
Abstract
Spinal cord injury (SCI) triggers neuropathic pain through inflammatory secondary injury. Olfactory ensheathing cells (OECs) hold therapeutic promise, but their pain-relieving mechanisms are unclear. This study investigated whether OEC transplantation alleviates pain by modulating macrophage extracellular trap (MET) formation and Na+–K+–Cl− cotransporter 1 (NKCC1) expression. Following ethical approval, a rat contusive SCI model was established. OEC-treated rats received a cell transplant post-injury. Motor and sensory functions were assessed over 35 days. Tissue was analyzed for NKCC1, CD68, and the MET marker H3Cit. In vitro, RAW264.7 macrophages were stimulated with TNF-α to induce METs. OEC transplantation significantly improved functional recovery and reduced mechanical/thermal hypersensitivity. Histology showed preserved tissue structure. Critically, OEC treatment suppressed MET formation (reduced H3Cit/CD68) and downregulated NKCC1 in vivo. In vitro, OEC-conditioned medium inhibited TNF‑α‑induced MET formation and NKCC1 expression in macrophages. Thus, OECs alleviate neuropathic pain and promote recovery after SCI by coordinately suppressing macrophage extracellular traps and regulating NKCC1 expression, revealing a novel immunomodulatory‑ion transport axis for pain intervention.
Keywords
Introduction
Spinal cord injury (SCI) represents a severe condition affecting the central nervous system, characterized by a biphasic pathological progression comprising primary and secondary injury phases. 1 Secondary injury involves inflammatory responses, neurodegeneration, and glial scar formation, which exacerbate the hostile microenvironment at the lesion site, leading to sensory, motor, and autonomic dysfunctions.2,3 Pain is a prevalent and debilitating consequence of spinal cord injury, affecting ~61% of patients. The onset of neuropathic pain (NeuP) is associated with the disinhibition of nociception. 4 Following injury to primary sensory neurons, resident macrophages in nerves proliferate, while peripheral macrophages are recruited to both the lesion site and the dorsal root ganglia. 5 These accumulated macrophages, particularly those of the M1 phenotype, produce a wide array of inflammatory mediators that activate multiple nociceptive signaling pathways, thereby sensitizing or exciting nociceptors.6–8 Notably, such inflammatory mediators, including tumor necrosis factor alpha and interleukin‑1 beta, also upregulate the expression of NKCC1. 9 As a key protein in maintaining intracellular chloride homeostasis in neurons, elevated NKCC1 expression leads to a rise in cytoplasmic chloride concentration, which attenuates GABAergic inhibitory signaling. This attenuation further exacerbates the disinhibition of nociception and facilitates the development of NeuP. 10
As a form of cell death, extracellular trap (ET) formation diverges mechanistically from classical programmed cell death and necrosis. ETs are extracellular web-like structures composed of DNA and proteins. When released by macrophages, they are termed macrophage extracellular traps (METs).11,12 Zhang et al. identified METs at the site of SCI and demonstrated that these structures promote the polarization of microglia toward the M1 phenotype, thereby intensifying local inflammation. 12 Given that METs originate from macrophages and that their proinflammatory properties are closely associated with the genesis of pain, modulating the formation or function of METs may alleviate the inflammatory response at the injury site and consequently improve NeuP following SCI.
Although cell transplantation holds promise for the treatment of SCI, its application to NeuP following SCI remains underexplored. OECs are specialized glial cells derived from the neural crest that exhibit functional properties reminiscent of astrocytes, Schwann cells, and oligodendrocytes.13,14 By modulating inflammation, secreting neurotrophic factors, and promoting axonal regeneration as well as remyelination, OECs improve the post injury microenvironment. 15 Notably, cell transplantation has demonstrated potential for alleviating pathological pain in other pain models. Lu et al. transplanted OECs into a rat model of trigeminal neuralgia and observed a marked increase in the facial mechanical pain threshold. 16 The proposed analgesic mechanism entails the secretion of anti‑inflammatory factors (such as interleukin‑1 receptor antagonist (IL‑1Ra)) by OECs, which suppress excessive macrophage activation and nociceptive hyperalgesia, thereby ameliorating the inflammatory microenvironment and relieving pain.17,18
Here, we show that transplantation of OECs alleviates NeuP in a rat model of SCI. We further reveal that the secretory proteome of OECs modulates the formation of METs, and identify a previously unrecognized mechanism by which OECs suppress MET generation post-injury, uncovering a novel anti-inflammatory pathway. Collectively, these findings not only propose a promising therapeutic strategy for NeuP after SCI, but also establish a conceptual framework for how OEC transplantation regulates inflammatory signaling.
Materials and methods
Ethical approval
All experimental protocols were reviewed and approved by the Animal Care and Use Committee of Capital Medical University (ethical approval no. AEEI-2024-239) and complied with the NIH Guide for the Care and Use of Laboratory Animals. The study rigorously adhered to the principles of Replacement, Reduction, and Refinement to minimize both the number of animals used and their potential distress.
Culture of RAW 264.7 macrophages and induction of MET formation
The immortalized RAW 264.7 macrophages, provided by the Cell Resource Center of the Institute of Basic Medical Sciences (CAMS/PUMC, Beijing, China) were grown under standard conditions at 37 °C in an atmosphere containing 5% CO2 and maintained in a humidified environment. The growth medium utilized was high-glucose DMEM (Gibco, USA), which was further supplemented with 10% fetal bovine serum (FBS; Servicebio, China) and 1% penicillin–streptomycin. For immunofluorescence staining, cells were seeded onto 12-mm coverslips 1 day prior to treatment. Macrophages were first incubated at 37 °C for 1 h and then stimulated with TNF-α (20 ng/mL; R&D Systems, Minneapolis, MN, USA) for an additional 4 h. After stimulation, cells were washed with 1× phosphate-buffered saline (PBS) and fixed with 4% cold paraformaldehyde before IF staining. For Western blot protein extraction, cells were cultured and METs were induced following the same protocol, excluding the paraformaldehyde fixation step.
Animals
Thirty-six adult female Sprague–Dawley (SD) rats (8 weeks old; 260 ± 15 g) were procured from the Institute of Rehabilitation Medicine, China Rehabilitation Science Institute (Beijing, China) and maintained under standard laboratory conditions (22 °C ± 2 °C; 55% ± 10% humidity; 12 h light/dark cycle) with ad libitum access to food and water. The animals were then randomly allocated to three experimental groups: Sham, SCI, and OECs groups. OECs transplantation was performed in SCI model rats. Prior to tissue collection, deep anesthesia was induced by intraperitoneal injection of pentobarbital sodium (50 mg/kg; Sigma-Aldrich, China).
Spinal cord contusion
Throughout surgery, rats were maintained under anesthesia with gaseous isoflurane in oxygen (RWD Life Science Co., Ltd., China; 5% induction, 2% maintenance). After preparation with hair removal and 75% ethanol disinfection, the T10 vertebra was accessed through a 2–3 cm midline incision. Subsequent retraction of the paravertebral muscles and a T9–T10 laminectomy exposed the dorsal spinal cord. The T8 and T11 spinous processes were clamped to stabilize the spine, and a 150 kdyn contusive injury was delivered at T10 using an Infinite Horizons Impactor (IH-400; Precision Systems and Instrumentation, Lexington, KY, USA) with a 2.5 mm tip. This force was selected based on preliminary studies indicating that it produces incomplete SCI accompanied by consistent NeuP behavior by 2–3 weeks post-injury.
In sham-operated rats, all surgical procedures were performed except for the impact. Following layered suturing of the muscle and skin and disinfection with iodophor, the rats recovered on a 30 °C heating pad under continuous monitoring until the anesthesia wore off. Postoperative care consisted of individual housing, daily subcutaneous injections of 2 ml sterile saline, a 3-day course of prophylactic antibiotics, and manual bladder expression performed at least twice daily until normal urination was restored. Female rats were exclusively used in this study to minimize urological complications. 19
Experimental design
We isolated primary OECs from the olfactory bulbs of 6-week-old SD rats and expanded them to passages 5–6. To establish an in vitro model of MET formation, we polarized RAW 264.7 macrophages with TNF-α and then cultured them in either standard conditioned medium (CM) or conditioned medium derived from OECs (OEC-CM). OEC identity and purity were confirmed by immunofluorescence and confocal microscopy for the OEC marker p75NTR. MET formation was assessed via Nuclear Green staining, immunofluorescence, and Western blotting for H3Cit and CD68. NKCC1 protein expression was evaluated in parallel by immunofluorescence and Western blotting, providing a comprehensive assessment of the modulatory effects of OECs and OEC-CM on MET formation and NKCC1 expression in vitro.
Thirty-six SD rats were randomly divided into three groups of 12: Sham, SCI, and OECs (SCI with OEC transplantation). Sham animals underwent laminectomy without spinal cord injury. Both SCI and OEC groups received a contusion injury at the T10 level. Thirty minutes post-injury, the SCI group received a 4 μl injection of OEC culture medium, while the OEC group was administered 4 μl of olfactory ensheathing cell suspension (1.0 × 105 cells/μl).
To evaluate the therapeutic effect of OEC transplantation on NeuP, motor and pain-related behaviors were assessed at 7, 14, 21, 28, and 35 days after surgery. Histological analysis using hematoxylin–eosin was performed to quantify lesion cavity area. Immunofluorescence and Western blot analyses were conducted to examine protein expression of CitH3, CD68, and NKCC1 in the spinal cord and dorsal root ganglia, thereby evaluating the influence of OEC transplantation on macrophage extracellular trap formation and NKCC1 signaling.
