
Abstract
Objectives
Regulatory T cells (Tregs) mediate immunomodulation and protect against atherosclerosis. It is considered that reducing the amount of pro-inflammatory mediators could be achieved by enhancing the anti-inflammatory response, and this may be considered one of the main targets for therapy development. The inhibitory cytokines secreted by Tregs mainly include interleukin-10 (IL-10) and transforming growth factor-beta (TGF-β). Based on its known immunosuppressive involvement with other inflammatory disorders, we hypothesized that the newly characterized cytokine interleukin-37 (IL-37) might be associated with the inhibitory functions of Treg in atherosclerosis. Immune regulatory functions of IL-37 have not been completely clarified. Accordingly, we speculated that IL-37 might play a regulatory role in the immunosuppression of Tregs in atherosclerotic disease.
Methods
Real-time polymerase chain reaction and enzyme linked immunosorbent assay were used to test gene expression and protein levels of IL-37 in peripheral blood and localized freshly resected arterial tissues from 84 patients with peripheral arterial occlusive disease and 50 non-atherosclerotic subjects. Results were correlated to disease hallmarks. We also evaluated the ability of recombinant IL-37 to modulate Treg cytokine secretion and T cell inhibition in relation to atherosclerotic disorder in vitro.
Keywords
Introduction
Atherosclerosis is a chronic inflammatory disease, characterized by generation and release of pro-inflammatory cytokines. 1 One of the most prevalent manifestations of atherosclerosis is peripheral arterial occlusive disease (PAOD), 2 which affects over 200 million people worldwide. 3 PAOD is a major health concern, and if management is not initiated immediately, the probability of disease progression and development of critical limb ischemia is high, which is an entity with a high mortality and high risk of limb amputation. Although treatment options have been improved in the last decades, including endovascular treatment possibilities, mortality and amputation rates are still relatively high. 4
It is well known that the equilibrium of the two opposites is mandatory for the maintenance of homeostasis in healthy individuals. While pro-inflammatory cellular responses are detrimental to the development of atherosclerosis, regulatory T cells (Tregs) mediate immunomodulation and protect against atherosclerosis. This balance can be broken in certain conditions including hyperlipidemia, hypertension, and diabetes, resulting in the development of atherosclerotic plaques and the onset of chronic lower limb ischemia. 5
Substantial evidence clearly demonstrates that decreased Tregs proliferation and/or dysfunction causes abnormal immune responses and pathologies. 6 Tregs are a specific subpopulation of T cells; the most typical Treg is CD4+CD25+. 7 These cells principally regulate immune response through the secretion of inhibitory cytokines including interleukin-10 (IL-10) and transforming growth factor-beta (TGF-β). 8 In fact, the balance between pro-inflammatory cells and cytokines on one hand, as well as Tregs and their anti-inflammatory cytokines on the other hand plays an essential role in the immunosuppressive process. 1
The newly discovered cytokine, interleukin-37 (IL-37), has recently been shown to have a promising role in atherosclerotic diseases as a potent inhibitor of inflammation by shifting cytokine equilibrium away from excessive inflammation. IL-37 is an IL-1 cytokine family member and was discovered and identified in the year 2000 by Kumar et al. 9 IL-37 can be detected in low levels in healthy human tissues 10 and in peripheral blood mononuclear cells (PBMCs) of healthy individuals. 11
Recently, an evidence of a significant regulatory role of IL-37 has been found in the development of inflammatory disorders, autoimmune diseases, and cancer immunology.10,12,13 It can suppress innate immune response, possibly by reducing the production of pro-inflammatory cytokines induced by Toll-like receptors (TLR). 14 In addition, Moretti et al. reported that IL-37 can significantly inhibit the activation of Th2/Th17 cells in mice with allergic aspergillosis, showing that IL-37 might also affect adaptive immunity. 15
Although research concerning the role of IL-37 and its mechanism in atherosclerosis is relatively scant, 16 recent studies have reported a significant increase of IL-37 in experimentally induced atherosclerosis.17,18 Moreover, a suggested role of IL-37 in inhibiting macrophage activation and its expression in the foam cells of coronary and carotid artery atherosclerotic plaques have been described. 19 Additionally, in apolipoprotein E−/− mice, IL-37 effectively decreased the area ratio between aorta plaque and vascular cavity, and the co-stimulation of IL-37 and oxidized low-density lipoprotein (ox-LDL) facilitated the transformation of macrophages into the anti-inflammatory subtype (M2), indicating that IL-37 can prevent atherosclerosis by modulating macrophage polarity. 20
Furthermore, a single nucleotide polymorphism of IL-37 (SNP rs3811047) has been identified to be associated with an increased risk of coronary artery disease (CAD). This allele variant is associated with decreased expression of IL-37, 21 indicating that IL-37 expression in humans has a role in the prevention of cardiovascular pathologies.
These studies indicated a potential protective role of IL-37 against atherosclerosis. However, until now, the biological functions and mechanism of immune regulatory functions of IL-37 have not been completely clarified. It is considered that reducing the amount of pro-inflammatory mediators could be achieved by enhancing the anti-inflammatory response, and this may be considered one of the main targets for therapy development. Therefore, we hypothesized that IL-37 may be able to inhibit the occurrence and progression of atherosclerosis by regulating Treg suppressive activity. In this study, we investigated if IL-37 can modulate the suppressive effect of Tregs and increase the secretion of their anti-inflammatory cytokines in atherosclerotic peripheral arterial disease.
Patients and methods
Study population and ethics
Eighty-four patients with chronic lower limb ischemia were recruited to the present study from the vascular surgery outpatient clinic in Alexandria Faculty of Medicine, between April 2017 and February 2019. Study protocols and the collection and use of blood and arterial samples were previously approved by the Medical Ethical Committee of Alexandria University, and bioethical research committee approval was taken. The study conforms to the principles outlined in the Declaration of Helsinki (1975) for use of human tissue or subjects. Signed informed consents were obtained from each subject included in the study.
Patients were assessed both radiologically (Figure 1) and clinically with specific consideration to comorbidities, and then scheduled for admission at the vascular surgery internal ward for further management of their atherosclerotic disease. Chronic lower limb ischemia diagnosis was determined by clinical assessment, color-encoded duplex sonography, and ankle brachial index (ABI), 22 according to the revised criteria of the Trans-Atlantic Inter-Society Consensus Document on Management of Peripheral Arterial Disease. 4 Categorization of PAOD severity was done using Rutherford classification. Chronic lower limb ischemia was defined as PAOD patients presenting with severe intermittent claudication (Rutherford III), ischemic rest pain (Rutherford IV), minor tissue loss (Rutherford V) or severe tissue loss or gangrene (Rutherford VI) in accordance to current guidelines. 23 In addition, 50 non-atherosclerotic subjects presenting to the vascular surgery unit for traumatic amputation were recruited to the study. Subjects who had a medical history of atherosclerotic disease including CAD or cerebrovascular arterial diseases were excluded. Amputated limbs’ arterial samples were used as non-atherosclerotic vessel controls and were prepared and stored in the same way as atherosclerotic arterial specimens.

Multi-slice CT arteriogram by injecting intravenous contrast with axial construction, showing a patient with chronic lower limb occlusive disease.
Exclusion criteria
Patients with acute ischemia or those with severe infection, as well as patients with previous history of vascular intervention or thrombolytic therapy in the previous month, were excluded. Patients with concomitant malignancies, inflammatory or autoimmune diseases, overt liver disease, and viral infections, such as HIV and hepatitis B, were not included in the study.
