
Abstract
Little is known about the molecular mechanisms of ascending thoracic aortic aneurysms (ATAAs). Abnormal extracellular matrix changes and variations of vascular smooth muscle cells (VSMCs) have been implicated in abdominal aortic aneurysm formation. Our objective was to investigate the alterations of collagen, stimulators of collagen synthesis and synthetic VSMCs in patients with ATAA. Surgical samples from ATAA were taken from 20 patients, and 18 control aortas were obtained during coronary artery bypass surgery. All aortic wall specimens were fixed for histology and immunohistochemistry for collagen, connective tissue growth factor (CTGF) and osteopontin. Realtime polymerase chain reaction was used to determine their mRNA expression. Histology and semi-quantitative analysis demonstrated that protein levels of collagen, CTGF and osteopontin significantly increased by 1.9-, 1.4- and 2.2-fold, respectively (P < 0.01 for all) in the ATAA group than in the control group. Similar results were shown in mRNA levels of type Iα1and IIIα1 collagen, CTGF and osteopontin. The protein levels of CTGF and osteopontin were positively correlated with aortic diameter (r = 0.67, r = 0.73; P < 0.01 for both). In conclusion, overexpression of aortic CTGF and synthetic VSMCs marker (osteopontin), which is likely to be responsible for elevated aortic collagen content, may provide a potential mechanism for aneurysmal enlargement.
Keywords
Introduction
Ascending thoracic aortic aneurysm (ATAA) is a chronic degenerative pathology with a very high mortality. 1 Its incidence is believed to be increasing due to a combination of an aging population and improved diagnostic tools. Two broad categories of ATAA include degenerative aneurysms and aneurysms associated with genetic syndromes. The latter is divided into syndromic and familial, such as Marfan syndrome, Loeys–Dietz and Ehler–Danlos syndrome, bicuspid aortic valves syndrome and Turner syndrome. 2
The mechanisms driving dilation of the ascending aorta during aneurysm formation are poorly understood and mostly extrapolated from those of abdominal aortic aneurysms. 3 However, the heterogeneous nature of the aortic aneurysms is underscored by epidemiological, genetic, transcriptional and histopathological distinctions between degenerative aneurysms of the thoracic and abdominal aortas. 4 Recent studies have begun to elucidate the cellular and molecular mechanisms responsible for ascending thoracic aneurysm development. Clinical and basic research studies have demonstrated an imbalance between the synthesis and destruction of structural matrix proteins, particularly collagen and elastin, in the wall of aortic aneurysms. 5 Collagen and elastin are the most abundant extracellular matrix (ECM) proteins in the aortic wall, and they play important roles in the maintenance of aortic wall elasticity and tensile strength. Recent studies concerning collagen content are variable in degenerative ATAA patients, and little is known about the alteration of collagen and stimulators of collagen synthesis in patients with degenerative ATAA. 1
Connective tissue growth factor (CTGF) is a cysteine-rich 36–38 kDa secreted protein, first described in 1991, which belongs to a set of structurally related proteins of the CCN family. 6 Studies have shown that CTGF stimulates the proliferation of fibroblasts and enhances ECM production. 7 Moreover, recent evidence indicated that CTGF appears to act downstream of transforming growth factor-β to induce ECM production. 8 CTGF has been implicated in the development of cardiac disease 9 and progressive fibrotic diseases 8 of diverse etiology. Studies of diseased tissues from human clinical specimens have also shown that CTGF is expressed in thoracic aortic dissection arteries. 10 However, the presence and localization of CTGF in degenerative ATAA remain unknown.
It is well established that vascular smooth muscle cells (VSMCs) have the ability to undergo profound changes in phenotype from contractile to synthetic in response to the changes of their extracellular environment, as occurs after vascular injury in both preclinical animal models and human subjects. 11 Osteopontin (OPN) is a highly phosphorylated sialoprotein which is a prominent component of the mineralized extracellular matrices of bones and teeth. 12 Recent studies have shown that the expression of osteopontin provides a useful marker which can be applied to distinguish synthetic VSMCs from all VSMCs. 13 Even though the research on total number of smooth muscle cells in ATAA formation has been widely investigated in vivo and in vitro,5,14 much less attention has been paid to the expression of synthetic VSMCs in ATAA specimens.
