
Abstract
Emerging contaminants (ECs) represent a risk for health and the environment at concentrations as low as nanograms per liter. In this study, acesulfame-K (ACK), acetaminophen (ACE), caffeine (CAF), and N, N-diethyl-m-toluamide (DEET) were selected based on their ubiquity and concentrations of 16.9 μg ACK/L, 4.7 μg ACE/L, 3.1 μg CAF/L, and 3.7 μg DEET/L measured in real domestic wastewater. Biological oxygen consumption by activated sludge in the presence of ECs, at the milligram per liter level, suggested a biodegradability degree of ACE > DEET > CAF > ACK, supported by removal of 100% ACE, 99% DEET, 97% CAF, and 90% ACK, verified by liquid chromatography on parent compounds. Metagenomic analysis of the activated sludge identified Aeromonas, Macellibacteroides, Ottowia, Tetrasphera, and Bdellovibrio among the most abundant bacterial genera. Sequential plate cultures, matrix-assisted laser desorption ionization-time of flight mass spectrometry, and 16S rRNA gene sequencing allowed us to report here for the first time four strains of Aeromonas tolerant to 100 mg DEET/L isolated from activated sludge. Also, single bacterial strains of Aeromonas, Shewanella, and Bacillus tolerant to 100 mg ACE/L were equivalently isolated and identified. These strains represent promising candidates for further studies in bioremediation and biodegradation of ECs. The findings enhance our understanding of ECs occurrence, fate, and biological degradation in wastewater treatment systems and highlight the potential of native microbial communities to mitigate pollutant persistence.
Introduction
The occurrence of emerging contaminants (ECs) in wastewater represents a potential risk to the environment, particularly to water bodies that receive treated water and serve as drinking water supply (Morin-Crini et al., 2022). Residual chemicals from personal care products, pharmaceuticals, industrial processes, repellents, and pesticides fall under the category of ECs (Almeida-Naranjo et al., 2023). Wastewater treatment plants (WWTP) remove many ECs through biological biotransformation in the activated sludge (AS) process or by adsorption in primary and secondary sludge (Gogoi et al., 2018; Kennes-Veiga et al., 2022; Núñez-Delgado and Arias-Estévez, 2023). However, some ECs, such as edulcorates, pharmaceuticals, antibacterial agents, and pesticides, have been reported as persistent after treatment in WWTP (Mofijur et al., 2024; Patel et al., 2019; Peña-Guzmán et al., 2019). For example, carbamazepine, acetaminophen (ACE), and triclosan have been detected in WWTP effluents in Argentina, Brazil, and India with concentrations of 13, 50, and 100–4,900 μg/L, respectively (Chaturvedi et al., 2021; Nieto-Juárez et al., 2021). In Mexico, 107.0 μg/L metformin, 67.2 μg/L ACE, 42.8 μg/L caffeine (CAF), 6.6 μg/L sulfamethoxazole, and 0.4–0.5 μg/L of perfluorinated carboxylic acids have been found in WWTP effluents and urban wastewater that is eventually used for crop irrigation (Lesser et al., 2018; Rodríguez-Varela et al., 2021).
Recent studies have reported the removal of ECs using advanced or hybrid treatment technologies, including adsorption systems, membrane nanofiltration, and photocatalytic processes (Kalimuthu et al., 2025; Li et al., 2025; Wang et al., 2025; Zhou et al., 2025). While these approaches can achieve removal efficiencies above 96.9%, they often require additional materials, energy input, or operational complexity compared with conventional biological secondary treatment. Strategies to optimize the removal of ECs within existing wastewater treatment infrastructure comprise a cost-effective and environmentally sustainable option to treat large volumes of wastewater under continuous operation.
For the most concentrated and persistent ECs, identifying microbes with potential to biodegrade these pollutants may lead to strengthening the effectiveness of WWTP as the last barrier for ECs dispersion in water bodies (Seshan et al., 2023). In this sense, bacterial genera such as Pseudomonas have been related to elimination of ACE and carbamazepine in AS (Li et al., 2013; Zhang et al., 2013). Also, the bacterial families Phyllobacteriaceae, Methylophilaceae, Bradyrhizobiaceae, and Pseudomonadaceae and the bacterial strains Bosea sp. and Chelatococcus sp., all isolated from AS, have been pointed out as potential degraders of artificial sweeteners (Kahl et al., 2018; Kleinsteuber et al., 2019).
Microbial isolation through selective pressure is typically performed with high concentrations of ECs to test tolerance and degradation potential of distinctive bacterial genera in terms of the applied organic compound. For instance, Bacillus dretensis strain S1 was enriched and showed maximal growth at 400 mg/L of ACE (Chopra and Kumar, 2020b). In addition, 100–250 mg/L of acesulfame-K (ACK) were utilized to isolate Bosea sp. and Chelatcoccus sp. bacterial strains (Huang et al., 2021; Kahl et al., 2018).
The objectives of this work are to identify the main ECs present in domestic wastewater and assess their chemical oxidation and biological degradation in independent experiments, besides isolating bacterial strains tolerant to the main ECs. The hypothesis of this work is that bacterial genera identified in the AS and isolated through selective pressure are expected to act as candidate degraders of the main ECs found in real wastewater. The main innovation of this work is to provide information about AS bacteria with the potential capability of removing ECs, focusing on improving wastewater treatment.
Materials and Methods
Sampling
AS was sampled from a municipal WWTP (2,250 L/s) in Jalisco, Mexico. Sampling was made in a thickener collecting sludge from secondary settlers after anaerobic, anoxic, and aerobic biological stages. Sludge samples were stored in amber sterile glass containers (1 L) and transported to the laboratory in coolers with ice. The sludge samples were immediately used as inoculum for the experiments, and aliquots were stored at −30°C for DNA extraction.