Tissue sampling and cell isolation from arterial specimens
Freshly resected atherosclerotic arterial tissues (parts of the arterial wall) were used for isolation of lymphocytes from arterial specimens (Figure 2). They were collected during revascularization surgery by arterial bypass in patients (stages III–V) from popliteal artery (supragenicular) or from common femoral artery (in those patients with markedly decreased popliteal artery lumen), or from popliteal artery (supra or infragenicular) in patients with stage VI during above or below knee amputation surgery, respectively. In addition, arterial tissues from amputated non-atherosclerotic control limbs were taken from popliteal artery (supra or infragenicular) during above or below knee amputation surgery. Around (100 mg) were cut and then enzymatically digested in RPMI 1640 medium containing 500 mg/ml liberase and 200 mg/ml DNase (Roche) for 45 min at 37°C with gentle shake every 10 min until all the tissue had resolved into a cell suspension. The resulting cell suspension was filtered through 70 ml cell strainers (BD Biosciences) and subjected to the density centrifugation as described. The specimens that were used for extraction of RNA were quickly frozen in liquid nitrogen. Those that were used for cultures were placed in RPMI 1640 medium.

Arterial sampling during revascularization surgery showing arterial biopsy during the surgery.
Gene expression of IL-37 by real-time polymerase chain reaction
Arterial specimens were removed quickly after perfusion with phosphate buffer saline (PBS) (Figure 3). TRIzol Reagent (Invitrogen, USA) was used for total RNA extraction from the arterial specimens according to the manufacturer’s instructions, and reverse transcription was performed with RNA-PCR Kit (Invitrogen, USA). RNA amounts and purity were determined by optical density measurements at 260 and 280 nm. First strand cDNA for qPCR analysis was then synthesized using random hexamer primers and RNase H-reverse transcriptase (Invitrogen). Gene expression levels of IL-37 were determined with SYBR Green Master Mix (Takara, Japan) on an ABI Prism 7500 sequence Detection System (Applied Biosystems, USA). Each sample was analyzed in duplicate and normalized to the house-keeping gene β-actin. Primers used were as follows: IL-37 – Forward: 5′-
Arterial specimen.
Blood sampling protocol and isolation of T cell subsets
PBMCs were separated from heparinized blood by gradient centrifugation at 1800 r/min for 30 min using Ficoll-Hypaque (1077) (Sigma-Aldrich). The diluted sample was overlayered gently over half its volume of Ficoll-Hypaque (1077). After centrifugation, the interface cells containing PBMCs were aspirated, pelleted, and resuspended in 1 ml medium (RPMI 1640) to determine cell count and viability. 24 PBMCs were immediately used for in vitro experiments or for further separation of CD3+ T-cells (CD3-labeled magnetic beads; MACS, Miltenyi Biotec, Bergisch Gladbach, Germany) as described elsewhere. Cell viability detected by Trypan blue staining was >95%.
Circulating serum levels of IL-10, TGF-β, and IL-37
Serum cytokine levels were detected strictly in accordance with the instructions of the commercially available enzyme linked immunosorbent assay (ELISA) kits. IL-37 ELISA kit was purchased from Abcam, UK (ab213798), while both IL-10 and TGF-β ELISA kits were purchased from Invitrogen, Thermo Fisher (IL-10: Catalog # EHIL10, TGF- β: Catalog # BMS249-4). Measuring was done at 450 nm on a Biocell HT1 ELISA microplate reader (TSZ Company, Minneapolis, USA). Minimal detectable concentration was 15.6 pg/ml for IL-37. Intra- and inter-assay coefficients of variation for all ELISAs were <5%. Samples were measured in duplicates.
Identification of T effector and Treg cells
For identification of CD4+CD25+ Tregs and CD4+CD25- effector T cells, we used a Treg isolation kit (Miltenyi Biotech, Auburn, CA, USA). Briefly, CD4+ T cells were first negatively selected on a separation column, then the CD4+CD25+ and CD4+CD25− subsets were purified using PE-anti-CD25 antibody and anti-PE magnetic beads (Miltenyi Biotech, Auburn, CA, USA) according to the manufacturer’s instructions. The purity of Treg CD4+CD25+ was checked using intracellular staining for Foxp3 and in all experiments was >90% as confirmed by FACS.
For suppression assays of CD4+CD25− cells, CellTrace™ Carboxyfluorescein succinimidyl ester (CFSE) Cell Proliferation Kit for flow cytometry was purchased from Invitrogen, Thermo fisher (Cat no.: C34554). Briefly, CFSE stock solution (5 mM) was prepared in DMSO. CD4+CD25− T cells were washed twice with cold PBS and resuspended (106/ml) in PBS containing 5 μM of CFSE. Cells were incubated in the dark for 30 min; then culture medium was added to stop the process of staining. After centrifugation, cells were pelleted, resuspended in fresh culture medium and incubated for 15 min, washed twice with PBS, counted, and used for in vitro suppression assay.
Treg cell culture and measurement of released IL-10 and TGF-β
Isolated Treg cells (2 × 106 cells/ml) were cultured in supplemented media stimulated with recombinant IL-37 or without IL-37 at 37°C in 5% CO2. Culture conditions included complete RPMI 1640 culture medium supplemented with 10% heat-inactivated fetal calf serum (Gibco BRL), 100 U/ml penicillin, 100 μg/ml streptomycin, anti-CD3 monoclonal antibody (mAb) combined with anti-CD28 mAb at a final concentration of 2 μg/ml for stimulation (BD Biosciences, Mountain View, CA, USA and Invitrogen) and phytohemagglutinin (10 μg/ml; Sigma). A measured quantity of 100 ng/ml of recombinant IL-37 (#9225-IL-025; R&D systems, Bio-techne) was added to one of the two panels of the culture plate for Treg stimulation. Following culture, part of Treg cells was collected by centrifugation at 1000 r/min for 5 min, and for the cytokine analysis, plates were centrifuged and supernatants were harvested. Released IL-10 and TGF-β were measured by ELISA, according to the manufacturer’s instructions and read at Biocell HT1 ELISA microplate reader (TSZ Company, Minneapolis, USA).
Co-culture and suppression assay
About 2 × 105 cells of CFSE-labeled CD4+CD25−T cells and CD4+CD25+Tregs were co-cultured in two panels in a 96-well culture plate in 1:4 ratio for 72 h at 37°C and 5% CO2. RIL-37 was added to only one panel of co-culture plate, at 24 h prior to the suppression assay. After stimulation, cultured CD4+CD25+ Tregs were used for suppression assay. After 72 h of co-culture, cells were collected and analyzed by flow cytometry to visualize CFSE dilution. Proliferation of CFSE-labeled responders was measured by the percentage of CFSE diluting responder cells.
Statistical analysis
Using SPSS statistical package (SPSS Inc., Chicago, IL, USA), the following statistical tests were used: Student’s t-test, analysis of variance, and a Mann–Whitney U test. A Pearson’s chi-square test was used for categorical variables. All data were presented as mean ± standard deviations (SD) (standard deviation of mean) and medians (for abnormally distributed parameters). They were compared with the tabulated probability value (P value) at the 0.05 level. The P value was considered significant if it is 0.05 or less. Correlations between two quantitative variables were assessed using Pearson’s coefficient and Spearman’s rho.
Results
Subjects’ characteristics
All subjects’ characteristics are outlined in Table 1. Patients were grouped according to Rutherford classification into four groups (stages III–VI). Regarding comorbidities, one-third of the patients had a diagnosis of type II diabetes (34.52%), 51.19% of patients had arterial hypertension, and 29.76% were with concomitant CAD.
