
Abstract
Objectives
The specific role of metabolites in mediating the relationship between gut microbiota (GM) and prostate cancer (PCa) remains unclear. In this study, we utilized Mendelian randomization (MR) analyses to investigate the causal relationship between 430 GM traits, 1,400 plasma metabolites, and PCa.
Methods
Inverse variance weighted (IVW) method is the primary analytical approach, with supplementary validation using MR-Egger, weighted median, and weighted mode methods. Mediation analysis was performed to assess the contribution of metabolites to the GM-PCa association.
Results
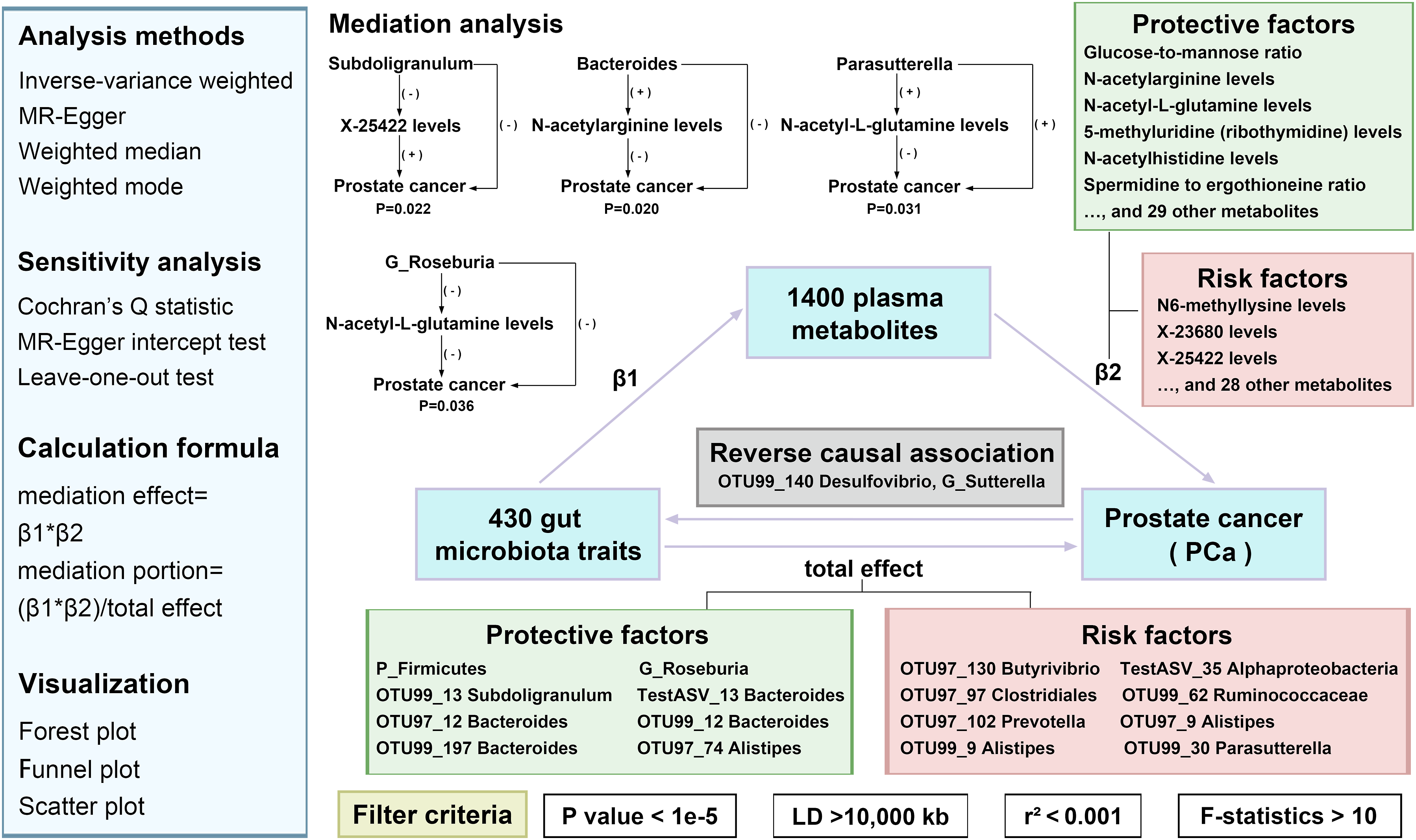
IVW analysis identified 18 GM traits and 66 plasma metabolites showing a causal association with PCa. For instance, P_Firmicutes exhibited a causally protective effect against PCa (IVW: OR=0.937, P=0.008), while Parasutterella showed a causally risk-increasing effect (IVW: OR=1.029, P=0.039). Notably, four metabolites mediated the causal effects of GM on PCa: Subdoligranulum via unidentified metabolite X-25422 (20.785% mediation proportion, P=0.022), Bacteroides via N-acetylarginine (6.706%, P=0.020), and Parasutterella and G_Roseburia via N-acetyl-L-glutamine (-9.382%, P=0.031 and -9.094%, P=0.036, respectively). No significant heterogeneity or horizontal pleiotropy was detected.