Primary OECs culture and conditioned medium collection
Primary OECs were obtained from neonatal SD rats. Briefly, olfactory bulbs were dissected, minced, and enzymatically digested in 0.25% trypsin-EDTA (Servicebio, China) for 25 min at 37 °C. Purification employed a modified differential adhesion method. Non-adherent cells were transferred after 18 h, and a second adhesion step was performed at 36 h to remove fibroblasts and astrocytes. The enriched OECs were plated on poly-L-lysine-coated dishes at 1 × 105 cells/ml in DMEM/F12 (Gibco, USA) with 15% FBS (Servicebio, China) and 1% penicillin–streptomycin, with medium refreshed every 2–3 days. For subculturing, OECs at 80%–90% confluence were trypsinized and passaged at a 1:2–1:3 ratio in DMEM/F12 (Gibco, USA) containing 10% FBS (Servicebio, China) and 1% penicillin–streptomycin. To produce OEC-CM, cells were maintained in serum-free DMEM/F12 for 48 h before the medium was collected. CM was prepared identically but without cells. All collected media were centrifuged at 1000g for 5 min to remove cell debris, followed by filtration through a 0.22 μm syringe filter for sterilization. Sterile samples were then concentrated using a centrifugal filter unit (Beijing Baiyang Medical Equipment, Beijing, China) with a 3 kDa molecular weight cut-off at 6000g. For long-term storage, the conditioned media were processed by concentration and aliquoting before being frozen at −80 °C.
OECs preparation
Rats subjected to spinal cord impact injury received immediate injections of OEC suspension (1.0 × 105 cells/μl; total 4 μl). Sham-operated and SCI control groups were administered an equivalent volume of OEC culture medium. We administered injections at four sites, each positioned ~2 mm rostral or caudal to the lesion center and 1 mm lateral to the midline. Using a microinjection system, 1 μl was infused per site at a rate of 1 μl/min to a depth of 0.8 mm.
BBB score
Motor function in rats was assessed using the 21-point Basso, Beattie, and Bresnahan (BBB) locomotor rating scale (0, no movement; 21, normal gait). 20 Hindlimb function was evaluated preoperatively and at days 0, 7, 14, 21, 28, and 35 post-injury by a blinded observer. In each session, animals were placed in an open field and allowed to explore freely for 4 min. Left and right hindlimb performances were scored independently and averaged. Successful SCI modeling was confirmed by sustained BBB scores of 0–1 during the first 3 days post-injury.
Paw withdrawal mechanical threshold
To evaluate mechanical sensitivity, von Frey filaments were employed. 21 Following a 30-min acclimation in a mesh-bottom enclosure, calibrated filaments (0.4–15 g; Stoelting 58011, USA) were applied perpendicularly to the mid-plantar hind paw for 5 s. Each trial was repeated three times, spaced 30 s apart, with a positive response defined as a brisk paw withdrawal or flinch. Using the up-down method, we calculated the paw withdrawal mechanical threshold (PWMT). The testing procedure commenced with the 2 g filament. Baseline measurements were recorded 1 day pre-injury, with subsequent testing on post-injury days 7, 14, 21, 28, and 35. NeuP-like behavior was defined as a sustained reduction in PWMT by two or more filament levels below baseline, accompanied by rapid foot retraction or licking.
Paw withdrawal thermal latency
An automated infrared heat stimulator (RWD Life Science Co., Ltd., China) was employed to assess thermal hyperalgesia. Rats were acclimatized in a quiet testing chamber for 30 min prior to assessment. A radiant heat stimulus (intensity = 30%) was directed to the hind paw plantar surface, and paw withdrawal latency (PWTL) was automatically recorded upon lifting or licking behavior. Three trials were conducted at 5 min intervals, with a mean PWTL calculated from all measurements. To prevent tissue damage, we set a 30 s cut-off. The baseline PWTL was measured prior to injury and then reassessed at 7, 14, 21, 28, and 35 days after injury. Established neuropathic hyperalgesia was defined as a sustained reduction in PWTL exceeding 2 s compared to baseline, a condition that typically developed between days 14 and 21 post-injury.
Harvesting of spinal cord tissues and dorsal root ganglia
Following the 5-week behavioral assessment, rats were euthanized and tissue samples were collected. Spinal cord segments (10 mm sections) containing the injury epicenter were harvested from all groups. Dorsal root ganglia (DRGs) were bilaterally isolated from lumbar segments L4–L6 and subjected to acute dissociation and primary culture as previously described. 22 We processed all tissue samples by snap-freezing in liquid nitrogen and archiving them at −80 °C for future analysis.
Western blotting
We extracted total protein from spinal cord tissue, DRGs, or RAW 264.7 macrophages using RIPA buffer (Beyotime, China) containing protease inhibitors (Beyotime, China). The homogenates were kept on ice for 15 min before a 10-min centrifugation at 12,000g to collect the supernatant. The resulting supernatant was collected, and protein concentration was determined using a BCA assay according to the manufacturer’s protocol (Beyotime, China). Protein samples were normalized using RIPA buffer, combined with 5× loading buffer (Beyotime, China), boiled for 5 min, and centrifuged (12,000g, 5 min, 4 °C). The prepared samples were stored at 4 °C until use. For electrophoresis, aliquots containing 15 μg of protein/lane were separated on 8% or 10% SDS–PAGE gels (Yeasen Biotechnology (Shanghai) Co., Ltd., China) at 200 V for 30 min. Subsequently, proteins were transferred onto 0.22 μm PVDF membranes (Millipore, USA) at a constant current of 300 mA for 50 min. Membranes were blocked with 5% skim milk for 1 h at room temperature, followed by three 5-min washes with TBST. Membranes underwent an overnight incubation at 4 °C in the presence of primary antibodies. These included anti-NKCC1 (ab303518, 1:1000; Abcam, UK), anti-CD68 (ab283654, 1:1000; Abcam, UK), anti-H3Cit (68345-1-Ig, 1:5000; Proteintech, China), α-tubulin (66031-1-lg, 1:20,000; Proteintech, China), or β-actin (AC026, 1:80,000; ABclonal, China). After rewarming at room temperature for 1 h and further washing, membranes were incubated for 1 h at room temperature with HRP-conjugated goat anti-rabbit IgG (AS014, 1:10,000; ABclonal, China) or HRP-conjugated goat anti-mouse IgG antibodies (AS003, 1:10,000; ABclonal, China). After washing three more times with TBST, the membranes were imaged on a Tanon 5200 Multi chemiluminescence system for band detection, and band intensity was quantified for analysis using GIS 1D software. Densitometric quantification was performed using ImageJ (National Institutes of Health, Bethesda, MD, USA), with background subtraction and normalization to α-tubulin or β-actin as loading controls.
Histology and immunofluorescence
Deep anesthesia was induced via intraperitoneal injection of pentobarbital sodium (50 mg/kg; Sigma-Aldrich, China). Prior to perfusion, a thoracotomy was performed. Animals were then perfused transcardially with 500 ml of ice-cold 0.9% saline, followed by 250 ml of 4% paraformaldehyde in phosphate buffer (pH 7.4) for fixation. Spinal cord segments containing the lesion epicenter and L4–L6 dorsal root ganglia were then dissected and post-fixed in the same fixative at 4 °C overnight.
For immunofluorescence analysis, the spinal cord and L4–L6 dorsal root ganglia were cut into 10 μm sections using a Leica CM1950 cryostat (Leica, Wetzlar, Germany). Sections were harvested in cold 0.01 M phosphate buffer (pH 7.4) and then processed to detect NKCC1, H3Cit, and CD68. We fixed the sections in 4% paraformaldehyde (Servicebio, China) for 10 min at room temperature, then rinsed them three times in PBS for 5 min each with gentle agitation. The sections were then permeabilized and blocked with 0.1% Triton X-100 (Beyotime, China) and 5% bovine serum albumin (BSA; Solarbio, China) in PBS for 2 h. The sections were subsequently incubated overnight at 4 °C with primary antibodies targeting NKCC1 (ab303518, 1:1000; Abcam, UK), Iba1 (ab283319, 1:200; Abcam, UK), CD68 (ab283654, 1:1000; Abcam, UK), or H3Cit (68345-1-Ig, 1:5000; Proteintech, China). Sections were washed three times in PBS (10 min/wash) and then incubated for 2 h in the dark with the corresponding fluorescent secondary antibody: either CoraLite594-conjugated goat anti-mouse IgG (SA00013-3, 1:250; Proteintech, China) or CoraLite488-conjugated goat anti-rabbit lgG antibodies (SA00013-2, 1:500; Proteintech, China). Following incubation, the sections were washed again in PBS. Finally, sections were mounted onto gelatin-coated slides using an anti-fluorescence quenching mounting medium with DAPI (Beyotime, China), dried, and imaged under an upright fluorescence microscope.
RAW 264.7 cells or OECs were fixed with 4% fixative solution (Servicebio, China) for 15 min and permeabilized with 0.1% Triton X-100 (Beyotime, China) for 10 min. After blocking with 5% BSA (Solarbio, China) in PBS for 30 min at 37 °C, samples were incubated overnight at 4 °C with primary antibodies against NKCC1 (ab303518, 1:1000; Abcam, UK), CD68 (ab283654, 1:1000; Abcam, UK), H3Cit (68345-1-Ig, 1:5000; Proteintech, China), or p75NTR (ab52987, 1:50, Abcam, UK). Cells were washed three times in PBS (1 min each) and then incubated for 30 min at 37 °C in the dark with either CoraLite594-conjugated goat anti-mouse IgG (SA00013-3, 1:250; Proteintech, China) or CoraLite488-conjugated goat anti-rabbit IgG antibodies (SA00013-2, 1:500; Proteintech, China). Following further PBS washes, nuclei were counterstained with DAPI, and samples were mounted in anti-fade medium (Beyotime, China) for confocal imaging. To assess lesion cavitation, a combined approach of confocal imaging and hematoxylin and eosin (H&E) staining (Beyotime, China) was used for histological evaluation. This involved incubating sections in hematoxylin for 2 min, followed by a brief 10-s differentiation. Excess stain was removed by rinsing the sections under running tap water, after which they were counterstained with eosin for 1 min. Following dehydration in a graded ethanol series (70%, 80%, 90%, and 100%), the sections were cleared, mounted with a neutral medium, and coverslipped. The cavitation area within the spinal cord was subsequently quantified.