Subjects’ characteristics.
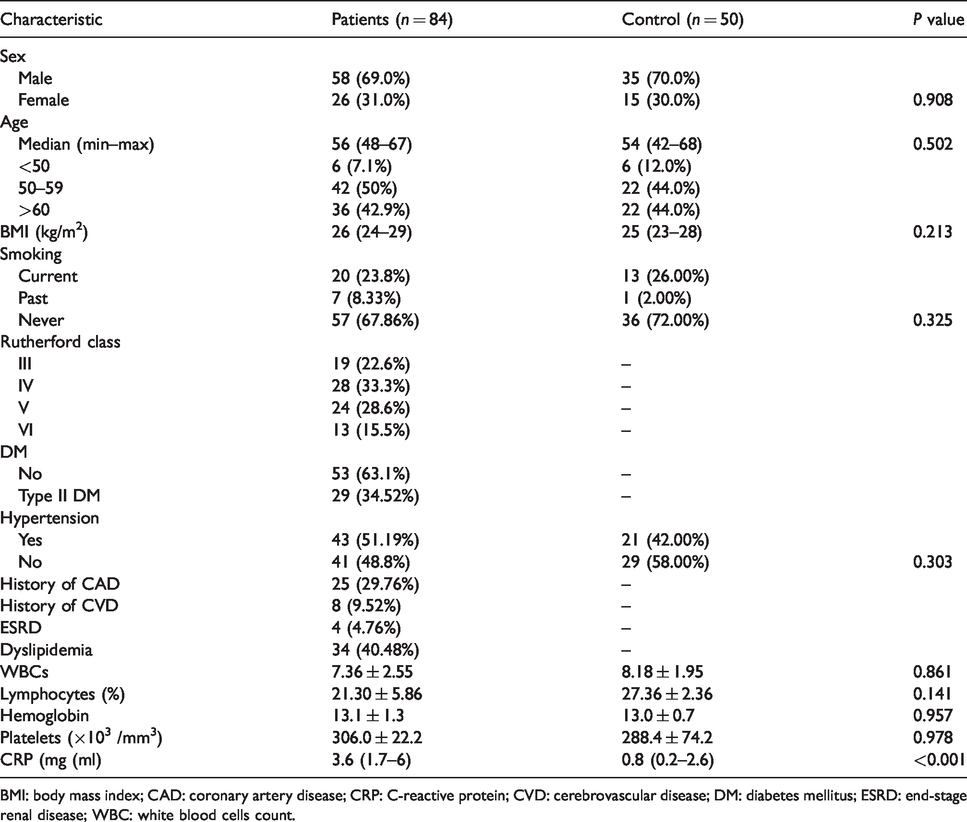
BMI: body mass index; CAD: coronary artery disease; CRP: C-reactive protein; CVD: cerebrovascular disease; DM: diabetes mellitus; ESRD: end-stage renal disease; WBC: white blood cells count.
Increased circulating IL-37 gene expression and levels in atherosclerotic patients
To examine the involvement of IL-37 in atherosclerosis, we considered measurements of IL-37 levels and mRNA expression in peripheral blood. We found that IL-37 mRNA expression in chronic lower limb ischemia patients was [18.5 (14–23.4)]-fold higher than those in control [7.4 (5.2–11.6)] (Figure 4). Simultaneously, IL-37 protein level in the serum of chronic lower limb ischemia patients was obviously increased compared with those in controls [(median= 1498.8 (1021.3–1976.3 pg/ml)] versus [median = 712.3 (532.5–864.3 pg/ml); P = 0.01] (Figure 5).

Comparison between atherosclerotic patients and control groups in gene expression of circulating (left) and arterial (right) IL-37 (folds): Data represented as means ± SDs and are summarized as dot plot. Each dot represents the mRNA expression of IL-37 for one individual. Right panel shows IL-37 folds expressed in arterial samples and left panel shows IL-37 folds in peripheral blood. The level of significance was set at *p ≤0.05.
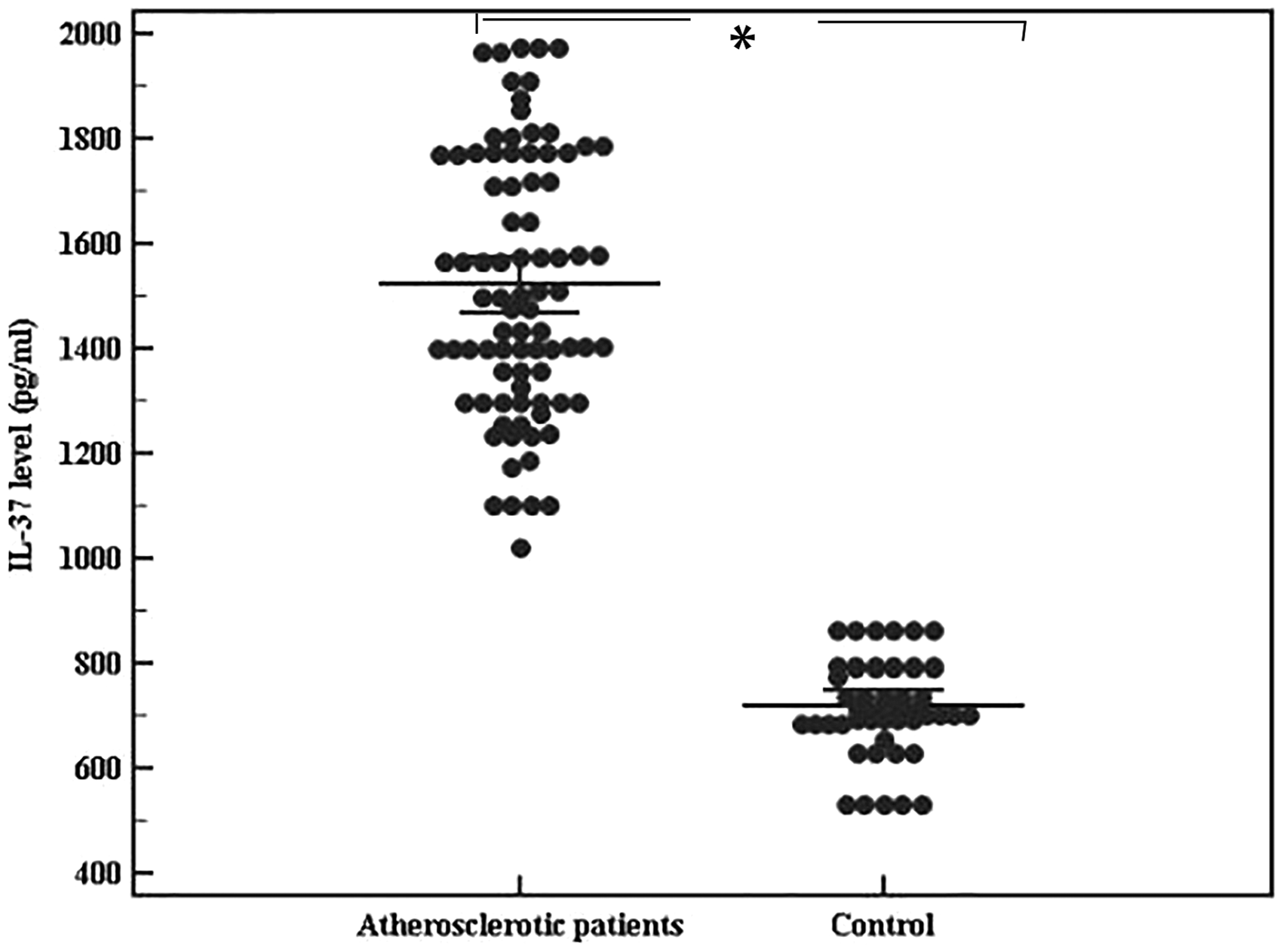
Comparison between atherosclerotic patients and control groups in circulating IL-37 level (pg/ml): Data represented as means ± SDs and are summarized as dot plot. Each dot represents the serum level of IL-37 for one individual. The level of significance was set at *p ≤0.05.
