
Abstract
Objectives
Accumulating evidence suggests that ursodeoxycholic acid (UDCA) has therapeutic potential for osteoarthritis (OA); however, the underlying molecular mechanisms remain poorly understood.
Methods
The chemical components of UDCA were retrieved from PubChem. Potential UDCA targets were collected from three databases. OA-related targets were obtained from GeneCards, OMIM (Online Mendelian Inheritance in Man), and TTD (Therapeutic Target Database) databases. Venny 2.1.0 was used to identify common targets between OA and UDCA. A protein-protein interaction network was constructed using the STRING database and subsequently analyzed using Cytoscape software to identify core genes. Gene Ontology and Kyoto Encyclopedia of Genes and Genomes pathway enrichment analyses were performed using DAVID. Molecular docking validation was conducted using AutoDock software, followed by molecular dynamics simulations using Amber to assess stability of interactions between UDCA and key targets.
Results
194 common targets associated with UDCA and OA were identified. Among these, Albumin (ALB), Epidermal growth factor receptor (EGFR), Matrix metalloproteinase-9 (MMP9), Proto-oncogene tyrosine-protein kinase Src (SRC), Caspase-3 (CASP3), and Estrogen receptor (ESR1) were identified as core targets. UDCA exerts its therapeutic effects on OA primarily through the modulation of protein hydrolysis, negative regulation of apoptosis, and promotion of cell proliferation and differentiation, as well as by influencing the PI3K-Akt, MAPK, and cancer-related signaling pathways. Molecular docking and molecular dynamics simulations demonstrated that UDCA exhibits good binding affinity and forms stable interactions with multiple core targets.
Conclusion
By targeting molecules such as ALB, EGFR, MMP9, SRC, CASP3, and ESR1, UDCA may interfere with the PI3K-Akt, MAPK, and other signaling pathways, thereby regulating the expression of inflammatory mediators, chondrocyte apoptosis, and cartilage matrix degradation and synthesis, thus contributing to OA treatment.
Keywords
1. Introduction
Osteoarthritis (OA) is a chronic joint condition characterized by progressive cartilage degeneration, synovial membrane inflammation, and osteophyte formation. It primarily affects the older population. Among people aged > 60 years, the prevalence of OA is 10% in men and 18% in women. 1 Given the global trend of aging populations and increasing obesity rates, the burden of OA is anticipated to increase significantly in the coming decades. If OA is not managed properly, it can lead to progressive functional limitations, disabilities, and various musculoskeletal complications. 2 These effects significantly diminish patient quality of life and impose a substantial financial burden on individuals and the national healthcare system. Traditional treatments, such as NSAIDs and joint injections alleviate symptoms but do not halt or reverse cartilage degeneration and may lead to side effects, including gastrointestinal issues and infection risks. In recent years, numerous emerging treatment approaches and pharmacological agents have been developed, including mesenchymal stem cell (MSC) therapy,3,4 platelet-rich plasma (PRP) therapy,5,6 gene therapy, 7 and the application of nanotechnology and innovative biomaterials.8,9 Furthermore, novel drugs targeting Bone Morphogenetic Protein-7 (BMP-7), Fibroblast Growth Factor-18 (FGF-18), Human Serum Albumin (HSA), Interleukin-1 (IL-1) inhibitors, and Beta-nerve growth factor (β-NGF) antibodies have also emerged. 10 However, most of these emerging drugs and therapies are still in the clinical trial stage; their efficacy remains uncertain, and they are extremely expensive, making them unaffordable to the general public.
Ursodeoxycholic acid (UDCA) is widely used to treat liver and gallbladder disorders. Moreover, it has exhibited potential in non-liver-related conditions such as cardiovascular diseases and organ transplantation. Recent studies have demonstrated that UDCA exerts multiple therapeutic effects, including alleviation of endoplasmic reticulum stress, promotion of chondrocyte proliferation, and anti-inflammatory and anti-apoptotic activities.11–15 Specifically, UDCA can inhibit the degradation of the cartilage extracellular matrix (ECM), ameliorate cartilage degeneration, and retard osteoarthritis progression.16–18 Therefore, UDCA is a potential therapeutic option for the treatment of OA. However, the specific molecular mechanisms underlying the protective effects of UDCA against OA remain unclear. Using network pharmacology, molecular docking, and molecular dynamics simulations, we explored the molecular mechanisms underlying the therapeutic effects of UDCA on OA.
2. Materials and methods
2.1. Screening of OA-related disease targets
Genes potentially associated with OA were identified and filtered from three publicly accessible databases: GeneCards 19 (https://www.genecards.org/), Online Mendelian Inheritance in Man 20 (OMIM, https://www.omim.org/), and the Therapeutic Target Database 21 (TTD, https://db.idrblab.net/ttd/), all accessed on August 17, 2025. These databases are frequently utilized in target identification research because of their extensive gene/disease annotations and disease-specific therapeutic target information. Searches were conducted using the search term “osteoarthritis” and restricting the organism filter to “Homo sapiens”. After duplicates were removed and the data were integrated, a distinct list of OA-associated genes was compiled.
2.2. Screening of UDCA-related targets
2D and 3D structures of UDCA were obtained from PubChem 22 (https://pubchem.ncbi.nlm.nih.gov/; CID: 31401). On August 17, 2025, potential UDCA targets were identified using three complementary databases: PharmMapper 23 (https://www.lilab-ecust.cn/pharmmapper/index.html) and SwissTargetPrediction 24 (https://www.swisstargetprediction.ch/), which facilitate computational target prediction based on pharmacophore and structural similarity, respectively, and DrugBank 25 (v.6.0; https://go.drugbank.com/), known for its complementary predictive methodologies and established credibility in drug target identification. Subsequently, a screening process was implemented: only those targets annotated for the organism “Homo sapiens” were selected. After integrating the results and removing duplicate entries, UDCA-related target genes were compiled. The targets were then merged with OA-related genes and analyzed for overlap using the Venn diagram tool 26 (v.2.1.0; https://bioinfogp.cnb.csic.es/tools/venny/).
2.3. Protein-protein interaction (PPI) network construction and identification of key target genes
The STRING database 27 (v.12.0; https://cn.string-db.org/), accessed on August 17, 2025, was used to construct a PPI network by uploading overlapping target genes of UDCA and OA. An in-depth analysis was conducted using Cytoscape 28 (v.3.10.4; https://cytoscape.org/) software to identify core target genes.
2.4. Gene ontology (GO) and kyoto encyclopedia of genes and genomes (KEGG) pathway analysis
The DAVID database 29 (v.6.8; https://davidbioinformatics.nih.gov/) was used (on August 17, 2025) to conduct GO and KEGG pathway enrichment analyses. The analyses sought to identify overrepresented categories in biological processes (BP), cellular components (CC), molecular function (MF), and significantly enriched KEGG pathways. The results were visualized using the bioinformatics tool 30 available at https://www.bioinformatics.com.cn, and an “enriched pathway-key target” network was constructed using Cytoscape software.
2.5. Molecular docking and visualization
The core target proteins identified in from the PPI network were subjected to molecular docking analysis. 3D structures of the target proteins were sourced from the RCSB PDB database 31 (https://www.rcsb.org/) (on August 17, 2025). These structures were selected due to their representation of high-resolution complexes (<2.5 Å) with known co-crystallized ligands, thus ensuring the reliability of active site geometry during docking procedures. Water molecules and non-essential ligands were excluded, polar hydrogen atoms were incorporated, Kollman partial charges were assigned, and non-polar hydrogen atoms were combined using AutoDock Tools 32 (v. 4.2.6; https://autodock.scripps.edu/). The 3D structure of UDCA was acquired from PubChem with the addition of hydrogen atoms and Gasteiger charges were assigned using AutoDock Tools. All molecular structures were stored in the PDBQT format. Blind docking simulations were performed using AutoDock Vina 33 (v.1.2.x; https://vina.scripps.edu/), with grid box dimensions and center coordinates specified in Supplemental Material 1, encompassing the entire protein surface. No specific amino acid residues were designated as active sites, which allowed the software to autonomously identify the global energy-optimal binding configuration. The grid point spacing for all systems was consistently set at 0.375 Å, the exhaustiveness parameter for docking was established at 12, and the number of resulting binding conformations was set to 9. The conformation with the lowest energy mode was chosen from among nine conformations. The binding affinity between UDCA and each target protein was evaluated based on the calculated binding energy, followed by comparative analysis using the co-crystallized ligands of each protein as controls. The binding modes were visualized using PyMOL 34 (v.2.5.7; https://pymol.org/) to illustrate molecular interactions during the docking process.
2.6. Molecular dynamics simulations
The complex interactions and dynamics of the system are captured by explicitly simulating each atom and chemical bond. Amber software 35 (v.24; https://ambermd.org/AmberMD.php) was used to assess the binding stability between the protein and the compound. The LEaP module was used to build a simulation system by loading the PDB structures of the wild-type protein and its mutants. The proteins were described using the ff14SB 36 force field and the compound was parameterized using the GAFF force field. The system was immersed in a TIP3P water box, maintaining at least a 10.0 Å gap between the protein surface and the box boundary. Charge neutralization was accomplished through the addition of Na+ ions, with the precise number of ions automatically computed and assigned using the ParmEd module within AmberTools 37 (v.25, https://ambermd.org/AmberTools.php). Following this, Na+ and Cl- ions were introduced to establish a physiological salt concentration of 0.15 M. A two-stage energy-minimization protocol was employed to resolve steric clashes and ensure structural stability. Initially, restraints were applied to the protein backbone while optimizing solvents and ions. Subsequently, these restraints were removed, allowing the entire system to undergo comprehensive minimization using the steepest descent and conjugate gradient algorithms. System equilibration involved two phases. First, the system was heated from a low temperature to 300.0 K under constant temperature conditions with backbone restraints. Subsequently, the system was equilibrated under constant pressure conditions to adjust solvent density, maintaining a temperature of 300.0 K and pressure of 1 bar in preparation for production simulations. A 100-ns molecular dynamics simulation was conducted under constant temperature and pressure conditions at 300 K and 1 bar, respectively, without positional restraints, to produce trajectories for analysis. CPPTRAJ was used to analyze trajectory data and calculate the root mean square deviation (RMSD), root mean square fluctuation (RMSF), hydrogen bond count, and principal component analysis (PCA). The binding free energies were estimated using the MM/GBSA method. Subsequently, per-residue decomposition of the binding free energy was performed to elucidate the contribution of individual amino acid residues to ligand binding. To compare the dynamic behavior of the selected ligand with that of a reference ligand, separate molecular dynamics simulations were performed for the reference ligand-protein complexes under identical simulation parameters.