The purpose of the present study was to determine mRNA and protein expression levels of CTGF, collagen and synthetic VSMCs marker in the aortic wall of degenerative ATAA patients compared with control aortas.
Patients and methods
Patient enrollment and tissue collection
This study was approved by the ethics committee of FuWai hospital. Written consent was obtained from all patients. Clinical data and aortic specimens were collected during a 12-month period (March 2009 to March 2010) from patients with ATAA undergoing replacement of ascending thoracic aorta (n = 20) and patients with non-aneurysmal aortas undergoing coronary artery bypass surgery (n = 18). All patients with ATAA underwent elective surgery. Relatively young patients (<55 years of age) and patients with aneurysms secondary to genetic syndromes such as Marfan, Ehlers-Danlos and Loeys-Dietz syndrome and patients with bicuspid aortic valves were excluded from this study. Patients’ preoperative ascending thoracic aortic diameter was recorded, as evaluated during the echocardiographic examination or computed tomographic scanning. Full-thickness biopsies containing all three layers of the aorta were obtained in the operating room from the right-lateral aspect of the ascending aorta (the greater curvature), and they were formalin-fixed and freshly frozen in liquid nitrogen and stored at −80℃ until analysis.
Histology and immunohistochemistry
Aortic specimens were fixed in 10% buffered formalin, embedded in paraffin. Serial 3-μm-thick sections were cut and stained with hematoxylin and eosin for examination of overall vessel morphology, Masson's trichrome to investigate the localization and distribution of collagen using standard techniques and reagents. Other sections were labeled with anti-CTGF (ab5097; Abcam plc, Cambridge, UK), anti-osteopontin (ab33046; Abcam plc, ), and the immunohistochemistry of osteopontin was used to determine the density of synthetic VSMCs.
For immunohistochemistry, sections were heated and fixed onto glass slides, deparaffinized in xylene and rehydrated through graded ethanol. The sections were then soaked in 3% hydrogen-peroxide methanol for 10minutes, washed five minutes three times in phosphate buffer solution (PBS), microwave antigen hot fixed 15 min in repair citrate solution, incubated the coverslips in fetal bovine serum at 37℃ for 20 minutes after washing in PBS (5 minutes×3). Primary antibody to CTGF (1:50 dilution) or primary antibody to osteopontin (8 μg/mL) was applied to tissue section for one hour at 25℃. Biotinylated goat anti-mouse antibody (GBI Inc., Mukilteo, WA, USA) and goat anti-rabbit antibody (GBI Inc.) were applied and incubated for one hour, respectively, followed by chromogenic substrate with 3,3′-diaminobenzidine. Slides were subsequently counterstained with hematoxylin. Antibody was replaced with PBS solution as a negative control.
The slides were examined with an optical microscope (Leica microscope; Frankfurt Am Main Area, Germany) and processed with coupled image-analysis software (leica qwin v3; Frankfurt Am Main Area). We graded fragmentation and disorientation of collagen from I (no or mild change) to III (severe change) with use of the following criteria: grade I, fewer than two foci with collagen fragmentation and disorientation in one microscopic field; grade II, 2–5 foci in one microscopic field; grade III, five or more foci in one microscopic field. To quantify the staining of collagen, we took images of three randomly selected high-power fields of each sample slide (original magnification:×40). The area occupied by collagen was quantified by color-detecting mode of the computer program on six regularly spaced areas in each aortic specimen. Brown particles or regions under the microscope were considered as the positive areas of immunohistochemical streptavidin-perosidase (SP) method, A semi-quantitative analysis of staining for CTGF and osteopontin was performed, evaluating the proportion of positive staining expressed as a percentage of the entire tissue. Value for each specimen was average from six sections. All sections were evaluated by two reviewers blinded to clinical data.