Overexpression of IL-37 in atherosclerotic arteries
We used real-time polymerase chain reaction to detect gene expression of IL-37 in arterial samples. As shown in Figure 4 (right), amplified IL-37 was detected in atherosclerotic arterial samples [20.4 (15.3–23.4 folds)]. On the other hand, only a slight amount of IL-37 was detected in normal arteries with a statistically significant difference [2.8 (1.9–6.8 folds)]. So, in this study, we demonstrate increased levels of IL-37 and increased expression of IL-37 mRNA, both systemically and within the lesion of patients with atherosclerotic disorder.
Circulating IL-37 mRNAs and serum protein levels were higher in patients with severe disease
We next investigated whether IL-37 was related to disease severity in PAOD patients as categorized by Rutherford’s classification. As seen in Figure 6, significant differences were observed in IL-37 mRNAs and protein levels between patients with higher stages (stage V + VI = 37 patients) who expressed more IL-37 mRNA in peripheral blood [19.4 (18.3–23.4 folds)] and those with less severe disease stages (stage III + IV = 47 patients) with almost [16.4 (14–20.5 folds); P <0.001]. Additionally, serum protein levels of IL-37 were [1774.5 (1510.7–1976.3 pg/ml)] versus [1356.3(1021.3–1574.9 pg/ml)] with the highest level in stage VI [1909.7 (1804.5–1976.3 pg/ml)] (Figure 7).

Comparison between (stage III + IV) and (stage V + VI) in gene expression of circulating IL-37 and arterial IL-37 (folds): Data represented as means ± SDs and are summarized as dot plot. Each dot represents the mRNA expression of IL-37 for one individual. The level of significance was set at *p ≤0.05.

Comparison between (stage III + IV) and (stage V + VI) in circulating IL-37 level (pg/ml): Data represented as means ± SDs and are summarized as dot plot. Each dot represents the serum level of IL-37 for one individual. The level of significance was set at *p ≤0.05.
Localized IL-37 expression does not correlate to disease severity
On the other hand, we did not observe a statistically significant difference in the expression of IL-37 mRNAs in the arterial samples between patients with (stage III+IV) = [20.3 (15.3–23.4 folds)] and patients with (stage V+VI) = [20.5 (18.4–23.4 folds); P =0.091] (Figure 6 (Right)).
Association between serum IL-37 protein levels and clinical comorbidities and laboratory values
In order to further assess the relationship between serum IL-37 protein levels and associated clinical features and laboratory values, serum IL-37 protein levels were compared among patients with and without certain concomitant diseases. We found no significant differences in serum IL-37 protein levels between patients with medical hypertension, diabetes mellitus, dyslipidemia, end-stage renal disease, and those with history of coronary or cerebrovascular ischemia, and patients without the above-mentioned clinical comorbidities (data not shown). In addition, we identified that neither smoking nor body mass index (BMI) had an association with IL-37 serum levels.
Our data suggested that serum IL-37 levels were negatively correlated with C-reactive proteins (CRP) (mg/ml), P = 0.048, but it lacked association with other laboratory values including hemoglobin (g/dl), white blood cell count (WBC), lymphocytes (%), and platelet count.
Correlation between circulating IL-37 level and levels of IL-10 and TGF-β
Serum levels of IL-10 and TGF-β were significantly lower in patients than in controls (data not shown) (P = 0.033 and 0.01, respectively). Results also show that levels of IL-10 released in cultured supernatants before stimulation with rIL-37 were lower in patients with chronic lower limb ischemia [median= 266.4 (198.4–299 pg/ml)] than in controls [309.4 (266.4–409.4 pg/ml); P <0.001]. In addition, TGF-β was shown to be largely reduced in cultures from patients [486.5 (299.4–582.6 pg/ml)] than in cultures of cells from healthy controls [712.3 (532.5–864.3 pg/ml)].
Interestingly, a significant direct correlation with circulating serum IL-37 level was found between IL-37 serum levels and TGF-β in cultures, P < 0.001, with no significant correlation with spontaneously released IL-10 in culture supernatants, P = 0.247 (Figures 8 and 9).
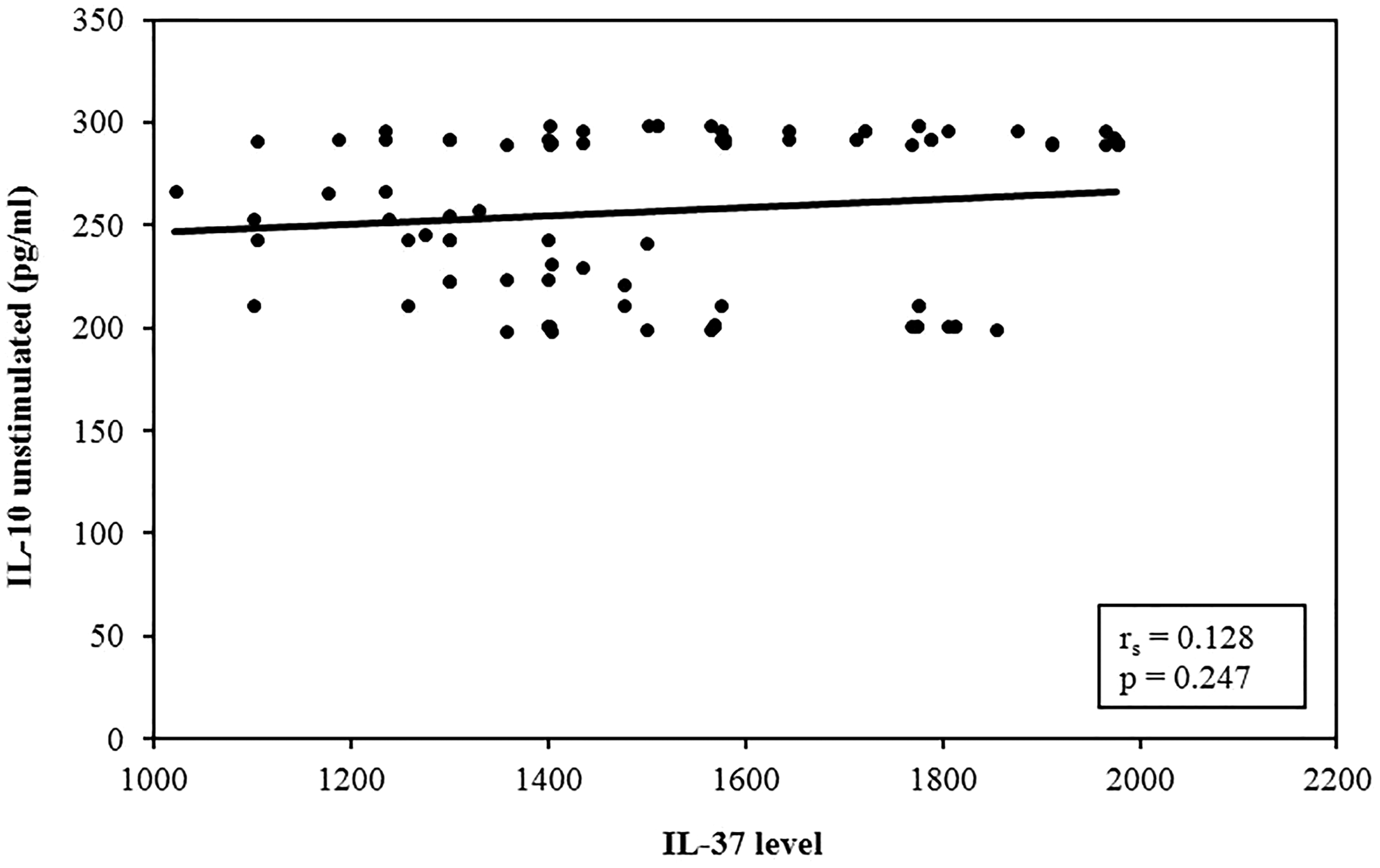
Correlation analysis between IL-37 level versus unstimulated IL-10 in atherosclerotic patients group (n = 84). Correlation was conducted by Spearman’s coefficient. The level of significance was set at *p ≤0.05.