3. Results
3.1. Collection of OA-related target genes
We identified 42 entries from OMIM, 5,205 from GeneCards, and 39 from TTD. Following the integration of these datasets and removal of duplicate targets, a non-redundant set of 5,258 OA-related target genes was established.
3.2. Collection of UDCA-related target genes and intersection with disease-associated genes
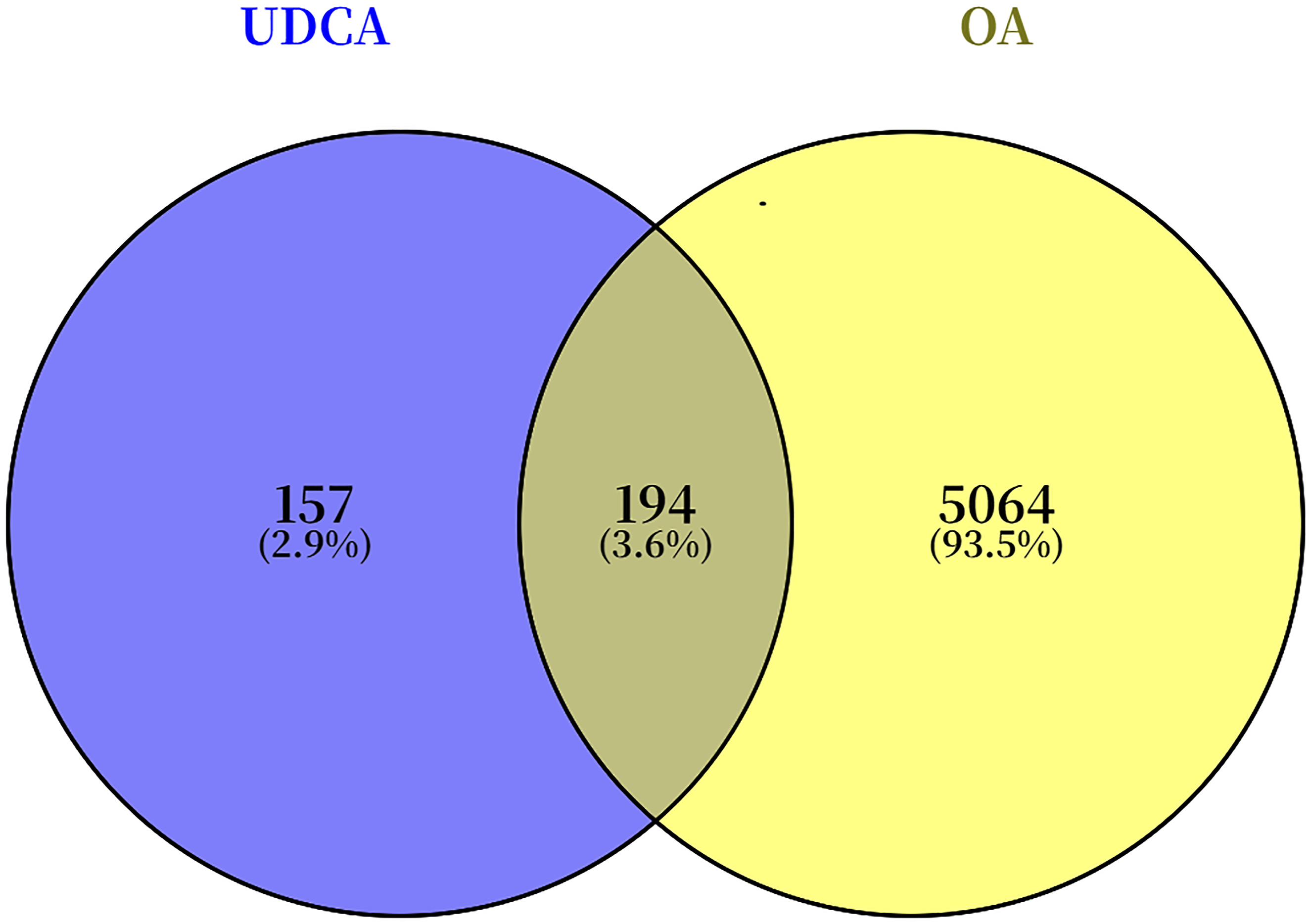
The chemical structure of UDCA in both 2D and 3D formats was obtained from PubChem (Figure 1). A total of 100, 4, and 290 UDCA-related target genes were identified from the SwissTargetPrediction, DrugBank, and PharmMapper databases, respectively. After integrating datasets and eliminating duplicates, a unique set of 351 UDCA-related target genes was identified. Common target genes of UDCA and OA were identified using the Venny analysis tool, resulting in 194 intersecting targets (Figure 2). Chemical structure of UDCA Venn diagram showing overlapping targets of UDCA and OA
3.3. Establishing PPI network and screening for essential genes
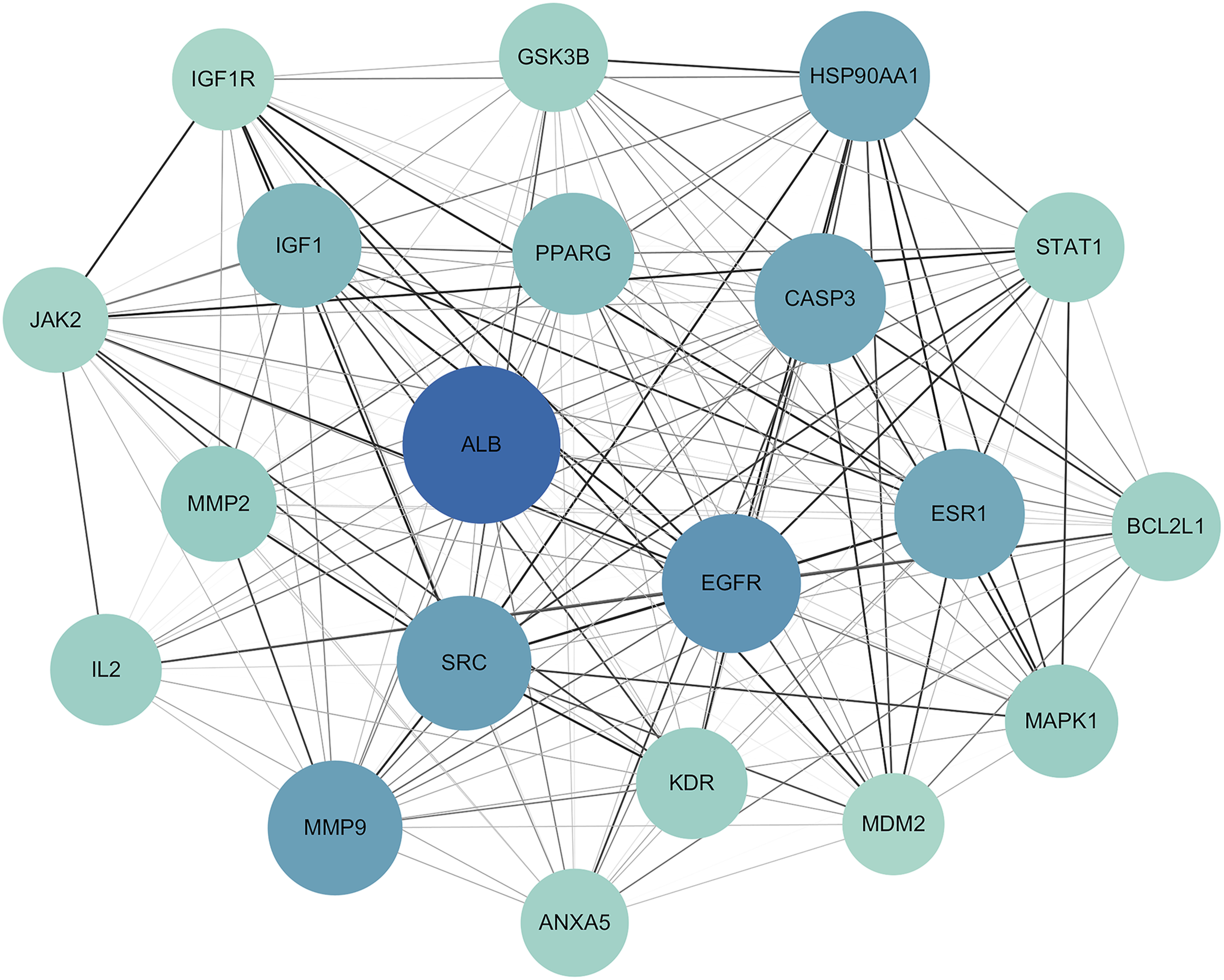
Intersecting genes were imported into the STRING database to construct a PPI network (Figure 3). The data were further analyzed using Cytoscape software, in which edges with higher combined score values were visualized with greater thickness or darker color, and nodes with higher degree values were displayed with larger size or darker color (Figure 4). Twenty target genes with the highest degree values were selected to generate a key gene network for UDCA in OA treatment (Figure 5), identifying Albumin (ALB), Epidermal growth factor receptor (EGFR), Matrix metalloproteinase-9 (MMP9), Proto-oncogene tyrosine-protein kinase Src (SRC), Caspase-3 (CASP3), and Estrogen receptor (ESR1) as the top six core genes. Potential target PPI network of UDCA in OA treatment Visualized PPI network topology Top-20 key genes ranked by degree value

3.4. GO and KEGG enrichment analysis
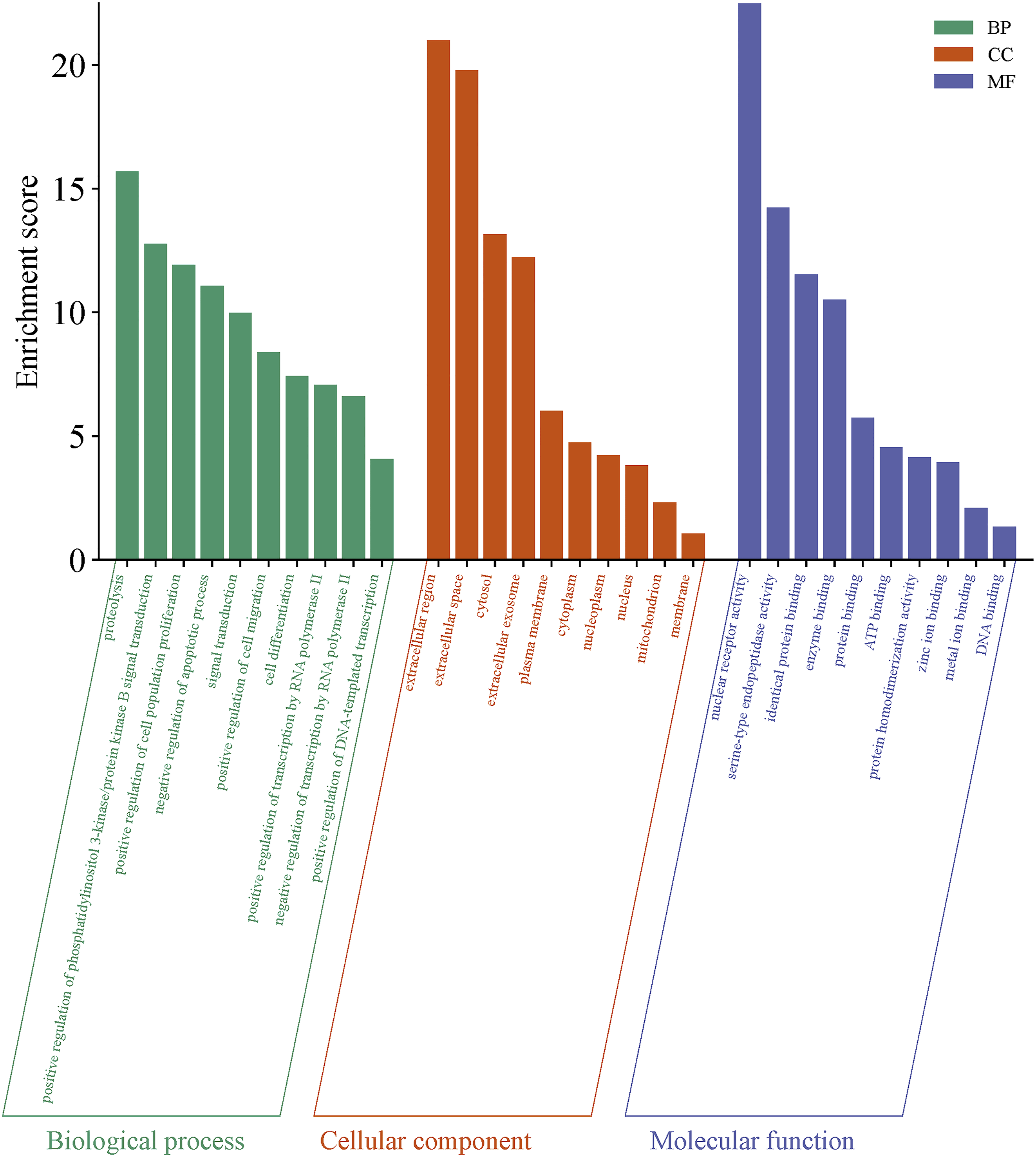
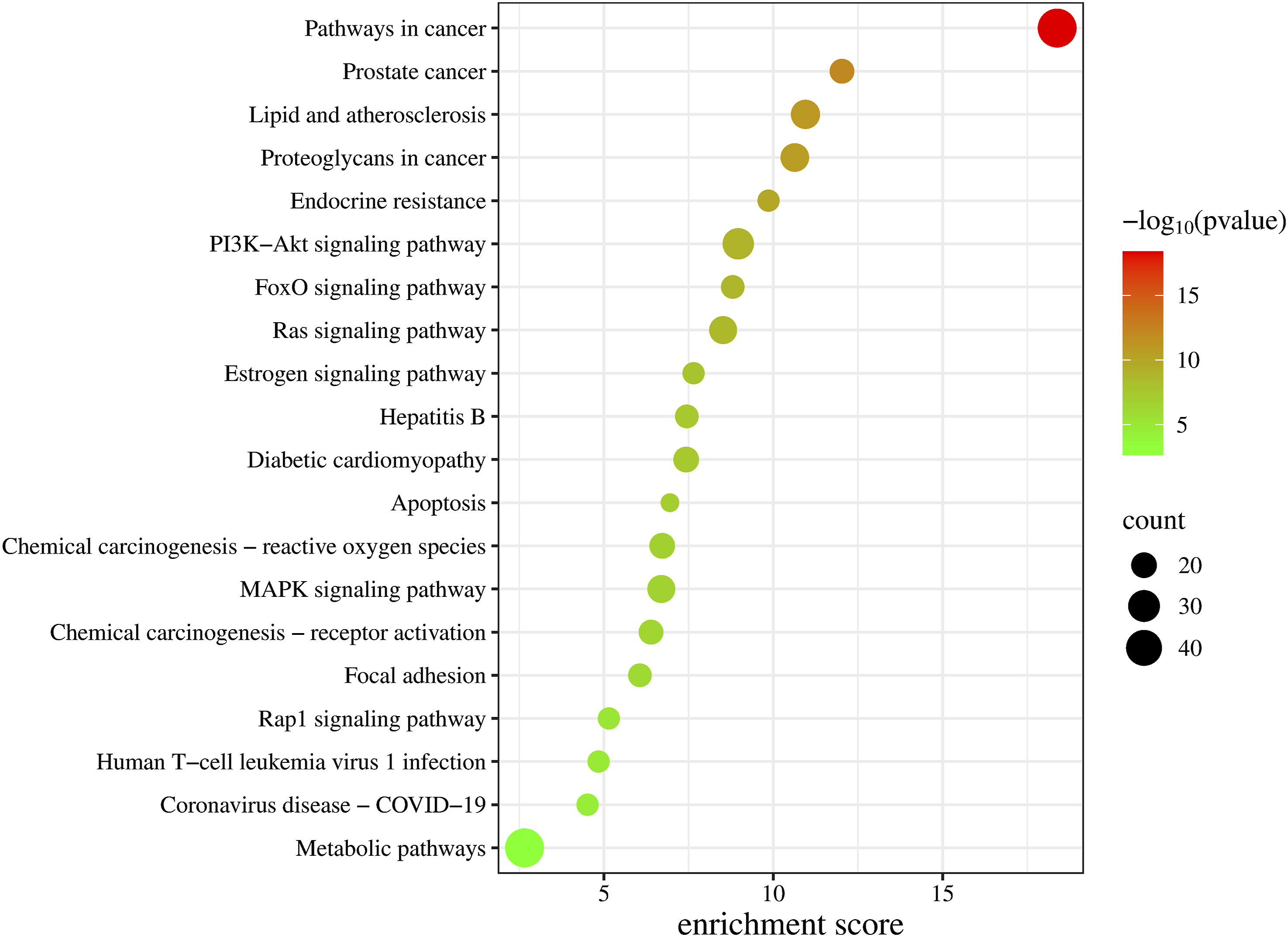
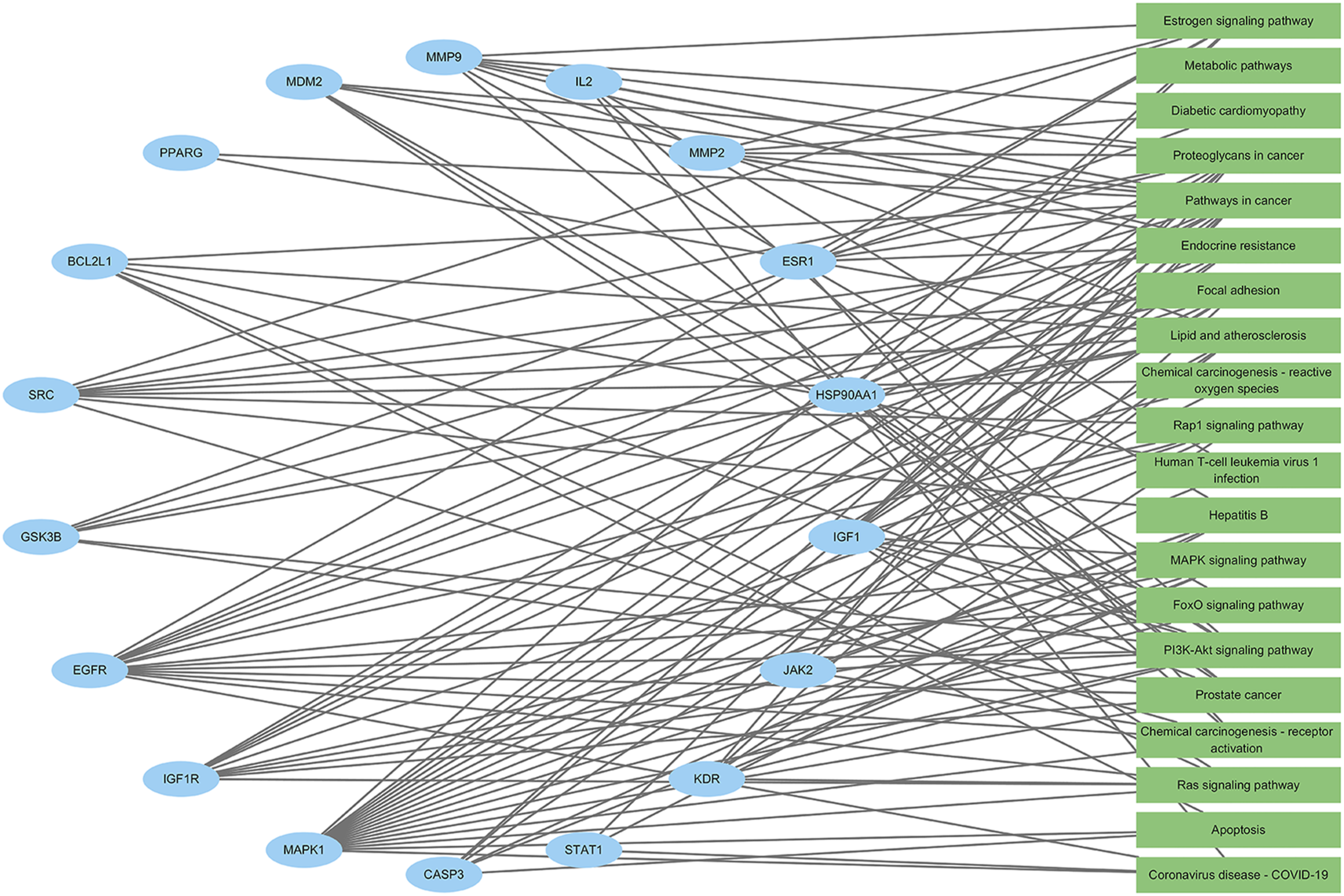
GO functional analysis of intersecting genes revealed that their BP was primarily associated with proteolysis, enhancement of PI3K-Akt signal transduction, stimulation of PI3K-Akt signaling, promotion of cell population growth, inhibition of apoptosis, and signal transduction; CC was mainly localized in the extracellular region, cytosol, and plasma membrane; and MF was predominantly related to nuclear receptor activity and serine-type endopeptidase activity (Figures 6 and 7). KEGG enrichment