Statistical analysis
All quantitative data are presented as mean ± SEM. Statistical analyses were performed using SPSS 25.0 (IBM, USA). All statistical analyses and graphing were performed with GraphPad Prism 10.0.0 (GraphPad Software, USA). For behavioral data (BBB scores, PWMT, PWTL) collected at baseline and on post-injury days 7, 14, 21, 28, and 35, a two-way repeated-measures ANOVA with Bonferroni correction was applied. Multiple group comparisons were conducted using one‑way analysis of variance, followed by Tukey’s post hoc test. Statistical significance was defined as a two-tailed p < 0.05.
Results
Morphological characterization and identification of OECs
To investigate whether OECs could suppress the formation of macrophage extracellular traps, we first established and characterized primary OEC cultures as described in the “Methods” section. Following purification and subculturing, OECs at passage six exhibited the characteristic flat soma morphology, predominantly adopting bipolar or multipolar shapes with slender, elongated cellular processes (Figure 1). Immunofluorescence staining confirmed the identity of the purified cells, showing positive expression of the p75 neurotrophin receptor (Figure 1(b)).
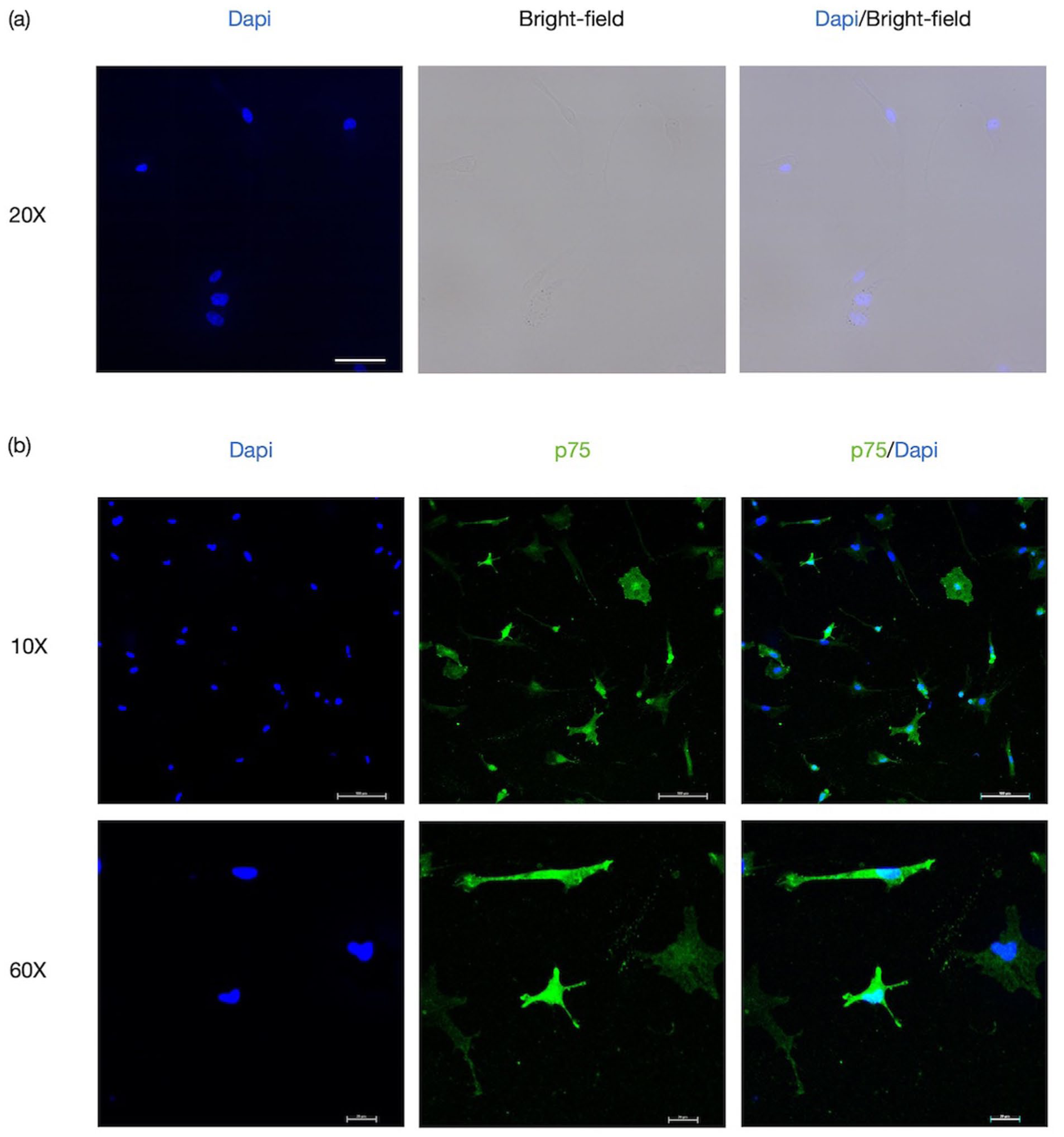
Morphological characterization and identification of olfactory ensheathing cells: (a) bright-field images show the representative morphology of OECs cultured up to the sixth generation. Scale bars: 50 μm and (b) immunofluorescence staining for p75, a marker expressed by purified OECs. Scale bars: 100 or 20 μm.
OEC transplantation enhanced locomotor recovery and attenuated NeuP in rats with SCI
After a 1‑week acclimation period, SD rats entered the study. To this end, two cohorts were established, with tissues collected at 1 and 5 weeks post‑surgery, respectively (Figure 2(a)). To investigate the effects of OEC transplantation on NeuP following SCI, OECs suspensions were injected into the lesion sites of SCI model rats. After acclimation, rats were randomized into Sham, SCI, or OEC-treated groups and sacrificed at 5 weeks post-surgery. As assessed by H&E staining, spinal cord tissue from SCI animals exhibited extensive cavitation. In contrast, OEC treatment substantially reduced lesion size relative to untreated SCI controls (Figure 2(b) and (c)).
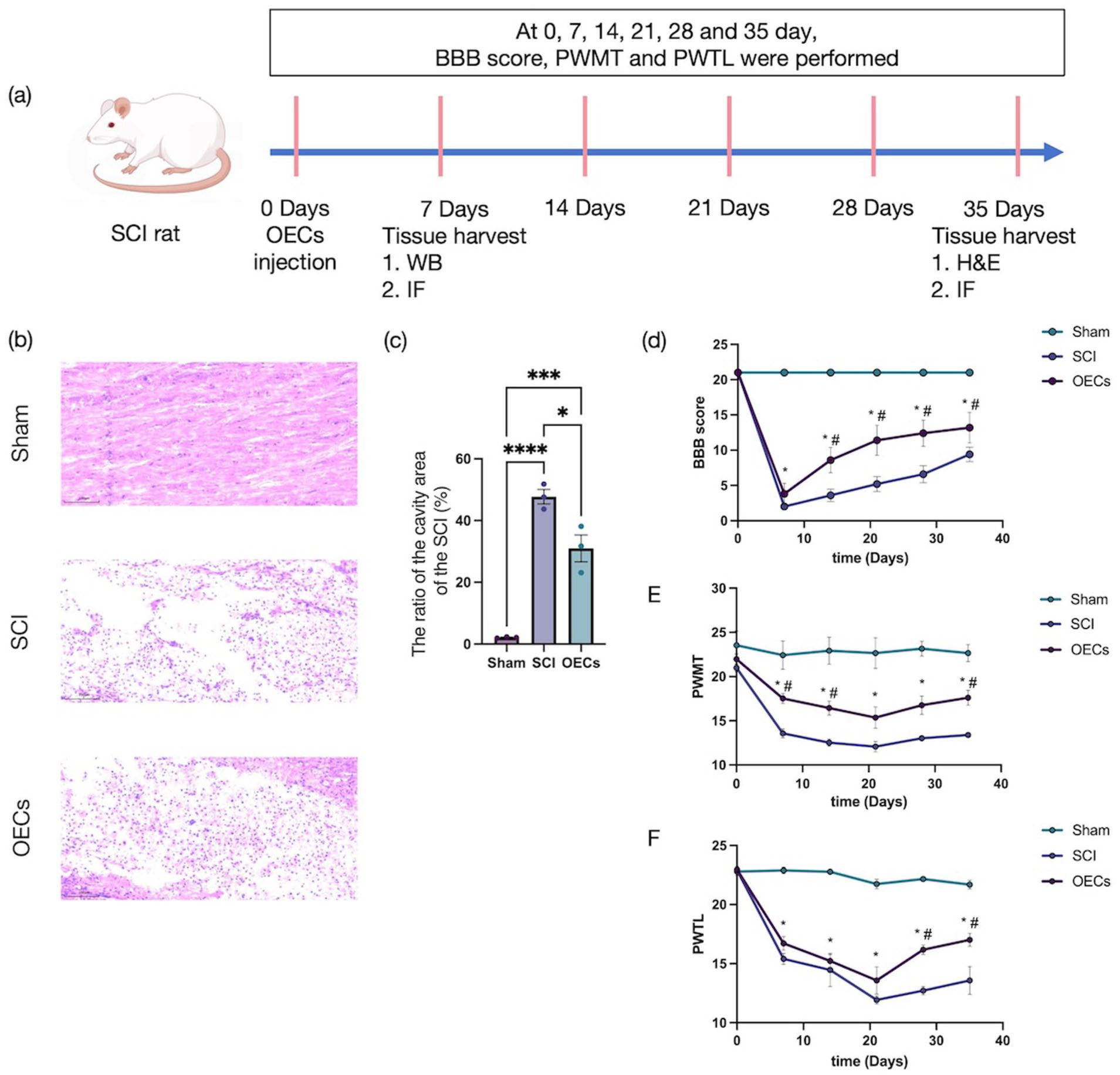
Functional recovery and pain sensitivity after spinal cord injury: (a) schematic of the in vivo experimental timeline, (b) representative hematoxylin and eosin-stained sagittal sections of the spinal cord lesion site (35 days post-injury). Scale bars: 100 μm, (c) quantification of the ratio of cavity area. Data are presented as mean ± SEM (n = 3). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 (one-way ANOVA followed by Tukey’s post hoc test), (d) BBB scores, (e) PWMT, and (f) PWTL assessed over 35 days post-injury. Data represent mean ± SEM (n = 6). *p < 0.05 versus Sham group; #p < 0.05 versus SCI group. Statistical analysis was performed using two-way ANOVA with Bonferroni post-hoc test.
