
Abstract
Background:
Perivascular adipose tissue (PVAT), found outside blood vessels, enhances vasorelaxation when endothelium-mediated vasorelaxation fails, as observed in the mesenteric arteries of rats with metabolic syndrome (MetS) and hypertension; however, whether high blood pressure contributes to PVAT dysfunction remains unclear. We, therefore, aimed to investigate vascular function and PVAT modulation in the mesenteric arteries of rats with MetS but no hypertension.
Methods:
Mesenteric arteries were isolated from both lean and obese male Zucker fatty (ZF) and ZF diabetes mellitus (ZFDM) rats at 20 and 30 weeks of age. Vasorelaxation was examined in arteries with or without PVAT; mRNA levels in PVAT and arteries were examined using the organ bath method and quantitative reverse transcription polymerase chain reaction, respectively.
Results:
Sodium nitroprusside-induced relaxations were lower in the obese versus lean ZF rats, and PVAT increased them to levels comparable to those in lean rats at 20 weeks. These enhancements disappeared after 30 weeks. The mRNA expression of apelin in PVAT and its receptor was upregulated in the arteries of obese rats, and its levels correlated positively with the enhancing effect of PVAT. In obese ZFDM rats, PVAT attenuated relaxation at both 20 and 30 weeks. Levels of chemerin mRNA expression in PVAT were higher in ZFDM versus ZF rats; however, treatment with chemerin or a chemerin receptor antagonist did not alter vasorelaxation in ZFDM rats.
Conclusions:
Apelin may contribute to compensatory PVAT modulation during vascular dysfunction in obese ZF rats. PVAT inhibits vasorelaxation by releasing a contractile factor other than chemerin in ZFDM rats. Thus, it may modulate vascular tone differently depending on MetS stage.
Introduction
Obesity and insulin resistance lead to several metabolic disorders. Metabolic syndrome (MetS) is a term used to characterize the accumulation of multiple such disorders. MetS promotes cardiovascular disease caused by atherosclerosis and type 2 diabetes through vascular dysfunction, for example, inflammation, oxidative stress, and impaired endothelial function.1,2 Perivascular adipose tissue (PVAT), located outside the blood vessels, increases as visceral obesity develops. This tissue regulates vascular homeostasis (including tone, inflammation, and remodeling) by releasing adipokines.3–5 Regarding PVAT modulation of vascular tone, Soltis et al. reported that it regulates contractile responses in normal rat aortas. 6 PVAT is now known to modulate vasomotor function by releasing vasorelaxing or vasoconstricting adipokines that contribute to MetS-associated cardiovascular disease.
A recent review divided adipokines into three domains 5 : Domain I adipokines are cardioprotective and secreted by healthy adipose tissue; Domain II adipokines are cardioprotective, and they are secreted by adipose tissue at higher levels in conditions such as obesity and visceral adiposity, as a compensatory mechanism; and Domain III adipokines are secreted by hypertrophied or hyperplastic inflamed adipocytes, causing cardiomyocyte dysfunction, cardiac and vascular hypertrophy, coronary microvascular disease, and hypertension. Apelin, the Domain II adipokine, 5 is an adipocyte-derived vasodilator that induces nitric oxide (NO)-dependent vasorelaxation to regulate blood pressure.7,8 Exogenous apelin enhances NO-induced relaxation in rat mesenteric arteries. 9 Its mRNA is expressed highly in adipose tissue, and its serum concentration is correlated with higher body mass indexes in obesity.10,11 Chemerin, the Domain III adipokine, 5 induces vasoconstriction by activating chemerin-like receptor 1 (CMKLR1) in rat thoracic aortas. 12 Exogenous chemerin-9, its active fragment, causes concentration-dependent contractions via CMKLR1 in isolated rat thoracic aortas, as well as mesenteric and pulmonary arteries.13–15 Circulating chemerin levels correlate positively with the accumulation of abdominal visceral fat 16 and are associated with obesity and MetS.17,18 Endogenous chemerin regulates blood pressure via vasoconstriction through CMKLR1 activation in smooth muscle cells.19,20