analysis indicated that the target genes were notably enriched in the PI3K-Akt and MAPK signaling pathways as well as cancer-related pathways (Figure 8). An “enriched pathway-key target” network was constructed (Figure 9). Bubble chart of GO functional enrichment analysis for potential targets Three-in-one chart of GO functional analysis for potential targets Bubble chart of KEGG pathway analysis for potential targets “Enriched pathways-key targets” network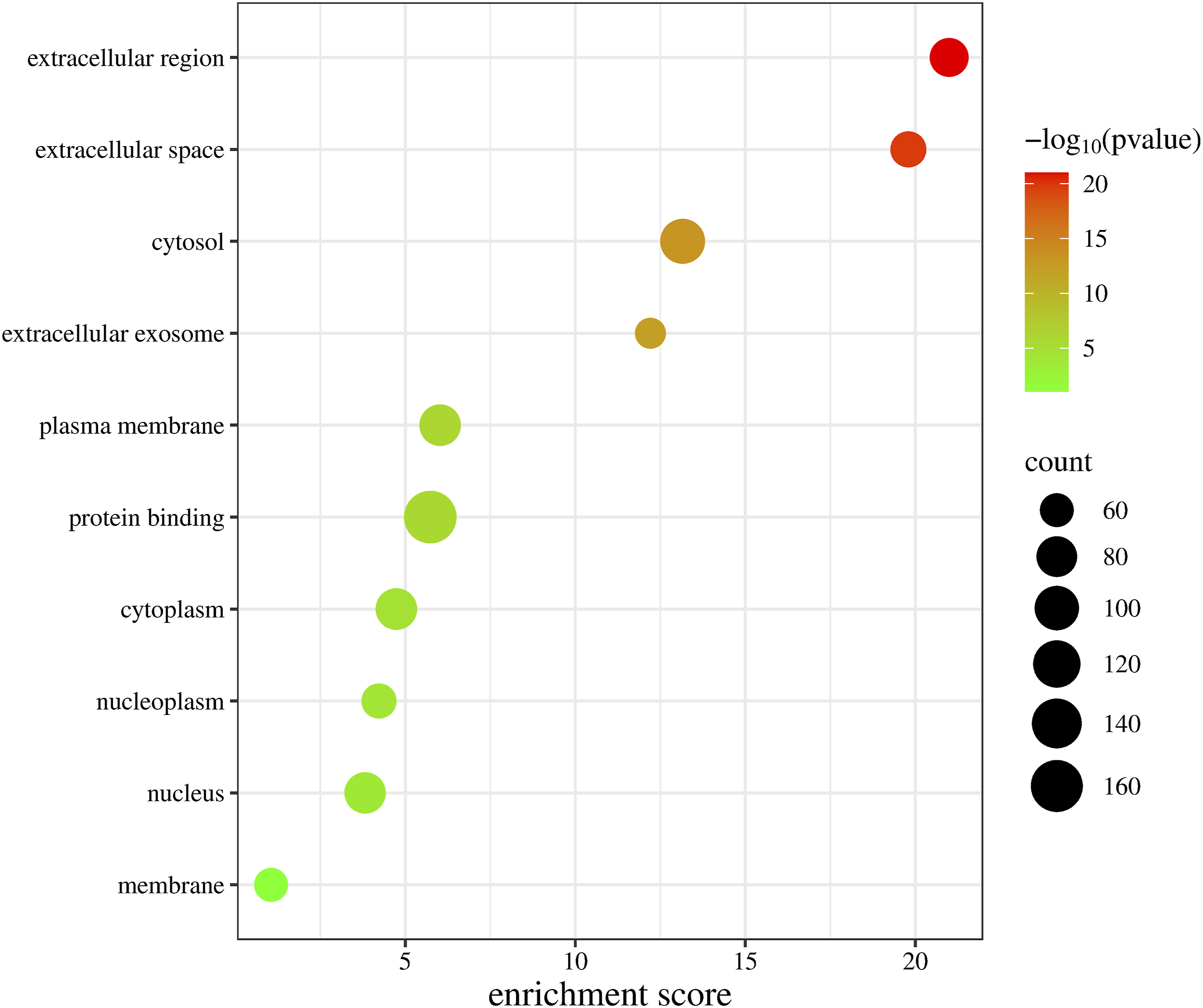
3.5. Molecular docking analysis
Docking at the molecular level was performed on the selected core targets, with their respective PDB IDs being 2BXH, 2ITY, 4WZV, 3G5D, 1PAU, and 3ERT. Docking outcomes were visualized in accordance with methodology described previously
38
(Figure 10). The binding free energy of UDCA was compared with that of the co-crystallized ligands of each protein as shown in Table 1. UDCA exhibited favorable binding energies across all six proteins (all < -6 kcal/mol), with an average binding energy of -7.4 kcal/mol. The binding energy with SRC was the lowest, recorded at -8.0 kcal/mol. The reference ligands exhibited an average binding energy of -9.0 kcal/mol. Although the binding energy of UDCA was marginally lower than the overall binding energy of the reference ligands, it nevertheless exhibited superior binding affinity for CASP3. Schematic diagrams of molecular docking between UDCA and core proteins Binding energy prediction of the interaction between UDCA and reference ligands with core target proteins.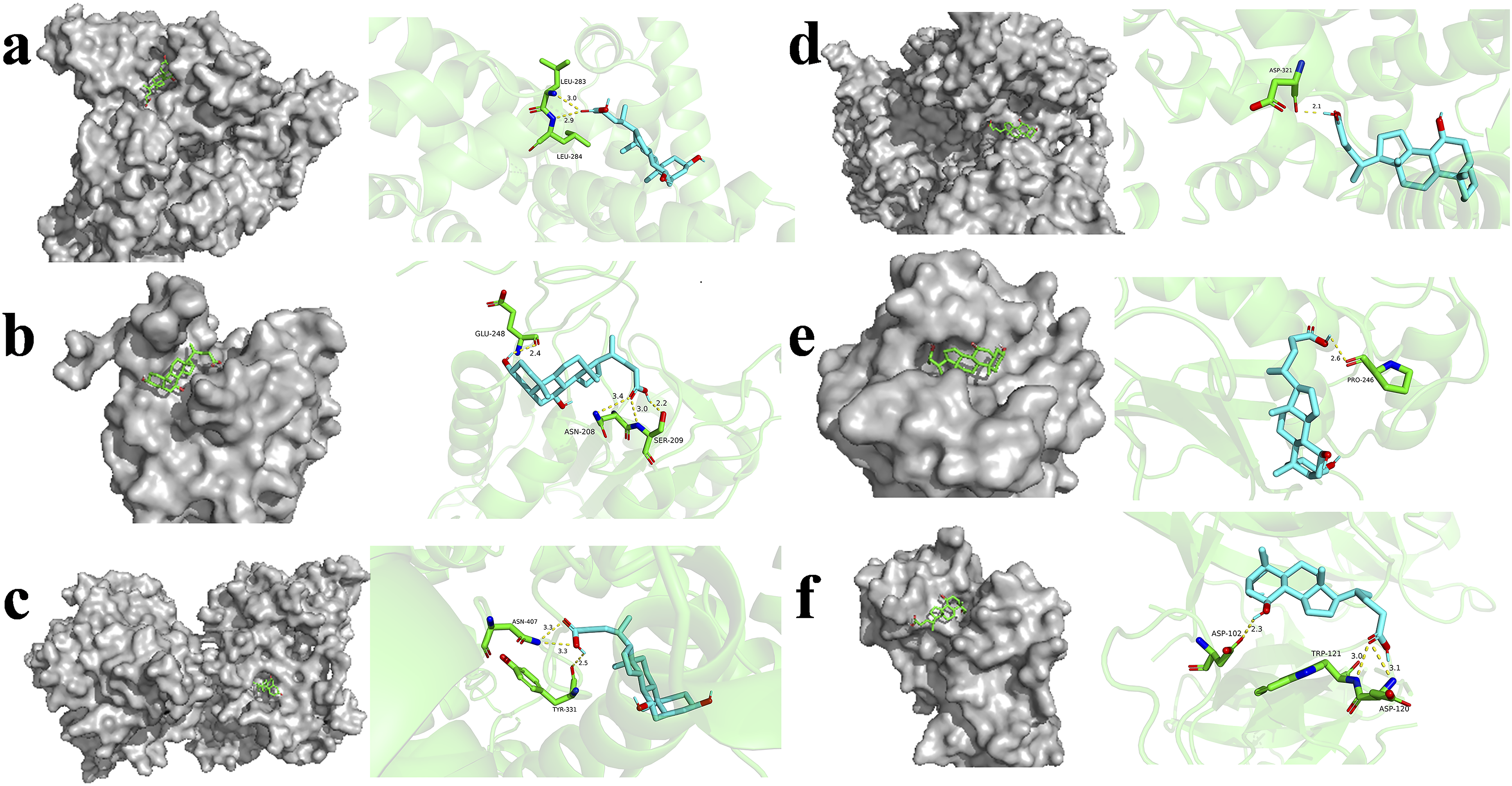
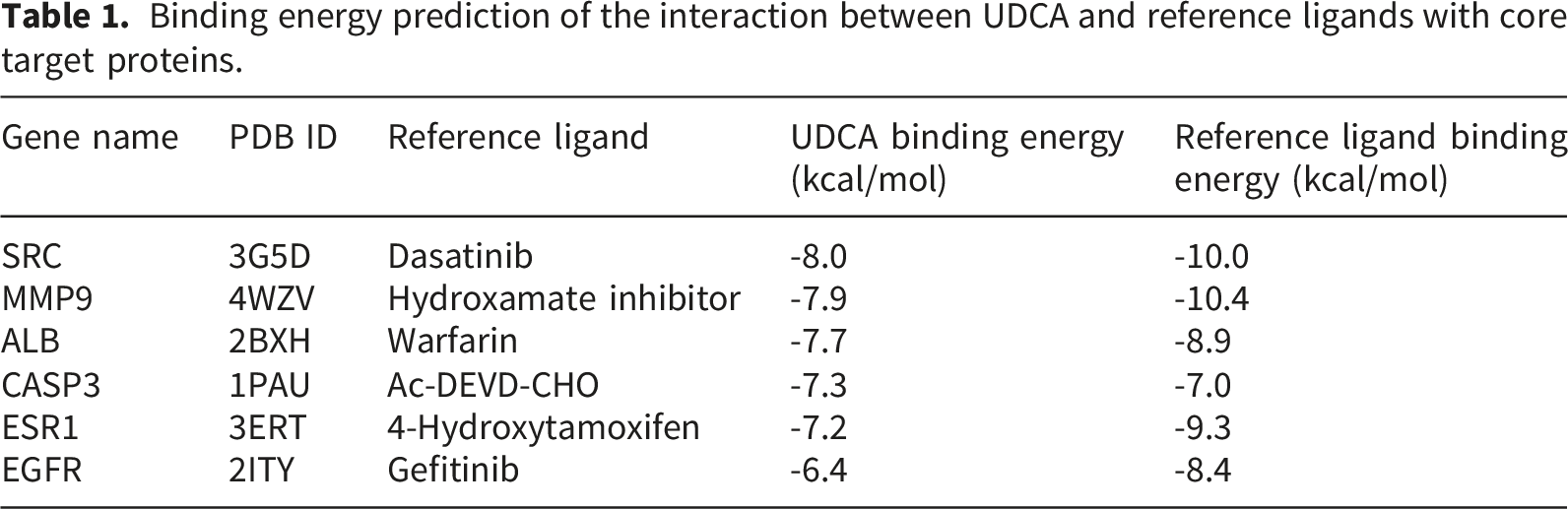
3.6. Molecular dynamics simulation analysis
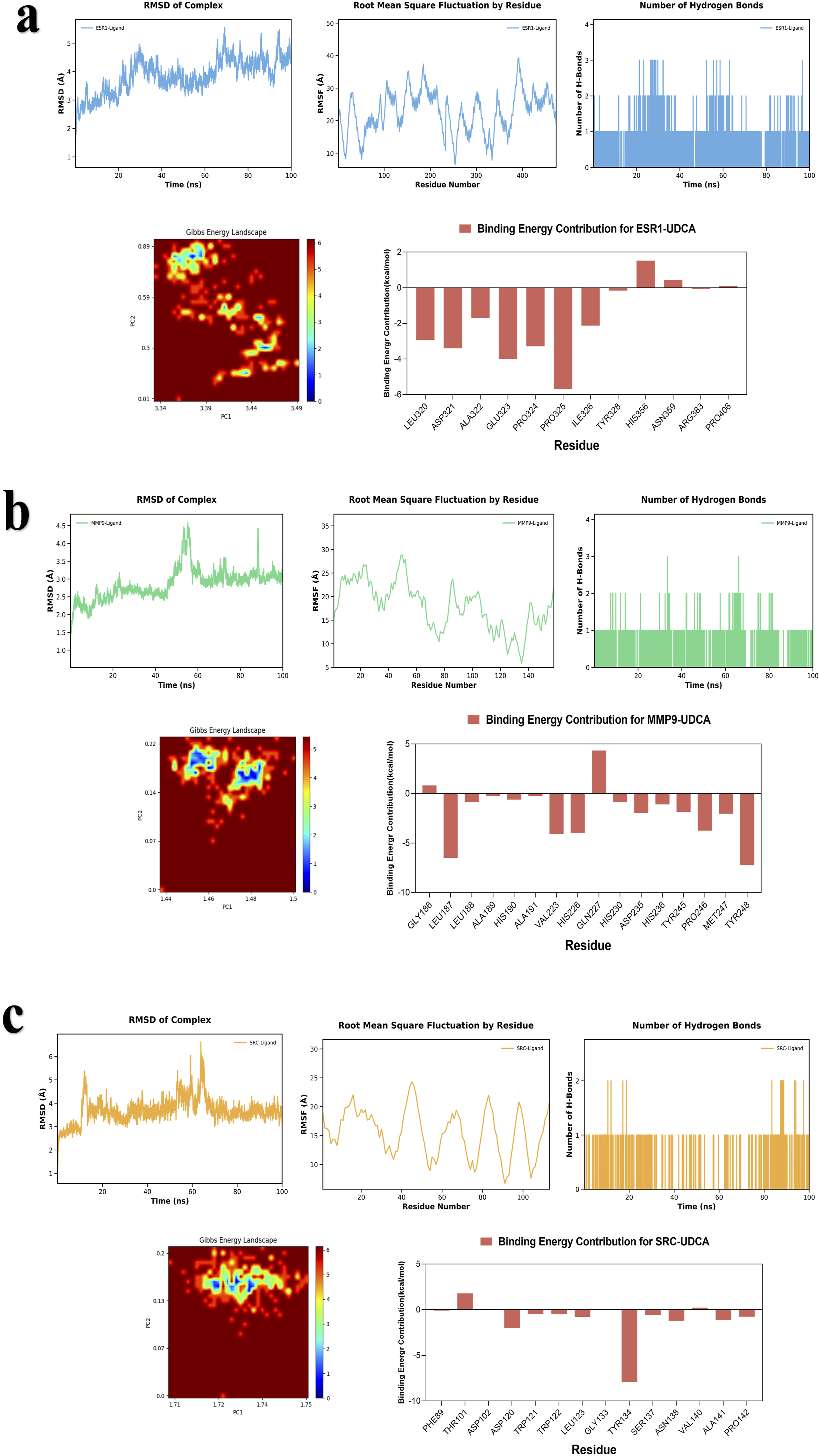
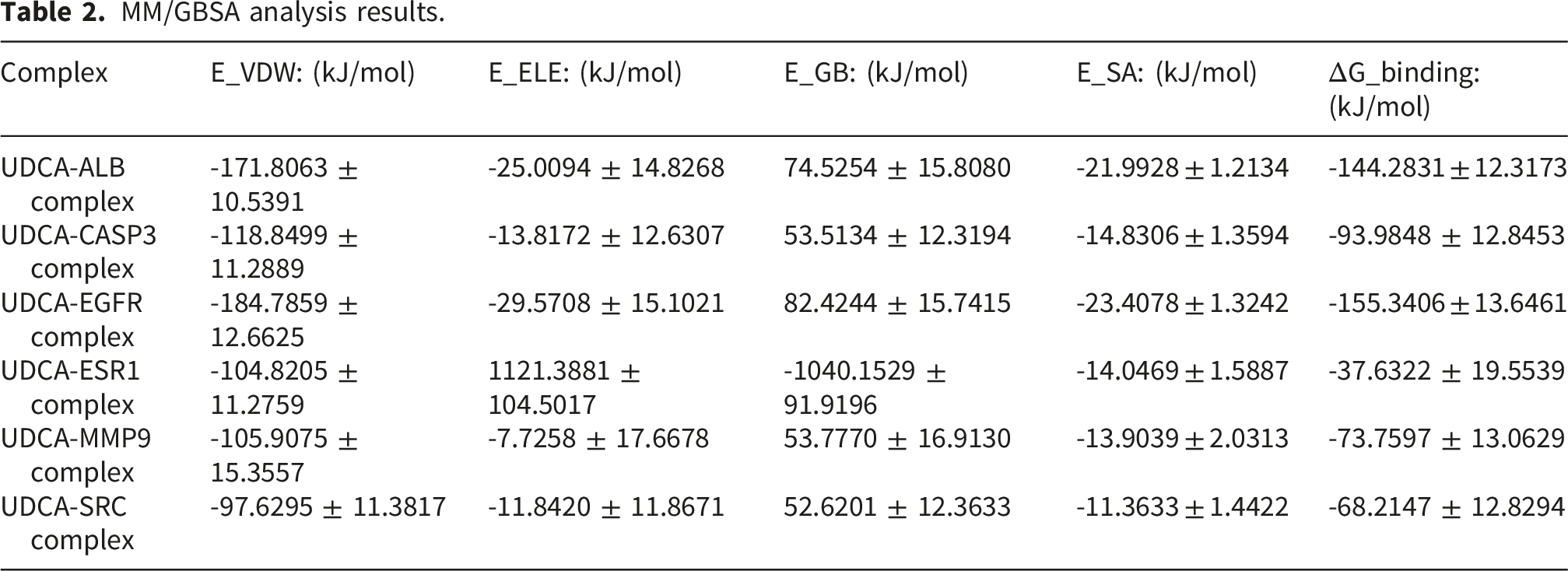
To systematically assess the structural stability, interactions, and binding thermodynamics of the complexes, 100-ns molecular dynamics simulations were performed on six docking complexes derived from the docking results (Figures 11 and 12) and subsequently compared with the reference ligand (Figure 13). Throughout the simulations, RMSD of all complexes remained relatively low, predominantly < 3 Å, indicating that the overall protein structure maintained stability following UDCA binding. Quantitative analysis revealed that the average RMSD of the UDCA complexes was approximately 3 Å, which is marginally higher than the 2 Å observed for the reference ligand complexes. This indicates that, although UDCA may induce partial conformational flexibility in proteins, the extent of these alterations remains constrained within a stable