Conclusion
These findings offer new insights into the metabolite-mediated pathways linking GM to PCa, which may serve as novel potential biomarkers and therapeutic targets for precision therapies.
1. Introduction
In 2022, prostate cancer (PCa) accounted for an estimated 1.467 million new cases and 397,000 deaths globally, ranking as the second most common malignancy and the fifth leading cause of cancer-related mortality in men. 1 In the United States, PCa represents the most common non-cutaneous cancer among men, with an estimated 299,010 new cases and 35,250 deaths projected for 2024 2 . Cancer remains one of the most lethal diseases in contemporary society, with modernization-driven factors, including lifestyle transformations and environmental degradation, have substantially amplified carcinogenic risks. 3 To address this escalating challenge, innovative pharmaceutical strategies have emerged at the forefront of oncological research. Notably, novel multidentate ligand metal complexes, particularly L-glutamic acid-based Cu (II) and Ru (III) coordination compounds, demonstrate potent cytotoxicity against diverse cancer cell lines. 4 Concurrently, thalidomide and its derivatives have experienced a remarkable renaissance following historical setbacks, now serving as critical therapeutic agents for metastatic PCa and other malignancies. 5 The development of intelligent nanocarriers has substantially enhanced drug delivery efficiency and therapeutic precision. 6 Furthermore, the biological applications of metal complexes extend beyond oncology, with pyrazoline-based Cu (II) and Ni (II) complexes demonstrating promising antifungal efficacy. 7
Despite substantial progress in imaging modalities, prostate-specific antigen screening, nanotechnology-enabled drug development, and multimodal therapeutic strategies, the prognoses for advanced PCa remain suboptimal.6,8 Most metastatic cases inevitably progress to castration-resistant PCa, an aggressive phenotype with limited therapeutic options and a 5-year survival of 30%. 9 Although several novel agents have demonstrated promising efficacy in in vitro studies of cervical cancer, hepatocellular carcinoma and other malignancies, the rising prevalence of malignancies has imposed a substantial socioeconomic burden globally.3-5 This underscores the need to elucidate novel pathogenic mechanisms, risk factors, and therapeutic targets.
The gut microbiota (GM), a dynamic and diverse microbial community in the gastrointestinal tract, has emerged as a key regulator of host metabolism and immunity.10,11 Dysbiosis has been linked to PCa initiation and progression, 12 with taxa such as Streptococcus 13 and Akkermansia 14 enriched in patients. These microbes may promote tumorigenesis by impairing intestinal barrier function, 15 modulating inflammatory mediators, 16 and reshaping the immune microenvironment, thereby influencing immunotherapy responses. 17
Metabolic reprogramming is a hallmark of cancer, and plasma metabolites reflect alterations in pathways including glycolysis, fatty acid oxidation, and amino acid turnover. 18 Changes in circulating myo-inositol, phosphate, and palmitic acid in lung cancer highlight disruptions in glucose and lipid metabolism. 19 In PCa, branched-chain amino acids, 20 sphingosine-1-phosphate, 21 and urinary glutamate and pseudouridine 22 have been proposed as biomarkers for diagnosis and prognosis. Deciphering the GM-metabolite-PCa axis could inform early detection strategies and precision therapeutics.
However, observational studies are often confounded by environmental and clinical variables. Mendelian randomization (MR), which uses genetic variants as instrumental variables (IVs), leverages their random allocation to minimize confounding and strengthen causal inference. 23 For example, Zhang et al used two-sample MR to link fatty acid profiles causally to dilated cardiomyopathy risk. 24 The causal interplay and mediating mechanisms among GM, plasma metabolites, and PCa remain largely unexplored. To address this gap, we integrate large-scale genome-wide association study (GWAS) datasets25-27 and represent the first use of mediation MR to quantify the mediating role of metabolites in the GM-PCa causal pathway. This offers novel causal evidence for microbiota-metabolite-tumor interactions in PCa.