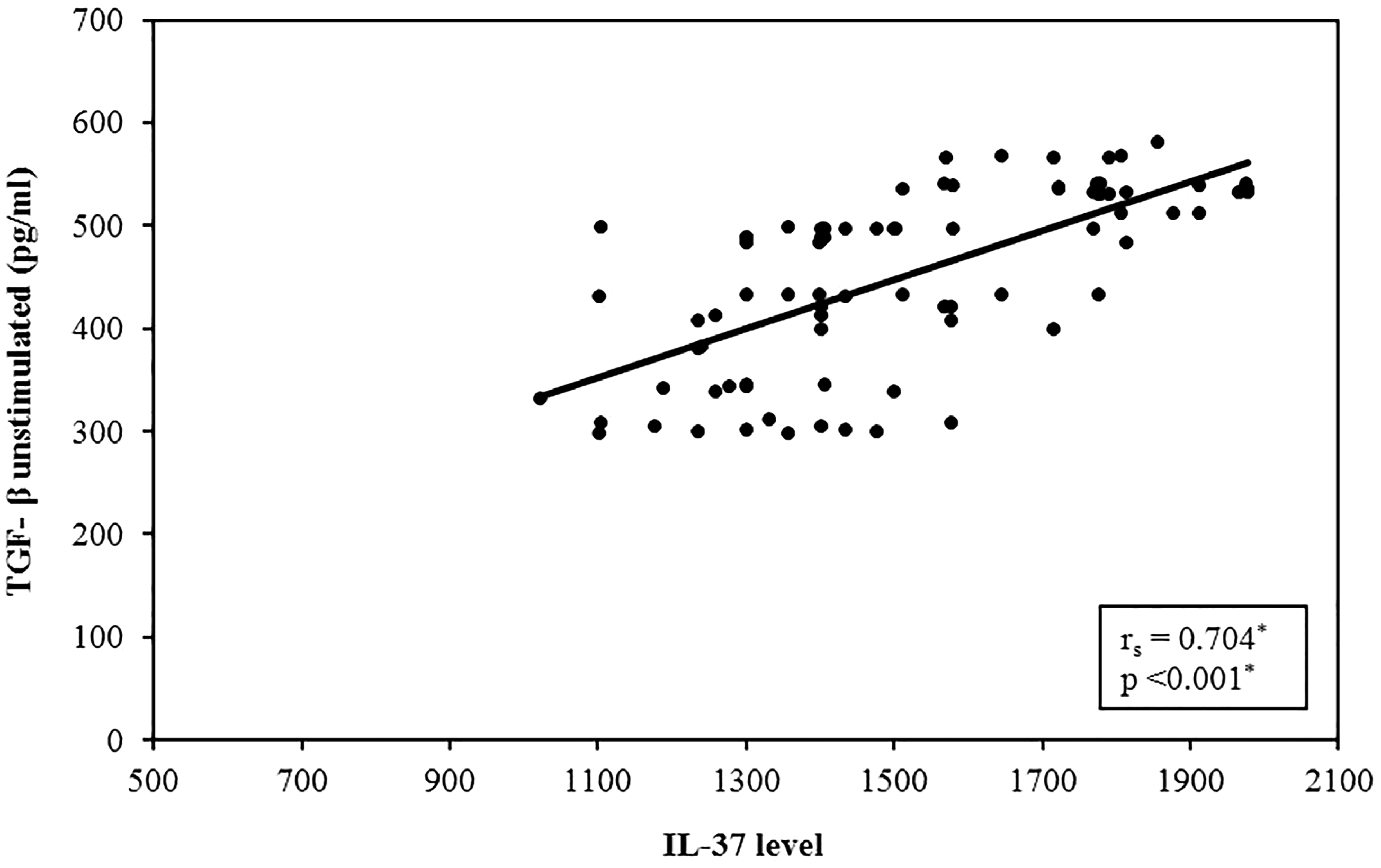
Correlation analysis between IL-37 level versus unstimulated TGF-β in atherosclerotic patients group (n = 84). Correlation was conducted by Spearman’s coefficient. The level of significance was set at *p ≤0.05.
Culture with recombinant IL-37 modulated IL-10 and TGF-β release from PBMCs of atherosclerotic patients
In agreement with previous results showing upregulation of IL-10 in the serum of IL-37-treated mice, 25 data from the present study indicated that stimulation with rIL-37 in human PBMCs markedly increased the released IL-10 in cultures from atherosclerotic patient group, P <0.001 (Figure 10), indicating that IL-37 not only suppresses expression of inflammatory mediators but also promotes the production of anti-inflammatory mediators. However, this significant increase in IL-10 was not seen in the controls.