Locomotor recovery was evaluated using the BBB scale. Compared with Sham controls, BBB scores in SCI rats declined sharply post-injury, indicating substantial motor impairment (Figure 2(d)). In contrast, OEC-treated animals exhibited a significant improvement in BBB scores from day 14 post-injury (p < 0.0001), which persisted throughout the observation period, suggesting that OEC transplantation partially restored motor function.
NeuP following SCI and the analgesic effects of OEC transplantation were further evaluated by measuring PWMT and PWTL (Figure 2(e) and (f)). Baseline PWMT and PWTL showed no significant differences between groups. In the subsequent 3 weeks, both SCI and OEC groups showed progressively heightened sensitivity, with significant decreases in PWMT (p < 0.01) and PWTL (p < 0.0001). These reductions indicate the successful induction of mechanical allodynia and thermal hyperalgesia. From day 21 onward, both PWMT and PWTL gradually increased. Notably, compared with the SCI group, OEC-treated rats demonstrated significantly higher PWTL from day 28 post-injury (p < 0.001) and a further increase in PWMT by day 35 (p = 0.025).
These findings indicate that NeuP manifests prominently ~3 weeks following spinal cord injury and that OEC transplantation not only facilitates motor recovery but also effectively ameliorates mechanical allodynia and thermal hyperalgesia.
OEC transplantation promotes neural repair and regeneration following SCI
Immunofluorescence analyses performed at 35 days post-injury revealed that, relative to sham-operated controls, spinal cord injury markedly reduced the expression of neurofilament 200 (NF200) while significantly increasing glial fibrillary acidic protein (GFAP) immunoreactivity (Figure 3). Notably, OEC transplantation partially reversed these injury-induced alterations. Compared with untreated SCI animals, OEC-treated rats exhibited significantly higher NF200 expression (p = 0.004) and markedly reduced GFAP levels (p < 0.001). These findings indicate that OEC transplantation attenuates glial scar formation while simultaneously promoting axonal regeneration, thereby facilitating neural repair following spinal cord injury.

Olfactory ensheathing cell transplantation promotes structural repair after spinal cord injury: (a) immunofluorescence staining for NF200 (red, CoraLite® 594) and GFAP (green, CoraLite® 488) in spinal cord sections at 35 days post-injury. Scale bars: 50 μm and (b, c) quantitative analysis of NF200 and GFAP immunofluorescence intensity. Data represent mean ± SEM (n = 3). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 (one-way ANOVA followed by Tukey’s post hoc test).
OEC transplantation suppresses MET formation in the spinal cord and DRG
At 7 days post-injury, immunofluorescence analysis revealed that the expression of H3Cit and CD68 in the spinal cord of rats with SCI was markedly upregulated compared with the sham operated group, and these markers exhibited colocalization (Figure 4(a) and (b)). Notably, OEC transplantation significantly attenuated this injury-induced increase, with the OEC-treated group exhibiting substantially reduced H3Cit and CD68 immunoreactivity relative to untreated SCI animals. Consistent with these observations, Western blot analyses of spinal cord tissue confirmed a pronounced elevation of H3Cit and CD68 protein levels following SCI, which was robustly suppressed by OEC transplantation (Figure 4(c)–(f)).
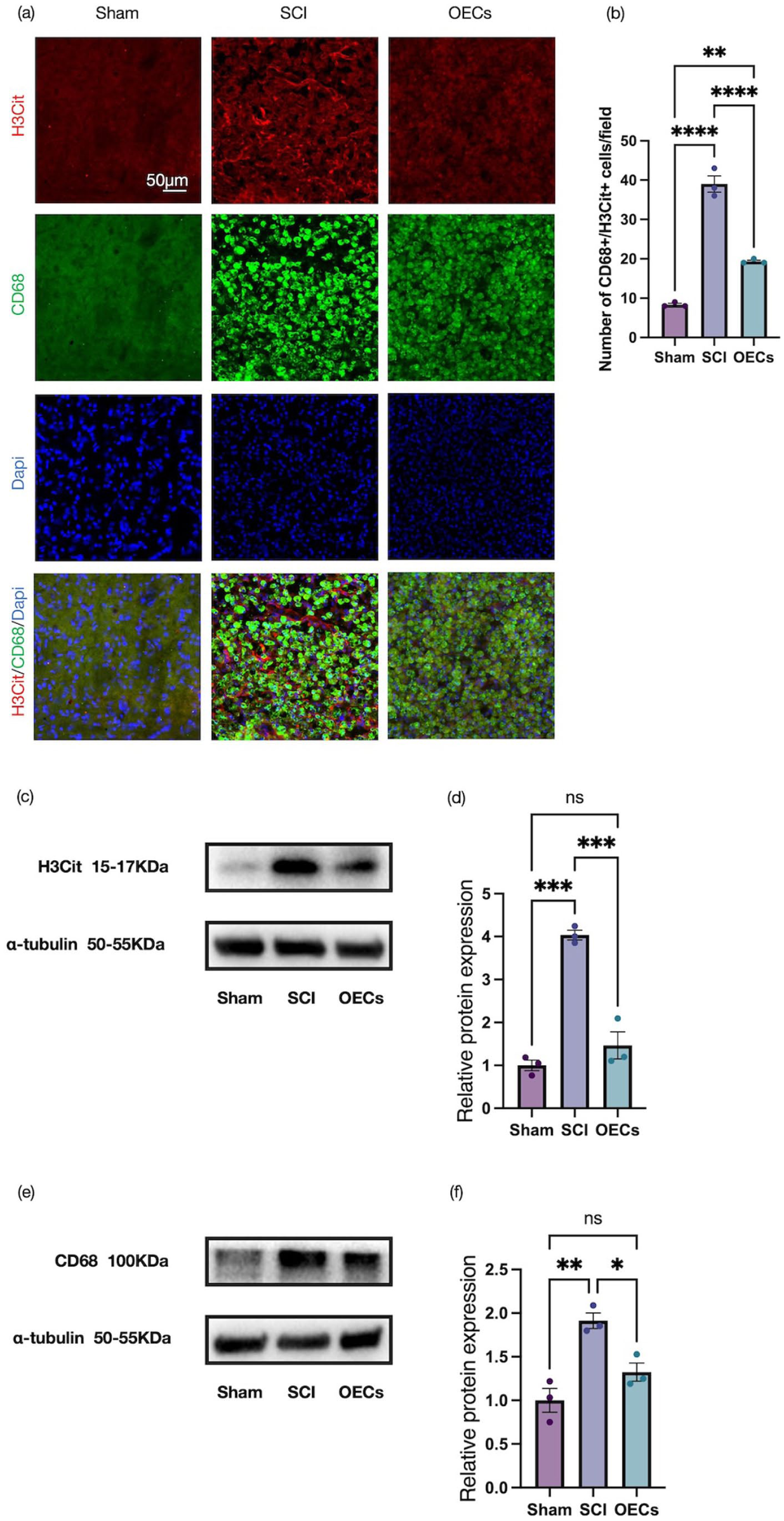
Olfactory ensheathing cell transplantation suppresses macrophage extracellular trap formation in the spinal cord: (a) representative immunofluorescence images of sagittal spinal cord sections stained for H3Cit (red, CoraLite® 594) and CD68 (green, CoraLite® 488) in the spinal cord at 7 days post-injury. Scale bar: 50 μm, (b) quantitative analysis of the number of CitH3+ CD68+ cells in three groups, (c, d) representative Western blots and quantification of H3Cit protein levels, and (e, f) representative Western blots and quantification of CD68 protein levels. Data are presented as mean ± SEM (n = 3). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 (one-way ANOVA followed by Tukey’s post hoc test).
A similar pattern was observed in the dorsal root ganglia at 7 days post-injury. Immunofluorescence staining demonstrated a significant increase in H3Cit and CD68 expression after injury, whereas OEC transplantation markedly reduced the levels of both markers (Figure 5(a) and (b)). These findings were further corroborated by Western blot analyses, which showed a significant decrease in H3Cit and CD68 protein abundance in the OEC-treated group compared with the SCI group (Figure 5(c)–(f)).