2. Materials and Methods
2.1. Data Sources
Detailed Information of the Three Datasets

2.2. Selecting Genetic Instruments
We selected single nucleotide polymorphisms (SNPs) as IVs based on the three core assumptions 28 : 1 strong association with the exposure, 2 no association with potential confounders, and 3 no direct effect on the outcome independent of the exposure. According to previous studies, 29 SNPs were selected using a significance threshold of p < 1×10-5. Linkage disequilibrium clumping was performed using the 1000 Genomes Project Phase 3 as the reference panel, with a clumping window of 10,000 kb and a linkage disequilibrium r2 threshold of 0.001. Data harmonization was implemented via the TwoSampleMR (v0.6.20) and ieugwasr (v1.1.0) packages. Strand alignment was inferred from allele frequency information using default settings, and palindromic SNPs with a minor allele frequency close to 0.5 were excluded to avoid alignment ambiguity. F-statistics were calculated using the formula F = Beta2/SE2, and only SNPs with F > 10 were retained to ensure instrument strength.
2.3. MR Analysis
2.3.1. Exploring the Causal Relationships Between GM, Plasma Metabolites, and PCa
Initially, we employed two-sample MR to assess the causal relationships between GM, plasma metabolites, and PCa, utilizing methods including inverse variance weighted (IVW), MR Egger, weighted median, and weighted mode approaches. IVW is considered the most precise and robust method. 29 The MR Egger method incorporates an intercept term to detect potential pleiotropy. 30 The weighted median method, as a supplementary approach, provides consistent estimates when at least 50% of the IVs are valid. 31 Our analysis was conducted using R version 4.3.1 with the “TwoSampleMR” and “ieugwasr” packages, with statistical significance set at p < 0.05. We applied the Benjamini-Hochberg method for false discovery rate (FDR) correction. Consistent with previous studies, 32 with an FDR-adjusted P-value (q value) < 0.2 deemed suggestive of a potential causal association and the q value < 0.05 considered indicative of a significant causal relationship. The primary evaluation metrics included odds ratios (ORs) and their corresponding 95% confidence intervals (CIs).
All reported β coefficients and ORs for GM and plasma metabolites correspond to the effect per 1-standard deviation (SD) increase in the exposure. MR effect estimates for microbial abundance traits reflect the change in the outcome per 1-SD increment in microbial abundance. For microbial prevalence traits, MR estimates denote the change in the outcome per unit increase in genetic liability to microbial presence. MR effect estimates for metabolites represent the change in the outcome per 1-SD increment in log-transformed metabolite levels.
2.3.2. Excluding Potential Reverse Causality Between GM and PCa
We performed a reverse MR analysis by treating PCa as the exposure and GM as the outcome. This approach ensured that the identified causal pathways followed the hypothesized direction from GM to plasma metabolites and subsequently to PCa.
2.3.3. Mediation Analysis
Additionally, we analyzed the causal relationships between candidate GM traits and plasma metabolites to investigate the potential mediating role of plasma metabolites in the GM-PCa causal pathway. The total effect was decomposed into a direct effect (not mediated) and an indirect effect (mediated via plasma metabolites). The indirect effect was then divided into the effect of GM on plasma metabolites (Beta1) and plasma metabolites on PCa (Beta2). The indirect effect was calculated using the formula: indirect effect = Beta1 × Beta 33 , with the ratio of indirect effect to the total effect defined as the mediation proportion.
2.3.4. Sensitivity Analysis
Heterogeneity was assessed using Cochran’s Q statistic under both the IVW and MR Egger methods, with P > 0.05 indicating no significant heterogeneity. 34 Horizontal pleiotropy was evaluated using the MR Egger intercept test, where the intercept reflects the average directional pleiotropic effect of genetic variants, with a P-value less than 0.05 suggests the presence of horizontal pleiotropy. 35 Additionally, symmetry in funnel plots was visually inspected as a qualitative indicator of potential pleiotropy. Leave-one-out analysis was performed to detect and correct influential outliers. Data download and MR analysis were completed in August 2025.