range. Further RMSF analysis revealed that fluctuations involving the key binding residues were relatively low. Among these, the UDCA-ALB complex exhibited the smallest fluctuation amplitude, suggesting higher conformational rigidity. Compared the reference ligand, the UDCA complexes exhibited a marginally higher average RMSF value; however, no significant local structural instability was observed. Hydrogen bond analysis revealed that the number of major hydrogen bonds between the ligand and the protein consistently ranged from one to two during the simulation, indicating stable hydrophilic interactions within the complexes. The free-energy landscape derived from PCA revealed that the majority of the UDCA complexes, along with the reference ligand complexes, displayed steep low-energy basins, indicative of satisfactory conformational convergence. However, UDCA-ALB and UDCA-CASP3 complexes exhibited a comparatively broad low-energy regions, indicating a marginally greater degree of conformational flexibility. Overall, UDCA, like the reference ligands, can stabilize proteins in a favorable conformational state. MM/GBSA calculations showed that the binding free energies for each complex were < -35 kJ/mol. The UDCA-EGFR complex had the lowest binding free energy, reaching -155.3406 ± 13.6461 kJ/mol, indicating strong binding affinity and high stability. Energy decomposition analysis suggested that van der Waals interactions and electrostatic complementarity were the primary driving forces for UDCA binding to the target proteins (Table 2). Further analysis of the decomposition of amino acid residues identified substantial variations among the UDCA complexes. Hydrophobic residues, specifically Leu, Val, Pro, and Phe, emerged as the primary driving forces in a majority of the complexes. In contrast, charged residues, including Lys, Glu, Asp, and His, made significant contributions to only some complexes. Analyses of RMSD, RMSF, number of hydrogen bonds, Gibbs energy landscape, and amino acid binding energy contribution of target protein-UDCA complexes during molecular dynamics simulations. The figures were created by the authors using Amber and AmberTools. ((a) UDCA-ALB; (b) UDCA-CASP3; (c) UDCA-EGFR). Analyses of RMSD, RMSF, number of hydrogen bonds, Gibbs energy landscape, and amino acid binding energy contribution of target protein-UDCA complexes during molecular dynamics simulations. The figures were created by the authors using Amber and AmberTools. ((a) UDCA-ESR1; (b) UDCA-MMP9; (c) UDCA-SRC). Molecular dynamics simulation of target protein-reference ligand complexes MM/GBSA analysis results.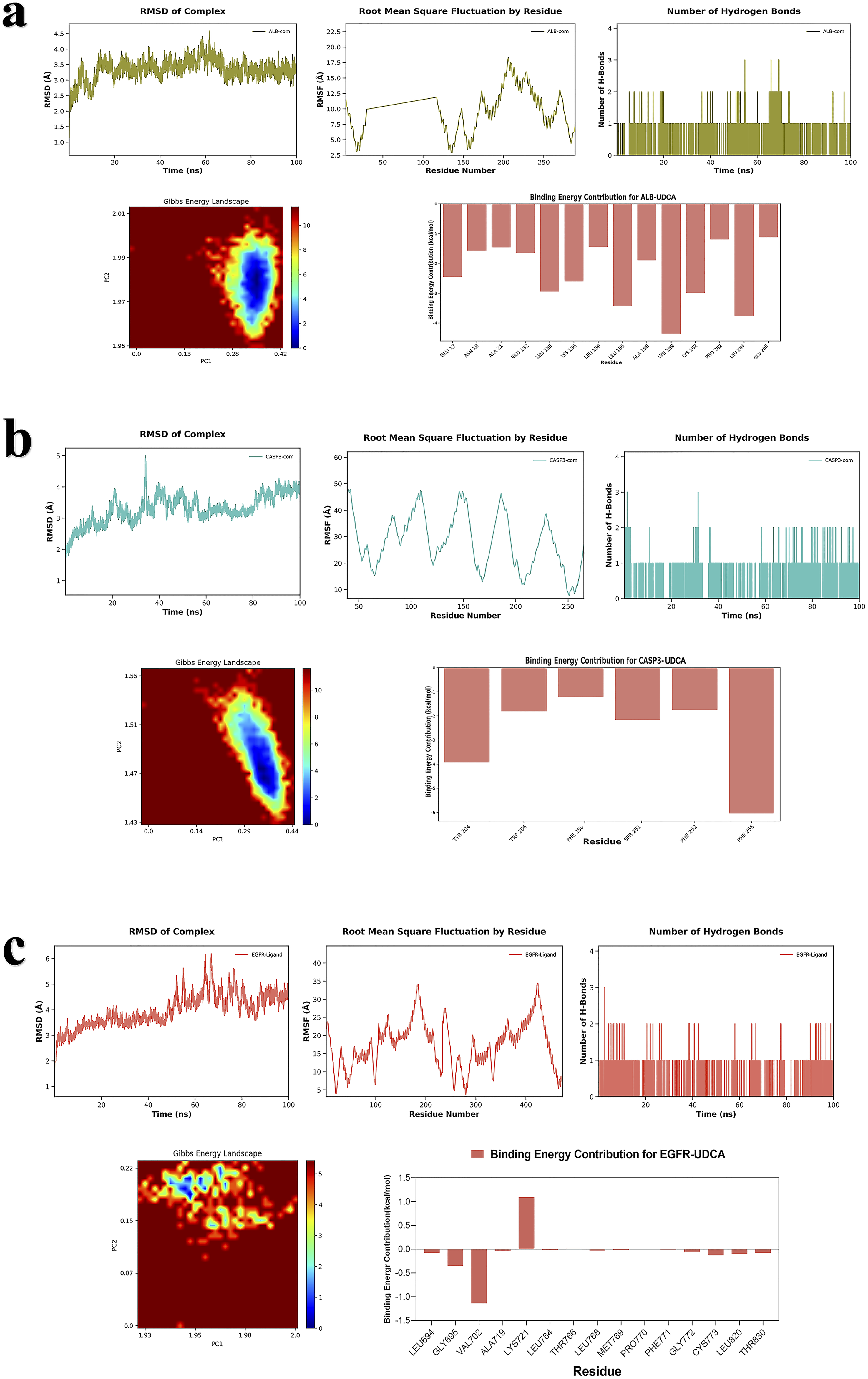