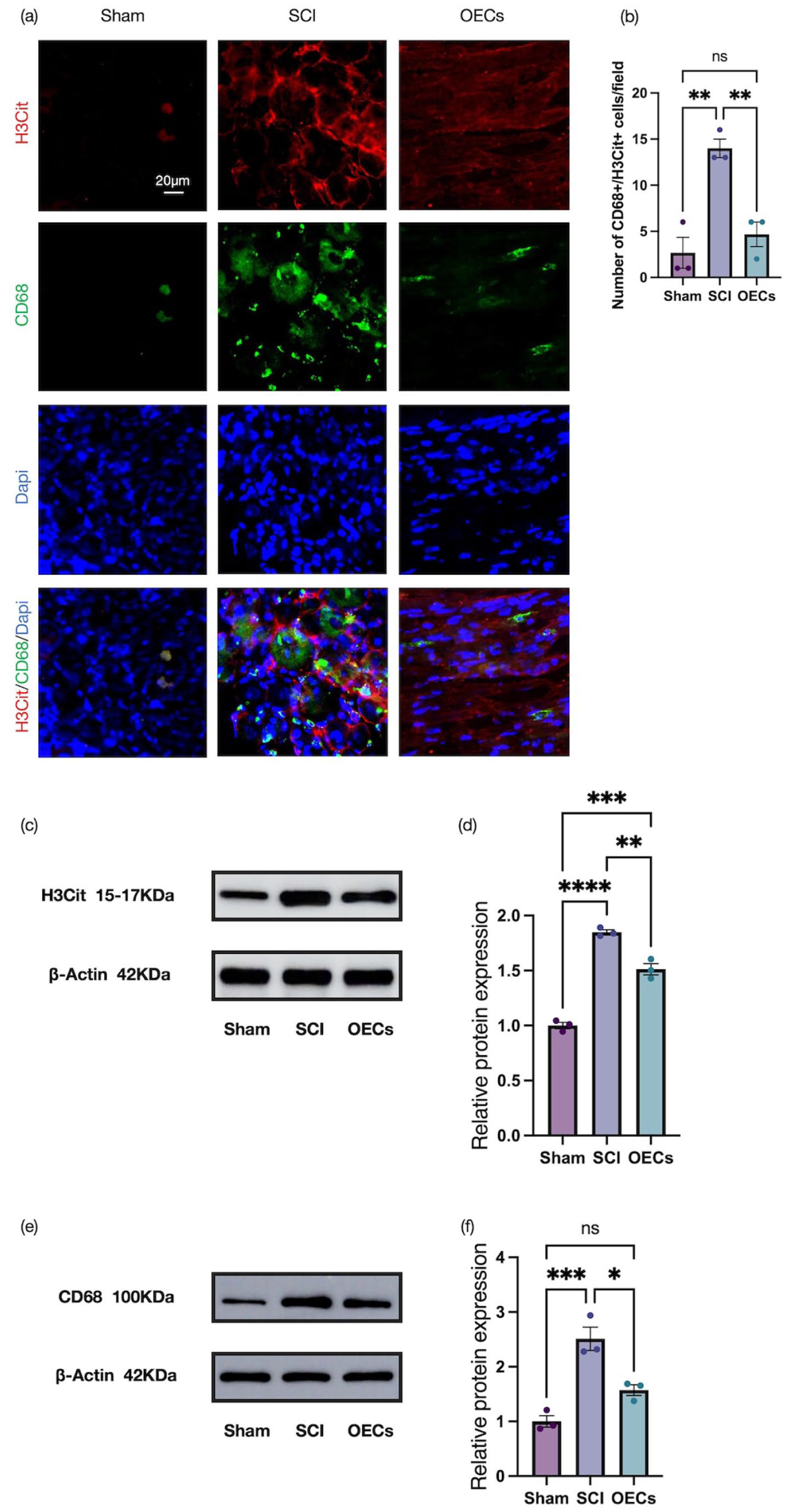
Olfactory ensheathing cell transplantation suppresses macrophage extracellular trap formation in the dorsal root ganglia: (a) representative immunofluorescence images of sagittal dorsal root ganglia sections stained for H3Cit (red, CoraLite® 594) and CD68 (green, CoraLite® 488) in the dorsal root ganglia at 7 days post-injury. Scale bar: 20 μm, (b) quantitative analysis of the number of CitH3+ CD68+ cells in three groups, (c, d) representative Western blots and quantification of H3Cit protein levels, and (e, f) representative Western blots and quantification of CD68 protein levels. Data are presented as mean ± SEM (n = 3). *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 (one-way ANOVA followed by Tukey’s post hoc test).
Collectively, these data demonstrate that OEC transplantation attenuates MET formation post-SCI, which is reflected in the decreased expression of MET markers in both spinal cord and DRGs.
OEC transplantation suppresses NKCC1 expression in the spinal cord and DRG following SCI
At 7 days post-injury, immunofluorescence staining for NKCC1 and Iba‑1 was performed on macrophages to evaluate NKCC1 expression in spinal cord tissue across experimental groups. As shown in Figure 6(a), SCI resulted in a marked increase in the fluorescence intensity of both NKCC1 and Iba‑1 per field of view, with colocalization observed. Notably, OEC transplantation significantly counteracted this injury induced upregulation, such that the expression levels of NKCC1 and Iba‑1 in the OEC treated group were substantially lower than those in untreated SCI rats (p = 0.0002). These findings were further substantiated by Western blot analyses, which confirmed a corresponding decrease in NKCC1 protein levels following OEC transplantation (Figure 6(c) and (d)).
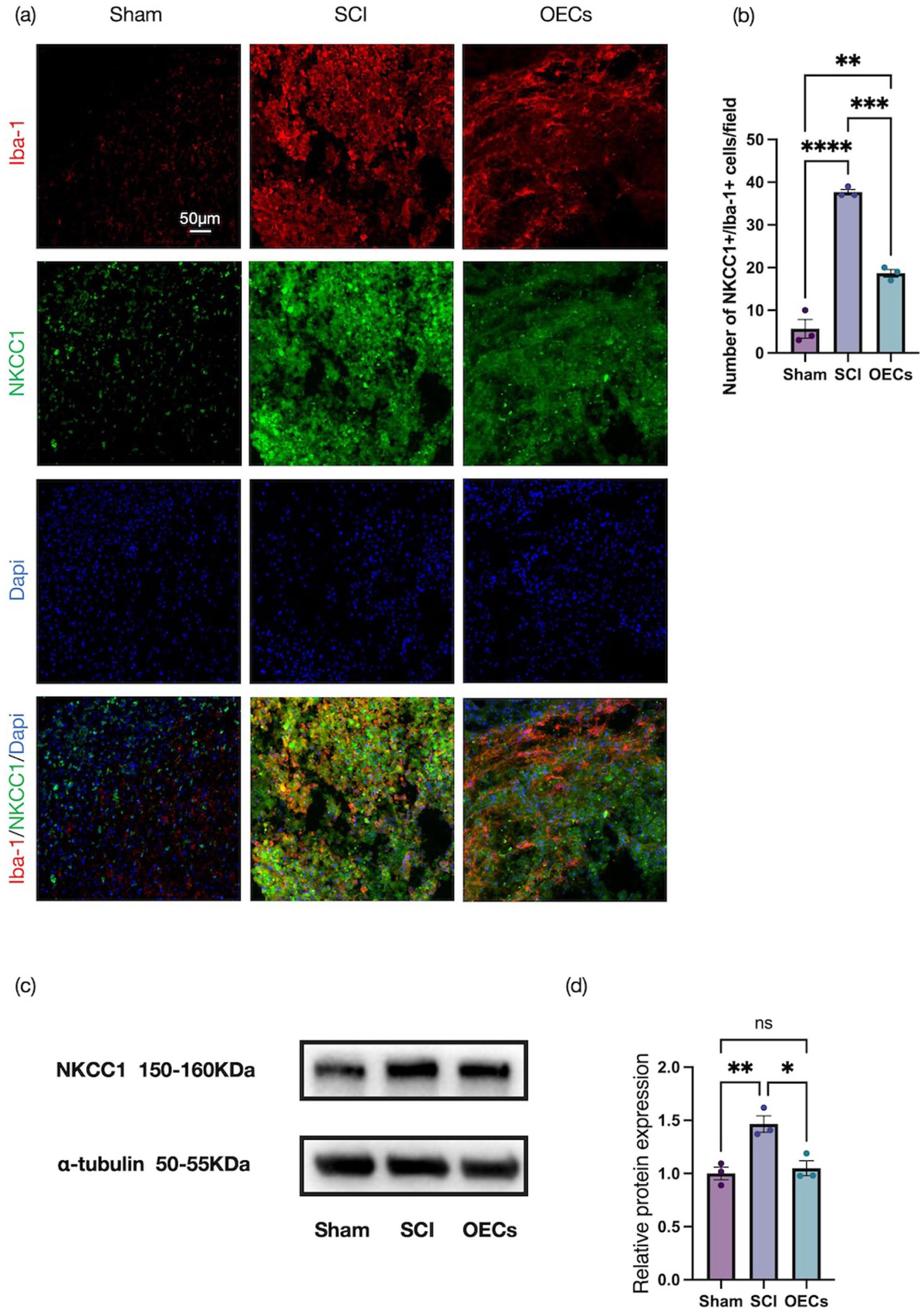
Olfactory ensheathing cell transplantation improves neuralgia after spinal cord injury: (a) representative immunofluorescence images of sagittal spinal cord sections stained for Iba-1 (red, CoraLite® 594) and NKCC1 (green, CoraLite® 488) in the spinal cord at 7 days post-injury. Scale bar: 50 μm, (b) quantitative analysis of the number of Iba-1+ NKCC1+ cells in three groups, and (c, d) Western blot analysis and quantification of NKCC1 protein expression in spinal cord tissue. Data represent mean ± SEM (n = 3). Statistical analysis was performed by one‑way ANOVA followed by Tukey’s post hoc test: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
To determine whether OEC transplantation exerted comparable effects within the peripheral sensory system, immunofluorescence analyses were conducted on dorsal root ganglia at 7 days post-injury (Figure 7). Compared with the sham operated control group, the SCI group exhibited a pronounced elevation in NKCC1 and Iba‑1 immunoreactivity in the DRG (p < 0.0001). In contrast, animals receiving OEC treatment showed significantly lower fluorescence intensity for both markers per field of view relative to the SCI control group (p < 0.0001). The upregulation of NKCC1 protein in DRGs following SCI, as confirmed by Western blot, was significantly attenuated by OEC transplantation (Figure 7(c) and (d)). The concordance between spinal cord and dorsal root ganglion findings suggests a coordinated regulation of NKCC1 expression across central and peripheral compartments after injury.