3. Results
3.1. Study Design
Our study was conducted in three steps, as illustrated in Figure 1. First, a bidirectional two-sample MR analysis was performed between GM and PCa to identify GM traits strongly associated with PCa risk and exclude potential reverse causality, thereby estimating the total effect. Second, plasma metabolites significantly associated with PCa risk were identified, obtaining Beta 2. In the third step, we assessed the causal relationships between the identified GM traits and candidate plasma metabolites, estimating Beta 1 and calculating the proportion of the mediation effect. Detailed information of the three GWAS datasets is presented in Table 1. A two-step MR study of GM on PCa mediated by plasma metabolites. MR, Mendelian randomization; GM, gut microbiota; PCa, prostate cancer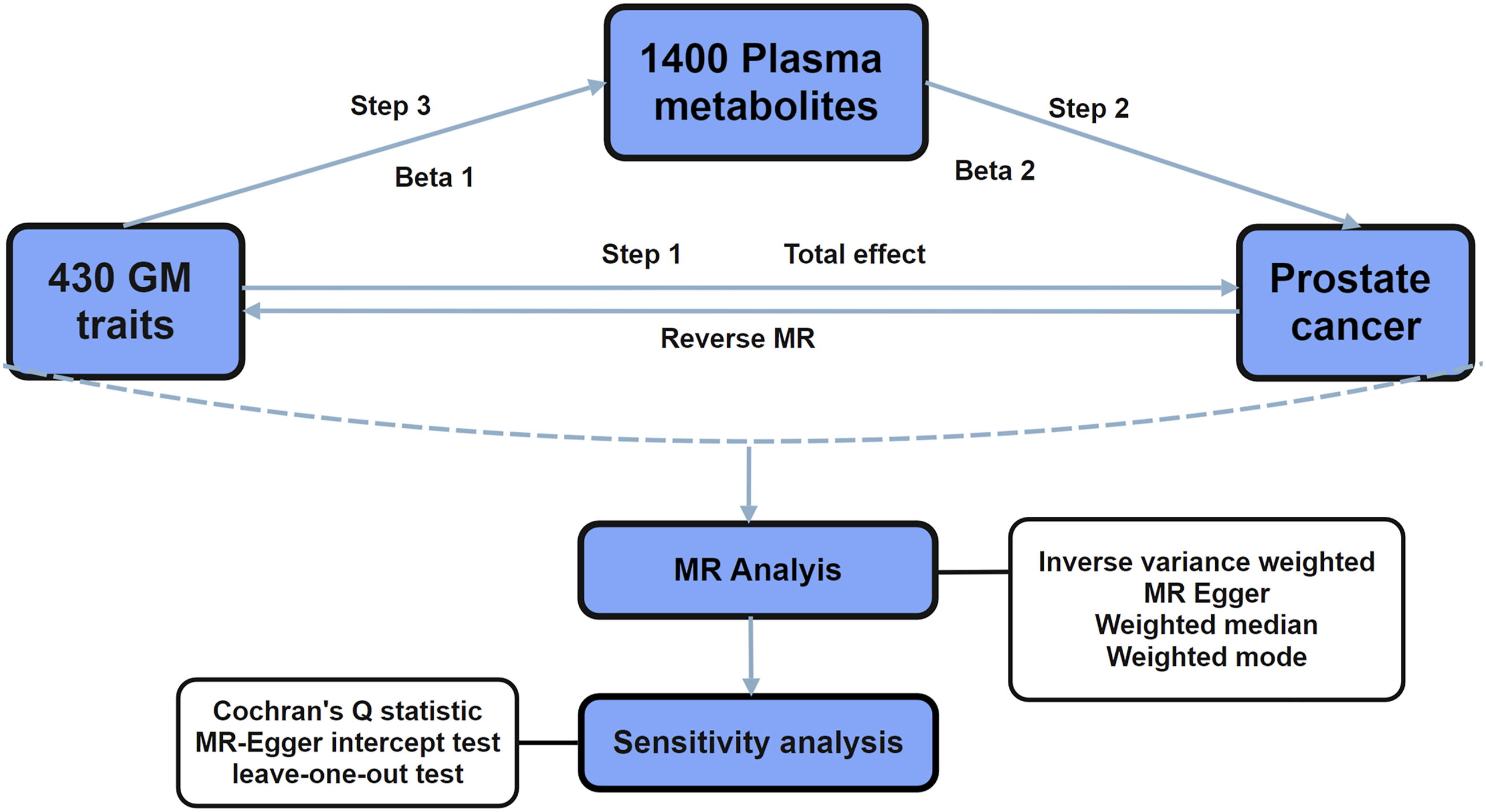
3.2. Bidirectional MR Analysis Between GM and PCa
Our rigorous MR analysis using the IVW approach identified 18 distinct GM traits with causal relationships to PCa (Figure 2 and Table S1). For example, P_Firmicutes (IVW: OR=0.937, 95% CI [0.893, 0.983], P=0.008), G_Roseburia (IVW: OR=0.945, 95% CI [0.899, 0.992], P=0.024), and Subdoligranulum (IVW: OR=0.946, 95% CI [0.903, 0.992], P=0.022) were found to be protective factors. Notably, the FDR-adjusted P-value (q value) for Bacteroides was 0.056, indicating that the corresponding causal inference is more credible than other findings. In contrast, Parasutterella (IVW: OR=1.029, 95% CI [1.001, 1.057], P=0.039), G_Sutterella (IVW: OR=1.034, 95% CI [1.003, 1.066], P=0.029), and Clostridiales (IVW: OR=1.047, 95% CI [1.006, 1.09], P=0.025) exhibited a positive correlation with PCa risk. Reverse MR revealed that G_Sutterella and Desulfovibrio had reverse causal relationships with PCa, leading to their exclusion from further analysis (Table S2). The causal relationships between 18 GM and PCa. GM, gut microbiota; PCa, prostate cancer