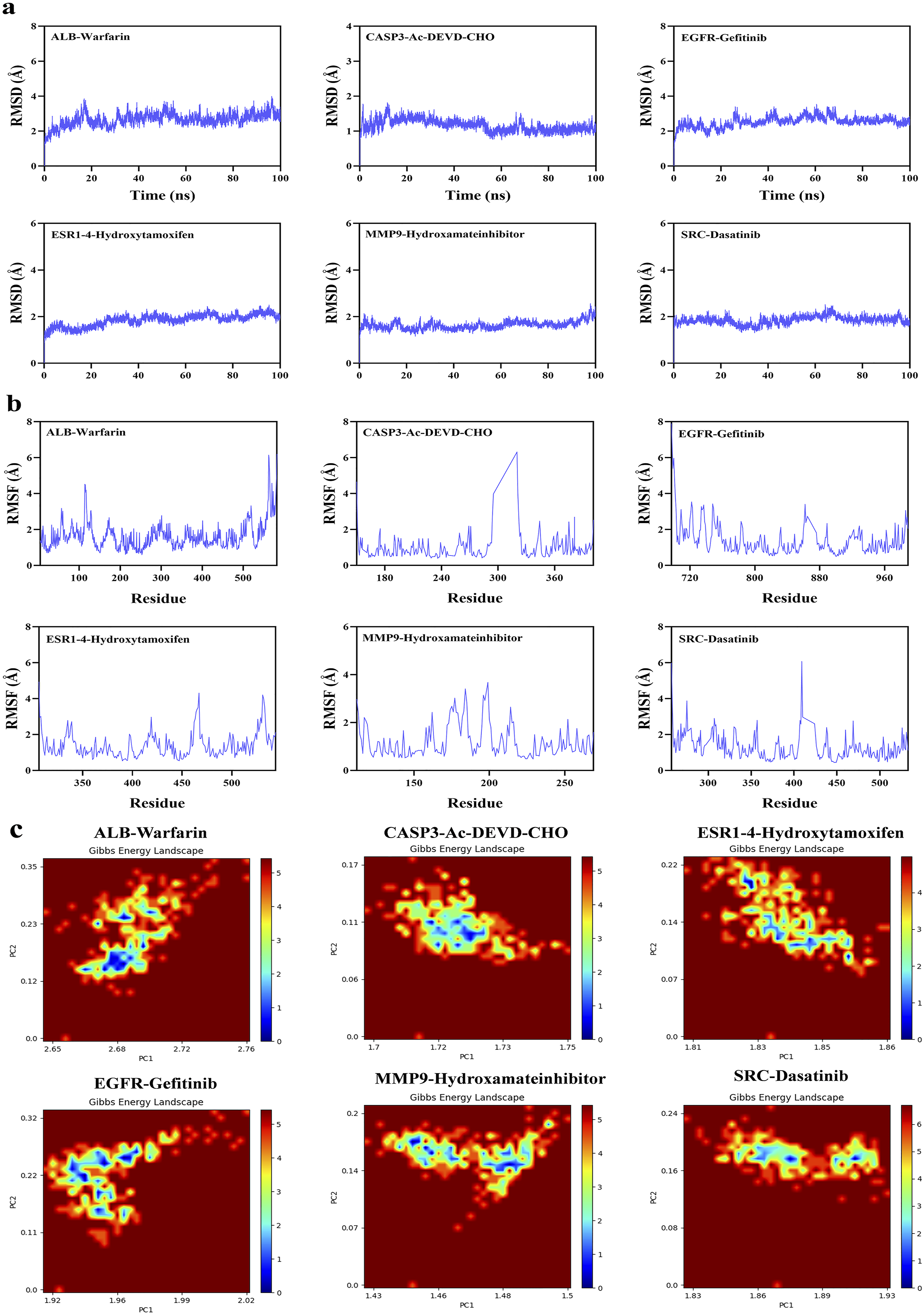
4. Discussion
4.1. Current status of OA treatment
With the continuous increases in average life expectancy and obesity rates, the incidence of OA is also increasing, making it a prevalent disease that significantly impacts human health and contributes to an increasing medical burden. Traditional OA treatment strategies primarily focus on pain relief and functional improvement, whereas patients with severe joint deformities often require artificial joint replacement surgery. Although these therapies have demonstrated efficacy in alleviating symptoms, they are unable to reverse articular cartilage damage and are frequently associated with adverse effects, such as gastrointestinal complications. Recently, innovative OA treatments have been developed, including autologous stem cell therapies derived from adipose tissue, and bone marrow-derived MSCs, PRP therapy, plasmid DNA-encoding long-acting Interleukin-10 (IL-10) variants, and nanoparticle-based targeted drug delivery systems. Additionally, therapies targeting key molecules such as BMP-7, FGF-18, HSA, and IL-1 have also been explored. However, most of these emerging therapies remain in the clinical trial phase, are costly, and face multiple challenges, including insufficient evidence of efficacy, lack of comprehensive safety evaluation, and lack of standardization of operational protocols for widespread clinical implementation.4,39,40
4.2. Therapeutic effects of UDCA in OA
UDCA, an active bile acid with choleretic and hepatoprotective properties, has been approved by the US Food and Drug Administration for primary biliary cholangitis treatment. UDCA has various biological effects, such as mitigating mitochondrial dysfunction and oxidative stress and restoring dopamine synthesis,41,42 which may have therapeutic applications in Parkinson’s disease (PD), non-alcoholic fatty liver disease (NAFLD), and type 2 diabetes.43,44 Recent studies have demonstrated that UDCA and its derivatives possess potential anti-OA properties. Yang and colleagues found that the levels of Glycoursodeoxycholic acid (GUDCA), a derivative of UDCA, are reduced in the blood of patients with OA, and this reduction is negatively correlated with disease severity. UDCA administration attenuates OA progression. 45 In a Destabilization of the Medial Meniscus (DMM)-induced mouse model of OA, UDCA treatment significantly reduced cartilage degradation and inhibited osteophyte formation. 