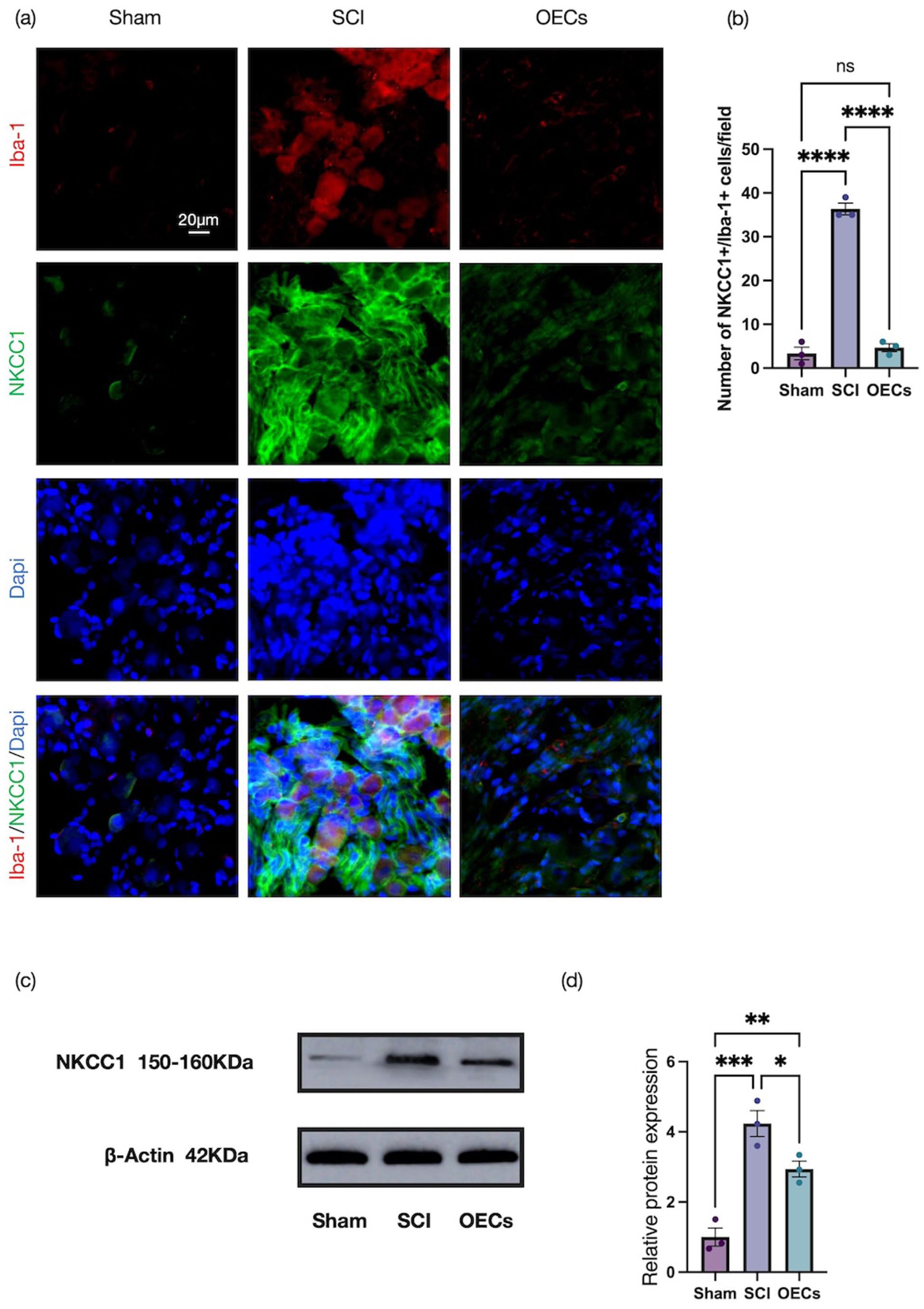
Olfactory ensheathing cell transplantation improves neuralgia after spinal cord injury: (a) representative immunofluorescence images of sagittal dorsal root ganglia sections stained for Iba-1 (red, CoraLite® 594) and NKCC1 (green, CoraLite® 488) in the spinal cord at 7 days post-injury. Scale bar: 20 μm, (b) quantitative analysis of the number of Iba-1+ NKCC1+ cells in three groups, and (c, d) Western blot analysis and quantification of NKCC1 protein expression in dorsal root ganglia tissue. Data represent mean ± SEM (n = 3). Statistical analysis was performed by one‑way ANOVA followed by Tukey’s post hoc test: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Taken together, these results demonstrate that OEC transplantation effectively suppresses NKCC1 expression within both the spinal cord and DRGs after injury, which underscores its role in mitigating associated chloride transporter dysregulation.
OEC-CM attenuates TNF-α-induced MET formation and suppresses NKCC1 expression in RAW 264.7 cells
We next investigated the effects of OEC-CM on MET formation in cultured RAW 264.7 cells. MET formation was assessed using Nuclear Green staining, which revealed that stimulation with TNF-α robustly induced MET generation in RAW 264.7 cells, whereas co-treatment with OEC-CM markedly attenuated MET formation (Figure 8(a)). To further explore the associated molecular changes, immunofluorescence analyses were performed to evaluate NKCC1 expression. Compared with the CM control group, TNF-α treatment resulted in a significant increase in NKCC1 immunoreactivity, while NKCC1 protein expression was substantially reduced in the presence of OEC-CM (Figure 8(b) and (c)). These observations were confirmed by Western blot analyses, which demonstrated a consistent suppression of NKCC1 protein levels following OEC-CM treatment (Figure 8(d) and (e)), indicating that OEC-CM effectively inhibits NKCC1 expression.
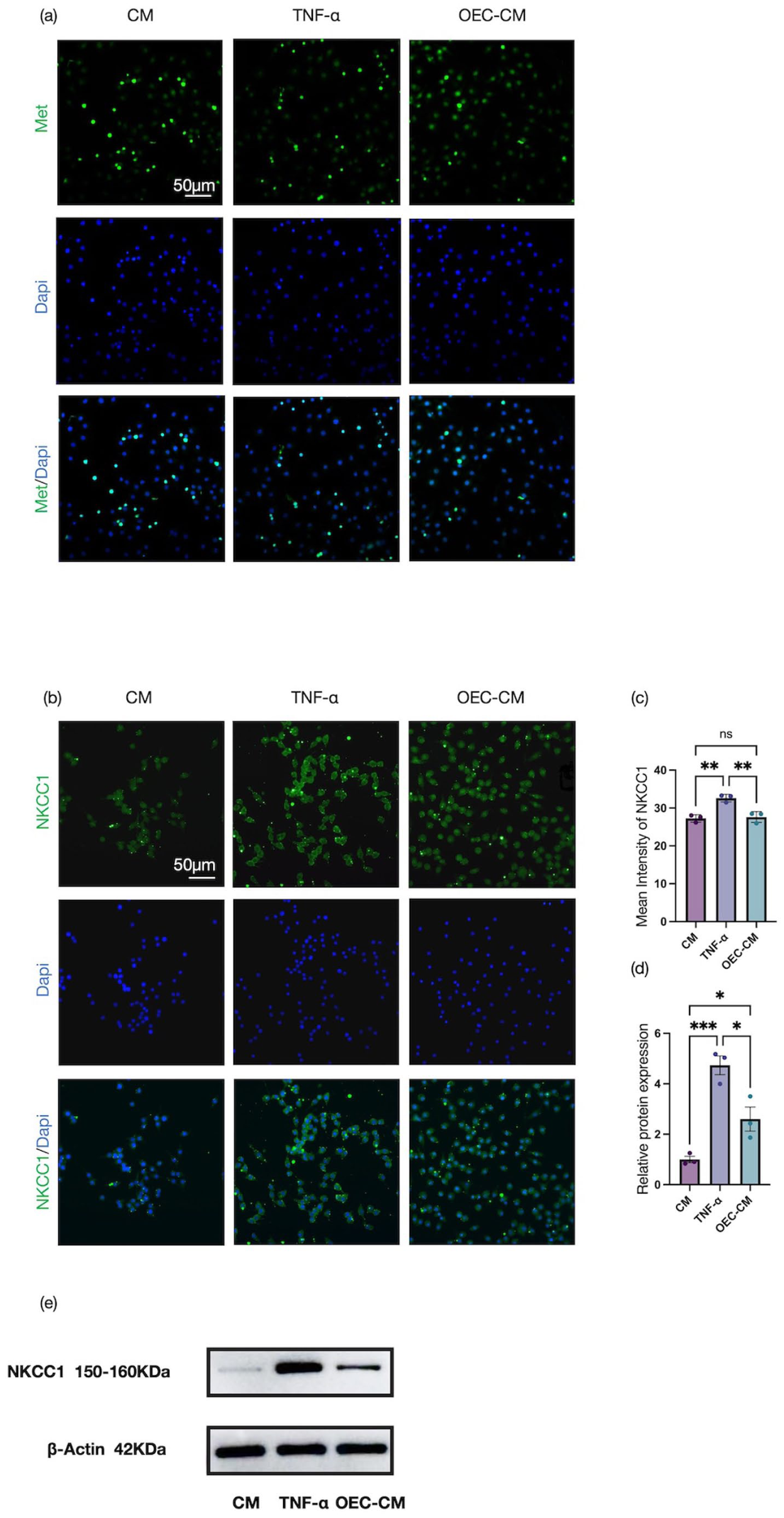
Detection of macrophage extracellular traps and NKCC1 expression in RAW 264.7 cells: (a) nuclear green staining of RAW 264.7 cells following TNF-α stimulation to visualize METs formation, (b, c) immunofluorescence analysis and quantitative assessment of NKCC1 expression in RAW 264.7 cells, and (d, e) Western blot analysis and quantification of NKCC1 protein expression in RAW 264.7 cells. Scale bars: 50 μm. Data represent mean ± SEM (n = 3). Statistical analysis was performed by one‑way ANOVA followed by Tukey’s post hoc test: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
In addition, MET-associated markers were examined by immunofluorescence staining and Western blotting. Immunofluorescence analyses showed that TNF-α stimulation markedly increased the expression of H3Cit and CD68 in RAW 264.7 cells compared with controls, whereas these elevations were reversed by OEC-CM co-treatment (Figure 9(a), (b), (e), and (f)). Consistently, Western blotting showed TNF-α-induced upregulation of H3Cit and CD68, which was markedly reduced by OEC-CM treatment (Figure 9(c), (d), (g), and (h)). Collectively, these results demonstrate that OEC-CM effectively suppresses TNF-α-induced MET formation in RAW 264.7 cells.