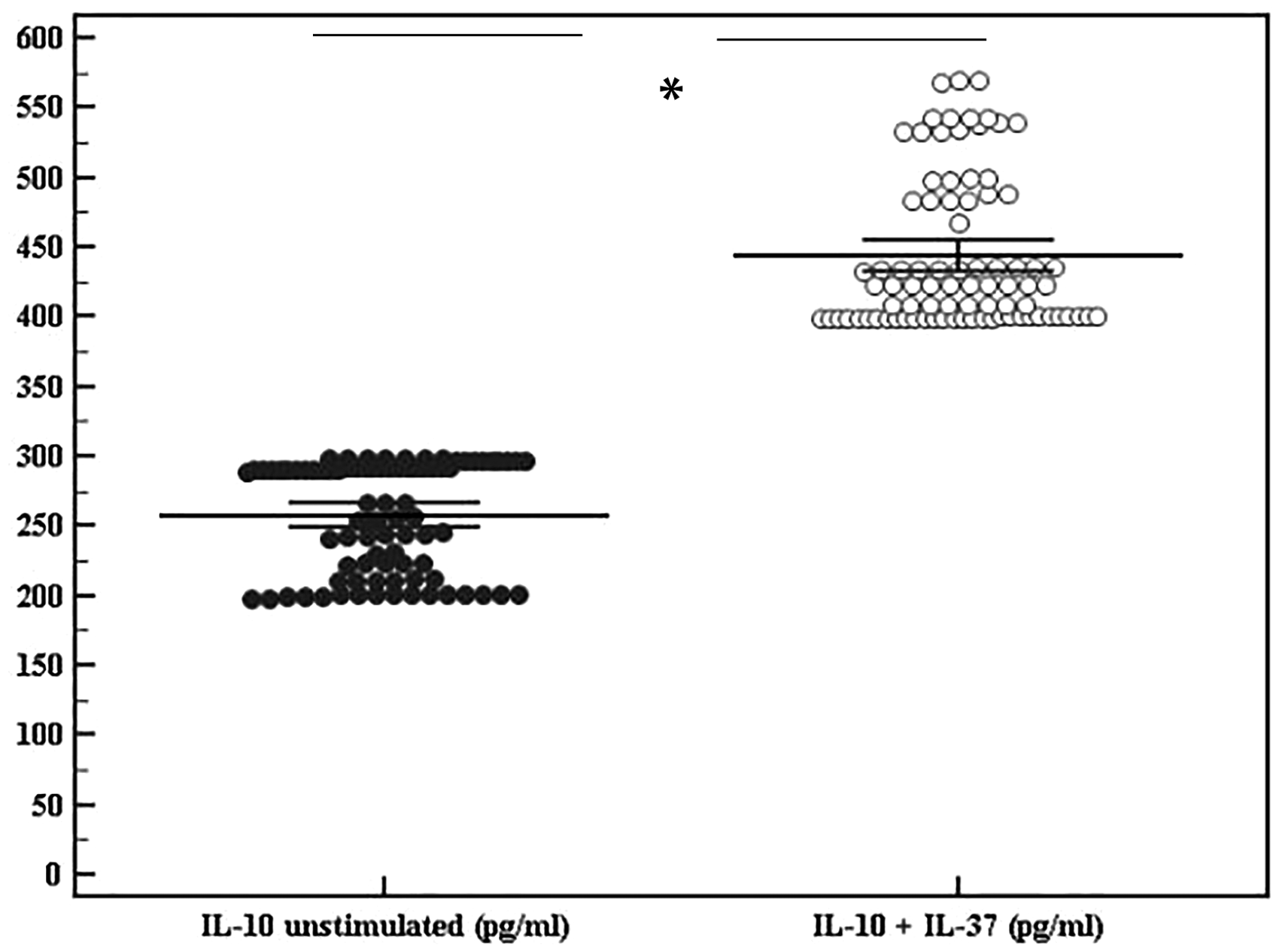
rIL-37 effect on IL-10 release. Data represented as means ± SDs and are summarized as dot plot. Each dot represents the serum level of IL-10 for one individual. Left dot plot shows unstimulated IL-10 (pg/ml) in atherosclerotic patients. Right dot plot shows cytokine level after addition of rIL-37. The level of significance was set at *p ≤0.05.
Likewise, levels of secreted TGF-β in cultures from patients were significantly increased after stimulation with rIL-37 and with a significant difference between patients and controls (Figure 11).

rIL-37 effect on TGF-β release. Data represented as means ± SDs and are summarized as dot plot. Each dot represents the serum level of TGF-β for one individual. Left dot plot shows unstimulated TGF-β (pg/ml) in atherosclerotic patients. Right dot plot shows cytokine level after addition of rIL-37. The level of significance was set at *p ≤0.05.
Correlation of IL-10 and TGF-β release to disease severity
Neither unstimulated IL-10 levels in culture supernatants nor the rIL-37 stimulated levels were correlated to disease stages (P = 0.345, 0.477 respectively). Conversely, TGF-β showed a significant relation to disease severity before stimulation with rIL-37, P < 0.001. However, after the addition of rIL-37 to cultures of Treg cells, a non-significant difference in the augmentation of the level was recorded between different disease stages, P= 0.146. Almost same results were obtained when we grouped stages together (Table 2).
Comparison between stages according to IL-10 and TGF levels in cultures (n = 84).

P represents p value for comparing between different stages; P1 represents p value for comparing between stages (III + IV) and (V + VI).
*Statistically significant at p ≤ 0.05.
IL: interleukin; TGF-β: transforming growth factor-beta.
The role of rIL-37 in enhancing Treg direct immunosuppressive ability in vitro
It was noteworthy that the percentages of CD4+CD25+Tregs in peripheral blood were found to be significantly reduced in chronic lower limb ischemia patients (4.02% ± 0.57%) than in control subjects (6.63% ± 1.01%; P < 0.001) (Figure 12).
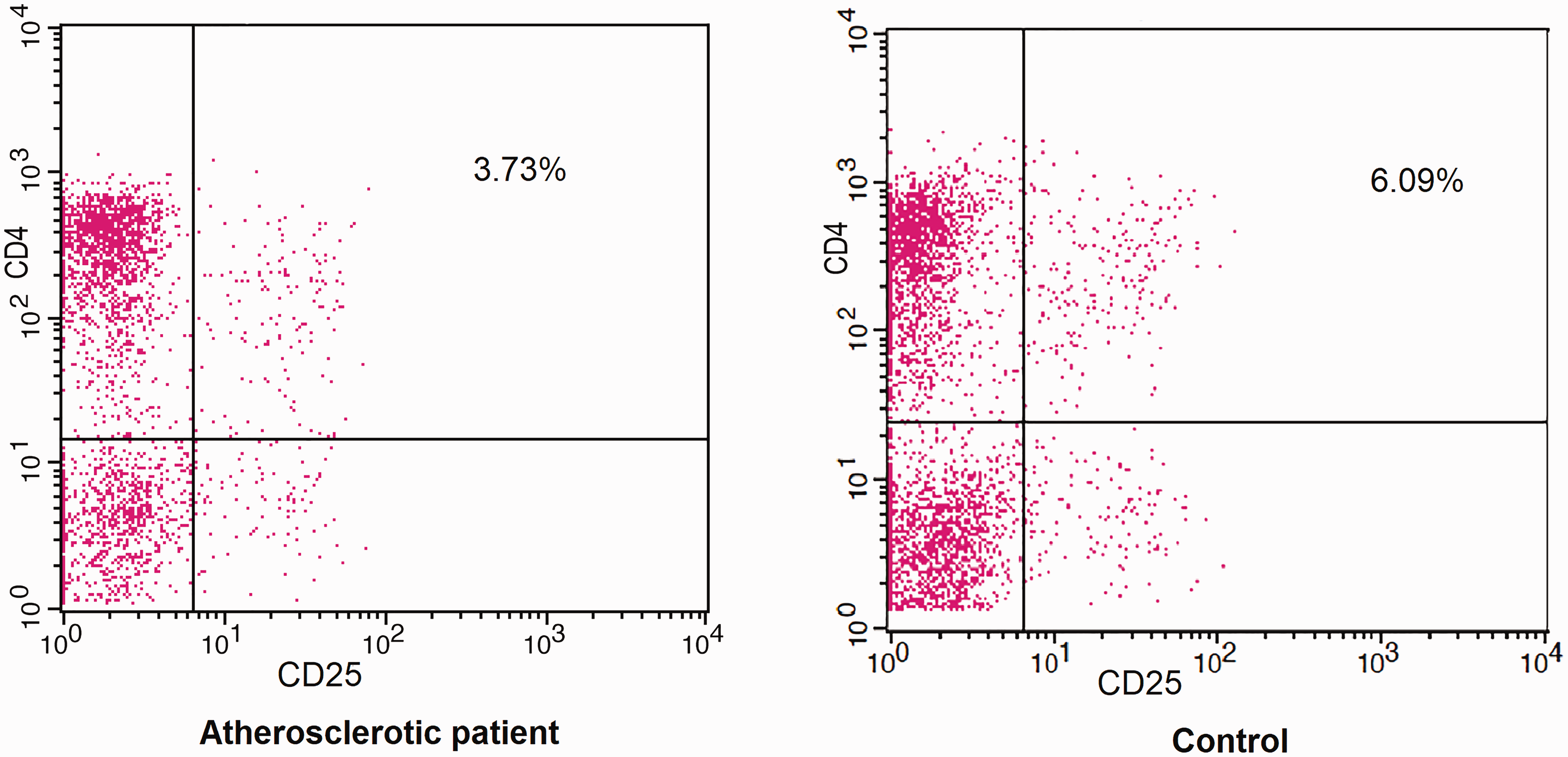
Percentages of CD4+CD25+ (Treg) cells from atherosclerotic patients and controls: Isolated CD3 T cells from an atherosclerotic patient were stained with anti-CD4 and anti-CD25 antibodies representing Treg cells in an atherosclerotic patient and non-atherosclerotic control subject.