3.3. MR Analysis Between Plasma Metabolites and PCa
We obtained the causality between 66 types of plasma metabolite levels and ratios and PCa using IVW method (Table S3). Notably, six metabolite levels and two metabolite ratios were associated with PCa risk across four MR methods, with concordant effect directions (Figure 3). Specifically, higher levels of glucose-to-mannose ratio (IVW: OR=0.955, 95% CI [0.930, 0.981], P=0.001), N-acetylarginine (OR=0.970, 95% CI [0.954, 0.986], P<0.001), N-acetyl-L-glutamine (OR=0.970, 95% CI [0.955, 0.986], P<0.001), 5-methyluridine (ribothymidine; OR=0.978, 95% CI [0.958, 0.999], P=0.043), N-acetylhistidine (OR=0.981, 95% CI [0.964, 0.998], P=0.030), and spermidine-to-ergothioneine ratio (OR=0.991, 95% CI [0.983, 0.999], P=0.028) were associated with a reduced risk of PCa. In contrast, elevated levels of N6-methyllysine (OR=1.021, 95% CI [1.006, 1.037], P=0.006) and unidentified metabolite X-23680 (OR=1.030, 95% CI [1.005, 1.056], P=0.018) were linked to an increased risk. The causal relationships between 8 plasma metabolites and PCa with four methods. PCa, prostate cancer
3.4. The Mediating Role of Plasma Metabolites
The MR analyses between 16 GM and 66 plasma metabolites identified 41 potential causal associations using IVW method (Figure 4 and Table S4). We then performed mediation analyses to calculate the mediation P-values, effect sizes (β), and proportions of mediation. Four potential mediation pathways were revealed (Table 2). Specifically, the causal effect of Subdoligranulum on PCa was mediated by unidentified metabolite X-25422 levels (20.785% mediation, P = 0.022); the effect of Bacteroides was mediated by N-acetylarginine levels (6.706% mediation, P = 0.020); the influence of Parasutterella