Real-time quantitative polymerase chain reaction
Aortic specimen was ground in liquid nitrogen and total RNA was extracted by homogenization in Trizol reagent (Invitrogen Corporation, Carlsbad, CA, USA) according to the manufacturer's protocol. The RNA underwent reverse transcription (RT) reactions with reverse transcriptase (M531A; Promega, Madison,WI, USA). Realtime quantitative polymerase chain reaction (PCR) reactions were prepared with Taqman PCR reagents (Bio-Rad Laboratories) and primers designed according to their recommendations, including CTGF, osteopontin, collagen type 1α-1, collagen type 3α-1 and GAPDH (glyceraldehyde-3-phosphate dehydrogenase). The primer sequences for the genes were as follows: CTGF: (forward) 5′ G T G C A C CG C CA A AG AT G G T 3′, (reverse) 5′ A A G G A C T C T C C G C T G C G GTA 3′); Collagen Iα1: (forward) 5′ C A A T G G T G C T CC T G G T A T T G C T 3′, (reverse) 5′ C T G T T A C C CT T G G G A C C A G G A 3′; Collagen IIIα1: (forward) 5′ G C C C C G T AT TA TGG AGA TGA 3′, (reverse) 5′ GG GG TT T TT ACG AGA ACC ATC 3′). Osteopontin: (forward) 5′ AG GAG GA GG CAG AG CA CA 3′, (reverse) 5′ CT GGT ATG GCAC AGG TGA TG 3′; and GAPDH: (forward) 5′ GG GA AGG TG A AG GTC GGA GT 3′, (reverse) 5′ TT GA GG TC AA T GA AG G GG TCA 3′. Realtime PCR was performed using 1 μL of target cDNAs for 95℃ 15 seconds, followed by 50 cycles at 95℃ for 15 seconds and at 60℃ for one minute and 95℃ for 15 seconds, then 95℃ for 15 seconds, 60℃ for 30 seconds and 95℃ for 15 seconds with the use of a SYBR Green Master mix Kit (TOYOBO, Tokyo, Japan) in a Bio-Rad CFX96 Real-Time PCR system (Bio-Rad Laboratories). For all samples, GAPDH RNA was amplified using an internal control. Relative Gene Expression was calculated using the 2−ΔΔCT method. Products were sequenced to ensure specificity. An iCycler and its system interface software were used to run both samples and analyzed data (Bio-Rad Laboratories).
Statistical analysis
Continuous variables are presented as the mean ± SD. Intergroup differences were analyzed by independent Student's t-test for comparisons between two groups. Chi-square test and Fisher's exact test (n < 40) analysis were used to examine the relationships between categorical variables. Spearman rank correlations and Pearson correlations were also performed to describe the association between different outcome variables when appropriate. All statistical analyses were performed in SPSS 13.0 (SPSS, Inc., Chicago, IL, USA). Two-tailed probability values of P < 0.01 were considered statistically significant for each test to ensure an overall study significance level of P < 0.05.
Results
Aortic specimens and patient characteristics
Patient characteristics
ATAA, ascending thoracic aortic aneurysm

(a, b) Representative serial sections stained with hematoxylin and eosin show smooth muscle cells are fragmented and disoriented in the aortic wall of ascending thoracic aortic aneurysm (ATAA) (b) as compared with control aortas (a). (magnification ×400). (c, d) Representative sections of aorta stained with Masson's trichrome demonstrate increased collagen and decreased elastin in the media of the aortic wall of ATAA patients (d) as compared with controls (c). Collagens are stained blue and elastins are stained red (magnification ×200)
Increased expression of CTGF in the aortic wall of ATAA patients
Analysis of realtime RT-PCR data showed that mRNA levels of aortic CTGF expression significantly increased in ATAA patients as compared with those in controls (P < 0.001, Figure 2b). The protein levels of CTGF were evaluated using semi-quantitative immunohistochemistry, which revealed increased positive staining for CTGF in ATAA (Figure 2c) than in controls (Figure 2d), and it was mainly localized in media and adventitia in both groups. Further analysis showed that positive area percentage for the ATAA samples is roughly 1.4-fold higher than those for the control samples, and these results are congruent with realtime PCR analysis.