To determine if rIL-37 modifies the suppressive activity of Tregs, we purified CD4+CD25+Tregs from patients’ PBMCs, and then we co-cultured them separately with autologous responder CD4+CD25−T cells at the 1:4 ratio in the presence of CD3/CD28 antibodies stimulation for 72 h. We further assessed proliferation of T cells using the extent of CFSE dilution of labeled autologous T cells (hereafter called responder cells). Results of CFSE staining demonstrated that co-culture of purified CD4+CD25+Tregs from patients’ blood and responder CD4+CD25−T cells showed a non-significant reduced capacity of CD4+CD25−T cells proliferation [(82.8% ± 8.3% with Treg) vs (96.2% ± 2.1% without Treg)]. By comparison, a significantly decreased proliferation of CD4+CD25−T cells was recorded under stimulation of rIL-37 (43 ± 6.5%; P < 0.05) (Figure 13). This indicates that IL-37 might be involved in the function of Tregs to suppress T cells in atherosclerosis, hence stimulating the immune suppressive capacity of Tregs. A significant decrease of responder T cell proliferation was seen in stage VI (30.5% ± 3.2%).
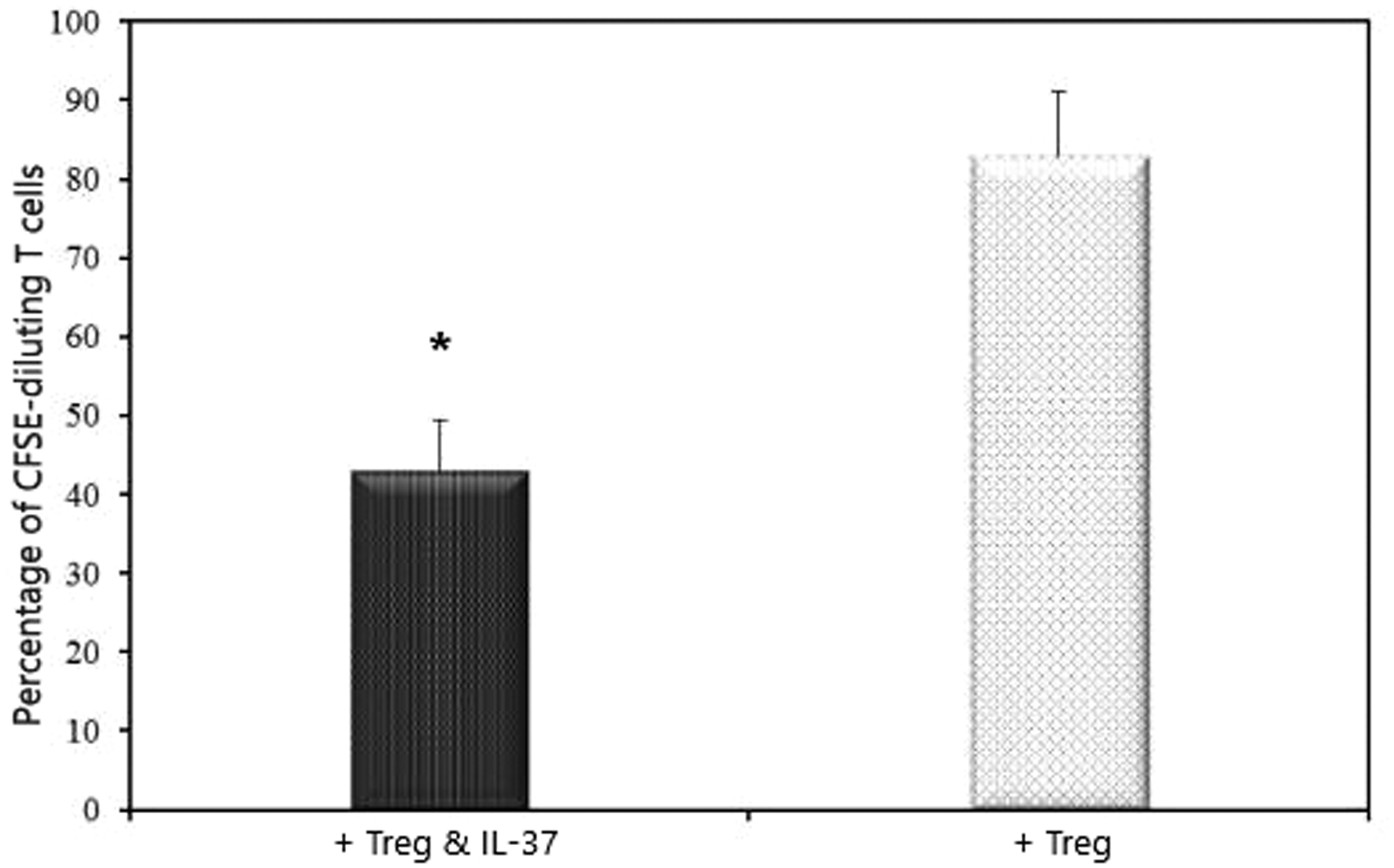
rIL-37 effect on the suppressive activity of CD4+CD25+ Tregs. CD4+CD25+ Tregs were mixed with CFSE-labeled CD4+CD25−T cells (responder T cells), and the proliferation of responder T cells was assessed by the percentage of CFSE diluting responder T cells. n = 84.
Discussion
Although mRNA expression and serum protein level of IL-37 have been found to be increased in a number of chronic inflammatory conditions, 26 , 27 as well as in patients with acute coronary syndrome 19 , 28 and atrial fibrillation, 29 however, data regarding their association with peripheral atherosclerosis in humans and their correlation to disease severity of chronic lower limb ischemia are lacking.
IL-37 may attenuate atherosclerosis, given that it is a critical regulator in the inflammatory response. 30 It was found to inhibit dendritic cells as well as IL-18-induced inflammatory response. 31 Since the first report of IL-37 in 2000, 9 many studies have tried to unravel the anti-inflammatory effects of endogenous IL-37 as the increased synthesis of pro-inflammatory cytokines was seen upon downregulation of IL-37 in vitro. 31 Also, IL-37 has been shown to inhibit the production of pro-inflammatory cytokines: TNF-α, IL-1, and IL-6 in PBMCs in vitro. 11 However, the potential significances and underlying mechanisms of IL-37 in mediating immune suppressive function of Tregs are poorly known.