Our group assessed the role of PVAT in vascular dysfunction using SHRSP.Z-Leprfa/IzmDmcr rats (SHRSP.ZF), which were established as an animal model of hypertensive MetS by crossing stroke-prone spontaneously hypertensive Izumo strain rats with Zucker fatty (ZF) rats. 21 The mesenteric PVAT of these rats retains vascular tonus in response to NO even under vascular dysfunction; however, this favorable PVAT function is lost during MetS progression.9,22 Apelin levels in PVAT contribute to this compensatory system in the mesenteric9,23 and renal arteries 24 of SHRSP.ZF rats; however, whether high blood pressure affects PVAT modulation and its dysfunction remains unclear. Therefore, we aimed to investigate its effects on vasorelaxation in two rat models of nonhypertensive MetS: ZF and ZF diabetes mellitus (ZFDM; a substrain of ZF). In addition, we sought to investigate whether aging affects PVAT modulation by comparing its function between rats aged 20 and 30 weeks, as the compensatory effects of PVAT have been observed at 20 weeks of age in male SHRSP.ZF rats22,25,26 but not in those aged 30 weeks. 22 We specifically focused on the roles of apelin and chemerin 5 to identify which adipokines are most involved in the effects of PVAT on the modulation of vascular tonus in MetS.
Materials and Methods
Experimental animals
All protocols were approved by the Animal Care Committee of the Mukogawa Women’s University (approval numbers P-12-2017-01-A, P-12-2023-01-A, and P-12-2024-01-A). ZF and ZFDM rats spontaneously exhibit MetS caused by obesity, resulting from a mutation in the fatty acid (fa) gene that generates a defective leptin receptor, although their systolic blood pressure levels are comparable to those of lean rats.27,28 ZF rats are generally less likely to develop diabetes because the insulin secretion ability of their pancreas is maintained. ZFDM rats were selectively bred to create a strain of naturally occurring diabetes from a colony of ZF rats. Herein, 20- and 30-week-old male ZF-Leprfa/fa (obese ZF, n = 6), normal control ZF-Lepr+/+ (lean ZF, n = 6), ZFDM-Leprfa/fa (obese ZFDM, n = 8), and normal control ZFDM-Leprfa/+ (lean ZFDM, n = 8) rats were used. All rats were purchased from Japan SLC, Inc. (Hamamatsu, Japan) and housed within an animal care center with a 12-h light–dark cycle, at a constant room temperature of 21°C–22°C and humidity of 35%–65%. They were provided standard chow (CE-2; CLEA Japan Inc., Tokyo, Japan) and water ad libitum during the experimental period. Each experiment was initiated after a 1-week habituation period.
Metabolic parameters
Body weight, body length, and waist length were measured under anesthesia (isoflurane). Afterward, blood was drawn from the abdominal aortas of nonfasting rats. Serum glucose and insulin levels were determined using commercial kits: glucose CII-test (FUJIFILM Wako Pure Chemical Co., Osaka, Japan) and a rat insulin detection kit (Morinaga Biochemistry Lab, Tokyo, Japan). Urine glucose levels were recorded using a UROPAPER III “Eiken” instrument (E-UR80, Eiken Chemical Co. Ltd., Tochigi, Japan) on a 5-point scale [minus (none detected) = 0; 50 mg/100 mL = 1; 100 mg/100 mL = 2; 250 mg/100 mL = 3; 500 mg/100 mL = 4]. Systolic blood pressure was measured via the tail-cuff method using an MK-2000 blood pressure monitor (Muromachi Kikai Co., Tokyo, Japan) 1 week before the vasodilation studies, according to a previously described protocol. 29
Vasodilation