(a,b) Aortas of ascending thoracic aortic aneurysm (ATAA) patients showed significantly increased connective tissue growth factor (CTGF) protein (a) and mRNA (b) levels compared with controls (mean values ± SD; *P < 0.01, independent Student's t-test). (c, d) Representative immunohistochemical sections of aorta demonstrate increased positive staining for CTGF in the medial vascular smooth muscle cells from ATAA patients (c) compared with controls (d) (magnification: ×400)
Elevated collagen in the aortic wall of ATAA patients
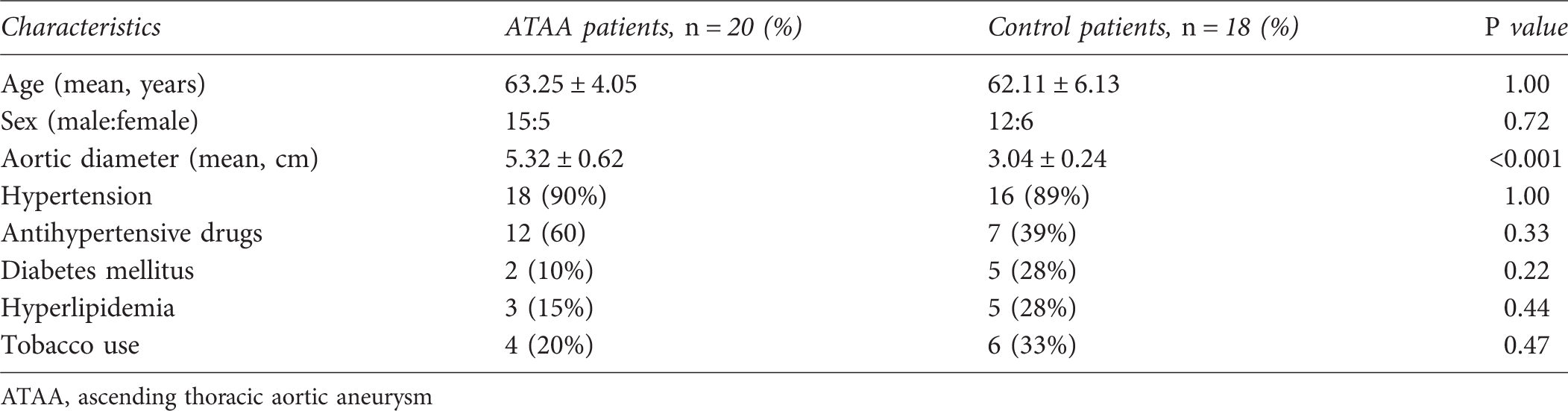
Compared with controls, mRNA levels of both collagen Iα1 and IIIα1 significantly increased in ATAA patients (P = 0.002 and P < 0.001, respectively; Figure 4b) than in controls (Figure 4b). To find the collagen levels in ATAA, the aortic wall was stained with Masson's trichrome, collagen was localized to the aortic media and adventitia in both groups, as shown in Figures 1c and d. The protein expression of collagen was then determined by quantitative image analysis. The area percentage of staining for collagen was significantly higher in aneurysms compared with non-aneurysmal aortas (P < 0.001, Figure 4a).
Enhanced expression of osteopontin in the aortic wall of ATAA patients
There was a significant increase in mRNA expression of osteopontin in the ATAA group as compared with the control group (P < 0.001, Figure 4b). Representative photomicrographs from immunohistochemical staining are shown in Figure 3. According to semi-quantitative results, the protein expression level of osteopontin (Figure 4a) was much higher in specimens taken from pathological aortas (area percentage: [24.06 ± 7.15]%) than in the control specimens (area percentage: [10.69 ± 5.41]%).