18 This finding was corroborated in an in vivo study. 46 Additional experimental evidence has confirmed the protective effects of UDCA, demonstrating that it can suppress articular cartilage degeneration and slow OA progression.45,46 Furthermore, a clinical cohort study shown that UDCA administration was associated with a decreased risk of joint replacement in patients with OA. 45 According to the aforementioned studies, UDCA demonstrates potential as a therapeutic agent for patients with OA; however, its specific molecular mechanism of action remains unclear. Our study provides an unreported molecular rationale for known therapeutic effects.
4.3. Core target genes
Our research identified ALB, EGFR, MMP9, SRC, CASP3, and ESR1 as the principal targets of UDCA in OA, confirming their potential as multi-target treatment options. These results emphasize that UDCA exerts its therapeutic effects in OA through multiple mechanisms, including the regulation of key signaling pathways via target genes, attenuation of inflammatory responses, inhibition of chondrocyte apoptosis, and modulation of cartilage matrix degradation and synthesis.
Albumin is not only the most abundant protein in the plasma, but is also found in synovial fluid. Albumin may modulate metabolic processes and functional activities of chondrocytes and synovial cells. In vitro studies have demonstrated that albumin preparations can induce bone marrow-derived MSCs to differentiate into chondrocytes and increase the expression of cartilage-related genes, such as aggrecan (ACAN) and type II collagen (COL2A1), and thereby helping maintain cartilage structure and function. 47 In addition, albumin may modulate the inflammatory response in synovial cells. For instance, Low Molecular Weight Fraction of 5% human serum Albumin (LMWF5A) has been found to inhibit the release of Tumor Necrosis Factor-alpha (TNF-α) induced by Lipopolysaccharide (LPS), suggesting its anti-inflammatory properties. 48 Molecular signaling pathway analyses indicate that albumin may regulate multiple OA-related pathways, including Cyclooxygenase-2 (COX-2), Mammalian Target of Rapamycin (mTOR), Cluster of Differentiation 36 (CD36), and Aryl hydrocarbon receptor (AhR), thereby influencing cellular inflammatory and metabolic processes and exerting therapeutic effects in OA. 49 Studies on chondrocytes further reveal that albumin can suppress the production of pro-inflammatory cytokines by inhibiting key pro-inflammatory signaling pathways such as Nuclear Factor Kappa B (NF-κB) and Signal Transducer and Activator of Transcription 1 Alpha (STAT1α), leading to reduced joint inflammation and diminished cartilage damage. 50 Moreover, albumin may interact with other biomolecules within the joint environment to control cell growth, specialization, and programmed cell death, thereby affecting the development of OA.