Superior mesenteric arteries with PVAT were harvested from the rats and prepared for myographic experiments following a previously described protocol. 22 Briefly, they were cleaned or not cleaned of surrounding PVAT, cut into approximately 3 mm-wide rings, and mounted isometrically at an optimal resting tension (0.3 grams in 10 mL organ baths). Isometric contractions in response to the cumulative addition of phenylephrine were recorded for each ring. After washout and equilibration at resting tension, each ring preparation was contracted with phenylephrine (2 μM) to ∼80% of its maximum. Once stable contractions were obtained, acetylcholine and sodium nitroprusside (SNP) were added cumulatively to the bath. Some ring preparations with or without ZFDM PVAT were treated with 1 μM human chemerin-9 (item 36760; Cayman Chemicals, Ann Arbor, MI, USA) or 10 μM 2-(α-naphthoyl) ethyltrimethylammonium iodide (αNETA; item AG-CR1-0151; AdipoGen Life Sciences, San Diego, CA, USA), a CMKLR1 antagonist, for 30 min; afterward, SNP-induced relaxation was measured. Isometric tension changes were detected using a force-displacement transducer (t-7; NEC San-Ei, Tokyo, Japan) coupled to a chart recorder (8K21; NEC San-Ei). Individual concentration–response curves were analyzed using nonlinear regression curve fitting of the relaxation–drug concentration relationships. Negative log(EC50) and Emax were determined using GraphPad Prism 10 for MacOS (GraphPad, San Diego, CA, USA). The relaxation-enhancing effects of PVAT were assessed based on the differences in the maximum responses of the mesenteric arteries with or without PVAT to 1 μM of SNP.
Expression of mRNA expression
Quantitative reverse transcription polymerase chain reaction (PCR) assays were performed to examine the mRNA expression levels of apelin (apln) and chemerin (Rarres2) in PVAT, as well as apelin receptor (apelin-R, aplnr), CMKLR1 (Cmklr1), endothelium NO synthase (eNOS, nos3), and soluble guanylyl cyclase (sGC, Gucy1b) in the mesenteric arteries of 20-week-old ZF and ZFDM rats, following a previously described protocol. 9 Real-time measurements of target gene expression were performed using a TaqMan RNA-to-CT 1-step kit and a LightCycler 1.5 instrument (Roche Diagnostics Japan K.K., Tokyo, Japan). Commercially available gene-specific probes were used (Roche Applied Science Universal Probe Library product IDs: apelin, 0486896001; apelin-R, 04685148001; ribosomal protein 18S, 04688937001; β-glucuronidase, 04688015001; β-actin, 04686900001), and gene-specific primers designed by Assay Design Center (Roche Applied Science) were purchased from Life Technologies Japan, Ltd. (Tokyo, Japan). TaqMan gene expression assays (a pair of PCR primers and TaqMan labeled with dye 4331182; product IDs: chemerin, Rn01451853_m1; CMKLR1; Rn00573616_s1) were purchased from Thermo-Fisher Scientific K.K. (Tokyo, Japan). A triad housekeeping gene expression approach (ribosomal protein 18S, β-glucuronidase, and β-actin) was used to normalize the sample material, and the efficiency levels for each primer set were included in all calculations. The amount of target gene was normalized to the reference gene to obtain the relative threshold cycle (ΔCT) and then related to the CT of the level in the 20-week-old lean ZF rats to obtain the relative expression level (2–ΔΔCT) of the genes.
Statistical analysis
Data were expressed as means ± standard errors of the mean. The means between groups were analyzed using one-way analysis of variance (ANOVA) followed by the Bonferroni post hoc test. Linear regression analysis was performed. All tests were performed in GraphPad Prism 10. Statistical significance was set at P < 0.05.
Results
The key phenotypic characteristics of the ZF and ZFDM rats at 20 and 30 weeks of age are summarized in Tables 1 and 2. The body weight in obese ZF was higher than that in lean rats at both ages (Table 1), whereas that in obese ZFDM was significantly lower than that in lean rats at 30 weeks of age (Table 2). This may be caused by long-term uncontrolled diabetes. Waist circumference and body length, indicating abdominal obesity, were significantly higher in the obese rats than in the lean ones in both strains at every age. Systolic blood pressure in the obese rats was lower than or comparable to that in the lean rats of both strains at 20 weeks of age. No significant differences were observed in serum glucose levels between the obese and lean rats; serum insulin levels were higher in the obese ZF rats, whereas serum glucose levels were higher in the obese ZFDM rats. High urine glucose levels were detected only in obese ZFDM rats in both age groups.