Representative aortic sections of immunohistochemistry for osteopontin in the media of pathological (b, d) and control (a, c) aortas. It reveals much stronger expression of immunopositive brown color reaction in the medial vascular smooth muscle cells of pathological aortas (b, d) than in controls (a, c) (a, b: magnification ×100; c, d: magnification ×400) (a) The protein level of collagen and osteopontin in aneurysm walls of patients and controls. (b) The mRNA level of collagen 1α1, collagen 3α1 and osteopontin in corresponding aneurysm walls. Bars represent group mean values ± SD. *P < 0.01, independent Student's t-test
Correlation analysis
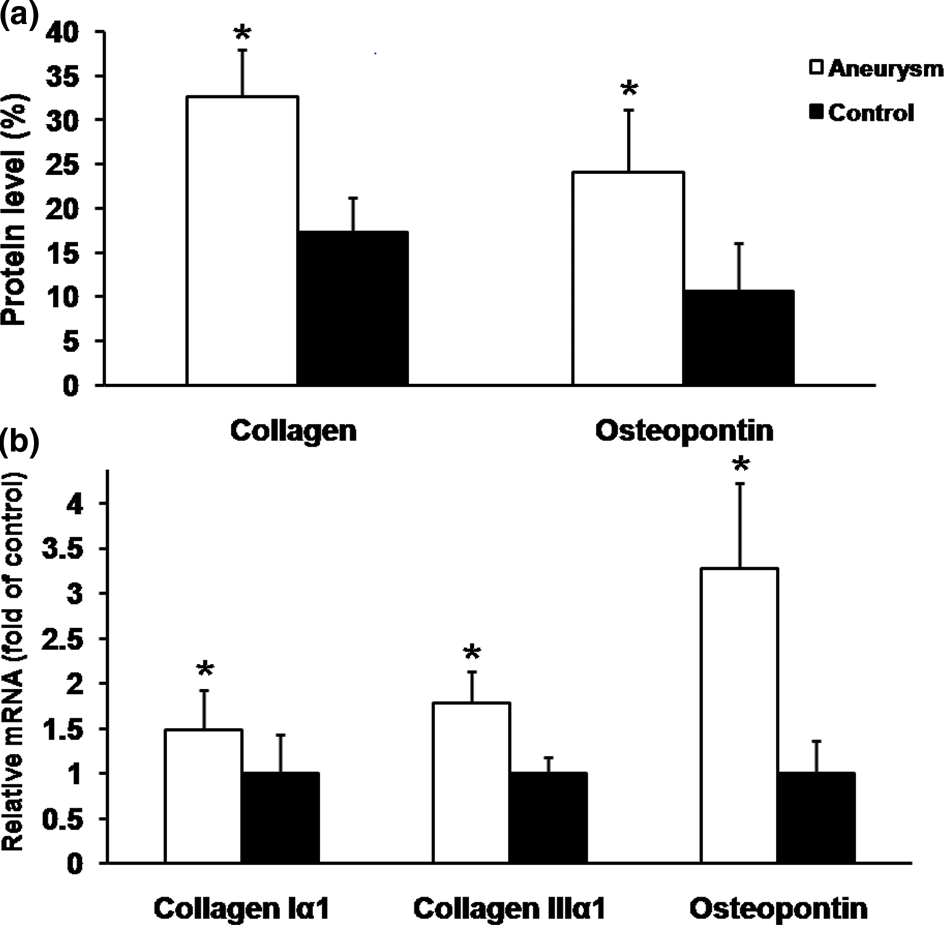
CTGF and collagen protein expression levels showed significant positive correlations with the size of the aneurysm (r = 0.67 and r = 0.73, respectively; P < 0.001 for both), as shown in Figures 5a and b, respectively. Osteopontin and collagen protein expression levels showed positive correlations with the CTGF protein levels (r = 0.59 and r = 0.57, respectively; P < 0.001 for both).
(a,b) Scatter plots of the protein expression levels of connective tissue growth factor (CTGF) (a) and osteopontin (b) with aneurysm size are shown. CTGF and osteopontin protein expression positively correlated with aortic dilation (r = 0.67, r = 0.73, respectively; P < 0.01, both)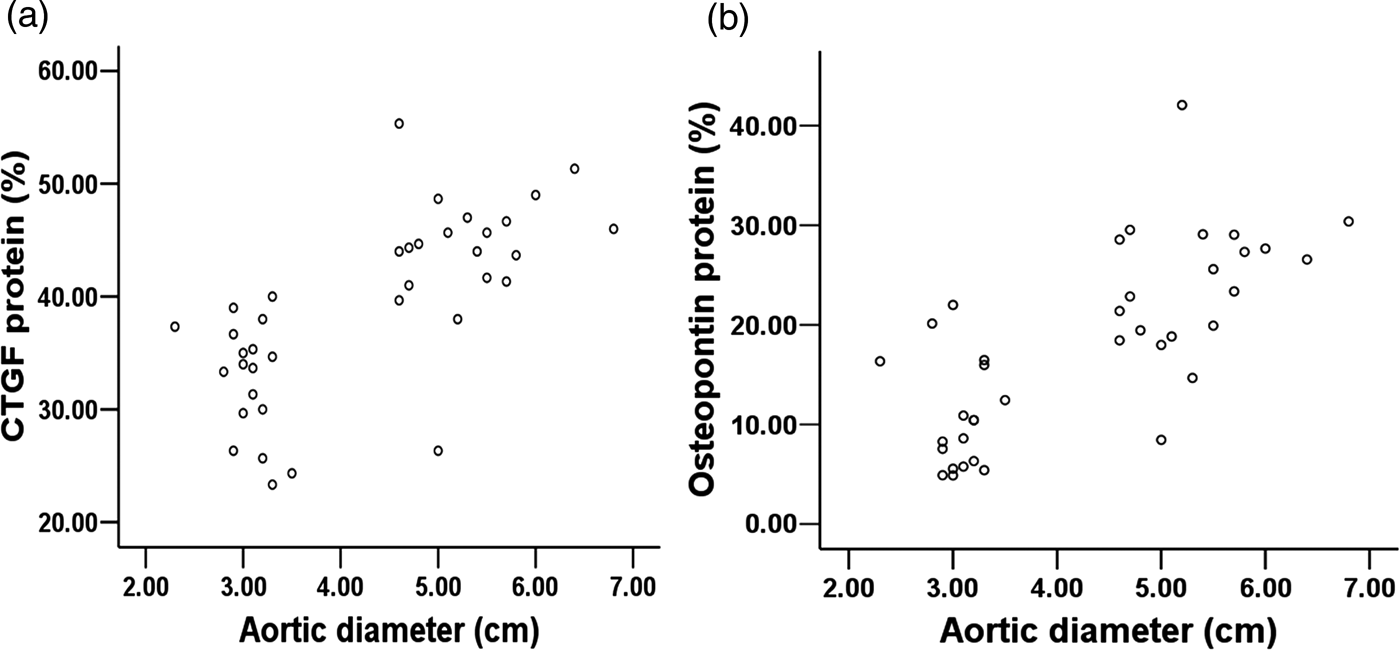
Discussion