Biodegradation essays
Biodegradation of ECs was estimated by the dissolved oxygen (DO) consumption following a standard procedure (OECD 301-A, 1992). Briefly, biochemical oxygen demand 300 mL bottles (KIMBLE, Germany) were filled with basal medium (g/L): 8.5 KH2PO4, 21.8 K2HPO4, 33.4 Na2HPO4 2H2O, 0.5 NH4Cl, 36.0 CaCl2, 22.5 MgSO4 7H2O, 0.25 FeCl3 6H2O, with pH adjusted to 7.4 using HCl and NaOH. Medium was saturated with oxygen from bubbling ambient air for 20 min, and the initial DO content was experimentally measured in all cases. Samples of AS collected in December 2023 and May 2024 were used as inoculum with an initial concentration of 0.5 mg chemical oxygen demand (COD)/L. ACK, ACE, CAF, and N,N-diethyl-m-toluamide (DEET) (≥98%, Sigma Aldrich, USA) were utilized as the sole organic carbon source with an initial concentration of 2.35 mg total organic carbon (TOC)/L. This concentration of EC was defined to match the initial DO, which would be just enough for the complete oxidation of the organic carbon. Control experiments were performed to quantify both endogenous respiration (basal medium and inoculum) and chemical oxidation (basal medium and ECs). Every 3 days, the concentration of DO was measured using an YSI MultiLab 4010-3W (YSI, Yellow Springs, OH, USA). All experiments were carried out in triplicate.
The percentage of biodegradation was calculated through the following equation:

The specific consumption rate of ECs between days 0–3, 3–9, and 9–15 was calculated as EC-TOC biodegraded according to equation 1, divided by the initial concentration of inoculum in mg COD/L and accounting for the experiment elapsed time with final units of mg-CEC/mg CODinoculum-day. The TOC in the utilized ECs was calculated from their chemical formula.
Compared with the concentrations used in the biodegradation experiments, concentrations of ECs in the environment might be orders of magnitude lower. However, detecting DO consumption by oxidation of ECs in, for example, micrograms per liter level, would have led to minuscule variations in DO, leading to a great probability of error in the measurements. Despite this, these experiments are presented to evaluate the potential capability of native microbiota in WWTP to degrade ECs.
DNA extraction
Genomic DNA was extracted from 0.3 g of centrifuged sludge (10733 g, 10 min, 4°C) using the FastDNA Spin Kit for Soil (MP Biomedicals, USA) according to the manufacturer's protocol. Extracted DNA was quantified using NanoDrop 2000 (ThermoFisher Scientific, MA, USA) and later purified (Genomic DNA Clean and Concentrator Kit, Zymo Research, CA, USA). The bacterial metagenome was sequenced using MiSeq Illumina at Macrogen (Seoul, Republic of Korea) with universal primers targeting the V4 region of the bacterial 16S rRNA gene with the universal primer set 515F/806R.
Bioinformatic analysis
Metagenomic data for 16S sequencing were analyzed as described. The number of reads ranged from 129,766 to 165,326. Data were quality trimmed using trimmomatic v0.39 (Q30 seqs comprised at least 76% of reads), then paired using flash 1.2.11. After dereplication, chimeras were eliminated, sequences were clustered using 97% identity, and subsequently singletons were eliminated using vsearch 2.14.2 and annotated against SILVA database v 138.1 pretrained using qiime2 for 16S. After this analysis, only 1,761 operational taxonomic units (OTUs) were obtained, and relative abundance was normalized for each sample. Table of OTUs was used for diversity index by means of Hill numbers as described previously (Chao et al., 2014). For such alpha diversity indexes, 1,000 bootstrap resampling with 95% subsamples was used to produce violin plots, which were graphed using MATLAB 2024b.
Isolation of DEET and ACE-resistant bacterial strains
Serial dilutions of AS were performed to inoculate 100 mL-aliquots on Luria Bertani (LB) agar medium (MCD Labs, Mexico) fortified with 100 mg/L of either DEET or ACE, then incubated for 24 h at 30°C under stationary inverted conditions. Distinctive colonies were isolated based on morphology (size, shape, texture, color) and subcultured individually by four rounds of streaking on fortified LB plates. The isolated organisms were subsequently identified by matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS, Bruker Daltonik Biotyper, Bremen, Germany) using the MALDI-Biotyper reference database. The obtained single colony was transferred to fresh LB liquid media containing 100 mg/L of DEET or ACE to confirm resistance. Bacterial biomass was recovered after centrifuging the liquid cultures (10733 g, 10 min, 4°C). DNA extraction was carried out with FastDNA Spin Kit for Soil (MP Biomedicals, USA) as described in the “Bioinformatic analysis” section. Genome sequencing on the Illumina platform, next-generation sequencing, was performed on the provided DNA samples (Psomagen, USA). The 16S rRNA gene sequence of each strain was amplified by PCR using the universal primer set 8F/1492R. The obtained sequences were used as a query for BLASTn against the NCBI nr database. The sequences were clustered using MEGA11.0 to construct the phylogenetic tree using the neighbor-joining method.
Chromatographical analysis
Liquid samples were extracted and concentrated by solid phase extraction using cartridges (Oasis HLB 20 cc Vac Cartridges, 5 g sorbent, Waters, USA) following the methodology reported previously (Wu et al., 2023) . Briefly, the cartridges were conditioned by rinsing the contents with 3 mL of methanol followed by 3 mL of Milli-Q acidified water (pH 2.0) and dried in a vacuum for 15 min. Wastewater samples (500 mL) or biodegradation essays aqueous suspension (300 mL) were filtered through fiberglass filters (11 μm pore size, circles 110 mm Æ, Whatman, UK) and pH adjusted to 2.0 with concentrated sulfuric acid. The samples were then passed through the conditioned cartridges. The analytes were eluted from the cartridges with 12 mL of methanol and 6 mL of acetone, and these extracts were concentrated to dryness under a gentle stream of nitrogen gas (Chromatographic grade, INFRA, Mexico) and finally resuspended in 1.0 mL of 0.1% formic acid in methanol (v/v) for analysis.
Separation of ACK, ACE, CAF, and DEET was performed using Ultra Performance Liquid Chromatography (UPLC) equipped with an Acquity I-class column (CSH C18, Waters, USA) coupled to a double quadrupole MS via heated electrospray ion source (Xevo TQ-S Mass Spectrometry, Waters, USA). The UPLC flow rate was 0.1 mL/min with a column temperature of 40°C and run time of 20 min. Detailed information on the instrument method, quality assurance, and method performance can be found elsewhere (Ibarra-Rodríguez et al., 2023) .