Our results provide a detailed analysis of IL-37 expression in chronic lower limb ischemia patients (Rutherford stages III–VI) and healthy controls. We demonstrated that IL-37 mRNAs expressions and serum protein levels were higher in patients with peripheral atherosclerotic ischemia than in non-atherosclerotic subjects. This could be explained by findings from Boraschi et al. 10 who suggested involvement of IL-37 in the development of atherosclerotic disease as it was expressed in the foam-like cells of atherosclerotic coronary and carotid artery plaques. This could imply that IL-37 is associated with the macrophage activation and the shift in macrophages to foam cells in atherosclerosis. 10
Alternatively, activated macrophages also act as antigen presenting cells to initiate T lymphocyte activation, and evidence has demonstrated that regulating the activation of macrophages is the main mechanism by which IL-37 controls inflammation and attenuates autoimmune and inflammatory diseases including murine model of myocardial ischemia/reperfusion (I/R) injury and colitis.32,33 Likewise, recent murine studies showed that IL-37 treatment reduced atherosclerosis development in mice, via reduction of macrophage content and increasing collagen and smooth muscle cell content. 20 Also, hematopoietic expression of IL-37 in atherosclerosis-prone LdLr-deficient mice reduced the circulating immune cells in vivo and the secretion of pro-inflammatory cytokines by peritoneal macrophages ex vivo; however, this reduced inflammatory state was not accompanied by reduced atherosclerosis development or reduced macrophage content of the lesions. 18
Further investigation in the current study revealed that IL-37 mRNAs and protein levels were not significantly higher in localized arterial tissues of patients with severe disease than in patients with mild to moderate severity, although a significant difference was seen in circulating levels. Alternatively, levels of anti-inflammatory cytokines were significantly lower in severe stages suggesting that a higher arterial expression of IL-37 may be required for achieving its atheroprotective effects in severe stages. This implies that there might be confounding factors that impede the circulating levels of anti-inflammatory IL-37 in severe stages, with the beneficial potential anti-inflammatory properties, to extend to the diseased arteries and decelerate the progression of inflammation. Taking this into consideration, we suggest the existence of other local factors that need to be further investigated.
IL-10 and TGF-β are the two cytokines that play the critical immunosuppressive role of Tregs. Although they are secreted by other cells besides Treg, Treg secretes the largest amount of them. Furthermore, both cytokines have been confirmed to exert anti-atherogenic effects.34–36 In the present study, the serum levels of IL-10 and TGF-β, as well as both IL-10 and TGF-β secreted in the supernatants of Tregs cultures, were significantly of a reduced amount in PBMCs of chronic lower limb ischemia patients than in PBMCs from healthy controls. Both IL-10 and TGF-β secreted levels, which were assessed in supernatants of Tregs as recombinant purified IL-37 stimulated, were significantly increased as to almost their release under normal condition. This result is not in agreement with the findings of Ye et al., 26 where rIL-37 inhibited pro-inflammatory cytokines in SLE patients with no effect on IL-10 or TGF-β production. 26
It was hypothesized that exogenous administration of IL-37 could not perform the anti-inflammatory powerful functions of endogenous IL-37 that was likely intracellular. 33 However, a recent report has revealed an expansion of IL-37 to include both intracellular and extracellular pathways involving the transcription factor Smad3 and binding to the orphan receptor IL-1R8, respectively. 16 Also, a previous animal study reported that exogenous IL-37 promoted a significant increase in the production of anti-inflammatory cytokines in myocardial ischemia model mice. 32 Moreover, in mice with I/R injury, reperfusion with rIL-37 protein has been found to decrease damage and recover organ function. 37 It also reduced infarct size, TLR-4 expression, and neutrophil infiltration in a mouse model of cardiac I/R injury. 25
Interestingly, the anti-inflammatory cytokines production in PBMCs of healthy controls in our study was unaffected by rIL-37, suggesting that anti-inflammatory actions of IL-37 are present only in the inflammatory disease conditions of atherosclerosis.
Consistent with these findings, the present study demonstrates that co-cultures of Tregs and effector T cells showed a significantly reduced proliferative capacity of effector T cells when rIL-37 was added. Recombinant IL-37 markedly amplified Tregs’ suppressive ability, which indicates that IL-37 might be involved in the function of Tregs to suppress T cells, thus promoting the immune suppressive capacity of Tregs. This could be attributed to its ability to modulate CTLA-4 levels on these cells, which is closely related to the development and function of Tregs. 38 In a mouse model of myocardial infarction, adoptive transfer of IL-37 significantly increased the number of Tregs and improved cardiac function. 39 This is in seeming contrast to another study, in which IL-37 expression did not influence the number of Tregs. 18
Levels of IL-37 in our study were directly correlated to TGF-β in chronic lower ischemia patients. It was shown that IL-37 expression is induced by TGF-β, which is dependent on the transcription factor Smad3. 40 IL-37 can also be upregulated by IL-10, and a positive feedback loop was suggested between IL-37 and IL-10. 11 However, blocking IL-10 signaling and not TGF-β significantly abolished the protective effects of IL-37 in a myocardial I/R injury mice model. 32
Taken together, whether IL-37 plays a protective role in atherosclerosis and what is the exact mechanism is still unknown. We propose at least two mechanisms that could explain the atheroprotective role of IL-37 in atherosclerosis: (1) at the molecular level, IL-37 ameliorates atherosclerosis via stimulation of the secretion of anti-inflammatory cytokines and (2) at the cellular level, IL-37 attenuates atherosclerosis through the stimulation of Treg cell to cell suppressive ability.
In summary, our study is the first to reveal an increased expression of IL-37 in peripheral atherosclerotic lower limb ischemia both peripherally and in the arterial lesions. The arterial expressed IL-37 did not correlate to disease stage and was expressed almost similarly in patients with different severity, while the circulating levels of this cytokine correlated well to disease stage. In addition, serum IL-37 levels were found to have no correlation to associated comorbidities, smoking, BMI, and selected laboratory parameters including Hb level, WBCs, lymphocytes (%), and platelet count. On the other hand, a significant direct correlation with TGF-β and a negative correlation with CRP were found. The study also revealed that recombinant IL-37 can promote the immune suppressive effect on T lymphocytes in association to Treg activation and could effectively augment the production of anti-inflammatory cytokines in supernatants of lymphocyte cultures from humans with peripheral atherosclerotic disease. We are convinced that one of the main targets for the development of new therapeutics for atherosclerotic disease might be the management of the inflammatory reaction, which is considered as the hallmark in development and progression of atherosclerosis, and that an in-depth study of IL-37 in atherosclerosis not only expands our understanding of the nature of atherosclerosis but also provides a novel therapeutic approach in atherosclerotic diseases in the future.
Conclusions
In conclusion, findings of the present study (summarized in Supplement 1) indicate that supplementation with exogenous IL-37 amplified levels of released anti-inflammatory cytokines: IL-10 and TGF-β secreted by Tregs in peripheral atherosclerotic patients in vitro. Results suggest a role for recombinant IL-37 in mediating anti-inflammatory functions in the atherosclerotic process, potentially involving enhancement of Treg inhibitory function and anti-inflammatory cytokine secretion with a particularly marked direct response in severe disease. Furthermore, the recruitment of the relatively high circulating IL-37 in severe stages of atherosclerotic disease to arterial lesions could be a target of modulation thereafter. Nevertheless, more investigation is required to study the precise mechanism underlying IL-37 on Tregs and the potential effect associated with host immune response in vivo.
Supplemental Material
sj-pdf-1-vas-10.1177_1708538120921735 - Supplemental material for The novel role of IL-37 to enhance the anti-inflammatory response of regulatory T cells in patients with peripheral atherosclerosis
Supplemental material, sj-pdf-1-vas-10.1177_1708538120921735 for The novel role of IL-37 to enhance the anti-inflammatory response of regulatory T cells in patients with peripheral atherosclerosis by Hassan Lotfy, Marwa Moaaz and Mai Moaaz in Vascular
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.