This study confirmed that patients with ATAA have increased CTGF expression in aortic wall tissues. Even though collagen fibers were fragmented and disoriented in ATAA tissues as compared with control tissues, we found the aortic walls of patients with ATAA had increased protein levels of collagen in both the media and adventitia and mRNA levels of collagen Iα1 and IIIα1. Our study also showed the number of synthetic VSMCs was increased in ATAA aortic wall tissues.
CTGF as an active growth factor can be secreted by human endothelial cells and VSMCs, and its expression becomes restricted to the atria and large blood vessels in the adult heart, 15 which make CTGF highly related to arterial diseases. CTGF promote fibroblast proliferation, angiogenesis and ECM synthesis and its expression is regulated by several factors, such as transforming growth factor-β, high glucose and angiotensin II.16,17 Our results showed that CTGF mRNA levels and protein levels were significantly increased in human specimens from ATAA patients as compared with control aortas, and the protein level was associated with the collagen level and size-diameter of the aneurysm. Previous studies have shown that CTGF may regulate the production of ECM proteins in VSMCs, as VSMCs expressing CTGF were localized predominantly in areas with ECM production. 18 Knockdown of the CTGF gene by CTGF-siRNA markedly reduced the synthesis of ECM proteins such as collagen type I. 19 We infer, therefore, that such a change in CTGF expression within the aortic tissues may play a role in pathogenesis and progression of ATAA.