on PCa was negatively mediated by N-acetyl-L-glutamine levels (-9.382% mediation, P = 0.031); and the effect of G_Roseburia was also negatively mediated by N-acetyl-L-glutamine levels (-9.094% mediation, P = 0.036). The causal relationships between GM and plasma metabolites. GM, gut microbiota Mediation Effect of GM on PCa via Plasma Metabolites GM, gut microbiota; PCa, prostate cancer.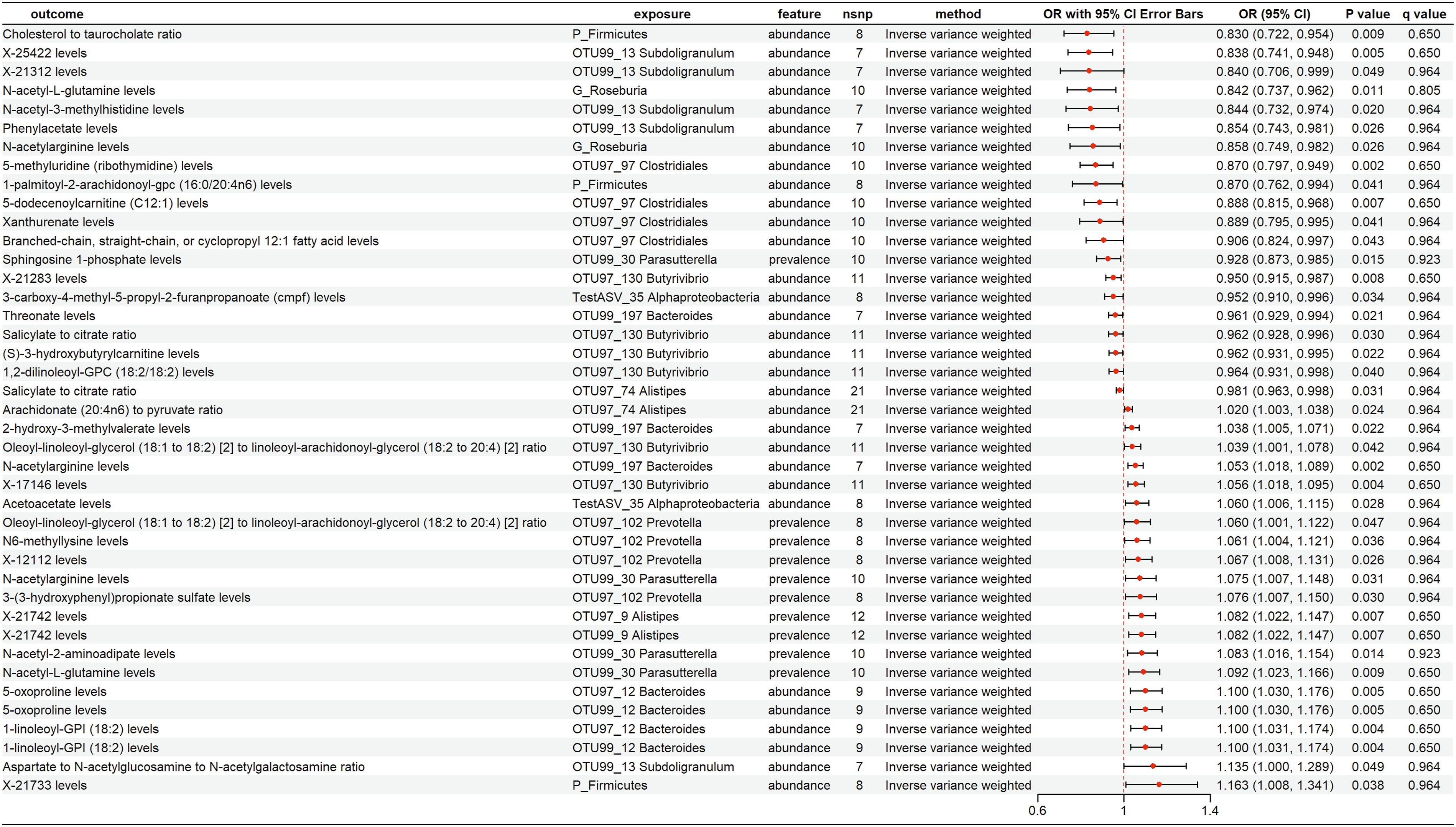

Positive mediation proportions indicate that the mediator amplifies the exposure-outcome association, while negative proportions indicate that the mediator attenuates this association. The negative mediation proportions of Parasutterella and G_Roseburia suggest that N-acetyl-L-glutamine acts as an anti-mediator, reducing the overall effect of these bacteria on PCa risk. Specifically, G_Roseburia was negatively associated with N-acetyl-L-glutamine, which in turn exhibited a protective effect against PCa. Consequently, the reduction in this protective metabolite partially counteracted the direct beneficial effect of G_Roseburia, leading to a negative mediation proportion. Parasutterella was positively associated with the protective metabolite N-acetyl-L-glutamine, which in turn reduced PCa risk and partially counteracted the pro-tumorigenic effect of Parasutterella.
3.5. Sensitivity Analysis and Visualization
In all our analyses, the p-values derived from Cochran’s Q statistic using both IVW and MR Egger methods were greater than 0.05, indicating no evidence of significant heterogeneity (Tables S5–S7). Similarly, the p-values from the MR Egger intercept tests exceeded 0.05, suggesting the absence of horizontal pleiotropy (Tables S5–S7). Corresponding forest plots, leave-one-out analyses, scatter plots, and funnel plots of the four significant mediation results are presented in supplementary figures S1–S4.