UPLC-grade solvents methanol, acetonitrile, isopropanol, formic acid, and acetone were acquired from Merck (Germany), except sulfuric acid (JT Baker, Phillipsburg, NJ, USA). Ultrapure water was used for all experiments (Milli-Q, Millipore, USA). Individual stock solutions of standards were prepared in methanol and stored in amber glass bottles at −30°C until used. Quantification limits (0.02–2.0 μg/L) were verified through concentration curves (μg/L) of 10, 50, 100, 200, 400, 500, 800, and 1,000, prepared with analytical grade standards (≥98%, Sigma-Aldrich, USA).
Statistical analysis
All experiments were performed in triplicate, and results are presented as mean values ± standard deviation. To evaluate differences between biological and chemical oxidation, individual one-way analysis of variance (ANOVA) tests were performed to evaluate differences between days of the experiment. Differences were considered significant when p < 0.05. Statistical analyses were conducted using Statgraphics Centurion (version XVI, Statgraphics Technologies, Inc., USA).
Results and Discussion
ECs in wastewater
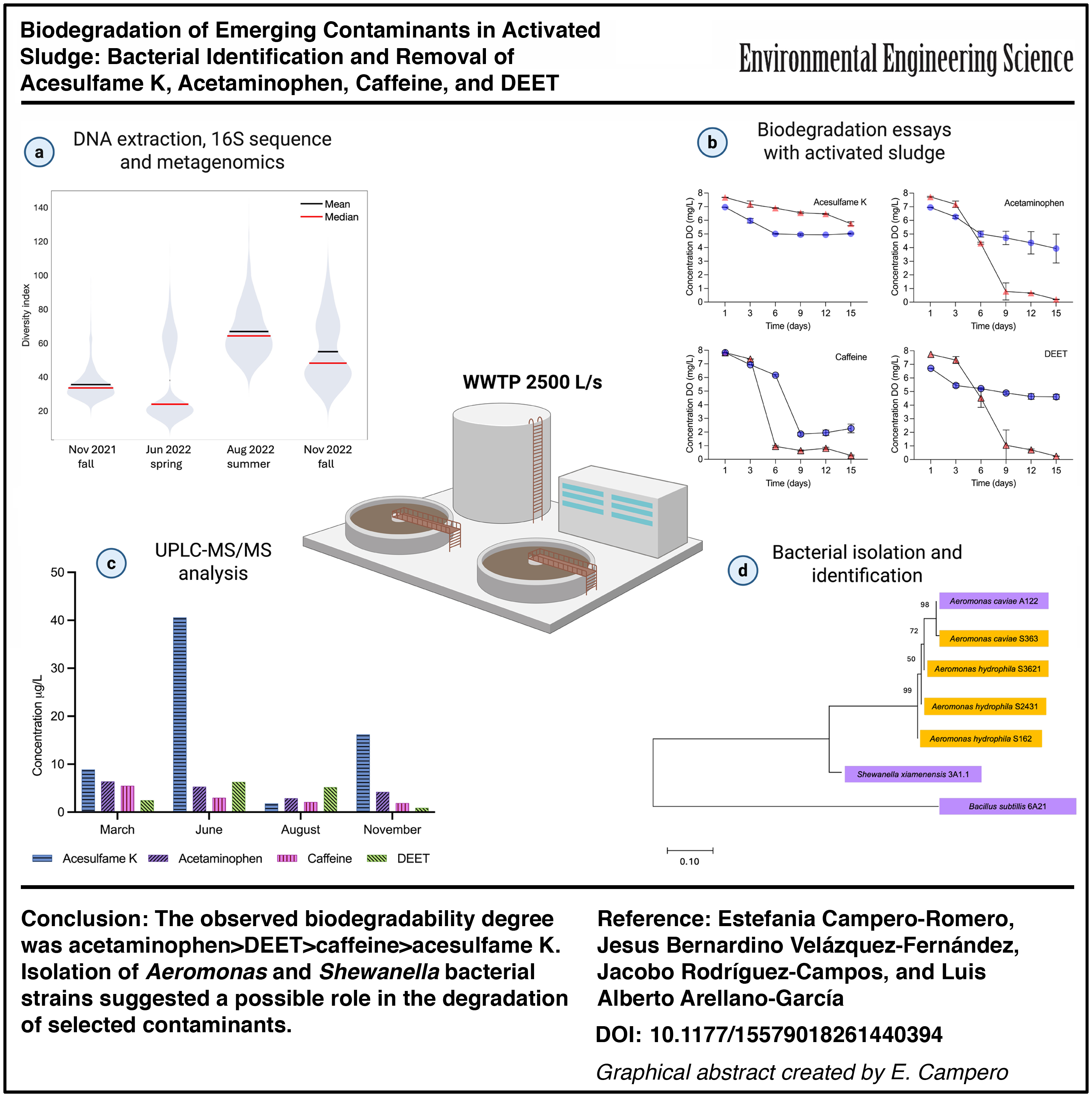
Average values of ECs detected with the highest concentration in the WWTP influent in March, June, August, and November 2022 were ACK 16.9 ± 16.9 μg/L, ACE 4.7 ± 4.4 μg/L, CAF 3.1 ± 1.6 μg/L, and DEET 3.7 ± 2.5 μg/L (Supplementary Table S1). Seasonal changes were observed in the rainy season (August 2022), where concentrations of ACK 1.8 μg/L, ACE 2.9 μg/L, CAF 2.1 μg/L, and DEET 5.2 μg/L were in general lower compared with the rest of seasonal samplings (Fig. 1). In addition, a peak concentration of ACE of 6.4 μg/L observed at the end of winter (March 2022) may be linked to the seasonal consumption of analgesics.

Concentration of the four most concentrated emerging contaminants in the influent of the WWTP in four different seasonal samplings. WWTP, wastewater treatment plant.
Other authors have reported WWTP influent concentrations of ACK 10–78 μg/L (Qiao et al., 2024; Tran et al., 2015), ACE 38–292 μg/L (Peña-Guzmán et al., 2019; Verlicchi et al., 2012), and DEET 15 μg/L (Marques Dos Santos et al., 2019). In Mexico, maximum concentrations of ACE 67.2 μg/L and CAF 42.8 μg/L have been found in urban wastewater (Vázquez-Tapia et al., 2022).