Key Phenotypic Characteristics of the ZF fa/fa (obese ZF) and ZF +/+ (lean ZF) Rats at 20 and 30 Weeks of Age
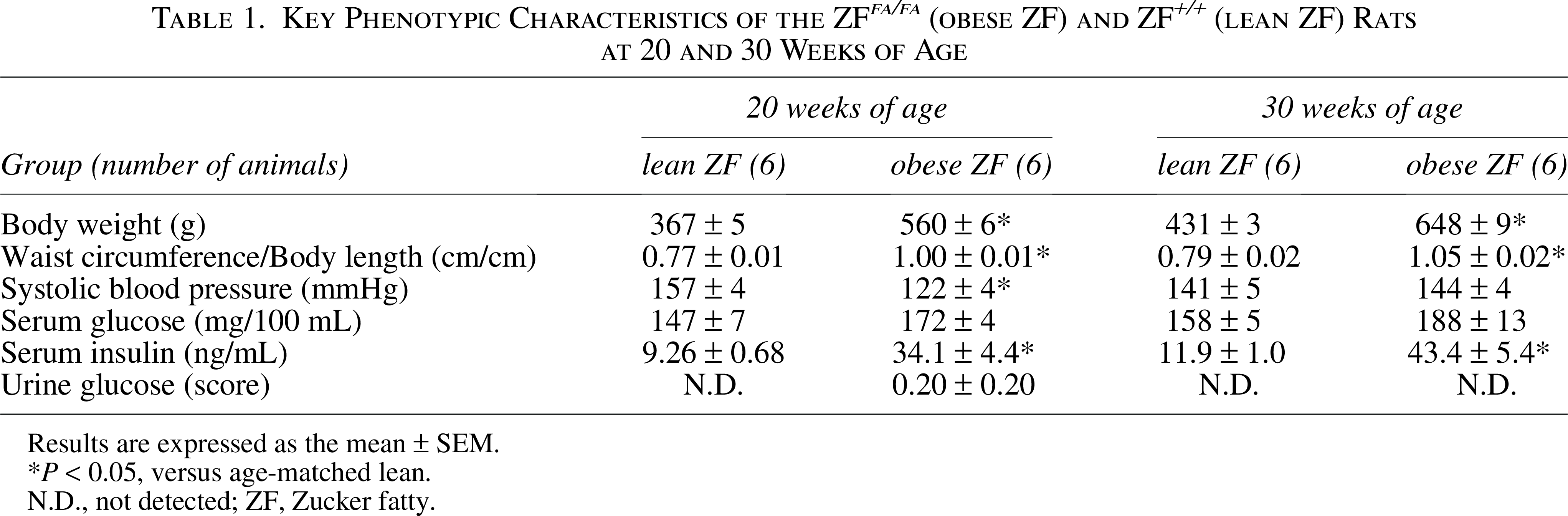
Results are expressed as the mean ± SEM.
P < 0.05, versus age-matched lean.
N.D., not detected; ZF, Zucker fatty.
Key Phenotypic Characteristics of the ZFDM fa/fa (obese ZFDM) and ZFDM fa/+ (lean ZFDM) Rats at 20 and 30 Weeks of Age

Results are expressed as the mean ± SEM.
P < 0.05, versus age-matched lean.
ZFDM, ZF diabetes mellitus.
Relaxation in response to acetylcholine, which induces endothelial NO-dependent relaxation, appeared unaffected by the presence or absence of PVAT in the mesenteric arteries of the ZF rats at both 20 (Fig. 1A) and 30 (Fig. 1D) weeks. However, the SNP-induced relaxations, which indicate the NO response in vascular smooth muscle cells, were lower in the arteries without PVAT harvested from the obese rats at 20 weeks. These relaxations increased to levels comparable to those of the lean rats by the addition of PVAT (Fig. 1B). This enhancing effect of PVAT—a favorable effect of PVAT in maintaining normal relaxation response—disappeared after 30 weeks of age (Fig. 1E). No significant differences were observed in the phenylephrine-induced contractile responses between the lean and obese rats of either age, with or without PVAT (Fig. 1C, F).