4. Discussion
Dysbiosis induced by a high-fat diet can compromise intestinal barrier integrity, allowing lipopolysaccharides to translocate into the circulatory system, thereby activating the Interleukin-6/STAT3 signaling pathway and promoting the progression of PCa. 36 Moreover, certain gut bacteria capable of synthesizing testosterone 37 or converting androgen precursors into bioactive androgens contribute to castration resistance in PCa. 38 These findings collectively support the existence of the “gut-prostate” axis, sparking continuous interest in the functional mechanisms of GM in PCa. Recent MR studies have further demonstrated a causal relationship between GM and PCa from a genetic perspective. 39 Building on a systematic evaluation of the causal relationships among GM, plasma metabolites, and PCa, our study is the first to introduce plasma metabolites as mediators to assess their bridging role in the GM-PCa causal pathway. We identified 18 GM traits significantly associated with PCa and, notably, revealed that four plasma metabolites act as key mediators in this causal chain.
In line with previous research, our study confirmed that Bacteroides and Ruminococcaceae exert a protective effect against PCa,40,41 while Butyrivibrio correlates with elevated PCa risk. 42 Many gut bacteria, primarily within the Firmicutes and Bacteroidetes phyla, are capable of fermenting dietary fibers and other indigestible carbohydrates into short-chain fatty acids (SCFAs), such as acetate, propionate, and butyrate. 43 For instance, Ruminococcaceae degrades resistant starch into SCFAs, 44 while Prevotella metabolizes hexose and pentose sugars into propionate via the succinate pathway. 45 Through the inhibition of histone deacetylases, SCFAs suppress tumor cell proliferation and migration, and to induce apoptosis. 46 Conversely, Matsushita et al suggested that SCFAs may promote PCa progression by activating the IGF1-MAPK/PI3K signaling pathway, 47 indicating a potentially dual role. However, these studies did not distinguish the specific biological effects of individual SCFAs. Our study found that SCFAs-producing bacteria, including Firmicutes and Ruminococcaceae, were negatively associated with PCa risk, whereas Clostridiales were positively associated, highlighting the complexity and context-dependence of SCFAs function. Moreover, Clostridiales are known to catalyze the side-chain cleavage of glucocorticoids into androgens, facilitating PCa progression, 48 which is consistent with our findings.
By disrupting glycolysis, decreasing mitochondrial membrane potential, increasing reactive oxygen species levels, and reducing ATP production, ultimately inducing mitochondrial apoptosis, mannose can inhibit the growth and proliferation of PCa cells. 49 Additionally, mannose can suppress the Warburg effect in tumor cells, thereby slowing their proliferation and enhancing sensitivity to chemotherapeutic agents. 49 Although vitro and animal experiments have preliminarily demonstrated its antitumor effects, 50 further investigation is required to validate its clinical efficacy. On the other hand, spermidine has been reported to promote ferroptosis in PCa cells by modulating proteasome activity. 51 We further identified that glucose-to-mannose ratio and the spermidine-to-ergothioneine ratio were protective factors against PCa and underscored the critical role of metabolite ratios in cancer interventions.
5-methyluridine, a key metabolite in the folate metabolic pathway, exerts antitumor effects by regulating the cell cycle and inhibiting cell proliferation. 52 Clinical studies have demonstrated that 5-methyluridine levels are significantly lower in patients with PCa compared to those with benign prostatic hyperplasia, 53 suggesting its potential as a biomarker for early diagnosis and prognosis. Our study further provides robust evidence supporting a negative association between 5-methyluridine and PCa.
