEK4 S461 with RRP1B S706 and RIF1 S2196, NEK4 S639 with CTDSPL2 S28 and PPP2R5D S573, and BAG3 T406 as the sole PsOP positively co-regulated with all four predominant NEK4 sites. Co-occurrence patterns among NEK4’s own phosphosites further support functional interdependence, as proposed by Li et al. (2017). Pairs like S461–S563 and S340–S563 displayed predominantly positive co-occurrence. Phosphosites in 376 known binary interactors of NEK4 (e.g., multiple phosphosites in TMPO such as S306 and T74, and in RRP1B such as S706) likewise exhibit high-frequency co-regulation with the predominant NEK4 sites. Predicted upstream kinases (e.g., CDK13, MARK3, RPS6KA1/3, PRKCD) and downstream substrates (e.g., MKI67 S538/S1207, INCENP T219/S197) display corresponding co-regulation patterns with NEK4 sites. Functional annotation of co-regulated PsOPs via PhosphositePlus consistently maps to cell-cycle regulation, apoptosis, autophagy, and growth.
These observations constitute data-driven conclusions supported directly by the integrated phosphoproteomic datasets which are discussed in subsequent sections.
The predominance of the four NEK4 sites suggests they may serve as key regulatory hubs, potentially modulating NEK4’s activity in response to cellular stresses, such as DNA damage or oncogenic signaling. The co-regulation analysis of PsOPs with NEK4’s predominant sites point to potential involvement in translational control and sister chromatid cohesion (e.g., NEK4 S563 with EIF4G2 T508 and ESCO2 S75), processes critical for genome stability during cell division. Similarly, co-regulation with proteins such as ACLY S455 (linked to metabolic reprogramming) and ZC3H4 S1275 (involved in RNA processing) at NEK4 S661 suggests isoform-specific functions, echoing earlier distinctions between NEK4.1 and NEK4.2 in DNA repair and apoptosis (Basei et al., 2015). The identification of BAG3 T406 as a PsOP that positively co-regulates all four predominant sites highlights a potentially conserved mechanism, possibly involving chaperone-mediated autophagy or stress responses, given BAG3′s role in protein quality control.
Co-occurrence patterns among NEK4’s own phosphosites imply cooperative phosphorylation that may enhance NEK4’s stability or activation. This is particularly relevant to DDR, where NEK4 interacts with the DNA-PK complex to facilitate nonhomologous end-joining (NHEJ) repair (Nguyen et al., 2012; Pavan et al., 2021). The observed patterns could explain how multisite phosphorylation coordinates NEK4’s recruitment to DNA damage sites, H2AX phosphorylation, and p53 signaling, thereby preventing premature dissociation of Ku70/Ku80 and bolstering cell cycle checkpoints.
Integrating binary interactors, upstream kinases, and downstream substrates into the analysis strengthens NEK4’s positioning within cancer-associated networks. Among interactors, multiple phosphosites in TMPO (e.g., S306, T74) and RRP1B (e.g., S706) showed strong positive co-regulation, suggesting NEK4 modulates nuclear lamina integrity and ribosomal RNA processing, processes disrupted in cancer. The lack of experimentally validated upstream kinases for these sites prompted our use of predictive tools, revealing candidates like CDK13, MARK3, and RPS6KA1/3, whose activating phosphosites (e.g., RPS6KA3 S227) co-regulate with NEK4 sites. These kinases are implicated in microtubule organization and EMT (Ding et al., 2018), aligning with NEK4’s context-dependent roles: tumor-suppressive in breast cancer via proliferation inhibition, yet pro-metastatic in lung cancer through EMT promotion. Downstream substrate predictions, including MKI67 (S538, S1207) and INCENP (T219, S197), link NEK4 to proliferation markers and centromere function, potentially explaining its ties to mitochondrial integrity and TRAIL sensitisation (Basei et al., 2022; Park et al., 2016).
Functional enrichment of co-regulated proteins emphasizes NEK4’s influence on cell cycle regulation, apoptosis, autophagy, and growth, and hallmarks of cancer. For example, co-regulation with activating sites in PKN1, PDPK1, and PRKCD suggests NEK4 amplifies PI3K/AKT/mTOR signaling, a pathway frequently dysregulated in tumors. This is consistent with NEK4’s overexpression promoting EMT and metastasis in lung adenocarcinoma (Ding et al., 2018), positioning it as a potential therapeutic target. However, the site’s roles in mitochondrial respiration and mtDNA protection (Basei et al., 2022) imply broader implications for metabolic reprogramming in cancer cells.
Limitations
Despite these insights, few limitations of the present study should be acknowledged. This study is based entirely on computational analyses, and thus all findings are predictive and correlative in nature. While the integration of large-scale phosphoproteomic datasets provides valuable insights into potential kinase–substrate relationships and phosphosite coregulation patterns, these associations do not establish causality. The observed coregulation may arise from direct interactions, indirect effects mediated by shared upstream regulators, or parallel responses to common signaling stimuli.
Importantly, the predicted upstream kinases, downstream targets, and functional implications of site-specific phosphorylation have not been experimentally validated (e.g., through kinase assays, phosphosite-specific antibodies, or mutagenesis studies). Therefore, the results should be interpreted with caution and considered as hypothesis-generating. Although the findings are biologically coherent and highlight plausible regulatory pathways, experimental validation such as confirming key kinase–substrate interactions or assessing the functional impact of specific phosphosites will be essential to substantiate these predictions. Future studies are required to validate these computational inferences and to elucidate the precise biological roles of the identified phosphorylation events.
Conclusions
In conclusion, our study provides the first comprehensive insight into the phospho-signaling networks mediated by the serine/threonine kinase NEK4, emphasizing its functional role in DDR and EMT. Through integrative phosphoproteomic analysis, we identified predominant phosphosites that exhibit significant regulatory alterations across diverse experimental conditions. We identified diverse phosphorylation signaling patterns associated with NEK4. The coregulation patterns revealed potential upstream kinases, downstream substrates and binary interactors associated with NEK4-mediated signaling, many of which are implicated in DDR pathways and transcriptional reprogramming during EMT. Furthermore, our analysis suggests that NEK4 phosphorylation events may coordinate cellular responses to genotoxic stress and contribute to the regulation of EMT-associated signaling networks that drive cancer progression. Collectively, these findings establish a foundation for future experimental investigations aimed at depicting the functional roles of NEK4 in DDR and EMT and underscore its potential as a therapeutic target in cancer biology.
Authors’ Contributions
S.S.K.: Formal analysis, investigation, visualization, writing original draft. A.P.: Formal analysis, writing, review and editing. L.D.: Review and editing. Visualization. A.P.G.: Data curation. A.C.R.: Supervision, review, editing. R.R.: Conceptualization, investigation, methodology, project administration, resources, supervision, review.
Data Availability Statement
All data generated or analyzed during this study are included in this published article and its Supplementary Data.
Footnotes
Acknowledgments
The authors thank Yenepoya (Deemed to be University) for providing infrastructure and other resources for the Center for Integrative Omics Data Science (CIODS).
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Supplemental Material
Supplemental Material
Supplemental Material
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.