Cartilage ECM is primarily composed of type II collagen and proteoglycans, which provide structural support and elasticity to the cartilage. During the later phases of OA, macrophages increase the secretion of MMP-9, suggesting an association between MMP-9 and OA. 51 Under the inflammatory conditions of OA, MMP-9 secreted by macrophages, together with other pro-inflammatory mediators, accelerates joint inflammation and promotes progressive cartilage degeneration and excessive subchondral bone growth. 52 Members of the matrix metalloproteinase family, particularly MMP-9, can specifically degrade type II collagen and other ECM components, ultimately leading to structural degradation and functional joint impairment. MMP-9 expression is influenced by inflammatory cytokines. In osteoarthritic chondrocytes, Interleukin-6 (IL-6) promotes MMP-9 expression by activating the MAPK/ERK signaling pathway, thereby contributing to ECM degradation. 53 Moreover, Interleukin-1 beta (IL-1β) serves as one of the primary drivers of MMP-9 expression in chondrocytes. 54
EGFR signaling plays diverse roles in OA progression. Sustained and moderate EGFR activation effectively suppresses the expression of proteases involved in ECM degradation, such as MMP9, Matrix metalloproteinase-13 (MMP13), and ADAMTS5, thereby inhibiting cartilage catabolism, promoting chondrocyte survival, and exerting protective effects. 55 On the other hand, EGFR signaling can also contribute to OA progression. For example, rabbit articular chondrocytes treated with epidermal growth factor (EGF) exhibit increased COX-2 expression and Prostaglandin E2 (PGE2) levels, while suppressing collagen and proteoglycan synthesis. 56 The EGFR ligand TGF-α further promotes cartilage degradation by enhancing MMP expression and downregulating SRY (sex determining region Y)-box 9 (Sox-9) and matrix-related gene expression.57–59 Additionally, EGFR ligands activate the MAPK pathway, which mediates Wnt-induced β-catenin activation 60 and stimulates the expression of MMPs, including MMP9, MMP13, and Matrix metalloproteinase-14 (MMP14), as well as the NF-κB receptor activator ligand (RANKL) in hypertrophic chondrocytes, ultimately leading to cartilage matrix degradation. 60 Therefore, EGFR signaling exerts distinct functional roles at specific stages or subtypes of OA.