In our mediation analysis, the unknown metabolite X-25422 levels accounted for 20.785% mediation proportion of the causal pathway between Subdoligranulum and PCa. Overall, Subdoligranulum exhibited a protective effect against PCa, whereas X-25422 levels were positively associated with PCa risk. X-25422 levels are downregulated by Subdoligranulum, thereby exerting its mediating role. Notably, Subdoligranulum is a major butyrate-producing bacterium in the gut, 54 and its metabolite butyrate—an essential energy source for colonic epithelial cells—plays a critical role in preserving intestinal mucosal integrity and barrier function. 55 Moreover, it mitigates inflammation by downregulating pro-inflammatory cytokines such as Interleukin-17 and upregulating anti-inflammatory cytokines like Interleukin-22, thereby contributing to a reduced risk of gastrointestinal tumorigenesis. 56
Our study identified N-acetylarginine and N-acetyl-L-glutamine as potentially critical protective mediators against PCa. Notably, N-acetylarginine is a key N-acetylated amino acid metabolite and serves as a central node in the arginine metabolic pathway. Serum metabolomic profiling demonstrated that the level of N-acetylarginine was statistically significantly lower in men who were subsequently diagnosed with PCa. 57 Furthermore, prospective serum metabolomic analysis validated a statistically significant reduction in N-acetylarginine levels in patients with T4 stage PCa. 58 In contrast, N-acetyl-L-glutamine, a core derivative of glutamine metabolism, has remained relatively understudied in the context of PCa. Further metabolomics-based studies are thus warranted to clarify the metabolic pathways and functional mechanisms of these identified metabolites in tumors.
Identification of novel GM-metabolite axes and metabolic ratio signatures linked to PCa pathogenesis enabled us to establish a framework for advancing PCa risk stratification and therapeutic optimization. The protective effects of Bacteroides, Ruminococcaceae, and Subdoligulum support probiotic supplementation or fecal microbiota transplantation as potential primary prevention strategies, while Butyrivibrio-based risk stratification tools could facilitate early screening of high-risk populations. As integrated biomarkers that better reflect metabolic network dynamics than individual metabolites, these metabolic ratios require validation of their supplementary diagnostic value in prostate-specific antigen screening-negative cohorts via prospective studies. For translational research, we propose a “microbiota-metabolite-host” trinity multi-omics platform, and recommend prospective nested case-control studies to verify causal associations and assess the safety and efficacy of metabolic interventions combined with standard therapies (e.g., androgen deprivation therapy or novel endocrine therapies) for PCa.
Our study provides compelling evidence supporting the pivotal role of GM and metabolites in PCa. Nonetheless, this study has certain limitations. In our present MR analysis, most associations did not meet the strict FDR threshold of < 0.05, likely due to limited statistical power, mild genetic heterogeneity, and minor measurement errors in exposure or outcome phenotypes. Notably, our findings are consistent with previous MR studies where marginally significant signals were later validated in larger datasets. Future multi-ancestry GWAS with expanded sample sizes will be critical to enhance statistical power and permit more robust identification of modest causal effects.
5. Conclusion
Our two-sample MR analysis identified 18 GM traits and 66 plasma metabolites with potential causal effects on PCa risk, among which 4 metabolites acted as mediators. These findings provide novel mechanistic insights into microbiota-driven PCa pathogenesis, highlighting the intricate crosstalk between the gut ecosystem and host metabolism in modulating PCa susceptibility. We further recognized protective microbial taxa (e.g., Bacteroides, Ruminococcaceae, Subdoligranulum) and metabolites (e.g., mannose, spermidine, N-acetylarginine), which represent promising targets for mechanism-guided preventive strategies. Future translational studies are warranted to validate these microbial and metabolic biomarkers in prospective cohorts and explore their utility as personalized intervention targets for PCa management.
Supplemental Material
Supplemental Material - Causal Association Between Gut Microbiota, Plasma Metabolites, and Prostate Cancer: Two-Step Mendelian Randomization Study
Supplemental Material for Causal Association Between Gut Microbiota, Plasma Metabolites, and Prostate Cancer: Two-Step Mendelian Randomization Study by Junxiong Li, Jian Hou, Haolin Liu, Shicheng Fan, Yumin Wang, Pinyao Liang, Jingbo Qin, Peng Gu, Xiaodong Liu and Xiangyang Wen in Dose-Response.
Footnotes
Acknowledgements
We acknowledge the use of summary-level data from the NHGRI-EBI GWAS Catalog. We thank all investigators and participants involved in these studies.
Ethical Considerations
This study leveraged publicly available GWAS datasets, which had already acquired ethical approval in the primary investigations.
Consent to Participate
Participant consent was obtained in the primary investigations.
Author Contributions
J.X. Li, J. Hou, and H.L. Liu conceived and designed the study. J.X. Li and S.C. Fan analyzed the data. H.L. Liu, Y.M. Wang, and P. Gu prepared the supplementary materials. P.Y. Liang, J.B. Qin, and X.D. Liu generated the figures and tables. X.Y. Wen supervised the project. J.X. Li drafted the manuscript. All authors contributed to manuscript revision and approved the final version.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The data utilized in this study were provided in the table and supplementary materials.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.