In native aortic vessels, the ECM is composed mostly of type I and III collagens, elastin and proteoglycans. These proteins play an important role in maintaining vessel structure by providing tensile strength (collagens) and elasticity (elastin). 20 In the steady state, the cyclic stretching of artery walls sustains a quiescent, contractile VSMC phenotype and slow turnover of ECM proteins. Once when physical or chemical injury started and aggravated, the arteries would react to the new environment by a process termed ‘vascular remodeling’, which involves the activation of a wide range of intracellular signaling pathways leading to the modulation of vascular cell changes as well as synthesis and degradation of the ECM. The balance between elastin and collagen, normally maintained by a slow process of protein production and degradation, is also destabilized. 20
Fragmentation and decreased content of elastin have been implicated in aneurysmal diseases by most studies.21,22 In contrast with elastin, which has a much lower turnover rate, collagen is synthesized continuously throughout life, reported to be in the range of decades. In the present study, we observed that patients with ATAA have elevated collagen content in aortic wall tissues, which was most probably due to increased production rather than reduced degradation, because collagen mRNA was also upregulated. Therefore, we speculate that collagen may play an important role in the process of ATAA vascular remodeling. Support for this result comes from Wang et al. 10 who have shown that aortic collagen types I and III levels are differentially increased in thoracic aortic dissection tissues. Another similar finding to our results was recently reported by Iliopoulos et al., 23 who observed a significant increased collagen density in the adventitia, however, there was an invariant density in the entire wall. The study of Tang et al. 5 also investigated the microstructure of collagen and elastin fibers in ATAA, reporting dissimilar alterations in ECM composition of the media layer compared with that seen in abdominal aortic aneurysms. They found that absolute total content of collagen may have actually increased, and there was an increase in types I and III collagen transcript expression in aneurysms merely not statistically significant since medial remodeling was most evident in comparisons between non-aneurysmal aortas versus smaller aneurysms and did not evolve further in larger aneurysms. Compared with only two cases whose aortic diameter is greater than 6 cm in our selected cases, they have half (aortic diameter >6 cm) of 29 ATAA patients, which may be the reason that the two research results are different.
In contrast with our results, Toumpoulis et al. 1 showed that collagen types I and III mRNA expression levels were similar in ATAA as compared with controls, and there was a significant decrease in their corresponding protein levels. We found that in the study by Toumpoulis et al., patients in the control group were younger and 16 in 25 ATAA patients have larger aneurysms, and compared with that study, the two subject groups in our study were age matched. In our study, further analysis showed that the collagen content is associated with the CTGF level which can stimulate collagen synthesis. As our clinical observation is limited by the nature of the study's cross-sectional design, our results do not distinguish the cause-and-effect relationship between them.
Together with the findings of collagen and CTGF overexpression, our study also showed excess synthetic VSMCs in aortic tissues of ATAA patients. We examined the expression of osteopontin which is a marker of synthetic VSMCs, 24 and found its expressions in mRNA and protein level were increased. VSMCs are not terminally differentiated and can alternate between a quiescent, contractile state when in a healthy artery and a proliferative non-contractile state (synthetic-mode) under circumstances of injury, repair and regeneration. 25 Recent studies5,26 have shown that the density of medial VSMCs is at least preserved in ATAA. Therefore, we speculate that increased synthetic VSMCs may be associated with excessive collagen production. Our data also showed that the expression of osteopontin was positively correlated with the expression of CTGF; it is tempting to suggest that CTGF overexpression may predispose to smooth muscle cell phenotype changes. An interpretation of our findings is that excessive CTGF stimulated phenotype conversion of VSMCs, accordingly, promoting the synthesis of collagen in synthetic VSMCs. In an early period, excessive collagen may be a protective substance; it could maintain the elasticity and tensile strength of the aortic wall. Once decompensation, as a result of collagen is not enough to resist the increasing wall stress, the aortic wall gradually enlarges and eventually ruptures. However, because of restrictions on cross-sectional study, we cannot make a conclusion, and specific signal pathways are not clear because of the complexity of pathogenetic mechanism on ATAA. An animal model will be essential to evaluate the causal relationship in which the effects of CTGF knockdown or overexpression on aortic collagen accumulation and smooth muscle cell phenotype change.
The principal limitation of this descriptive and comparative study is that our analyses were made on only a few sections from a representative portion of surgically removed aortic wall segments. This study can only determine whether collagen content as well as CTGF expression play a role in end-stage disease but we cannot conclude earlier developmental stages of aneurysm formation from our observations. Another limitation is that we only selected a limited number of patients and tissue samples and we did not measure the varieties of elastin
In conclusion, our results demonstrate that CTGF and collagen are overexpressed in the aortic wall of ATAA patients. We also report increased osteopontin expression in thoracic aortas of ATAA patients. Excessive aortic synthetic VSMCs and collagen suggesting smooth muscle cell phenotype changes and increased collagen synthesis may have been an initial adaptive response to vascular dilation, and CTGF may play an important role in this process.
Footnotes
Acknowledgements
Supported by the National Natural Science Foundation of China (81070252).