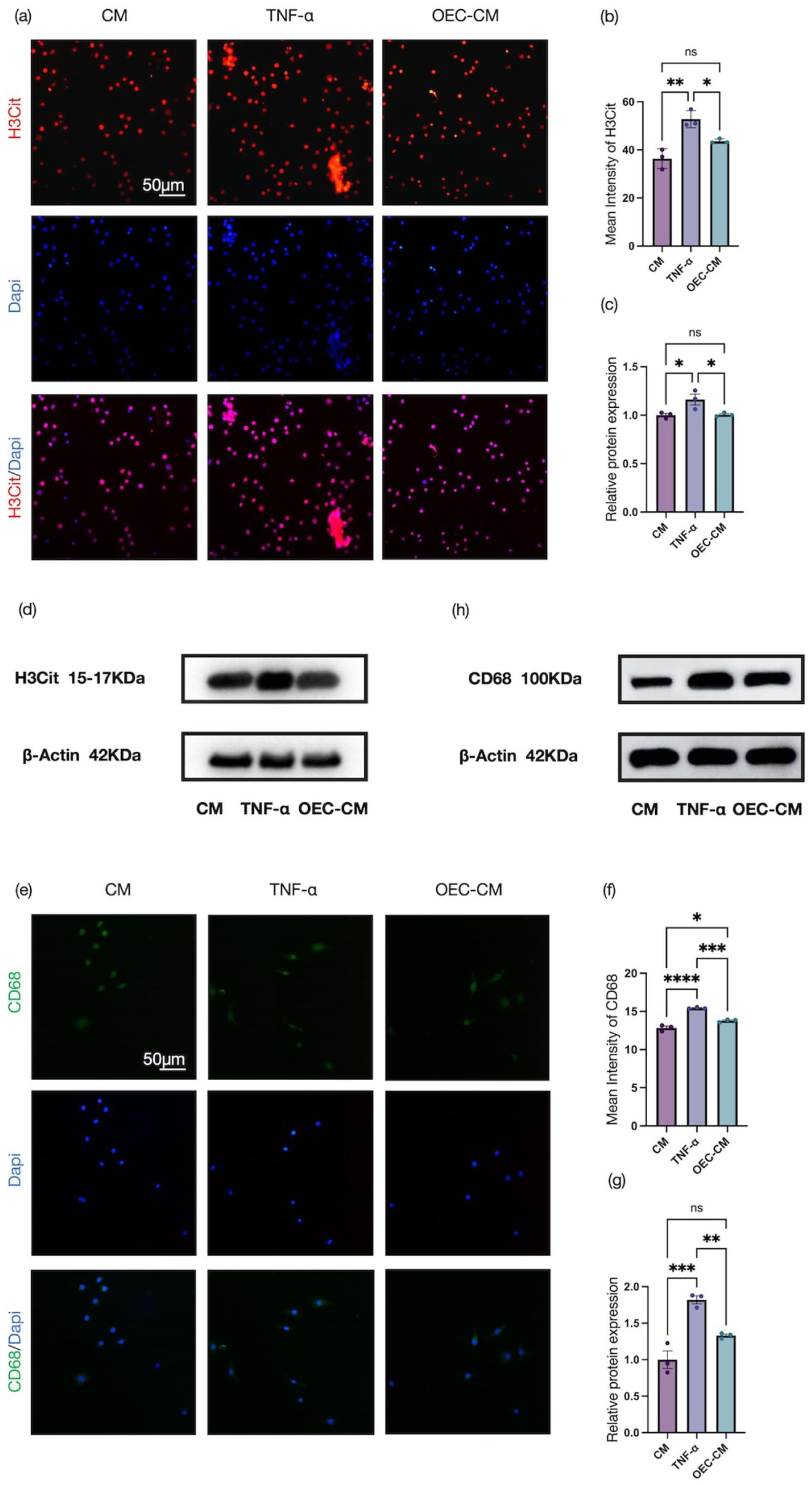
Effects of OEC-CM on macrophage extracellular trap formation in RAW 264.7 cells: (a, b) representative immunofluorescence images and quantitative analysis of H3Cit (red, CoraLite® 594) in RAW 264.7 cells, (c, d) representative Western blots and quantification of H3Cit protein levels, (e, f) representative immunofluorescence images and quantitative analysis of CD68 (green, CoraLite® 488) in RAW 264.7 cells, and (g, h) representative Western blots and quantification of CD68 protein levels. Scale bars: 50 μm. Data represent mean ± SEM (n = 3). Statistical analysis was performed by one‑way ANOVA followed by Tukey’s post hoc test: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Discussion
This study demonstrates that transplantation of rat-derived OECs effectively alleviates post-injury NeuP and promotes the recovery of motor function in a rat model of SCI. The therapeutic benefits of OEC transplantation are manifested through the regulation of chloride ion homeostasis within the injured spinal cord, attenuation of glial scar formation, and facilitation of axonal regeneration. Mechanistic studies further indicate that OECs alleviate NeuP by suppressing MET formation, thereby reducing NKCC1 expression and activity. Furthermore, using an in vitro cell model, we further validated the inhibitory effect of OEC‑CM on METs formation and NKCC1 expression in TNF‑α‑induced RAW 264.7 macrophages.
OEC transplantation ameliorates NeuP following spinal cord injury
OECs ensheathe olfactory axons as they traverse from the peripheral nervous system into the central nervous system and represent, to date, the only known population of glial cells capable of crossing the peripheral–central nervous system boundary while retaining regenerative capacity in adulthood. 23 Characterized by phenotypic and functional traits of both central and peripheral glia, OECs represent a promising candidate for cell-based therapies aimed at treating spinal cord injury, peripheral nerve injury, and various neurological disorders. 14 During therapeutic intervention, OECs express a diverse repertoire of trophic and regulatory factors with the potential to suppress inflammation, modulate pain signaling, and promote neural regeneration.24–29 Transplantation of OECs into rat models of spinal cord injury as well as sciatic nerve injury has been shown to significantly alleviate NeuP-like behaviors.29,30 In addition, our previous studies demonstrated that inhibition of NKCC1 expression in the spinal cord and dorsal root ganglia attenuates NeuP and facilitates motor functional recovery in SCI rats. 31 Despite these findings, it has remained unclear whether OECs participate in pain modulation through the regulation of NKCC1 expression. Here, we provide the first evidence that OEC transplantation mitigates NeuP after spinal cord injury by modulating NKCC1, thereby promoting neural repair and functional recovery.