ACK molecule is resistant to enzymes present in the human body, and after ingestion, it is excreted without undergoing any chemical change (Danner et al., 2023). However, it has been reported that ACK can undergo biological degradation in WWTP under oxic or anoxic conditions (Castronovo et al., 2017). CAF is rapidly absorbed into the gastrointestinal system, and around 95% is oxidized to paraxanthine, theobromine, and theophylline (Reddy et al., 2024). However, ∼1% to 5% of ingested CAF is not metabolized in the human body and is excreted in the urine, reaching the sewage system (Marasco Júnior et al., 2019). ACK, due to its recalcitrancy and CAF due to the volume of consumption, have been proposed as tracers of wastewater anthropogenic contamination hot spots (Nödler et al., 2016).
In Mexico, analgesics are commonly accessed as over-the-counter medications, and self-medication is a prevalent practice throughout Latin America. This trend is primarily driven by regulatory shortcomings that allow the sale of medications without the necessity of a medical prescription (Santa-Ana-Tellez et al., 2016). Despite the widespread availability of ACE in Mexico, data on the consumption are quite limited (García Aguirre et al., 2019). Although ACE is one of the most widely used medicines worldwide, pharmaceutical consumption patterns differ by location, region, or country, such as country income; these disparities significantly affect its environmental occurrence (Patel et al., 2019).
DEET, as the active ingredient in insect repellents, has been detected in wastewater, surface water, and groundwater, mainly from consumer use, with higher concentrations during the summer months, which correspond to the season of mosquito prevalence. In an urban WWTP, CAF and DEET were the two predominant ECs detected at levels of micrograms per liter with a frequency above 50% in monthly samplings on seven sites for 9 months (Hattaway et al., 2024).
Biodegradation experiments
The rate of biological DO consumption, with AS as inoculum, was similar for ACE, CAF, and DEET, which significantly consumed more DO compared with the chemical oxidation control (Fig. 2, Table 1). In general, the highest consumption of DO and consequently the estimated consumption rate of pollutants occurred between days 3 and 9, suggesting this was the most active phase for biodegradation (Table 1).

Concentration of dissolved oxygen during biological (red triangle) and chemical (blue circle) experiments with acesulfame K, acetaminophen, caffeine, and DEET over 15 days at 25°C. The biological oxygen consumption was corrected by deducting the endogenous respiration. Each marker corresponds to the average of three independent replicates, while the error bars indicate the standard deviation. DEET, N, N-diethyl-m-toluamide.
Mean Dissolved Oxygen Consumption (±SD) for the Four Selected Emerging Contaminants During the Biodegradation Experiment
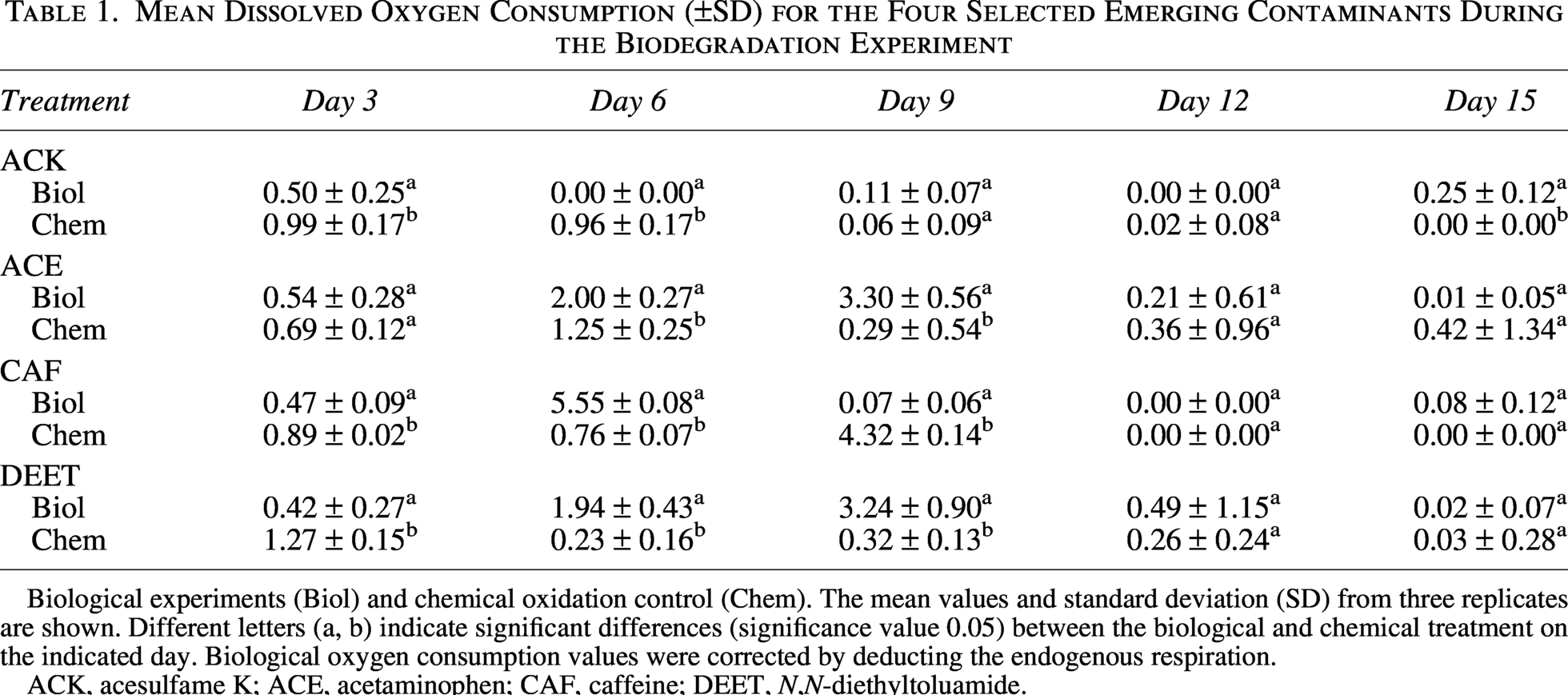
Biological experiments (Biol) and chemical oxidation control (Chem). The mean values and standard deviation (SD) from three replicates are shown. Different letters (a, b) indicate significant differences (significance value 0.05) between the biological and chemical treatment on the indicated day. Biological oxygen consumption values were corrected by deducting the endogenous respiration.