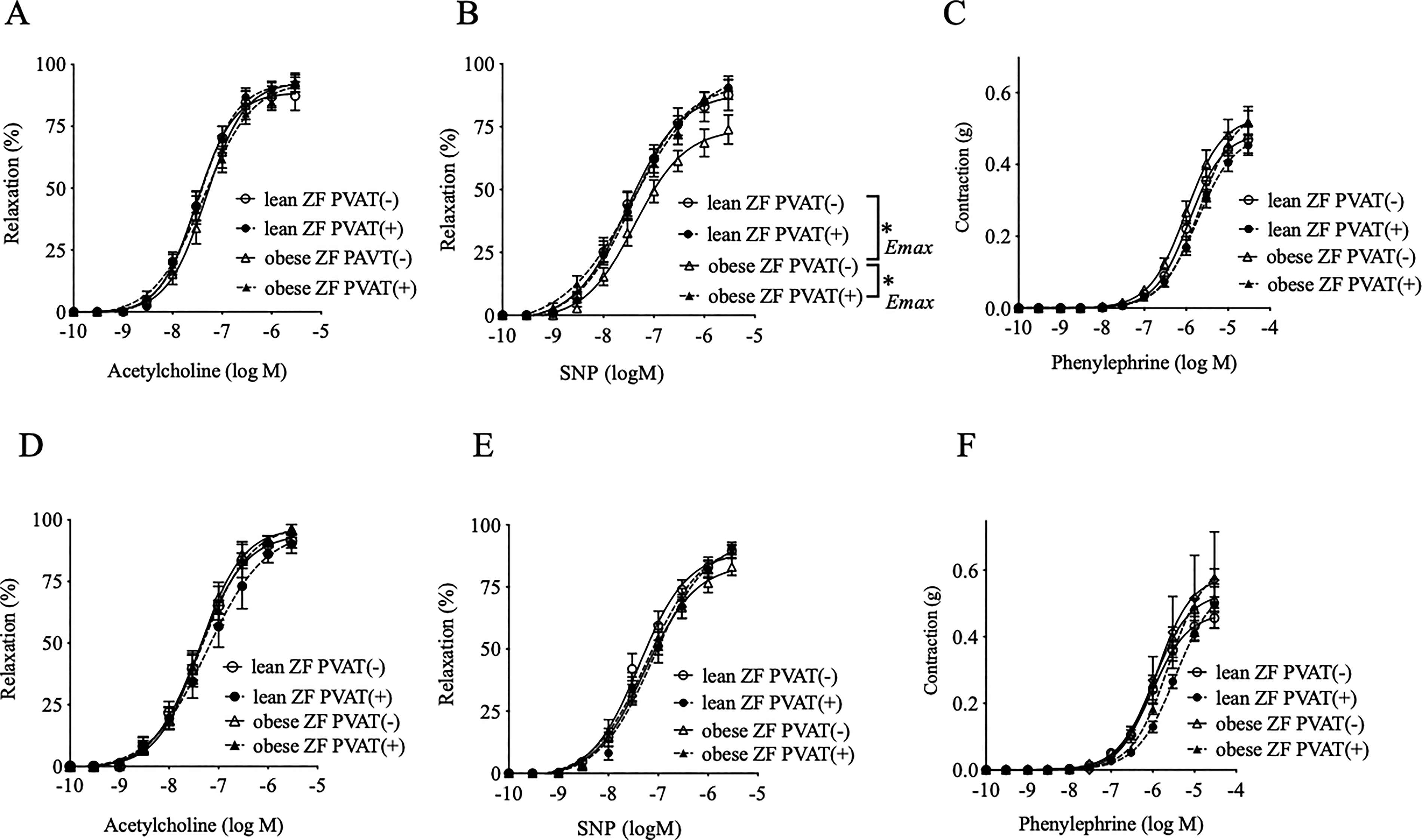
Vascular relaxations in response to acetylcholine
Acetylcholine-induced relaxation (Fig. 2A, D) and phenylephrine-induced contractions (Fig. 2C, F) were comparable between the obese and lean ZFDM rats at both 20 and 30 weeks, regardless of the presence or absence of PVAT. NO-donor-induced relaxation was unchanged in the arteries without PVAT but attenuated by its presence in the obese rats at 20 weeks of age (Fig. 2B); the unfavorable/inconvenient effect continued until 30 weeks of age (Fig. 2E).