SRC is a non-receptor tyrosine protein kinase that serves as a central hub in the cellular signaling network and transmits signals from receptors such as G protein-coupled receptors, growth factor receptors, and integrins to downstream effectors to regulate core cellular functions. Fyn, a member of the SRC family, is significantly upregulated in OA cartilage of both humans and mice. 61 It accumulates in chondrocytes of articular cartilage and interacts with β-catenin, thereby activating the β-catenin signaling pathway and accelerating cartilage degradation and OA progression. Using next-generation sequencing (NGS) and bioinformatic approaches, studies on fibroblast-like synoviocytes (FLS) from patients with OA have revealed that genes such as SRC kinase-associated phosphoprotein 2 (SKAP2) exhibit similar expression patterns in both FLS and OA synovial tissue datasets. These findings suggest that SKAP2 and related genes contribute to OA pathogenesis and further indicate the involvement of SRC-associated signaling pathways in the initiation and progression of OA. 62 Research has shown that inhibiting SRC may be a new approach for OA treatment. 63
CASP3, a member of the cysteine protease family, is activated through multiple signaling pathways and subsequently cleaves a series of intracellular substrates, triggering the apoptotic cascade. In an IL-1β-induced chondrocyte injury model, CASP3 activity is significantly elevated, forming a vicious cycle of inflammation and apoptosis. 64 Knockdown of CASP3 has been shown to modulate key proteins such as B-cell leukemia/lymphoma 2 (Bcl-2), MMP-13, and β-catenin, producing effects that reduce inflammation, apoptosis, and ECM degradation. 64 However, the role of CASP3 in the different stages of OA remains controversial. Some studies have proposed that activation of CASP3 during the early stages of OA may serve as a protective mechanism by eliminating damaged cells and preserving intra-articular homeostasis. In contrast, the excessive activation of CASP3 during the later stages of OA may promote chondrocyte apoptosis and accelerate disease progression. However, further studies are required to substantiate this hypothesis.
The ESR1 gene encodes estrogen receptor alpha (ERα), a crucial nuclear receptor involved in mediating estrogen signaling. Accumulating evidence from investigations carried out in vitro and in vivo demonstrates that ERα-mediated regulation of inflammatory responses exerts a protective effect against synovitis. 65 ERA-63, a selective ERα agonist, alleviates OA symptoms and associated joint pathology by significantly reducing the levels of Interleukin-12 p40 subunit (IL-12p40,) IL-6, IL-1β, Regulated upon Activation, Normal T-cell Expressed and Secreted (RANTES), and Protein Kinase C (KC) in the synovium. Furthermore, ERA-63 inhibits the estrogen-mediated activation of NF-κB, 66 blocks its nuclear translocation and transcriptional activity, and consequently suppresses the expression of downstream pro-inflammatory mediators and MMPs. Manolagas et al. reported that ERα activates the Wnt signaling pathway to promote osteogenesis. 67 Zha and co-workers demonstrated that ERα enhances the chondrogenic differentiation of MSCs through activation of the Jun N-terminal kinase 1/2 (JNK-1/2) signaling pathway. 68 Additionally, ESR1 upregulate the expression of Insulin-like Growth Factor Binding Protein-2 (IGFBP-2), 69 which indirectly promotes chondrocyte proliferation and matrix synthesis by modulating IGF signaling.
Collectively, UDCA may exert therapeutic benefits in OA by influencing the expression of these factors.
4.4. GO and KEGG pathways enrichment analysis
To further clarify the biological roles of potential target genes of UDCA in OA treatment, these genes were subjected to GO and KEGG pathway enrichment analyses.
According to functional insights from GO enrichment analysis, UDCA was found to primarily modulate nuclear receptor activity, serine-type endopeptidase activity, and protein hydrolysis during OA treatment. Multiple nuclear receptors, such as Liver X Receptors (LXRs) and Farnesoid X receptor (FXR), are dysregulated in OA tissues and are implicated in anti-inflammatory responses, inhibition of tissue degradation, and suppression of osteoclast activity.70–72 Serine-type endopeptidases represent a class of protein hydrolases, among which High Temperature Requirement Serine Protease A1 (HtrA1) is significantly upregulated in OA cartilage and indirectly exacerbates cartilage matrix degradation. 73 Therefore, UDCA may exert therapeutic effects in OA by modulating various nuclear receptors and regulating key protein hydrolysis processes involved in OA progression.
Further analysis of the KEGG pathways revealed that UDCA affected OA through the PI3K/Akt and MAPK signaling pathways. The PI3K/Akt signaling pathway is crucial in the pathological processes of OA, such as cartilage degeneration and synovitis.74,75 By inhibiting this pathway, UDCA can regulate chondrocyte autophagy, maintain cartilage homeostasis, suppress inflammation, and alleviate OA.76,77 Moreover, activation of the PI3K/Akt signaling pathway promotes chondrocyte proliferation and exerts protective effects against OA.77–79 The MAPK signaling pathway subfamilies implicated in OA pathogenesis include JNK, p38 mitogen-activated protein kinase (p38), and ERK, and the activation of these pathways ultimately leads to ECM degradation, inflammatory responses, and chondrocyte apoptosis.80–83
4.5. Molecular docking and molecular dynamics simulations
To assess the ability of UDCA, a naturally occurring bile acid, to establish stable binding interactions, we comprehensively investigated its potential binding sites across the protein surface using blind docking methodologies. Molecular docking demonstrated that UDCA exhibited strong binding affinities for key targets, including ALB, EGFR, MMP9, SRC, CASP3, and ESR1. These findings provide a molecular-level theoretical foundation for understanding the regulatory effects of UDCA in OA. Furthermore, variations in binding energy compared to the reference ligands may be attributed to structural differences between UDCA and the inhibitors of co-crystallized ligands, potentially resulting in divergent binding modes. As for further molecular dynamics simulations, RMSD analysis revealed that the overall structural stability of UDCA complexes was marginally lower than that of the reference ligand. This difference may be attributed to fundamental disparities involving their chemical structures. The reference ligand, a well-established active molecule, has undergone extensive optimization and validation over numerous years, resulting in enhanced geometric congruence with the binding pocket of the target protein. In contrast, UDCA’s steroid skeleton and functional group distribution, which was not specifically designed, confer a relatively limited ability to constrain protein conformation. RMSF analysis further indicated that UDCA induced slightly greater fluctuations in protein residues than the reference ligand did, suggesting that UDCA exerted a comparatively weaker stabilizing effect on the local conformation of the binding pocket. Nonetheless, the key binding residues did not exhibit significant destabilization, implying that UDCA was still capable of maintaining its binding to the target protein. Analysis of hydrogen bonding suggests that hydrophilic interactions stabilize auxiliary functions in the binding process. Examination of the free energy landscape revealed that the conformations of the reference ligands were highly convergent. In contrast, partial UDCA complexes exhibited a more extensive low-energy region and greater conformational heterogeneity, which is consistent with the observation that the binding energy of UDCA is generally weaker than that of the reference ligand. Despite the increased conformational flexibility of UDCA, which results in reduced thermodynamic stability compared with that of the reference ligand, this flexibility may underpin its capacity for adaptive binding across different target proteins. MM/GBSA calculations demonstrated that UDCA exhibited spontaneous binding to each target protein, with a particularly high affinity for EGFR. The binding interactions between UDCA and the target proteins are primarily driven by hydrophobic effects, supplemented by electrostatic and hydrogen bonding interactions, which align with the intrinsic characteristics of bile acids.
In conclusion, the binding stability and thermodynamic properties of UDCA for target proteins were generally inferior to those of the optimized reference ligand. Nonetheless, UDCA exerts a therapeutic effect in OA by effectively and stably binding to multiple targets.
5. Limitations
This study relied primarily on database-derived information, which may have introduced potential bias. Furthermore, limitations of the algorithms employed could result in false-positive or false-negative results. Additionally, the absence of integrated in vitro and in vivo experimental validation and functional studies at the cellular level limit the ability to confirm the actual regulatory roles of predicted targets within physiological contexts. Moreover, clinical trials verifying the clinical efficacy of UDCA are currently lacking. The current findings lacked sufficient clinical evidence from large-scale trials to substantiate the therapeutic efficacy of UDCA in patients with OA. Future studies should focus on establishing cell-based and animal models combined with histopathological and molecular testing to verify the regulatory effects of key targets of UDCA and its regulatory impact on core pathways, thereby further strengthening the chain of evidence.
6. Conclusion
The role of UDCA in treating OA may involve its potential effects on specific targets such as ALB, SRC, EGFR, ESR1, MMP9, and CASP3. It intervenes in the expression of related signaling pathways, such as MAPK and PI3K/Akt, thereby regulating the levels of relevant inflammatory mediators, inhibiting chondrocyte apoptosis, and modulating the balance between cartilage matrix degradation and synthesis, thereby comprehensively reducing pathological damage to joint tissues. Therefore, UDCA may serve as a potential drug for the treatment of OA and warrants further research.
Supplemental material
Supplemental material - Exploring the mechanism of ursodeoxycholic acid in the treatment of osteoarthritis based on network pharmacology, molecular docking, and molecular dynamics simulations
This supplemental material provides a table of coordinate centers and mesh sizes used for molecular docking in this study.
Supplemental material
Supplemental material - Exploring the mechanism of ursodeoxycholic acid in the treatment of osteoarthritis based on network pharmacology, molecular docking, and molecular dynamics simulations
Supplemental material for Exploring the mechanism of ursodeoxycholic acid in the treatment of osteoarthritis based on network pharmacology, molecular docking, and molecular dynamics simulations by Xin Li, Jinhe Xiong in European Journal of Inflammation
Footnotes
Acknowledgements
We acknowledge the use of public databases for gene targets, open-source software for data analysis and visualization, and open-access resources. We also thank the authors of the cited literature for providing the theoretical foundation during the writing process. We would like to thank Editage (![]() ) for English language editing.
) for English language editing.
Ethical considerations
This article does not contain any studies with human or animal participants.
Author contributions
All authors reviewed the manuscript.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Supplemental material
Supplemental material for this article is available online.
Appendix
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.