ACK, acesulfame K; ACE, acetaminophen; CAF, caffeine; DEET, N,N-diethyltoluamide.
In the biological oxidation of ACE, 44% and 90% of DO were consumed by days 6 and 9, suggesting that ACE removal occurred primarily through biological oxidation because the chemical oxygen exhaustion was significantly lower (Table 1). The maximum ACE consumption rate based on oxygen consumption was observed between days 3 and 9, with 0.66 ± 0.48 mg CACE/mg COD-d.
For CAF, the consumption of DO was faster compared with ACE and DEET, with 94% of the DO depleted after 6 days. The biological DO consumption was significantly higher than the chemical oxygen depletion except for days 1–3. The highest estimated consumption rate for CAF also occurred between days 3 and 9, with values of 0.70 ± 0.04 mg CCAF/mg COD-d.
In the case of DEET, 42% of DO was consumed on days 6 and 9. Biological oxidation consumed significantly more DO compared with chemical oxidation after day 6 (Table 1), suggesting a predominant role of biological transformation over chemical degradation. The maximum calculated consumption rate of DEET of 0.65 ± 0.87 mg CDEET/mg COD-d was observed between days 3 and 9.
Notably, biological respiration in the presence of ACK consumed less DO compared with the chemical oxidation control, where only 25% of the initial DO was consumed biologically by day 15, compared with 28% consumed chemically over the same period. Specifically, the maximum consumption rate of ACK occurred between days 0 and 3, with an estimated consumption rate of 0.13 ± 0.18 mg CACK/mg-day.
According to current literature, the order of biodegradability among the selected ECs is ACE > DEET > CAF > ACK, with ACE being the most readily biodegradable and ACK the most persistent (Hattaway et al., 2024; Manyepa et al., 2024; Qiao et al., 2024). This biodegradability also coincides with the biological degradation constant (Kbiol) values and the octanol–water partitioning (log Kow), where low values signify low absorption in solid compartments that also limit biodegradation (Yildirim and Köse, 2025) (Table 2). Here, the removal of parent compounds in the biodegradation experiments confirms the tendency shown in the literature, with 100% of ACE, 99% of DEET, 97% of CAF, and 90% of ACK removed, based on final concentrations of 0.0 mg ACE/L, 25.0 mg DEET/L, 73.5 mg CAF/L, and 6.1 mg ACK/L experimentally determined.
Characteristics of Selected Emerging Contaminants
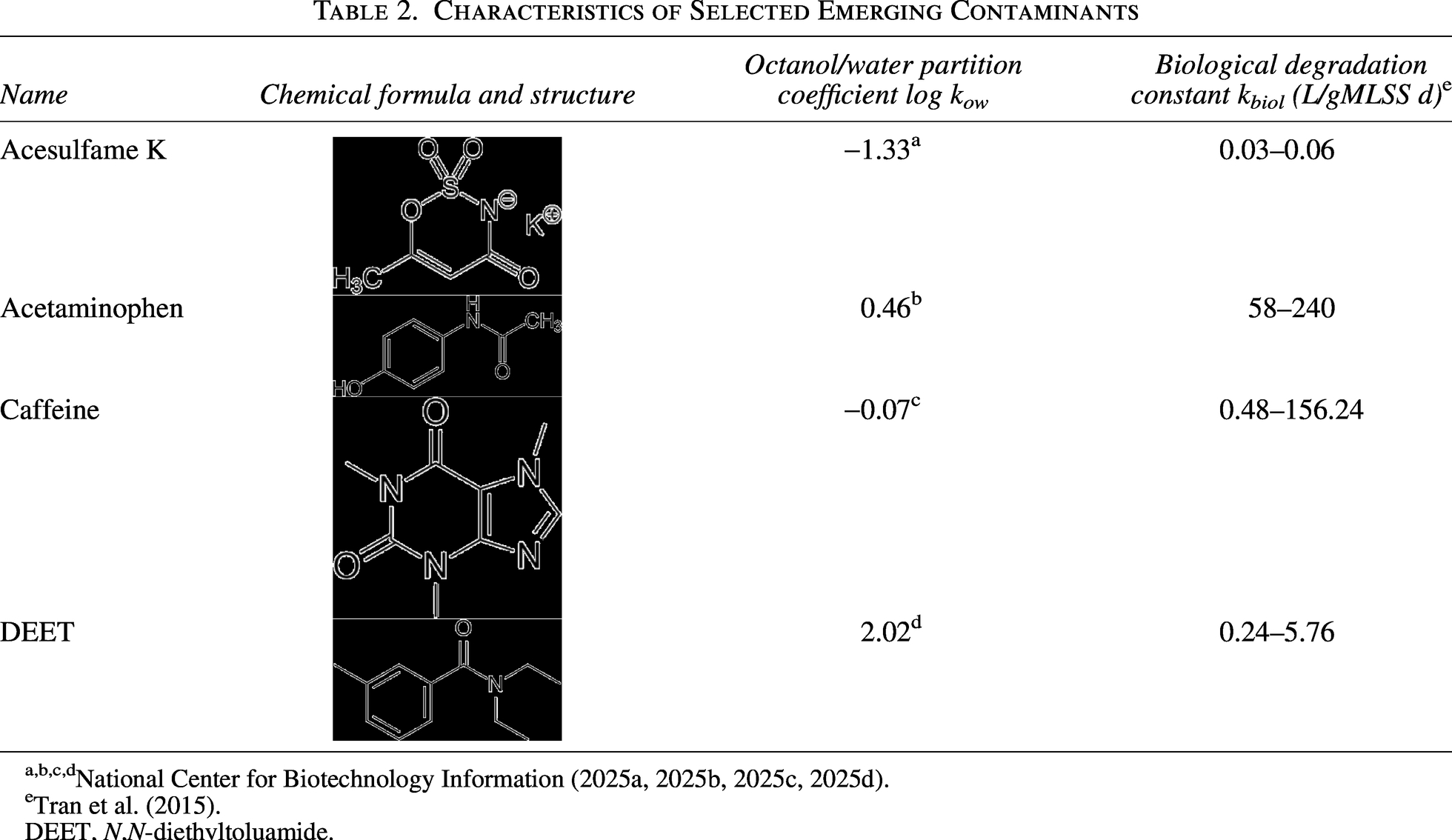
Tran et al. (2015).