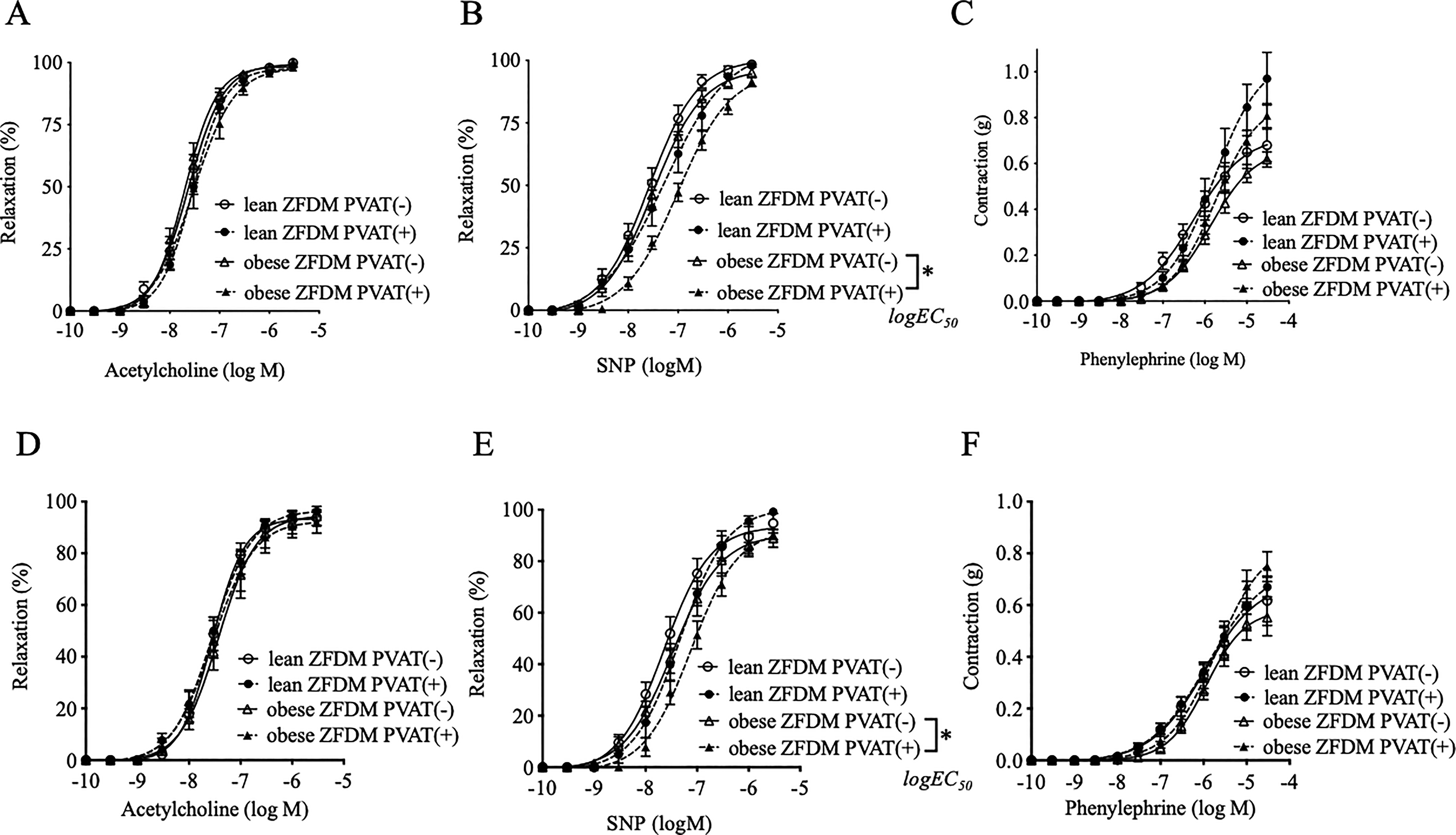
Vascular relaxations in response to acetylcholine
Apelin mRNA expression in PVAT (Fig. 3A) and its arterial receptor (Fig. 3B) was upregulated in the tissues of obese ZF rats and correlated positively with the enhancing effect of PVAT (Fig. 3C). The expression of eNOS mRNA (Fig. 3D) was increased, whereas that of sGC (a smooth-muscle NO receptor) was decreased in the arteries of obese ZF rats (Fig. 3E). Chemerin mRNA expression was higher in PVAT from ZDFM versus ZF rats (Fig. 4A); CMKLR1 mRNA levels were comparable in their arteries (Fig. 4B). SNP-induced relaxations were unchanged by treatment with chemerin-9 (Fig. 4C) and the αNETA chemerin receptor antagonist (Fig. 4D) in both the lean and obese ZFDM rats.

Expression levels of apelin mRNA in PVAT

Expression levels of chemerin mRNA in PVAT
Discussion
This study demonstrated that, in MetS, mesenteric PVAT regulates vasodilation by releasing apelin to compensate for impaired NO-induced vasodilation; however, the favorable PVAT effects disappeared with aging. However, at the diabetes stage—where MetS includes insulin resistance—PVAT inhibits vasodilation by releasing vasoconstrictive factors, and the unfavorable/inconvenient PVAT effects continue at a later age. As chemerin was not a significant factor in this study, we concluded that adipokine-based vascular modulation by PVAT differs depending on the stage of MetS.
Notably, there was no significant difference in endothelial NO-derived acetylcholine-induced relaxation between obese and lean ZF rats at both 20 and 30 weeks of age. Similarly, acetylcholine-induced relaxation does not impair the mesenteric arteries of obese ZF rats at 8–40 weeks of age and is comparable to that in lean rats aged 28–40 weeks. 