Excessive excitability of sensory neurons is widely recognized as a principal driver of NeuP following SCI. 32 SCI induces neuronal hyperexcitability, resulting in sustained release of excitatory neurotransmitters and consequent activation of glial populations, including microglia and astrocytes. 32 Activated astrocytes not only contribute to glial scar formation – thereby impeding axonal regeneration and reconnection – but also secrete pro-inflammatory mediators that further exacerbate neuronal hyperexcitability, ultimately intensifying NeuP. 32 In the present study, behavioral assessments revealed that injured rats developed pronounced mechanical allodynia and thermal hyperalgesia between 7 and 21 days after SCI. Notably, transplantation of OECs significantly increased both the mechanical withdrawal threshold and thermal withdrawal latency, concomitant with marked improvements in motor functional recovery. To clarify how OEC transplantation exerts its analgesic effects at the molecular level, we performed comprehensive morphological and immunohistochemical analyses. At 35 days post-injury, OEC transplantation markedly reduced GFAP expression within the lesion epicenter, while significantly upregulating NF200 expression. These changes indicate a robust suppression of astrocytic overactivation, attenuation of glial scar formation, and promotion of axonal regeneration and repair, in agreement with previous reports. 27 These results collectively suggest that OECs exert analgesic effects by dampening glial and immune cell activation and limiting the release of pronociceptive mediators, including pro-inflammatory cytokines and neuropeptides. 33
Beyond neuronal hyperexcitability, disinhibition of nociceptive signaling represents a critical mechanism underlying pain sensitization. 4 Following neural injury, inhibitory neurotransmission mediated by γ-aminobutyric acid and glycine is markedly attenuated, accompanied by a disruption of the intracellular–extracellular chloride ion gradient. This imbalance compromises inhibitory control within sensory neuronal networks, leading to hyperalgesia and aberrant pain perception.34–37 NKCC1, a major chloride transporter, is implicated in NeuP by modulating both its expression and functional activity. 10 Previous studies have reported a significant upregulation of NKCC1 protein levels in SD rats between 2 and 14 days after SCI. 38 Building on these observations, we investigated the effects of OEC transplantation on NKCC1 expression in both the spinal cord and dorsal root ganglia. Immunofluorescence and Western blot analyses performed 7 days after injury revealed that OEC transplantation significantly decreased NKCC1 protein levels in both the injured spinal cord and DRGs. These findings suggest that OECs may suppress the development of hyperalgesia by restoring chloride homeostasis through modulation of NKCC1 expression and function, thereby influencing both central and peripheral mechanisms involved in the initiation and maintenance of SCI-induced pain. Collectively, our results indicate that OEC transplantation alleviates ionic homeostasis imbalance following spinal cord injury, contributing to the mitigation of post-injury NeuP.
OECs may alleviate NeuP by suppressing MET formation
ETs represent a distinct form of cell death that differs from classical programmed cell death and necrosis, involving the release of extracellular reticular structures composed of DNA and proteins. When these structures are released by macrophages, they are specifically termed METs.11,12 To date, investigations into the mechanisms underlying METs formation remain relatively limited; however, given their structural similarity to neutrophil extracellular traps (NETs), researchers have often drawn insights from the mechanisms of NETs formation. 39 Consequently, numerous studies have revealed that METs formation is similarly associated with the nicotinamide adenine dinucleotide phosphate (NADPH)/reactive oxygen species (ROS) system and peptidyl arginine deiminases (PADs).40–47 Of note, existing evidence indicates that NETs formed in the spinal cord can promote pain development. 48 Specifically, following local infiltration, neutrophils release NETs, which in turn facilitate the release of inflammatory mediators and exacerbate peripheral inflammation. These peripheral stimuli are further transmitted to the spinal cord, triggering microglial activation and NF‑κB phosphorylation, thereby enhancing neuroinflammation and promoting pain sensitization. 49 Conversely, inhibition of peripheral NETs alleviates peripheral inflammation in mice with inflammatory pain and reverses mechanical and thermal hyperalgesia by suppressing spinal microglial activation. 49 Although a direct link between METs and pain has yet to be elucidated, the above regulatory mechanisms of NETs in pain provide an important theoretical basis for the present study, which seeks to investigate whether METs are involved in neuropathic pain following spinal cord injury.
Previous studies have demonstrated that mesenchymal stem cells can inhibit the formation of ETs, yet whether OECs exert therapeutic effects through the suppression of ETs remains unreported.50,51 Macrophages are known to play a proinflammatory role in neuropathic pain, and their depletion from the peripheral nervous system has been shown to alleviate mechanical hyperalgesia.52,53 However, a direct link between METs, which are proinflammatory structures derived from macrophages, and pain has yet to be established. In the present study, we found that OEC transplantation significantly inhibited the formation of METs following SCI. In vivo experiments revealed that at day 7 post injury, rats exhibiting NeuP showed markedly increased fluorescence intensity of the METs markers H3Cit and CD68 in both the spinal cord lesion site and the dorsal root ganglia. Following OEC transplantation, the expression of these two markers was substantially reduced, indicating that METs formation was suppressed. Given that OEC transplantation concurrently inhibited METs formation and alleviated NeuP, these results suggest the involvement of METs in pain modulation. Furthermore, our study revealed that OEC transplantation downregulated the expression of NKCC1, a key transporter for maintaining chloride homeostasis, whose upregulation weakens GABAergic inhibitory signaling and thereby promotes nociceptive disinhibition. 10 Because upregulation of NKCC1 is frequently associated with a proinflammatory milieu characterized by elevated levels of tumor necrosis factor alpha and interleukin‑1 beta, 9 and because METs formation exacerbates such inflammation, 12 we hypothesize that the inhibitory effect of OECs on NKCC1 is achieved indirectly through the suppression of METs formation and the consequent reduction of inflammatory responses. In summary, we propose a potential mechanistic pathway by which OECs alleviate neuropathic pain. OECs inhibit the formation of METs, thereby attenuating macrophage mediated inflammation. This in turn reverses the neuronal hyperexcitability that arises from NKCC1 upregulation, ultimately producing an analgesic effect.
OEC-CM suppresses MET formation and NKCC1 expression
Stem cell-derived conditioned media have emerged as a promising cell-free therapeutic strategy in regenerative medicine, offering the advantage of circumventing risks associated with direct cell transplantation, such as immune rejection and graft-related complications.54,55 To elucidate the mechanisms by which OEC-CM regulates MET formation and NKCC1 expression, a series of in vitro experiments were carried out. The murine RAW 264.7 cell line, a macrophage-like model widely employed to recapitulate macrophage inflammatory responses and functional phenotypes, was used in this study. 42 Consistent with previous reports, stimulation with TNF-α resulted in a pronounced upregulation of MET-associated markers, including H3Cit and CD68. Notably, co-culture of TNF-α-stimulated RAW 264.7 cells with OEC-CM markedly attenuated the expression of both H3Cit and CD68, indicating a robust suppression of MET formation. In parallel, TNF-α exposure induced a significant increase in NKCC1 expression in RAW 264.7 cells, an effect that was similarly reversed by OEC-CM treatment. These findings suggest that the OEC conditioned medium may contain certain constituents with anti‑inflammatory properties, capable of modulating the phenotype and function of macrophages, reducing the release of METs, and downregulating the expression level of NKCC1, thereby exerting both anti‑inflammatory and analgesic effects.
Limitations
We evaluated the potential of OEC transplantation to alleviate NeuP following contusive SCI in rats. The present study has two main limitations. First, the specific bioactive constituents within OECs and OEC‑CM that mediate the analgesic effects remain unclear. Previous research has shown that OECs secrete anti‑inflammatory factors such as IL‑1Ra, which can inhibit microglial activation and downregulate the levels of proinflammatory cytokines including interleukin‑1 alpha and interleukin‑1 beta, thereby alleviating neuroinflammation. 18 We hypothesize that these secreted factors may play a critical role in modulating MET formation and associated inflammatory signaling pathways. Future studies could employ proteomic analysis or microRNA sequencing to investigate OEC‑CM, aiming to identify key effector molecules and perform further functional validation. Second, the present study did not specifically intervene in METs or NKCC1 in the animal model. Therefore, whether OECs alleviate pain behaviors by suppressing METs and consequently modulating NKCC1 requires verification through in vivo experiments. Subsequent research could adopt myeloid‑specific PAD4 knockout mice to directly block MET formation, combined with assessments of pain behavior and inflammatory markers, to clarify the causal relationship between METs and NKCC1 in the analgesic effect. Alternatively, NKCC1 overexpression and knockout experiments could further validate the causal role of NKCC1 in the analgesic effects of OECs. These investigations will help to elucidate the mechanism underlying the H3Cit‑CD68‑NKCC1 signaling axis.
Conclusions
Following SCI, the formation of METs contributes significantly to the development of NeuP. Our findings suggest that OEC-CM may protect RAW 264.7 macrophages in vitro by inhibiting the H3Cit/CD68/NKCC1 signaling axis, thereby suppressing TNF-α-induced MET formation and downregulating NKCC1 expression. In vivo, OEC transplantation reduced MET-associated marker expression, partially reversed astrocytic overactivation, attenuated glial scar formation, and promoted axonal regeneration and neural reconnection. These effects collectively dampened sensory neuron hyperexcitability, alleviated NeuP, and enhanced motor functional recovery. This study demonstrates that OEC transplantation is therapeutically effective for SCI. Furthermore, it highlights OEC-CM as a promising cell-free strategy for treating NeuP, underscoring its potential for clinical translation.
Footnotes
Acknowledgements
We gratefully acknowledge the contributions of all researchers involved in this study.
Author contributions
HL, TL, YY, and SS performed animal models and data collec‑tion and statistical analysis. HL conceived the study, acquired the data, interpreted the results, and drafted the article. FW performed funding acquisition, conceptualization, writing original draft, project administration, writing review and editing, methodology, supervision, and resources. The final manuscript has been reviewed and approved by all authors.
Data availability statement
The raw data are available from the authors without restriction. Access requests may be directed to
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Key R&D Program of China (project no. 2021YFF0501600 and subject no. 2021YFF0501604).
Ethical considerations
This study was approved by the Animal Care and Use Committee of Capital Medical University (ethical code: AEEI-2024-239).
Consent to participate
Not applicable.
Consent for publication
Not applicable.