DEET, N,N-diethyltoluamide.
In the case of ACK, concentration and acclimation of biomass are considered the main reasons for differences in the removal between the cited articles and this study. For example, removal up to 94% of ACK was reported in batch experiments seeded with AS after 27 h. However, the concentration of biomass is not explicitly declared (Castronovo et al., 2017). In WWTP around the world, 20–90% of ACK removal have been reported in Canada (influent: 15.6–28.4 μg/L; effluent: 0.2–21.0 μg/L), 7–50% in China (influent: 0.8–11.8 μg/L; effluent: 0.6–10.5 μg/L), and 49–58% in Vietnam (influent: 10.0–24.0 μg/L; effluent: 5.1–10.0 μg/L) (Li et al., 2018; Nguyen et al., 2018; Qiao et al., 2024; Van Stempvoort et al., 2020). Sludge concentrations of 2,000–6,000 mg of volatile suspended solids per liter are typical in the AS process (Metcalf and Eddy Inc., 2002). Meanwhile, up to 180 days have been used to satisfactorily obtain acclimated ACK-degrading bacterial strains Bosea sp. and Chelatococcus sp. (Huang et al., 2021; Kleinsteuber et al., 2019).
In this study, 0.5 mg COD/L of nonacclimated AS was utilized to measure the instantaneous consumption of oxygen in biomass at its actual state in the sourced WWTP. Cometabolism of ECs was minimized by the low concentration of AS; besides, exogenous organic carbon or nitrogen source essays were not supplemented. Also, to explain the lower consumption of oxygen in the biological experiment with ACK compared with the chemical control (Fig. 2, Table 1), it is hypothesized that this artificial sweetener inhibited the biological consumption of oxygen. For instance, a recent report showed that for two bacterial strains, the half maximal inhibitory concentration for ACK was between 22 and 26 mg/L, while a reduced bacterial activity was observed starting from 8.9 mg/L (De Dios et al., 2023).
It is known that cometabolism is a typical pathway for the removal of recalcitrant compounds in wastewater treatment, where the supply of organic carbon in the AS process promotes the biotransformation of a variety of compounds, including ECs (Kennes-Veiga et al., 2022). Evidence suggests that nonspecific enzymes such as ammonia monooxygenase intervene in the cometabolic degradation of ECs, contributing significantly to the removal of micropollutants (Li et al., 2024; Yu et al., 2018). Given that nitrification is present in most full-scale WWTP, this pathway represents a key mechanism for the degradation of ECs in these systems (Bhatt et al., 2022).
Biodegradation efficiency and kinetics vary substantially depending on the compound and the degree of microbial adaptation. In a laboratory scale, Achromobacter, Cupriavidus, and Pseudomonas were enriched from activated sludge and achieved complete removal of 100 mg/L of CAF, ACE, and ibuprofen within 1 week (Suleiman et al., 2023). Likewise, microalgal-bacterial consortia dominated by Chlorella spp., and Scenedesmus spp., achieved removal above 85% of sulfamethoxazole and 65–70% for metoprolol, fluoxetine and diclofenac and 30–40% for carbamazepine within 6 days of hydraulic retention time (Mantovani et al., 2024). In addition, sulfate and nitrate-reducing consortia can remove 80–84% of compounds such as ciprofloxacin and 17β-estradiol after 8 days (Martins et al., 2018).
Metagenomic
Activated sludge is the globally dominant system for wastewater treatment, and its effectiveness and treatment quality rely on the balance of bacterial species, which can be influenced by not only wastewater temperature and composition but also treatment process design, aeration time, and chemicals added (Peces et al., 2022).
Seasonal variation in the relative abundance of the dominant bacterial genera in AS studied is shown in Table 3. In November 2021 (fall), Aeromonas was the most abundant bacterial genus. The genus Aeromonas commonly occurs in sewage, can grow both aerobically and anaerobically, and utilizes a wide range of carbohydrate sources besides its ecological role in nitrogen cycling (Taabodi et al., 2020; Vuković Domanovac et al., 2019). Aeromonas spp. has been shown to degrade organophosphate insecticide chlorpyrifos and the synthetic estrogen 17α-ethinylestradiol (Balarabe et al., 2025; Palma and Costa, 2024). Gene sequences encoding enzymes sulfatases and amidases have been identified in Aeromonas (Seshadri et al., 2006) (Uniprot database A0KLD1, A0KPV3), suggesting the capability of ECs biotransformation, as these enzymes have proven to be involved in the biodegradation of ACK by bacteria (Bonatelli et al., 2023). Moreover, the next most abundant genera Macellibacteroides, Bacteroides, Anaerosinus, and Propionivibrio are typically involved in the hydrolysis and acidogenesis phases of organic matter degradation in anaerobic conditions, which may indirectly support ECs degradation by providing intermediary substrates or redox conditions (Daims et al., 2006; Mei et al., 2020).
Top Five Bacterial Genera in Activated Sludge Across Four Different Seasons. Values Indicate Relative Abundance Percentage (%) at the Genus Level

The microbial dominant genera shifted in June 2022 (spring), with OLB8 (2.1%) being the more abundant. This genus has been found in WWTP worldwide, related to its ability to breakdown organic compounds and participate in nitrogen and phosphorus removal (Kondrotaite et al., 2022; Peces et al., 2022). Ottowia, the second most abundant bacterial genus, includes denitrifying bacteria intervening in the nitrogen cycle (Spring et al., 2004). The detection of the halotolerant Halomonas (1.1%) in June 2022 suggests an increase in salinity or osmotic stress within the system (Arahal et al., 2002). Meanwhile, the detection of Novosphingobium (0.7%) suggests an increase in aromatic compound degradation capacity, as this genus is known for its ability to metabolize xenobiotic compounds (Kertesz and Kawasaki, 2010). A comparative biodegradation study of 10 ECs found that Novosphingobium was among the genera statistically associated with higher overall removal efficiency across treatments (Coll et al., 2020).