30 However, endothelium-independent NO-donor-induced relaxation was decreased in 20-week-old obese ZF rats. This suggests that the NO response is impaired in the vascular smooth-muscle tissue of obese ZF rats, although endothelial NO synthesis is increased. This was supported by our findings that sGC mRNA levels were downregulated, whereas eNOS levels were coincidentally upregulated in the arteries of the obese ZF rats. Furthermore, PVAT enhanced NO-mediated relaxations in the obese rats. Upregulated apelin mRNA levels in the PVAT, which correlated with higher apelin-R mRNA levels in the arteries, were observed in the obese ZF rats. In addition, PVAT apelin mRNA levels correlated positively with the enhancing effects of PVAT on NO-induced vasorelaxation. Thus, apelin released from PVAT may contribute to the PVAT compensatory system to counteract the impaired vascular smooth muscle NO response in obese ZF rats. Exogenous apelin enhances NO-dependent relaxation in rat mesenteric arteries, 9 and endogenous apelin induces NO-dependent vasorelaxation in animals and humans.7,8 However, this compensatory apelin system was attenuated in obese ZF rats at 30 weeks of age. This PVAT and vascular bed apelin system may play a compensatory role that maintains normal vascular tonus during the early stages of MetS. Its disturbance may reduce blood supply to the organ, causing damage during the later MetS stages. Circulating apelin levels are significantly lower in patients with cardiovascular disease. 31 The apelin system has been noted in treatment approaches for cardiovascular diseases, 32 with apelin analogs and apelin receptor agonists showing therapeutic potential for MetS-related cardiovascular and kidney dysfunction. 33 However, further studies are warranted to identify which specific metabolic abnormalities affect PVAT functions in obese ZF rats. As a similar compensatory system involving PVAT apelin has been found in SHRSP.ZF rats with hypertensive MetS,9,23 high blood pressure can be excluded as a significant contributing factor to PVAT-based arterial modulation in ZF rats.