In August 2022 (summer), the supremacy of genera Bdellovibrio and OM27 from the Bdellovibrionaceae family, capable of infecting and lysing other bacteria (Rotem et al., 2014), was detected, followed by genera Denitratisoma, Nitrospira, and Dechloromonas, which are known to participate in the nitrogen cycle as documented previously (Petriglieri et al., 2021). The presence of Dechloromonas is particularly noteworthy; recent genomic studies have shown that strains within this genus encode key detoxification enzymes, including glutathione S-transferases and 4-oxalocrotonate tautomerases, both associated with the degradation of aromatic and xenobiotic compounds (Zhang et al., 2021).
In November 2022 (fall), Tetrasphera (2.8%) and Ottowia (1.6%) were the dominant genera. Tetrasphera is known for its role in enhanced biological phosphorus removal and glycogen storage metabolism, suggesting potential seasonal optimization of phosphorus removal processes (Kristiansen et al., 2013; Otieno et al., 2022). Moreover, the genus Tetrasphera has been identified among the growing genera through a long-term (3–5 years) sampling in WWTP (Peces et al., 2022). The persistence of Ottowia reinforces the importance of denitrification in the system, showing that this genus is fundamental for nutrient removal in wastewater treatment.
Despite the genera Macellibacteroides, Bacteroides, Anaerosinus, Propionivibrio, OLB8, Ottowia, Halomonas, Bdellovibrio, OM27, Denitratisoma, Nitrospira, and Tetrasphera cannot be directly linked to ECs degradation, they seem to play supportive roles in maintaining functional stability, nutrient cycling, and redox balance, thereby facilitating co-occurrent biodegradation processes.
Analysis of the four sequencing data sets showed similar richness, evenness, and dominance based on results of Hill numbers q = 0 (Supplementary Fig. S1), q = 1 (Supplementary Fig. S2), and q = 2 (Supplementary Fig. S3), except for the June 2022 sample, which Hill numbers were all lower. Despite two subpopulations being depicted in the June 2022 sample, mean, and median are both in the dominant population. The sequencing of 16S rRNA in DNA extracted from AS in the studied WWTP, along with the subsequent characterization of microbial bacterial communities, has provided insights into their taxonomic composition and relative abundance. This analysis has also highlighted the potential role in the degradation of ECs, specifically of identified bacterial genera capable of transforming organic compounds.
Bacterial isolates
Seven bacterial strains tolerant to 100 mg/L of either DEET or ACE were isolated and identified (Fig. 3, Supplementary Table S3). Here, genus and species-level identification by MALDI-TOF MS was confirmed by 16S rRNA gene sequence, except for two bacterial strains initially classified as Acinetobacter towneri and Aeromonas veronii, which were both reclassified as Aeromonas hydrophila.
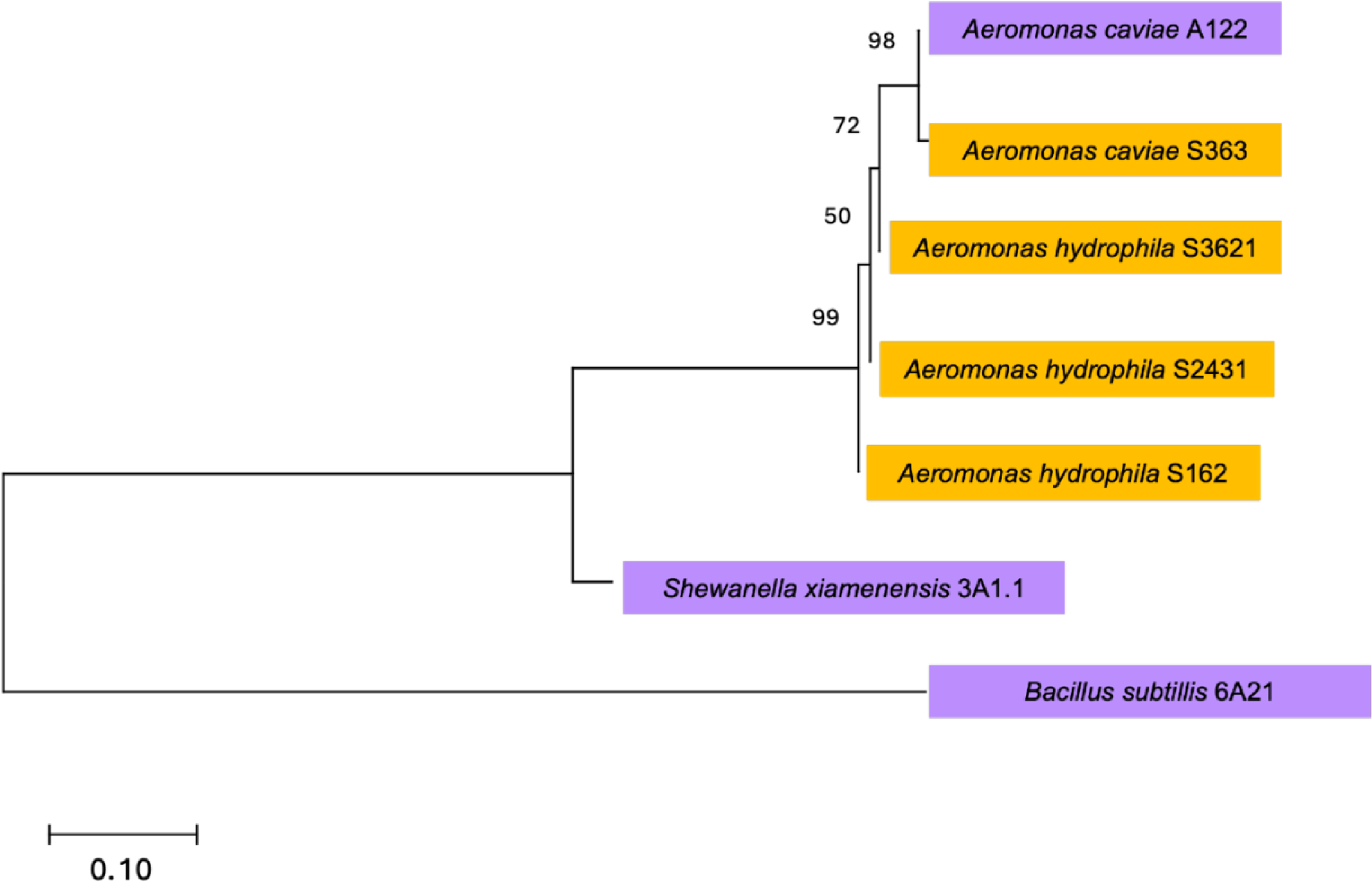
Phylogenetic tree of 16S rRNA genes of bacteria resistant to acetaminophen (purple) and DEET (yellow) based on the neighbor-joining method. Numbers at the nodes represent bootstrap values (%) from 1,000 replicates, indicating branch support. The scale bar (0.10) shows the number of nucleotide substitutions per site. DEET, N,N-diethyl-m-toluamide.