Acetylcholine-induced relaxations are progressively impaired in the mesenteric arteries of obese ZFDM rats aged 28–40 weeks versus 8–12 weeks and significantly smaller than those in lean rats aged 28–40 weeks. 30 Impaired acetylcholine-induced relaxation, but unaltered SNP-induced relaxation, has been reported in the mesenteric arteries of obese ZFDM rats aged 21–23 weeks. 34 The anticontractile effect of mesenteric PVAT has been previously reported in obese ZFDM rats aged 38 weeks, characterized by high blood glucose and insulin levels. 35 Our study showed unchanged contractile and relaxation responses; however, PVAT inhibited SNP-induced relaxations in response to NO in the smooth muscle cells of obese ZFDM rats, which had higher glucose but comparable serum insulin levels versus lean rats, at both 20 and 30 weeks of age. Our ZFDM rats (Hoshino Laboratory Animals Inc., Ibaraki, Japan) were previously reported to have higher serum insulin levels in obese rats (homozygous) than in lean rats (heterozygous) at 12 weeks of age, but these became comparable at 21 and 36–38 weeks, although serum glucose levels rose continuously over weeks 12–38. 27 Such serum insulin and glucose level differences may explain the variety of vascular and PVAT functions observed in ZFDM rats.
Chemerin treatment decreases relaxations induced by SNP and an sGC stimulator in aortas isolated from Wistar rats. 36 Exogenous chemerin-9, an active fragment, causes concentration-dependent contraction via CMKLR1 in isolated rat arteries.13–15 Chemerin mRNA expression is higher in the aortic PVAT of Dahl salt-sensitive rats; CCX832, a selective CMKLR1 receptor antagonist, inhibits PVAT-related vasomotor response effects. 12 Similar inhibitory actions of CCX832 on PVAT modulation have also been reported in the arteries of Sprague-Dawley rats.13,19 In this study, PVAT levels of chemerin mRNA were higher in ZFDM rats than in ZF rats, but no such difference was observed between obese and lean ZFDM rats. Treatment with chemerin-9 and αNETA, a chemerin receptor antagonist, did not affect arterial relaxation in obese ZFDM rats. Thus, it may not contribute to PVAT-induced inhibitory modulation in ZFDM rats with obesity and diabetes. A lack of modulatory role for PVAT-derived chemerin has also been reported in human coronary arteries. 37
This study has some limitations. The factors that affect PVAT function in obese ZF and ZFDM rats remain unclear, and the contributing PVAT-derived contractile factors in obese ZFDM rats remain to be elucidated. Whether the appearance or disappearance of PVAT-based vascular tonus modulation leads to cardiovascular disease in MetS also diabetes merits further study.
Conclusions
This study demonstrated that PVAT compensates for impaired NO-induced vasorelaxation by releasing apelin during early-stage MetS but releases other factors that impair vasorelaxation during the later diabetes stage. The adipokine-based regulation of vasomotor function by PVAT also varies according to MetS stage.
Authors’ Contributions
S.K. designed the experiments; S.K., K.M.-F., and A.N. performed the experiments and analyzed the data; S.K. and K.S. acquired the funding for the study and supervised its execution; and S.K. interpreted the data and wrote the original draft of the article, as well as reviewed and edited it. All of the authors have read and agreed to the final submitted version of the article.
Data Availability
The data presented in this study are available upon request from the corresponding author.
Footnotes
Acknowledgments
The authors thank Ms. Miho Shimari, Ms. Yui Yamada, Ms. Chinatsu Shiga, Ms. Ayana Nakata, Ms. Ayaka Fujiwara, and Ms. Seina Izumo from the Mukogawa Women’s University for their technical support. The authors thank Editage for their professional assistance with editing the article for proper English.
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
This study was partially supported by the JSPS KAKENHI (grant number JP21K06777, awarded to S.K.).