By aligning the 16S rRNA gene sequences against the NCBI nr database through BLASTn, the seven strains were classified into three main genera, including the gram-negative Aeromonas (5), gram-positive Bacillus (1), and gram-negative Shewanella (1). The dominance of Aeromonas species was consistent with the results from the metagenomic analysis presented in the “Metagenomic” section. Phylogenetic analysis of the isolates using the neighbor-joining method (Fig. 3) revealed distinct evolutionary clusters. Aeromonas caviae A122 and Aeromonas caviae S363 strains clustered together with 98% bootstrap support, indicating a close relationship. Similarly, the grouping of Aeromonas spp. (hydrophila S3621, S2431, and S162) suggests a shared evolutionary adaptation.
The identification of Aeromonas species in isolates tolerant to DEET and the positive consumption of DO in biological experiments suggest that members of this genus may possess mechanisms relevant for DEET transformation. This hypothesis is supported by the Aeromonas hydrophila oxidoreductive and hydrolytic enzymes, such as laccase, involved in breaking down aromatic compounds (Srinivasan and Sadasivam, 2021). In contrast, ACE-tolerant isolates identified as bacteria were more phylogenetically diverse. Among these, Shewanella xiamenensis features enzymes catalase, laccase, and manganese peroxidase (Ng et al., 2015). Degradation of ACE by immobilized laccase of fungal origin has been demonstrated for a concentration of 18 mg ACE/L (Sotelo et al., 2022). Peroxidase obtained from bacterial cultures was successfully applied for the degradation of up to 300 mg ACE/L (Baratpour and Moussavi, 2018). In the same genus, Shewanella putrefaciens performed degradation of organic compounds in aerobic conditions, where enzyme dioxygenase might play a key role in the cleavage of benzene rings (Li et al., 2024).
In the case of Bacillus subtillis, it is a well-known environmental bacterium and is often associated with bioremediation due to its metabolic versatility. Gene annotation analysis in this species revealed the capability for xenobiotic degradation, involved in the breakdown of polyaromatic compounds (Saikat et al., 2024).
Commonly, concentrations between 100 and 400 mg/L have been utilized to successfully isolate bacterial strains in presence of ECs, where degrading ability has been demonstrated by analyzing biodegradation transformation products (Huang et al., 2021; Kahl et al., 2018). Bacillus species, including B. dretensis, B. licheniformis, and B. pumilus, have been reported to degrade ACE through oxidative enzymatic pathways involving oxidases, peroxidases, and amidases (Chopra and Kumar, 2024, 2023, 2020a, 2020b), supporting the plausibility that Bacillus intervened in the ACE degradation observed here.
Despite the results presented in this study confirming the presence and activity of specific bacteria that may be key for degradation of ECs, functional assays and metabolite analyses are required to confirm the specific role of the isolated strains in either DEET or ACE biodegradation. Nonetheless, the analysis presented here enhances our understanding of how different microbial communities contribute to the biodegradation of ECs and provides valuable insights for the development of microorganism-based water treatment technologies.
Conclusions
This study assessed the biodegradation of the emerging contaminants ACK, ACE, CAF, and DEET using activated sludge as inoculum. The assayed contaminants were selected based on the most concentrated compounds measured in municipal wastewater. ACK exhibited strong recalcitrance, while the other pollutants showed varying degrees of biodegradability (ACE > DEET > CAF > ACK) under laboratory conditions. Metagenomic analysis of the activated sludge used as inoculum identified Aeromonas, Macellibacteroides, Ottowia, Tetrasphera, and Bdellovibrio among the most abundant bacterial genera. In addition, seven bacterial strains tolerant to DEET or ACE were isolated through sequential plate culture, of which five belong to the Aeromonas genus, suggesting a possible role in contaminant transformation. These results underscore the environmental persistence of certain contaminants and highlight the biotechnological potential of native bacterial genera such as Aeromonas and Shewanella for bioaugmentation strategies in WWTP. Future work will focus on enzyme activity tests or biodegradation assays using the bacterial strains isolated here to discern their practical value in enhancing contaminant removal in engineered systems.
Authors’ Contributions
E.C.-R.: Writing—original draft, review and editing, investigation, methodology, data curation, formal analysis, and conceptualization. J.B.V.-F.: Software, data curation, formal analysis, and writing—original draft, review and editing. J.R.-C.: Methodology, investigation, and writing—original draft, review and editing. L.A.A.-G.: Writing—original draft, review and editing, conceptualization, project administration, and supervision.
Footnotes
Acknowledgments
The authors would like to thank SECIHTI Mexico for funding this work through the grant Secretaría de Ciencia Humanidades Tecnología e Innovación. Ciencia de Frontera 2019 Grant No. 102967 and for Estefania Campero PhD scholarship No. 815481. The authors acknowledge the technical assistance of Dra. Diana Ibarra-Rodríguez in sampling and sample processing.
Author Disclosure Statement
No competing financial interests exist.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.